阅读材料:细胞膜的结构模型
初中生物细胞膜结构(含学习方法技巧、例题示范教学方法)

初中生物细胞膜结构第一篇范文细胞膜,作为生命体系中最基础的组成部分,承担着维持细胞内外环境平衡、物质交换和信息传递等至关重要的功能。
初中生物教学中,对细胞膜结构的认识,有助于学生理解生命现象的本质,培养科学素养。
细胞膜的组成细胞膜主要由脂质、蛋白质和糖类三种生物大分子构成。
其中,脂质是细胞膜的基本骨架,占据了细胞膜的绝大部分。
蛋白质则承担着细胞膜的功能性作用,如通道蛋白、受体蛋白等。
糖类则与脂质和蛋白质结合,形成糖脂和糖蛋白,参与细胞识别和信号传递等过程。
细胞膜的结构细胞膜的结构模型有多种,如流动镶嵌模型、单位膜模型等。
流动镶嵌模型认为,细胞膜中的脂质和蛋白质都是可以流动的,形成了动态的、非均质的结构。
单位膜模型则认为,细胞膜由两层脂质分子层组成,中间夹杂着蛋白质。
细胞膜的功能细胞膜的功能可以概括为维持细胞内外环境平衡、物质交换和信息传递三个方面。
首先,细胞膜通过选择性通透性,控制细胞内外物质的交换,保证细胞内部环境的稳定。
其次,细胞膜上的蛋白质通道和载体,参与了多种物质的主动运输和被动扩散。
最后,细胞膜上的受体蛋白,能够接收外部信号,引发细胞内部的生化反应,调控细胞生长、分化和死亡等过程。
教学启示在初中生物教学中,教师应当注重细胞膜结构与功能的结合,通过生动的实例和模型,帮助学生理解细胞膜的重要性和复杂性。
同时,鼓励学生进行实验和探究,提高学生的实践能力和科学素养。
细胞膜的研究,是现代生物科学的前沿领域之一。
通过对细胞膜结构与功能的研究,我们可以深入了解生命现象的本质,为疾病的诊断和治疗提供新的思路和方法。
希望初中生能够在学习中,感受到生物科学的魅力,激发对生命科学的热爱。
第二篇范文:初中学生学习方法技巧对于初中学生而言,学习生物细胞膜结构这一知识点,不仅需要理解抽象的理论知识,还需要掌握一定的学习方法和技巧。
本文旨在为初中学生提供一套系统化的学习策略,以提高他们对细胞膜结构的理解和记忆。
学习前的准备在学习细胞膜结构之前,学生应该具备一定的基础知识,如细胞的结构和功能、脂质和蛋白质的基本概念等。
第1节:细胞质膜的结构模型

第一节:细胞质膜的结构模型细胞质膜的结构模型模型(整体)基本结构磷脂双分子层镶嵌部分膜蛋白如何镶嵌:脂筏p84(看①)结构决定功能蛋白的类型、蛋白分布的不对称性及其与脂分子的协同作用赋予生物膜各自的特性与功能膜的性质二维溶液:生物膜可以看成是双层脂分子中嵌有蛋白质的二维溶液。
但是膜脂和蛋白质在一定程度上受到限制。
p85膜脂分三类(看②)甘油磷脂、鞘脂、固醇运动方式(4种)沿膜平面的侧向运动、脂分子围绕轴心的自旋运动、脂分子尾部的摆动、双层脂分子之间的翻转运动脂质体利用脂分子的性质制备的人工膜,可有效导入DNA 或药物p88膜蛋白分三类(看③)外周膜蛋白、整合膜蛋白、脂锚定蛋白内在膜蛋白与膜脂结合方式与生化内容α螺旋例子一致(看④)去垢剂是一端亲水另一端疏水的两性小分子,SDS 、Triton X-100(p92)①脂筏:以甘油磷脂为主体的生物膜上,胆固醇、鞘磷脂等形成相对有序的脂相,如同漂浮在脂双层上的“脂筏”一样,载着执行某些生物学功能的各种膜蛋白。
脂筏最初可能在内质网或高尔基体上形成,最终转移到细胞膜上。
有些脂筏与膜下细胞支架蛋白交联。
推测一个直径100nm的脂筏可载有600个蛋白分子。
②膜脂分类:④内在膜蛋白的结合方式1、内在膜蛋白的跨膜结构域与脂双层分子的疏水核心的相互作用;(最主要的)2、内在膜蛋白的跨膜结构域两端携带正电荷的氨基酸或者负电荷的氨基酸,正电荷则与极性头(带负电荷)形成离子键,带负电荷则通过Ca2+、Mg2+等阳离子与极性头(带负电荷)相互作用;3、通过细胞质基质一侧的半胱氨酸残基上共价结合的脂肪酸分子插入到膜双层之间。
P91(α螺旋外侧非极性链,内测极性链,形成跨膜通道(这里的内侧和外侧值得是α螺旋自身,并非伸向细胞质和细胞外液的部分);孔蛋白还具有β折叠片→生化内容)。
细胞膜的结构及探索历程

二、生物膜的成分:
(一)膜脂: 生物膜的基本组成成分,主要包括的类型有: 磷脂:膜脂的基本成分,含量>50% 糖脂:含量<5% 胆固醇:含量 <1/3
Figure. The parts of a phospholipid molecule. Phosphatidylcholine (PC,磷脂酰胆碱), represented schematically (A), in formula (B), as a
于水。
与膜结合紧密,用去垢剂破坏膜结构才可 以分离出来。
占全部膜蛋白的70-80%。 例如:MN血型糖蛋白
Figure. A typical single-pass transmembrane protein. Note that the polypeptide chain traverses the lipid bilayer as a right-handed a helix and that the oligosaccharide chains and disulfide bonds are all on the noncytosolic surface of the membrane. Disulfide bonds do not form between the sulfhydryl groups in the cytoplasmic domain of the protein because the reducing environment in the cytosol maintains these groups in their reduced (-SH) form.
