中国野生大豆生物学研究(庄炳昌)思维导图
大豆遗传图谱的构建

大豆遗传图谱的构建摘要大豆是严格的自花授粉作物,是由古四倍体演变而来的二倍体,基因组较大,染色体又很小,难于进行细胞遗传学研究,因此构建分子遗传图谱并对重要性状进行QTL定位,对于重要性状基因的图位克隆及分子标记辅助育种具有重要意义。
就大豆遗传图谱构建过程和研究进展进行综述,并对有待进一步研究领域进行了分析讨论,以期促进大豆遗传图谱的构建,促进育种工作的发展。
关键词大豆;遗传图谱;分子标记大豆是人类优质蛋白和脂肪的重要来源,同时也是饲料蛋白的重要来源[1-2]。
因此,大豆的遗传研究一直受到广泛的重视。
但由于大豆的遗传变异程度低,基因组较大并含有广泛的复制区和丰富的重复序列,染色体又很小,难于进行细胞遗传学观察等研究,导致大豆的遗传研究尤其遗传作图明显滞后于其他作物。
随着分子遗传学的发展和RFLP、RAPD、SSR、ARLP等分子标记技术的发展,尤其是高密度遗传图谱的构建,不但对数量性状位点(QTLs)进行精确定位,而且能利用图位克隆克隆出控制数量性状的基因。
因此,可以利用高密度的大豆遗传连锁图谱,结合现代分子操作技术,对有重要价值的性状进行QTL定位,尽可能地挖掘有利用价值的等位基因,将分子标记辅助选择用于育种实践,以培育优质高产的新品种。
1遗传图谱的构建过程遗传图谱是数量性状基因定位(QTL)、基因图位克隆、比较基因组学研究以及分子标记辅助育种等的基础。
图谱的构建过程主要包括:①选择建立适合的作图群体;②选择适合作图群体的遗传标记;③确立连锁群;④基因排序与遗传距离的确定。
1.1群体的选择适合于作图的遗传群体有F2、BC、NIL、DH、RIL等,作图群体的选择取决于使用的标记类型。
由于SSR标记具有较高的多态性,可以用于在亲缘关系较近的亲本甚至一些近交获得的群体中构建图谱。
另外,对于共显性的标记使用F2群体可以获得最多的遗传信息,而对于显性标记,使用DH、RIL则可以获得最多的遗传信息。
1.1.1F2群体或其衍生的F3、F4家系。
大豆起源

精心整理一、栽培大豆起源于中国是国内外学者所公认的栽培大豆起源于中国,早已为国内外学者所公认。
正像《美国大百科全书》所指出的:“中国古文献认为,在有文献记载以前,大豆便因营养价值高而被广泛地栽培。
同时在公元2000年前大豆便被看作是最重要的豆科植物。
”《苏联大百科全书》更明确指出:“栽培大豆原产于中国,中国在5000年前就开始栽培这个作物,并由中国向南部及东南各国传播,于16世纪进入欧洲。
”苏联着名的学者瓦维洛夫在《主要栽培植物的世界起源中心》一书中指出:“第一个最大的独立的世界农业发源和10“大豆的,本(● 1712● 1740● 1790● 1873 ● 1840● 1822● 1910● 1915● 1929年美国已种植大豆400万亩。
● 1941年二战期间,美国由于缺乏食用油,开始大规模种植大豆。
到1944年美国大豆种植面积达6000多万亩。
1972年美国大豆种植面积达4.28亿亩,产额达到1,234亿斤。
二、关于栽培大豆起源地的几种假说栽培大豆起源于中国,早已为国内外学者所公认,但是,栽培大豆究竟起源于中国何地何时却是众说纷纭的问题。
我们在这里对主要的几种假说作以下介绍并加以评论:1.黄河中下游起源说有的学者根据古代文献、考古文物、栽培大豆品种资源和野生大豆的分布,对栽培大豆的起源进行了分析(1985),提出栽培大豆起源数千年前,最早记载栽培大豆的地区是黄河中下游的河南、山西、陕西等地。