• 脂质体: 膜蛋白在脂双层二维溶液中的运动是自发的热运动,不需提供能量。
GPI锚定膜蛋白 内在膜蛋白的跨膜结构域是与膜脂结合的主要部位,或者形成α螺旋,或者形成β折叠片结构。
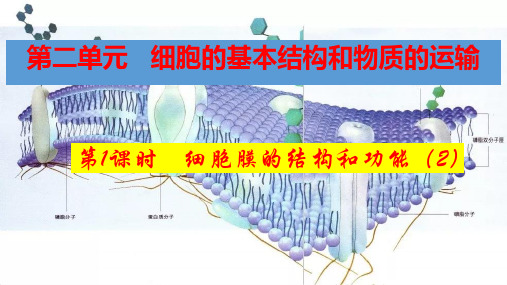
细胞膜的结构和功能- (解析版)

第3章细胞的基本结构第1节细胞膜的结构和功能一、【学习目标】1.从系统与环境关系的角度,阐释细胞膜作为系统的边界所具有的功能。
2.分析细胞膜组成成分与结构的关系,说明细胞膜结构的物质基础,概述流动镶嵌模型的主要内容。
3分析对细胞膜成分与结构的探索历程,认同科学理论的形成是一个科学精神、科学思维和技术手段结合下不断修正与完善的过程。
自主学习一、细胞膜的功能1.将细胞与分隔开,保障细胞内部环境的相对稳定。
2.控制物质进出细胞。
(1)一般来说,细胞需要的可以从外界进入细胞;细胞,不容易进入细胞。
(2) 等物质在细胞内合成后,分泌到细胞外,细胞产生的__也要排到细胞外;但是,细胞内却不会轻易流失到细胞外。
(3)细胞膜的控制作用是_,环境中一些有可能进入;有些进行细胞间的。
3.进行细胞间的信息交流(1)通过化学物质传递信息。
(2)通过直接接触传递信息。
(3)通过细胞通道传递信息。
4.对细胞膜成分的探索(1)1895年欧文顿发现细胞膜对不同物质的通透性不一样:溶于脂质的物质更容易穿过细胞膜,据此推测细胞膜是由组成的。
(2)科学家利用哺乳动物的红细胞制备出细胞膜,得知组成细胞膜的脂质有磷脂和胆固醇,其中含量最多。
(3)1925年,荷兰科学家发现,细胞膜中的磷脂分子排列为连续的。
(4)1935年,英国学者丹尼利和戴维森研究细胞膜的张力,发现细胞表面张力明显低于油-水界面的表面张力,据此推测细胞膜中除含有脂质分子外,可能还含有。
二、对细胞膜结构的探索1.细胞膜的成分(1)细胞膜主要由脂质和蛋白质组成,此外,还有少量的。
在组成细胞膜的脂质中,磷脂最丰富,此外还有少量的胆固醇。
(2)蛋白质在细胞膜行使功能方面起重要作用,功能越复杂的细胞膜,蛋白质的种类与数量就越。
2.对细胞膜结构的探索历程三、流动镶嵌模型的基本内容1.基本内容:(1)基本骨架:。
(2)蛋白质的分布:以不同方式镶嵌在磷脂双分子层中。
2.结构特点:具有一定的。
高一生物必修1_分子与细胞_阅读材料:细胞膜的结构模型

细胞膜的结构模型19世纪中叶K.W.Mageli发现细胞表面有阻碍染料进入的现象,提示膜结构的存在;1899年E.Oveiton发现脂溶性大的物质易入胞,推想应为脂类屏障。
1925年荷兰人E.Gorter和F.Grendel用丙酮抽提红细胞膜结构,计算出红细胞膜平铺面积约为其表面积的两倍,提出脂质双分子层模型.成立前提:a.红细胞的全部脂质都在膜上;b.丙酮法抽提完全;c.RBC平均表面积估算正确。
(70%~80%偏低);40年后Bar重复这一试验发现红细胞膜平铺面积应不是70%~80%,而是1.5倍还有蛋白质表面,同时干膜面积是99μm2,湿膜面积则为145μm2。
两项误差相抵,结果基本正确。
根据细胞的生理生化特征,曾先后推测质膜是一种脂肪栅、脂类双分子层和由蛋白质-磷脂-蛋白质构成的三夹板结构。
同时电镜观察也证实质膜确实呈暗-明-暗三层结构。
随后冷冻蚀刻技术显示双层膜中存在蛋白质颗粒;免疫荧光技术证明质膜中蛋白质是流动的。
据此S.J.Singer等人在1972年提出生物膜的流动镶嵌模型,结构特征是:生物膜的骨架是磷脂类双分子层,蛋白质分子以不同的方式镶嵌其中,细胞膜的表面还有糖类分子,形成糖脂、糖蛋白;生物膜的内外表面上,脂类和蛋白质的分布不平衡,反映了膜两侧的功能不同;脂双层具有流动性,其脂类分子可以自由移动,蛋白质分子也可以在脂双层中横向移动。