还有学者从农业的起源、以及古代文献、考古资料等方面对大豆起源进行分析以外,还联系野生大豆的分布和性状演化作了分析(1989),他们在全国野生大豆资源考察中发现黄河中下游的陕西、山西、河南,不仅野生大豆分布普遍,而且群落大,类型最为丰富,在野生的环境中,有多种进化程度的野生、半野生大豆存在。
因此,认为栽培大豆的主要起源地应该是黄河中下游地区。
2.河北东北部及东北中南部起源说有的学者认为,中国的北方地区不仅有考古发掘最早的大豆实物,最早的文字记载,而且也是我3.4.,加栽培大豆起源于黄河中下游的假说,所引用的古代文献和考古资料是不全面的,不准确的。
野生大豆种质资源的综合应用

科
野生大豆种质资源 的综 合应用
位昕 禹 ‘ 杨 德 光 ‘ 吴纪 安 陈祥 金 于跷 光 崔 杰印 z 吴 俊 彦 2 ・ 。
(、 1东北 农业 大 学农 学 院 , 龙 江 哈 尔滨 10 3 2 黑 龙 江省农 业科 学 院黑 河分 院 , 黑 50 0 、 黑龙 江 黑 河 140 ) 63 0
摘 要:介 绍 了野 生 大豆 的性 状 . 并分 别介 绍 了野 生大 豆在 大 豆育 种 中 , 料作 物 上及 药用 方 面 的应 用价 值 。 阐述 了利 用野 生 大豆 所取 得 的成 果 , 饲 并 对野 生 大豆 大豆 资 源的进 一 步利 用提 出 了建议 。 关键 词 : 生 大豆 ; 用 ; 源 野 应 货 野生大 豆为一 年生草 本. 落豆秧 、 俗称 野大 豆 、 源研究 所利用野 生大豆 为亲本与 栽培大 豆进行 杂交 对肿 瘤 、 动脉硬化 等多种 疾病都具有预防疗效 。野生 中野 1 和“ 号” 中野 2 大豆新品种' 号” 该品种 大豆 的异黄酮 含量较栽 培大豆高, 原因是 人 主要 们很 属于豆科 、 蝶形花亚科大豆属提 栽培大豆的近缘野 培育出“ 早就意 识到大 豆异黄酮 是导致 大豆食品苦 涩味产生 生种。羽状三 出复叶/、 J 叶薄纸质 、 、 卵形 卵状 椭圆形 具有耐早耐盐喊高蛋白。 或卵状披针形. l6Ⅱ 13m 长 _ c堪£-c o总状花序腋生, 花 栽培大豆资源过于单一化' 遗传基础狭窄, 野生 的抗营养因子, 因此在大豆品种选择改良的过程中人 小敞 紫色和白 花粤钟状; 色 花冠蝶形旗瓣近圆形, 结 大豆种质资源丰富,挖掘抗病基因是一条有效的途 工选择很 自然地会淘汰那些 口味差的大豆种魔间接 荚习性已无限型居多. 也有少数的亚有限型。籽粒色 径。中国科学院遗传所乖
高清版新课标高中生物思维导图
过程
光反应 暗反应
场所,条件 物质变化 能量变化
实质、意义
影响光合作用 的因素及应用
内部因素 外界因素
叶面积指数 叶龄 光照强度 温度 二氧化碳浓度 必需矿质元素
水
ATP的主要来源
概念
有氧呼吸
方式
比较
无氧呼吸
实质
意义
影响呼吸作用 的因素及应用
内部因素 环境因素
遗传因素 温度 氧气浓度 CO2浓度
含水量
DNA
分类 RNA
功能
糖类
元素 种类及作用
脂质
元素 种类及作用
有氧呼吸主要场所 光合作用的场所
线粒体 叶绿体
双层膜 细胞器
对来自内质网的蛋白质 进行加工、分类和包装
高尔基体
增大膜面积,与蛋白质、脂质 和糖类的合成有关,蛋白质 的运输通道
维持渗透压,使细胞膨胀
内质网 液泡
单层膜 细胞器
含有多种水解酶,可 分解细胞器和病毒
染色质
结构 功能
遗传物质的载体
是遗传信息库,是细胞代谢和遗传的控制中心
细胞膜 细胞壁
细胞的基 本结构
2/20
细胞骨架
组成 蛋白质纤维组成的网状结构
作用
在真核中维持细胞形态; 保持细胞内部结构的有序性
细胞质基质
成分
水、无机盐、脂质、糖类、 氨基酸、核苷酸和多种酶
状态 呈不断流动的胶质状
作用 活细胞代谢的主要场所
保护、物质交换、能量转换、 信息传递等;提供酶的附着位点; 使细胞内区域化
功能
生物膜系统
研究意义 实验:体验制备细胞膜的方法
纤维素和果胶 成分
保护和支持作用 功能
高中生物知识结构网络图(完整版)
预祝2017级高考学子梦想实现。
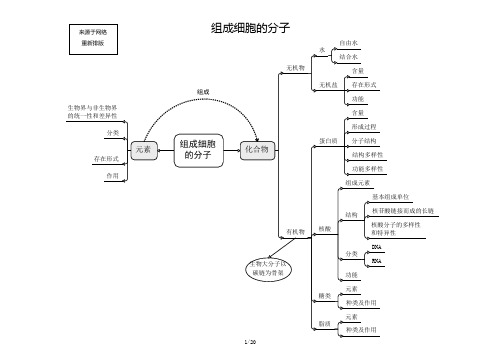
高中生物知识结构网络图第一单元 生命的物质基础和结构基础(细胞中的化合物、细胞的结构和功能、细胞增殖、分化、癌变和衰老、生物膜系统和细胞工程)1.1化学元素与生物体的关系1.2生物体中化学元素的组成特点1.3生物界与非生物界的统一性和差异性1.4细胞中的化合物一览表1.5蛋白质的相关计算设 构成蛋白质的氨基酸个数m ,构成蛋白质的肽链条数为n ,构成蛋白质的氨基酸的平均相对分子质量为a , 蛋白质中的肽键个数为x , 蛋白质的相对分子质量为y ,控制蛋白质的基因的最少碱基对数为r ,则 肽键数=脱去的水分子数,为 n m x -= …………………………………①蛋白质的相对分子质量 x ma y 18-= ………………………………………②或者 x a ry 183-=………………………………………③ 1.6蛋白质的组成层次1.7核酸的基本组成单位1.8生物大分子的组成特点及多样性的原因1.9生物组织中还原性糖、脂肪、蛋白质和DNA的鉴定1.10选择透过性膜的特点1.11水被选择的离子和小分子其它离子、小分子和大分子亲脂小分子高浓度——→低浓度不消耗细胞能量(ATP)离子、不亲脂小分子低浓度——→高浓度需载体蛋白运载消耗细胞能量(ATP)1.12线粒体和叶绿体共同点1、具有双层膜结构2、进行能量转换3、含遗传物质——DNA4、能独立地控制性状5、内含核糖体6、有相对独立的转录翻译系统7、能自我分裂增殖1.13真核生物细胞器的比较1.14细胞有丝分裂中核内DNA、染色体和染色单体变化规律1.15理化因素对细胞周期的影响注:+ 表示有影响1.16细胞分裂异常(或特殊形式分裂)的类型及结果1.18已分化细胞的特点 1.19分化后形成的不同种类细胞的特点G 21.20分化与细胞全能性的关系1.211.22癌细胞的特点分化程度越低全能性越高,分化程度越高全能性越低分化程度高,全能性也高分化程度最低(尚未分化),全能性最高扁平梭形 球形成纤维细胞癌变如癌细胞膜糖蛋白减少,细胞黏着性降低,易转移扩散。
野生大豆资源的研究与利用

野生大豆资源的研究与利用胡小梅;林红;刘广;阳;杨雪峰;刘丽艳;张俐俐;张必弦;朱延明;来永才;李炜;李琬;毕影东;肖佳磊;齐宁【摘要】对野生大豆资源的分布、起源、分类、研究与利用5方面进行了阐述,并从野生大豆的产量品质、生理性状、杂种优势的优良特性及其在生物技术中的应用角度介绍了野生大豆资源研究利用的情况.%Thedistribution,origin,classifying,studies and utilizes of wild soybean(dycine soja)were expounded,and the studies and utilization condition of wild soybean were introduced from the angles of yield,quality,physical properties,hybrid advantages and biotechnology apply.