尽管目前还没有一种能够直接观察膜的分子结构的较为方便的技术和方法,但从研究中30年代以来提出了各种假说有数十种,其中得到较多实验事实支持而目前仍为大多数人所接受的是美国的S.J.Singer和G.L.Nicholsom于1972年提出的流体镶嵌模型(fluid mosaic model)。
这一假想模型的基本内容是:膜的共同结构特点是以液态脂质双分子层为基架,其中镶嵌着具有不同分子结构、因而也具有不同生理功能的蛋白质,后者主要以α-螺旋或球型蛋白质的形式存在。
细胞膜的结构模型与细胞功能

02
在细胞膜上载体的帮助下,通过消耗ATP,将某种物质逆浓度梯度进行转运的过程。
03
逆浓度梯度转运;
04
耗能(ATP) 。
05
细胞生理
主动转运 (据提供能量方式)
01
02
03
04
原发性主动转运
继发性主动转运
直接利用ATP水解产生的能量进行离子的跨膜转运。如Na+的转运
05
能量不是直接来自ATP的水解,来自膜外的高势能Na+。
1、片层结构模型
细胞的表面张力显著低于油-水界面的表面张力,因此,细胞膜不可能是单纯由脂类构成的,可能还吸附有蛋白质。
1935年J. Danielli和H. Davson提出了第一个膜的分子结构模型——片层结构模型 。
2、单位膜模型
1959年,J.D.Robertson用电镜观察细胞膜,发现细胞膜呈三层式结构。内外两侧为电子密度高的暗线,中间为电子密度低的明线,即所谓“两暗一明”,进而提出单位膜模型。
20世纪初,Irving Langmuir 将红细胞的脂提取后铺展在Langmuir 水盘(Langmuir Trough)的水面上,研究了脂的展层行为,提出脂单层的设想。
1925 年,E. Gorter & F. Grendel 用有机溶剂提取了人类红细胞质膜的脂类成分,将其铺展在水面,测出膜脂展开的面积二倍于细胞表面积,因而推测细胞膜由双层脂分子组成。
(3)卵磷脂与鞘磷脂比值:
卵磷脂越高流动性越强。
(4)膜蛋白的影响:
卵磷脂
脂肪酸不饱和程度高,相变温度低。
鞘磷脂
脂肪酸饱和程度高,相变温度高。
(二)膜蛋白分子在质膜中的运动
利用细胞融合技术观察蛋白质运动
周辉校本研修:细胞膜的分子结构模型2008.09.05

细胞膜的分子结构模型――生物教研组周辉整理根据大量的实验研究和分析,人们提出了关于多种生物膜的分子结构模型。
1.片层结构模型或三明治模型:1935年由J.D. Davson和J. Danielli提出“双分子片层”结构模型,或三明治模型。
他们认为膜的骨架是脂肪形成的脂双结构,但是,为了降低活性细胞的表面张力,他们假定在脂双层的内外两侧都有一层蛋白质包被,即蛋白质-脂-蛋白质的三层结构,内外两层的蛋白质层都非常薄。
他们还认为,蛋白质层是非折叠、完全伸展的肽链形式包在脂双层的内外两侧。
这一模型提出后,很快又进行了一些细微的修改,认为在腊上有一些二维伸展的孔,孔的表面也是由蛋白质包被的,这样孔就具有极性,提高了水对膜的透性。
该模型是第一次用分子术语描述的膜结构,并将膜结构同所观察到的生物学性质联系起来,对后来的研究有很大的启发。
(2).单位膜模型随着电镜的问世和技术的不断改良,1959年Robertson用电子显微镜镜对各种膜结构进行了详细研究,在电子显微镜下发现细胞膜是类似铁轨结构(railroadtrack),两条暗线被一条明亮的带隔开,显示暗-明-暗的三层,部厚度为7.5nm,中间支为3.5 nm,内外两层各为2 nm。