【期刊名称】《安徽农业科学》【年(卷),期】2011(039)022【总页数】3页(P13311-13313)【关键词】野生大豆;资源;草本植物【作者】胡小梅;林红;刘广;阳;杨雪峰;刘丽艳;张俐俐;张必弦;朱延明;来永才;李炜;李琬;毕影东;肖佳磊;齐宁【作者单位】东北农业大学生命科学学院,黑龙江哈尔滨150030;黑龙江省农业科学院作物育种研究所,黑龙江哈尔滨150086;黑龙江省农业科学院作物育种研究所,黑龙江哈尔滨150086;黑龙江省农业科学院作物育种研究所,黑龙江哈尔滨150086;黑龙江省农业科学院作物育种研究所,黑龙江哈尔滨150086;黑龙江省农业科学院生物技术研究所,黑龙江哈尔滨150086;黑龙江省农业科学院生物技术研究所,黑龙江哈尔滨150086;东北农业大学生命科学学院,黑龙江哈尔滨150030;黑龙江省农业科学院博士后工作站,黑龙江哈尔滨150086;东北农业大学生命科学学院,黑龙江哈尔滨150030;黑龙江省农业科学院博士后工作站,黑龙江哈尔滨150086;黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086;黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086;黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086;黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086;黑龙江省农业科学院耕作栽培研究所,黑龙江哈尔滨150086;黑龙江省农业科学院作物育种研究所,黑龙江哈尔滨150086【正文语种】中文【中图分类】S643.7野生大豆(Glycine soja)多为一年生或多年生草本植物,是栽培大豆(Glycine max)的祖先物种[1]。
高中生物必修二知识点整理思维导图
高中生物必修二 知识点整理第一章 遗传因子的发现相对性状概念同一种生物的同一种性状的不同表现类型显性性状与隐性性状性状分离:在杂种后代中出现不同于亲本性状的现象显性基因与隐性基因等位基因:决定1对相对性状的两个基因 (位于一对同源染色体上的相同位置上)纯合子与杂合子纯合子:由相同基因的配子结合成的合子发育成的个体(能稳定的遗传,不发生性状分离)杂合子:由不同基因的配子结合成的合子发育成的个体如Aa (不能稳定的遗传,后代会发生性状分离)表现型与基因型表现型:指生物个体实际表现出来的性状。
分显性和隐形基因型+环境→表现型基因型:与表现型有关的基因组成杂交与自交测交:让F1与隐性纯合子杂交(可用来测定F1的基因型,属于杂交)孟德尔实验成功的原因正确选用实验材料豌豆是严格自花传粉植物(闭花授粉),自然状态下一般是纯种具有易于区分的性状由一对相对性状到多对相对性状的研究(从简单到复杂)对实验结果进行统计学分析严谨的科学设计实验程序:假说-演绎法第二章 基因和染色体的关系减数分裂减数分裂的概念进行有性生殖的生物,在产生成熟生殖细胞时进行的染色体数目减半的细胞分裂染色体只复制一次,而细胞分裂两次。
结果是成熟生殖细胞中的染色体数目比原始生殖细胞减少一半减数分裂的过程精子形成的过程(场所:精巢/睾丸)精原细胞曲细精管中初级精母细胞(减数第一次分裂)间期:染色体复制(包括DNA复制和蛋白质的合成)前期:同源染色体联会;四分体时期;四分体中的姐妹染色单体发生交叉互换中期:排列在赤道板上后期:同源染色体分开;非同源染色体自由组合末期:细胞质分裂,形成两个子细胞减数第二次分裂(无同源染色体)前期:染色体排列散乱中期:每条染色体的着丝粒都排列在细胞中央的赤道板上后期:姐妹染色单体分开,成为两条子染色体。
并分别移向细胞两极末期:细胞质分裂,每个细胞形成2个子细胞,最终共形 成4个子细胞精细胞(1个精原细胞产生4个精细胞)精子:由精细胞分化呈蝌蚪状,头部含有细胞核,尾长,能够摆动卵细胞形成过程(场所:卵巢)卵原细胞初级卵母细胞次级卵母细胞和极体卵细胞和极体减数第一次分裂减数第二次分裂精子和卵细胞形成过程的异同不同点精子形成部位精巢(哺乳动物称睾丸)过程有变形期子细胞数一个精原细胞形成4个精子卵细胞形成部位卵巢过程无变形期子细胞数一个卵原细胞形成1个卵细胞+3个极体相同点精子和卵细胞中染色体数目都是体细胞的一半注意点同源染色体形态、大小基本相同一条来自父方,一条来母方精原细胞和卵原细胞的染色体数目与体细胞相同。
高中生物思维导图必修三部分

必修三专题一内环境及稳态盐津二中2019届高考生物基础内容强化必修二育种及必修三(黄照南整理) 育种又叫IXU 勰U 技剜®4拼技技术・它J 酗黒人 ffJMIK,覺 种I H ]以 修饰A'&,称阚朋 州:物幽跑内,鈔哋 攻堆牛剜適f 我狀._ * _______________________________________亦概念 生产辛丙I 用踌耐1 ---------------射训巴©足和IIiff irjx^iisr附作物計胖乩啪懾墓降悔m 昔材宦的动物忧内屁古弁布tsss卫虫方面核酸内瀨斗作用攻畅性|抻特宦的峡ENT 乍列.并㈱址的如沏剽悚X"WW- 伏晶业-朋 DNA SHtWWi—I//f获刑分卿-谢桂炎的”悝址坐an 临巫如鞠;Hl DDT 的mt 甘为人瓷卩廉勘询tt 挪柬刊¥1極物理因童1ttV Zfit. mm起的考查。
緞处圖面在业姑方附骂.・.S 臓金性厂極wim樓计牖」「中变因索J-町臭矗哭变顾率:能较甌时風MS 性軟;igjaao打好吆需处理大*的 电物材料.MHjjW 堵霄飾建人黄需要 肢片英鱼师优帥种〈塞因工程》从杂交育种到基因工程诱变 育种W,育种殽肌目的卑閑~tr科的缈与世戟広瞄舍 稚H 骑革X,人经讳桂庭—© -------------日的輒刃的椅鶴*丧追-f 育种过程尸 将两个亲甬杂交.那到F.再娄 衣自更逐牝选弁・霞到紂淞 状不幟q 讲离的理酚E 种丽N 能嫌谿优联状如一体L 髀牧业]\抽*撤优良跆的时,重点关注五种育种的原理、优缺点以及将育种和基因的自由组合定律整合在人体器官、硫 ftg 別地迖『缈特松态的调代机制机休进行正當生命 活幼的邨箜条件机体进打IE 常4渝常型的羽强家丼倍内細胞柑外环*ft 遊行物瓯空换过和细胞外液的 理化性质厂定又\界皆卩祥烦匱栈时忒的吸引力港透圧J-灌透压丄 的大小[31 6t 时为 TT (Wu,)丄姿4无机盐和黄 血粉抽的諒曲的*办艾•址軽外灌津透d —V“户…工:肖汉加「- W 讯:「-址持因索5 o一地kt 曲比u实暫r 业物呻 借持pH 沁 存祁S神毎BH*□小Illg NilIjRVN*刑 食i 