并推测:暗层是蛋白质,透明层是脂,建议将这种结构称为单位膜。
单位膜与片层结构膜有许多相同之处,最重要的修改是膜脂双分子层内外两侧的蛋白质存在的方式不同。
单位膜模型强调的是蛋白质为单层伸展的β折叠片状,而不是球形蛋白。
另外,单位膜模型还认为膜的外侧表面的膜蛋白是糖蛋白,而且膜蛋白在两侧的分布是不对称的。
这一模型能够解释细胞质膜的一些基本特性。
(3).流动镶嵌模型1966年,lenarnard和Singer发现膜蛋白的氨基酸中的30%是螺旋形式弯曲存在,而不像片层结构模型那样完全伸展在两侧。
若将膜蛋白展开后铺展在膜的外层,其厚度是7~8 nm的几倍,显然与所观察的结果不符。
另外从Singer的观察,如果蛋白质是完全伸展,则需要消耗大量的能量以维持疏水氨基酸暴露在水性环境中,事实上蛋白质总是倾向于折叠,让亲水和疏水的不同基团处于不同折环境中,以维持最低的能量状态。
高中生物 细胞膜的分子结构假说和模型

细胞膜的分子结构假说和模型关于膜的结构,从2019开始一直到现在,科学家们提出了很多假说和模型。
下面举几个比较流行的模型加以说明。
1.单位膜模型这种模型于1935年提出,到20世纪50年代加以修正,随后经罗伯特桑(Robertson)的电镜观察加以完善。
这种模型表示,细胞膜由脂质双分子层及在其内外两侧各覆盖一层蛋白质所组成,脂质分子相互平行,与膜垂直。
蛋白质是以β-折叠形式结合在膜的内外两侧,形成网状。
从而形成蛋白质――磷脂――蛋白质的三层结构。
罗伯特桑于1959年指出,所有生物膜的厚度基本上是一致的,这种三层结构的膜普遍存在于细胞中,他叫这样的膜为单位膜。
但到20世纪60年代以后,由于应用了一系列新技术,科学家证实膜的脂质双分子层中也有蛋白质颗粒,并证实膜蛋白主要不是β-折叠结构,而是α-螺旋结构等。
科学家根据这些事实,对生物膜的单位膜模型理论提出了修正。
单位膜模型的主要不足在于:把膜结构描述成静止的、不变的,这显然与膜功能的多样性相矛盾。
2.流动镶嵌模型这一模型是S.J.Singer和G.Nicolson于1972年通过对已有的模型进行修正而提出的,这是细胞生物学的重要进展之一。
科学家发现细胞膜不是静态的,而是膜中的脂质和蛋白质都能自由运动。
这种模型叫做流动脂质—球蛋白镶嵌模型,现已广泛被接受。
这是个动态模型,表示细胞膜是由脂质双分子层和镶嵌着的球蛋白分子组成的,有的蛋白质分子露在膜的表面,有的蛋白质分子横穿过脂质双分子层。
这种模型主要强调的是,流动的脂质双分子层构成了膜的连续体,而蛋白质分子像一群岛屿一样无规则地分散在脂质的“海洋”中。
后来,不少实验都证实膜脂的“流动性”是生物膜结构的基本特性之一,因此这种模型比较普遍地被大家所接受和支持。
但是,这种模型也有不足之处,它比较忽视了蛋白质分子对脂质分子流动性的控制作用,以及其他因素对脂质分子运动的影响。
3.晶格镶嵌模型由于液态镶嵌模型有上述不足之处,沃利奇(Wallach)于1975年提出了晶格镶嵌模型。
第三章第1节 细胞膜的结构和功能

我确信哪怕一个最简单的细胞,也比迄今为止设计出的任何 智能电脑更加精巧。
——引自翟中和院士等主编的《细胞生物学》
第3章 细胞的基本结构
第1节 细胞膜的结构和功能
学生自主阅读第40-46页, 思考以下几个问题
第1节
细胞膜的结构和功能
本节聚焦 1、细胞膜有哪些主要功能? 2、流动镶嵌模型的基本内容是什么? 3、通过对细胞膜结构的探索过程的分析,你对科学的过程 和方法有哪些领悟?