止就人血灿0衽(^)Na\艸WCn,警枸应 *護中内煤沖微聚・H 有 —定荊視神廉力」 厂人体细屜外液的珮质融址持371C 左右 o_MSfil 幼年〉成年> V\^; k4>!B 性专题二动物和人的生命活动调节专题三植物激素调节'檄索诚节的发现Z 总外界博韋力主 itlMHh 弟均対亠 鼻勰井坐绘』、匀勺 一曲性迄型'唱嗾力向的头界赫潦「为勺尽間定向站唧 柚物皿好地适应环境[[腳年,达尔文实叭 I9IH 年,加檬齐©・”统的调芳通过神经系的调节通过激素目加匸舉的叫取力性.墨的 甘社力性命活动的调节〔动物和人体生Hffit4U/J植物的廉性、 向性运动1松性葩件用机理彳喘檢養的发叹(射咖肪的烟丸]迥%啲实屮卅科I 岀2勺,工札富的■认)概念及种类激素调W 的轻点神经调ff 与体液调节的比较寒净*克越环班中的你悽调节液训节的关系神经调节与体人体的水捷 半fti 调W 両屯応盐轲节即又歩下丘脑的功濫 1. 血糖调节、体温调节、水盐平衡调节一直都是神经一体液调节中的重点和热点内容。
全套高中生物思维导图word清晰打印版

思维导图高中生物
快乐学习,一目了然
全套高中生物思维导图(清晰打印版)
•
•
•
•
•
•
•
•
•
例
•第2节生物膜的流动镶嵌
模型
•第3节物质跨膜运输的方
式
•第5章细胞的能量供应
和利用
•第1节降低化学反应活化
能的酶
•第2节细胞的能量“通
货”──ATli
•第3节ATli的主要来源
──细胞呼吸
•第4节能量之源──光与
光合作用
•第6章细胞的生命历程•第1节细胞的增殖
•第2节细胞的分化•第3节细胞的衰老和凋亡•第4节细胞的癌变
(选学)
•第5章基因突变及其他
变异
•第1节基因突变和基因重
组
•第2节染色体变异
•第3节人类遗传病
•第6章从杂交育种到基
因工程
•第1节杂交育种与诱变育
种
•第2节基因工程及其应用
•第7章现代生物进化理
论
•第1节现代生物进化理论
的由来
•第2节现代生物进化理论
的主要内容
定性
•第1节生态系统的结构
•第2节生态系统的能量流
动
的洗涤效果
•课题3 酵母细胞的固定
化
•专题5 DNA和蛋白质技
术
•课题1 DNA的粗提取与
鉴定
•课题2 多聚酶链式反应
扩增DNA片段
•课题3 血红蛋白的提取
和分离
•专题6 植物有效成分的
提取
•课题1 植物芳香油的提
取
•课题2 胡萝卜素的提取。
大豆遗传连锁图构建

阳!,
摘要:利用大豆栽培品种科丰 ! 号和南农 !!’( ) " 杂交得到的重组近交系 *+,-./,通过 ,012、33,、,425 和 4012 6 种分子标记的遗传连锁分析,构建了包含 "6 个连锁群、由 7%" 个遗传标记组成的大豆较高密度连锁图谱,该图谱覆盖 " ’"$ # 789,平均图距 " # %89。33, 标记 的 多 态 性 较 高 ,在 基 因 组 中 的 位 置 相 对 稳 定 ,可 以 作 为 锚 定 标 记 ,有 利 于 连 锁 群 的 归 并和不同图谱的比较整合;而 4012 标记对于增加图谱密度效率较高,但其容易出现聚集 现象,从而造成连锁群上有很大的空隙(:;<)。另外,在连锁群中有 "! # 7 = 的分子标记出 现偏分离。该图谱为基因定位、比较基因组学和重要农艺性状的 >?1 定位等研究打下了 基础。 关键词:大豆;遗传图谱;分子标记;基因组分析 中图分类号:>7&& 文献标识码:4 文章编号:$’7% ) 6!7(" "$$!)!! ) !$&! ) !!
张德水等[6]以栽培品种长农 . 为母本和半野生品种新民 6 为父本杂交得到 +5 群体, 构建了国内第一张大豆的分子标记连锁框架图。其中 *+,- 标记 64 个,*%-D 标记 $ 个, 组成 5" 个连锁群,覆盖大豆基因组长度为 ! ..6 H $78。刘峰等[C]用以此为组合的重组自 交系群体(*G,),构建了包含 5." 标记的遗传图谱。这两张图谱标记密度较低,不能满足 大豆基因组研究的需要。贺超英等用 99* 标记对本文所用的群体进行了遗传结构的合 理性评估[$],本文用 *+,-、*%-D、99* 和 %+,- 标记对重组近交系 LM*G/N 进行了分析,构 建了较高密度的遗传连锁图谱,并定位了与大豆抗病性和重要农艺性状有关的分子标记。
第5代加强版

理论研究 预期成果
1
2
3
4
1.在实验中筛选出比 较合适的提取大豆籽 粒中蛋白含量的方法。
2.筛选出一定数量多 态性较好引物(引物 用于PCR技术),为 后续研究奠定基础。
3.结合前人研究成果 进一步分析大豆蛋白 含量的影响因素。
4.完成或超常完成相 关实验,使学生相关 知识层面有所提高, 完成项目结题报告。
11
经费预算
12
经费预算
本项目总经费预算是1550元
1.实验材料500元。 2.劳务费加管理费850元。 3.其它200元。
13
个人总结
14
在本项目中我们拥有的优势和不 足
在能力有所欠缺的 情况下我们可能会 产生一些大的实验 误差,所以以我们 也要不断进行重复 实验。
我们有专业的老 师从国外学习, 在项目期间会进 行相关辅导
本项目研究的目的和 意义
6
基本概念 项目意义
根据大豆蛋白质 QTL 定位的结果,大豆中拥有效应较大的主 效 QTL 位点,而且加性效应占很大比例,有助于通过标记辅助选 择培育高蛋白品种。分别位于 20 号染色体( I 连锁群) 、15 号染 色体( E 连锁群) 和 6 号染色体( C2 连锁群) 上的 QTL 区段A688 - Satt239、Satt151 - Satt045、Satt281 - Satt291在多个环 境和多个群体中均检测到,且最高贡献率达 56% 。通过标记辅 助选择,将这些位点的有利等位基因聚合到同一品种中,从而提 高品种的蛋白质含量,将是可行的。有些位点的有利等位变异存 在于低蛋白亲本中,通过标记辅助选择,有可能将其导入目标品 种,提高其蛋白含量,也就是说低蛋白亲本在分子育种中也可能 作为有利等位基因的供体。这些都是常规育种难以达到的。
中国野生大豆遗传多样性中心

第26卷第5期作 物 学 报V o l.26,N o .52000年9月A CTA A GRONOM I CA S I N I CASep .,2000中国野生大豆遗传多样性中心X董英山1,2 庄炳昌1 赵丽梅1 孙 寰1 张 明1 何孟元2(1吉林省农业科学院,吉林公主岭,136100;2东北师范大学,吉林长春,130024)提 要 本文采用群体遗传学方法,研究分析了6172份中国野生大豆资源的数量、遗传多样性指数(Shannon 指数)、12个性状的综合变异系数及其地理分布。
结果表明:①42~47°N ×122~127°E 的东北中南部野生大豆资源分布极广、遗传多样性丰富、综合变异系数高;②34~35°N ×113~115°E 、38~39°N ×113~115°E 和34~35°N ×107~109°E 的黄河中下游和秦岭山区野生大豆次之;③26~27°N ×119~121°E 的东南沿海地区野生大豆遗传多样性、综合变异系数均很高,但分布较少。
根据遗传多样性、综合变异系数及资源的地理分布,提出了中国野生大豆遗传多样性中心及多样性扩散,据此推测了中国野生大豆的起源模式,为栽培大豆的起源提供了一个重要依据。
关键词 野生大豆;遗传多样性;富集中心;起源The Geneti c D i versity Cen ters of Annua l W ild Soybean i n Ch i n aDON G Y ing 2Shan 1,2 ZHUAN G B ing 2Chang 1 ZHAO L i 2M ei 1 SUN H un 1 ZHAN G M ing1H E M eng 2Yuan2(1J ilin A cad e m y of A g ricultural S ciences ,136100,Gongzhuling ,J ilin ;2N orth E ast N or m al U niversity ,130024,Changchun ,J ilin )Abstract T he genetic diversity ,comp rehen sive coefficien t of variability of 12traits