1972年,辛格和尼科尔森提出——流动镶嵌模型。
科学方法——提出假说
根据已有的知识和信息提出解释某一生物
1.细胞膜主要是由磷脂分子和蛋白质分子构成的。 2.磷脂双分子层是膜的基本支架,其内部是磷脂分子的
学问题的一种假说,再用进一步的观察与 实验对已建立的假说进行修正和补充。
疏水端,水溶性分子或离子不能自由通过,因此具有屏障作用。
(1)为什么两类药物的包裹位置各不相同? 由双层磷脂分子构成的脂质体,两层磷脂分子 之间的部分是疏水的,脂溶性药物能被稳定地 包裹在其中;脂质体的内部是水溶液的环境, 能在水中结晶的药物可稳定地包裹其中。
脂质在细胞膜中只有排列成连续的两层,铺展成单分子层的面积 才会恰为红细胞表面积的2倍。
思考·讨论:P42
3.磷脂分子在水中能自发地形成双分子层,你如何解释 这一现象?由此,你能否就细胞膜是由磷脂双分子层 构成的原因作出分析?
由于磷脂分子有亲水的“头部”和疏水的“尾部”,在水溶液中,朝 向水的是“头部”,“尾部”受水的排斥。当磷脂分子的内外两侧均 是水环境时,磷脂分子的“尾部”相对排列在内侧,“头部”则分别 朝向两则水的环境,形成磷脂双分子层。
成分:纤维素和果胶 功能:支持和保护 特点:全通透性 细胞壁没有生命, 不属于细胞的边界。
高三生物一轮复习课件细胞膜的结构和功能
题后总结
知识点02
3.细胞膜流动性的意义和影响因素
细胞的融合实验
细胞膜具有一定的流动性,这是生物膜 正常功能 的必须条件。例如,细胞的物质运输、细胞识别、 细胞免疫、细胞分化与信息转导等都与膜 流动性 有密切关系。温度影响分子运动,因而影响膜的流动 性。一定范围内,温度升高,膜的流动性 加大,有利于生理功能的进行;但温度过高,膜流动性过大, 会破坏 膜结构,不利于生命活动的进行,甚至使细胞死亡;若温度过低,膜流动性下降,黏度增加,运 输功能下降,严重的使膜结构破坏,内容物 大量排出 ,引起细胞死亡。
知识点01 对细胞膜结构的探索
【典例1】 下列关于细胞膜探索历程的说法,正确的是( B )
A. 用电镜观察细胞膜,看到的是亮-暗-亮三层,其中暗的部分是蛋白质,亮的是磷脂 B. 提取人的红细胞中的脂质,单分子层面积为红细胞表面积的2倍,证明细胞膜中的脂质分子为两层 C. 人鼠细胞融合实验中,放射性同位素标记的蛋白质发生流动,可证明细胞膜具有流动性 D. 磷脂是甘油、脂肪酸和磷酸组成的,其中“磷酸”头是疏水的,“脂肪酸”尾是亲水的
(5)细胞膜的外侧是___M_____(选填“M”或“N”)侧,判断的依据是_M___侧__有___多__糖__与___蛋__白___质__结。合形成的糖蛋 白,糖蛋白位于细胞膜外侧
(6)细胞膜的这种结构模型被称为_流___动__镶___嵌__模__型___。
知识点04 细胞膜的组成及功能的实验探究
2.细胞膜静态结构模型 3.细胞膜的流动镶嵌模型
细胞膜的功能和结构

b.在37 ℃下经过40 min后两种颜色的荧光_均__匀__分__布_。 c.结论:细胞膜具有流__动__性__。
(3)1972年,辛格和尼科尔森 ①方法:新的观察和实验证据。 ②结果:提出为大多数人所接受的流__动__镶__嵌__模__型__。
特别
提醒
【科学方法】
【提出假说】细胞膜结构的探索过程,反映了提出假说这一科学 方法的作用。科学家首先根据已有的知识信息,提出解释某一生 物学问题的一种假说,再进一步观察与实验对已建立的假说进行 修正和补充。一种假说最终被接受或被否定,取决于它是否能与 以后不断得到的观察和实验结果相吻合!
b
亲水的头 疏水的尾
【思考】磷脂分子在空气——水界面上会怎样排布? 空气
水 磷脂分子的结构(P42)
教材1深925挖年:,将两位人荷的兰肝科细学胞家中戈特的和磷格脂伦全德部尔提用丙取酮出从来人,的铺红成细胞单中分提子取层脂,质其,在面积 是肝空细气胞—表水面界面积上的铺2倍成单吗分?子为层什,么测?得单层分子的面积恰为红细胞表面积的2倍。
大大超过2倍,肝细胞具有多种具膜的细胞器和细胞核
【问题1】该实验得出了怎样的推论? 1层细胞膜含2层磷脂分子 【问题2】细胞膜上的磷脂分子是怎样排b 列的?
水 水
水 水
讨论:如果将磷脂分子置于水—苯的混合溶剂中,磷脂分子将会如何分布?
b
如果将磷脂分子置于水—苯的混合溶剂中,磷脂的“头部”将与水接触, “尾部”与苯接触,磷脂分子分布成单层。
【科学方法】
【提出假说】细胞膜结构的探索过程,反映了提出假说这一科学 方法的作用。科学家首先根据已有的知识信息,提出解释某一生 物学问题的一种假说,再进一步观b 察与实验对已建立的假说进行 修正和补充。一种假说最终被接受或被否定,取决于它是否能与 以后不断得到的观察和实验结果相吻合!
细胞膜结构的液态镶嵌模型

细胞膜:液态镶嵌的奇妙结构细胞膜是细胞中最重要的结构之一,具有控制细胞物质进出的重要功能。
近年来,液态镶嵌模型成为了描述细胞膜结构的一种重要理论。
本文将从不同角度介绍液态镶嵌模型的成因、特点以及应用。
液态镶嵌模型是如何产生的呢?我们知道细胞膜是由两层磷脂分子排列而成,这个结构看起来似乎很简单,但其实内部组成是极其复杂的。
每一层磷脂分子都能自由旋转,而且细胞内外环境的化学成分不同,在不同条件下,磷脂分子之间的相互作用也会发生改变。
这种动态的变化导致了细胞膜内外层形成了许多微小的区域,这些小区域大小不同、形状不一、含有不同的蛋白质和其他生物大分子,这些区域统称为“液态镶嵌体”。
液态镶嵌体优越的物理化学性质,使其在许多细胞过程中发挥着至关重要的功能。
例如,某些类型的液态镶嵌体上有一些接受信号的受体,这些受体可以接受到化学分子或电子信号,并将这些信号传递到细胞内部。
此外,液态镶嵌模型还有利于细胞的自我修复,并提供了更加灵活的细胞移动方式。
更加深入地理解液态镶嵌模型,需要了解其特点。
首先,液态镶嵌体内部不是均匀的,有一定的局部有序性。
其次,液态镶嵌体整体上稳定性较高,但组成其的磷脂分子是可以自由翻转的。
最后,液态镶嵌体可以被其他细胞结构和细胞外物质所影响,这种相互作用可能会导致液态镶嵌体的构成发生改变,并对细胞功能产生影响。
液态镶嵌模型的应用领域非常广泛。
例如,在药物研究中,液态镶嵌模型可以被用来描述药物分子和细胞膜之间的相互作用,从而预测药物分子在细胞中的吸收、分布和代谢。
此外,液态镶嵌模型还可以被用来设计新型的生物传感器和药物递送系统,从而提高其稳定性和可控性。
综上所述,液态镶嵌模型是研究细胞膜结构的一种重要理论。
通过了解液态镶嵌模型的成因、特点以及应用,可以帮助我们更好地理解细胞膜的功能和生命过程,同时也为药物研究和生物技术的发展提供了新的思路。
细胞膜的分子结构模型

细胞膜的结构模型
细胞膜的构造
1.按组成元素分:构成细胞膜的成分有磷脂和糖蛋白。
2.按组成结构分:磷脂双分子层是构成细胞膜的的基本支架。
细胞膜的主要成分是蛋白质和脂质,含有少量糖类。
其中部分脂质和糖类结合形成糖脂,部分蛋白质和糖类结
合形成糖蛋白。
3.化学组成:细胞膜主要由脂质、蛋白质和糖类等物质组成;其中以蛋白质和脂质为主。
在电镜下可分为三层,即在膜的靠内外两侧各有一条厚约2.5nm的电子致密带,中间
夹有一条厚2.5nm的透明带,总厚度约7.0~7.5nm左右这种结构不仅见于各种细胞膜,细胞内的各种细胞器膜如:线粒体、内质网等也具有相似的结构。
细胞膜功能
(1)分隔、形成细胞和细胞器,为细胞的生命活动提供相对稳定的内部环境,膜的面
积大大增加,提高了发生在膜上的生物功能;
(2)屏障作用,膜两侧的水溶性物质不能自由通过;
(3)选择性物质运输,伴随着能量的传递;
(4)生物功能:激素作用、酶促反应、细胞识别、电子传递等。
细胞膜的结构模型

03
1959年,J.David和A.Danielson通过电子显微镜观察到细胞膜的脂质双层结构
脂质双层模型的基本特点
细胞膜主要由磷脂和胆固醇组成,形成脂质双层结构
磷脂分子形成双层结构,胆固醇调节膜流动性
蛋白质镶嵌在脂质双层中,实现细胞膜的功能
脂质双层模型的局限性
未能解释细胞膜的选择性通透性和信号传导等功能
02
信号传导
• 接收细胞外信号,传导至细胞内,引发生物学效应
• 通过受体蛋白、信号传导蛋白等实现信号传导
03
物质转运
• 负责细胞内外物质的转运,包括被动转运和主动转运
• 通过通道蛋白、转运蛋白等实现物质转运
细胞膜与细胞内外环境的相互作用
与细胞外的相互作用
与细胞内的相互作用
• 与周围细胞、组织、器官等相互作用,实现生物学功能
节机制
04
细胞膜的流动镶嵌模型
流动镶嵌模型的发展历程
1972年,
S.J.Singer和
G.L.Nicolson
提出流动镶嵌
模型,认为细
胞膜由脂质双
层和镶嵌其中
的蛋白质构成,
具有流动性
1975年,科学