and thegeograph ical distributi on of 6172accessi on s co llected from Ch ina w ere studied by quan titativegenetics in th is paper .It show ed that :①A nnualw ild s oybean in 42~47°N ×122~127°E p l o tsare the richest in accessi on s ,genetic diversity and comp rehen sive coefficien t of variability .②A nnual w ild s oybean in 34~35°N ×113~115°E ,38~39°N ×113~115°E and 34~35°N ×107~109°E of Yell ow R iver V alley and Q in ling M oun tain s take second p lace .③T he geneticdiversity and comp rehen sive coefficien t of variability of annual w ild s oybean w ere h igher in 26~27°N ×119~121°E of s outheast coast ,but the distributi on of accessi on s w as less.T he genetic diversity cen ters ,s p read modes of genetic diversity w ere put fo r w ard acco rding to the distributi on of accessi on s ,genetic diversity and comp rehen sive coefficien t of variability .F rom th is the o rigin mode of w ild s oybean in Ch ina w as inferred .T he results p rovided an evidence fo r o rigin of cultivated s oybean in Ch ina .Key words Genetic diversity cen ters ;O rigin ;A nnualw ild s oybean野生大豆为G ly cine W illd 属,S oja 亚属植物,分布于东亚中北部的日本、朝鲜、俄罗斯远东地区及中国[1]。
大豆起源

一、栽培大豆起源于中国是国内外学者所公认的栽培大豆起源于中国, 早已为国内外学者所公认。
正像《美国大百科全书》所指出的:“中国古文献认为, 在有文献记载以前, 大豆便因营养价值高而被广泛地栽培。
同时在公元2000 年前大豆便被看作是最重要的豆科植物。
”《苏联大百科全书》更明确指出:“栽培大豆原产于中国, 中国在5000 年前就开始栽培这个作物, 并由中国向南部及东南各国传播, 于16 世纪进入欧洲。
”苏联着名的学者瓦维洛夫在《主要栽培植物的世界起源中心》一书中指出:“第一个最大的独立的世界农业发源和栽培植物起源地是中国的中部和西部山区及其毗邻的低地。
”在“栽培植物的中国起源中心”中第10 项作物就是“大豆”。
日本学者星川清亲在《栽培植物的起源与传播》一书中也指出:“大豆的原始野生种起源于中国北部……从中国的东北到西伯利亚、黑龙江流域, 古代人们就已开始栽培, 不久又传到中国内地, 作为五谷之一而受到人们的重视。
”日本的栽培大豆, “似乎是在弥生初期作栽培作物从中国传入日本”。
栽培大豆在世界各地的传播情况是:大约在公元前200 年从中国引入朝鲜, 而后自朝鲜引至日本(日本学者永田忠男说)。
而日本南部的大豆, 可能直接由商船自华东一带引去。
大豆引至欧美则是近代之事。
●1712 年德国植物学家首次从日本将大豆引入欧洲。
●1740 年法国传教士曾将中国大豆引至巴黎试种。
●1790 年英国皇家植物园首次试种大豆。
●1873 年奥地利的维也纳博览会上试种从中国和日本引入的19 个大豆品种, 其中有4个品种结粒。
●1840 年美国从中国和日本引入大豆试种。
●1822 年美国种植大豆主要作饲料作物。
●1910 年美国已经引入280 个中国大豆品种。
●1915年美国首次用本国产的大豆榨油。
●1929 年美国已种植大豆400 万亩。
●1941 年二战期间, 美国由于缺乏食用油, 开始大规模种植大豆。
到1944 年美国大豆种植面积达6000 多万亩。