家通过荧光染
料标记实验证
实了细胞膜的
流动性
01
02
流动镶嵌模型的基本特点
细胞膜由脂质双层和镶嵌其中的蛋白
功能
05
细胞膜的其他结构模型
板块镶嵌模型
01
板块镶嵌模型认为细胞膜蛋白质以独立板块形式存在,与脂质双层相互作用
02
板块镶嵌模型解释了细胞膜蛋白质的有序排列和稳定性
03
板块镶嵌模型与流动镶嵌模型相互补充,共同解释细胞膜的结构和功能
细胞膜的液态镶嵌模型的特点

细胞膜的液态镶嵌模型的特点细胞膜是细胞的外部边界,其主要功能是控制物质的进出和细胞内外环境的维持。
细胞膜的液态镶嵌模型是一种描述细胞膜结构的模型,它具有以下特点。
1. 液态性:液态镶嵌模型认为细胞膜是由磷脂双层组成的,其中磷脂分子具有液态特性。
这意味着细胞膜具有流动性,磷脂分子可以在膜内自由移动,而不是固定不动的。
2. 镶嵌性:液态镶嵌模型认为细胞膜中的蛋白质是嵌入在磷脂双层中的,形成了一种镶嵌结构。
这些蛋白质可以是跨膜蛋白,也可以是与膜表面相接触的膜蛋白。
蛋白质的镶嵌使得细胞膜具有多样的功能,如传递信号、运输物质等。
3. 不对称性:液态镶嵌模型认为细胞膜的内外两侧具有不同的组成和功能。
磷脂分子和蛋白质在膜内外的分布是不均匀的,形成了不对称的结构。
这种不对称性使得细胞膜能够实现选择性通透和特定的功能分区。
4. 动态性:液态镶嵌模型强调细胞膜的动态性,即磷脂分子和蛋白质可以通过扩散和流动来改变它们的位置。
这种动态性使得细胞膜能够适应不同的环境和功能需求,例如在细胞分裂和细胞运动过程中。
5. 可塑性:液态镶嵌模型认为细胞膜具有可塑性,即它可以通过改变磷脂分子和蛋白质的组成来调整自身的性质和功能。
这种可塑性使得细胞膜能够适应不同的生理和病理条件,维持细胞内外环境的平衡。
细胞膜的液态镶嵌模型在描述细胞膜结构方面具有一定的局限性,仍然存在一些争议和未解之处。
例如,液态镶嵌模型无法解释一些具有特殊结构的细胞膜,如红细胞膜中的蛋白质组织方式。
此外,液态镶嵌模型也没有考虑到细胞膜中其他重要成分的作用,如胆固醇等。
因此,科学家们还在不断探索和研究中,以完善对细胞膜结构和功能的理解。
细胞膜的液态镶嵌模型具有液态性、镶嵌性、不对称性、动态性和可塑性等特点。
这个模型为我们理解细胞膜的结构和功能提供了一个重要的框架,但仍然需要进一步研究来验证和完善。
只有深入理解细胞膜的特点和机制,我们才能更好地理解细胞的生命活动和疾病发生的机理,为相关领域的研究和应用提供科学依据。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
细胞膜的结构模型
19世纪中叶K.W.Mageli发现细胞表面有阻碍染料进入的现象,提示膜结构的存在;1899年E.Oveiton发现脂溶性大的物质易入胞,推想应为脂类屏障。
1925年荷兰人E.Gorter和F.Grendel用丙酮抽提红细胞膜结构,计算出红细胞膜平铺面积约为其表面积的两倍,提出脂质双分子层模型.成立前提:a.红细胞的全部脂质都在膜上;b.丙酮法抽提完全;c.RBC平均表面积估算正确。
(70%~80%偏低);40年后Bar重复这一试验发现红细胞膜平铺面积应不是70%~80%,而是1.5倍还有蛋白质表面,同时干膜面积是99μm2,湿膜面积则为145μm2。
两项误差相抵,结果基本正确。
根据细胞的生理生化特征,曾先后推测质膜是一种脂肪栅、脂类双分子层和由蛋白质-磷脂-蛋白质构成的三夹板结构。
同时电镜观察也证实质膜确实呈暗-明-暗三层结构。
随后冷冻蚀刻技术显示双层膜中存在蛋白质颗粒;免疫荧光技术证明质膜中蛋白质是流动的。
据此S.J.Singer等人在1972年提出生物膜的流动镶嵌模型,结构特征是:生物膜的骨架是磷脂类双分子层,蛋白质分子以不同的方式镶嵌其中,细胞膜的表面还有糖类分子,形成糖脂、糖蛋白;生物膜的内外表面上,脂类和蛋白质的分布不平衡,反映了膜两侧的功能不同;脂双层具有流动性,其脂类分子可以自由移动,蛋白质分子也可以在脂双层中横向移动。
尽管目前还没有一种能够直接观察膜的分子结构的较为方便的技术和方法,但从研究中30年代以来提出了各种假说有数十种,其中得到较多实验事实支持而目前仍为大多数人所接受的是美国的S.J.Singer和G.L.Nicholsom于1972年提出的流体镶嵌模型(fluid mosaic model)。
这一假想模型的基本内容是:膜的共同结构特点是以液态脂质双分子层为基架,其中镶嵌着具有不同分子结构、因而也具有不同生理功能的蛋白质,后者主要以α-螺旋或球型蛋白质的形式存在。
其局限性在于未表达出流动性不均一,Jain与White提出了“板块与镶嵌模型”。
1、单位膜模型
J. Danielli & H. Davson 1935 发现质膜的表面张力比油-水界面的张力低得多,推测膜中含有蛋白质,从而提出了”蛋白质-脂类-蛋白质”的三明治模型。
认为质膜由双层脂类分子及其内外表面附着的蛋白质构成的。
1959年在上述基础上提出了修正模型,认为膜上还具有贯穿脂双层的蛋白质通道,供亲水物质通
过。
J. D. Robertson 1959 用超薄切片技术获得了清晰的细胞膜照片,显示暗-明-暗三层结构,厚约7.5nm。
这就是所谓的“单位膜”模型。
它由厚约3.5nm的双层脂分子和内外表面各厚约2nm的蛋白质构成。
单位膜模型的不足之处在于把膜的动态结构描写成静止的不变的。
2、流动镶嵌模型
流动镶嵌模型突出了膜的流动性和不对称性,认为细胞膜由流动的脂双层和嵌在其中的蛋白质组成。
磷脂分子以疏水性尾部相对,极性头部朝向水相组成生物膜骨架,蛋白质或嵌在脂双层表面,或嵌在其内部,或横跨整个脂双层,表现出分布的不对称性.
质膜的结构模型
3、晶格模型
流动镶嵌模型的补充,强调流动的整体性。
4、脂筏模型
脂筏(lipid raft)是质膜上富含胆固醇和鞘磷脂的微结构域(microdomain)。
大小约70nm左右,是一种动态结构,位于质膜的外小页。
由于鞘磷脂具有较长的饱和脂肪酸链,分子间的作用力较强,所以这些区域结构致密,介于无序液体与液晶之间,称为有序液体(Liquid-ordered)。
在低温下这些区域能抵抗非离子去垢剂的抽提,所以又称为抗去垢剂膜(detergent-resistant membranes,DRMs)。
脂筏就像一个蛋白质停泊的平台,与膜的信号转导、蛋白质分选均有密切的关系。
从脂筏的角度来看,膜蛋白可以分为三类:①存在于脂筏中的蛋白质;包括糖磷脂酰肌醇锚定蛋白(GPI anchored protein),某些跨膜蛋白,Hedgehog蛋白,双乙酰化蛋白(doubly acylated protein)如:非受体酪氨酸激酶Src、G蛋白的G α亚基、血管内皮细胞的一氧化氮合酶(NOS);②存在于脂筏之外无序液相的蛋白质;③介于两者之间的蛋白质,如某些蛋白在没有接受到配体时,对脂筏的亲和力低,当结合配体,发生寡聚化时就会转移到脂筏中。
脂筏中的胆固醇就像胶水一样,它对具有饱和脂肪酸链的鞘磷脂亲和力很高,而对不饱和脂肪酸链的亲和力低,用甲基-β-环糊精(methyl-β-cyclodextrin)去除胆固醇,抗去垢剂的蛋白就变得易于提取。
膜中的鞘磷脂主要位于外小页,
而且大部分都参与形成脂筏。
据估计脂筏的面积可能占膜表面积的一半以上。
脂筏的大小是可以调节的,小的独立脂筏可能在保持信号蛋白呈关闭状态方面具有重要作用,当必要时,这些小的脂筏聚集成大一个大的平台,在那里信号分子(如受体)将和它们的配件相遇,启动信号传递途径。
如致敏原(allergen)能够将过敏患者体内肥大细胞或嗜碱性细胞表面的IgE抗体及其受体桥联起来,形成较大的脂筏,受体被脂筏中的Lyn(一种非受体酪氨酸激酶)磷酸化,启动下游的信号转导,最终引发过敏反应。
细胞表面的穴样内陷(caveolae)具有和脂筏一样的膜脂组成,不含笼形蛋白(clathrin),含有caveolin(一种小分子量的蛋白,21KD)。
大量存在于脂肪细胞、上皮细胞和平滑肌细胞。
这种结构细胞的内吞有关,另外穴样内陷中还富含某些信号分子,说明它与细胞的信号转导有关。