第四章细胞质膜
第四章 细胞质膜
影响中枢神经系统,甚至危及生命。
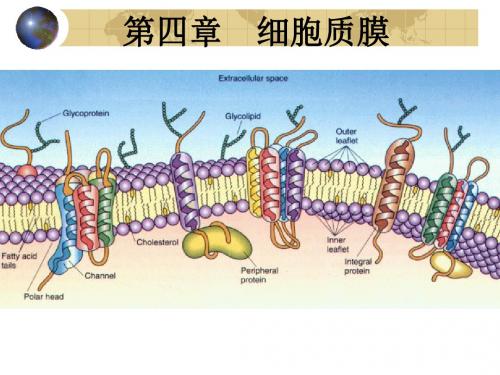
糖脂
其极性头部共价结合到鞘氨醇上的糖分子或寡糖链。 普遍存在于原核和真核细胞的质膜上,约占膜脂总 量的5%以下,但神经细胞膜上糖脂含量较高,约 5-10%。 最简单的糖脂是半乳糖脑苷脂;最复杂的糖脂是神 经节苷脂。家族性白痴病和神经节苷脂的异常降解 有关。
糖脂,是鞘氨醇的衍生物
膜的流动性低于一定的阈值时,许多酶的活动和跨
膜运输将停止.
流动性过高,又会造成膜的溶解。
(二)细胞膜的不对称性
膜内外两层组分和功能的差异,称膜的不对称性。 (二)细胞膜的不对称性 样品经冰冻断裂处理后,各断面命名为:ES,细胞外表面;
EF ,细胞外小页断面; PS ,原生质表面; PF ,原生质小
甘油磷脂
(3-磷酸甘油的衍生)
2、鞘脂
主要在高尔基体合成。 是含有鞘氨醇骨架的两性脂,具有一条 长链脂肪酸,一条烃链 ,和一个头部。 根据头部基团的不同,可分鞘磷脂、脑 苷脂以及神经节苷脂三种;其中,鞘磷 脂中含有磷酸基团,脑磷脂和神经节苷 脂中含糖残基而不含磷酸基团。
鞘 磷 脂 p56
鞘磷脂(sphingomyelin)是最普通的鞘脂,其极 性头是磷酰胆碱或磷酰乙醇胺。 在高等动物的组织中含量极为丰富,细胞内鞘磷脂 的水解有赖于磷脂酶,磷脂酶的先天性缺损便导致 鞘磷脂在组织内堆积,可造成肝、脾肿大,严重者
白比较容易纯化、分析。
红细胞经过低渗处理,质膜破裂,内容物释放,留下一 个保持原形的壳,称为红细胞影(血影)。红细胞是研究膜 骨架的理想材料。
(二)红细胞质膜蛋白及膜骨架
SDS聚丙烯酰胺凝胶电泳分析:血影的蛋白成 分包括:血影蛋白、锚蛋白、带3蛋白、带4.1 蛋白、肌动蛋白和血型糖蛋白。
第四章细胞质膜
2、糖脂
•是含糖而不含磷酸的脂类,含量 约占脂总量的5%以下,在神经细 胞膜上糖脂含量较高,约占5-10% 。 •糖脂也是两性分子,其结构与SM
glycolipid s
人红细胞膜的血型糖蛋白 为膜内在蛋白(跨膜蛋白),决定血型。
红细胞质膜中糖蛋白对ABO血型的决定作用
❖ 人的血型是A型、B型、AB型还是O型,是由红 细胞膜脂或膜蛋白中的糖基(也称为ABO血型抗 原,ABO血型决定子)决定的。
组成。 3) 常含有不饱和脂肪酸(如油酸)。
Phospholipids
极性 头 部
(亲 非极性尾部 水 (疏水) )
胆碱磷脂酰碱基
磷酸
甘油
脂脂 肪肪 酸酸
磷 脂 )
磷 脂 酰 胆
碱
(
卵
(1)、甘油磷脂
• 以甘油为骨架的磷脂类,在骨架上结合两个脂肪酸链一个 磷酸基团,胆碱、乙醇胺、丝氨酸或肌醇等分子籍磷酸基 团连接到脂分子上。主要类型有: ① 磷脂酰胆碱phosphatidylcholine,PC,旧称卵磷脂 ② 磷脂酰丝氨酸phosphatidylserine,PS ③ 磷脂酰乙醇胺phosphatidylethanolamine,PE,旧称 脑磷脂 ④ 磷脂酰肌醇phosphatidylinositol,PI ⑤ 双磷脂酰甘油Diphosphatidylglycerol, DPG,旧称心磷 脂
3. 推测细胞膜由连续的脂类
4. 物质组成。
2. E. Gorter & F. Grendel 1925 用有机溶剂提取了人的红细胞
质膜的脂类成分,将其铺展在水面,测出膜脂展开的面积二倍 于细胞表面积,因而推测细胞膜由双层脂分子组成。
Langmuir水盘中展
细胞生物学 名词解释 第四章 细胞质膜

lipid anchored protein
通过与之共价连接的脂分子插入膜的脂双分子层中,从而锚定在细胞质膜上。与脂肪酸结合的脂锚定蛋白分布在质膜内测,与糖脂结合的脂锚定蛋白分布在质膜外侧,GPI脂锚定膜蛋白分布在质膜外侧。
研究膜蛋白的分析技术
低温电镜、X射线晶体衍射技术、电镜三维重构
研究膜蛋白的难处
1、表达量低
2、分离纯化困难
3、难以形成三维晶体
细胞膜流动性பைடு நூலகம்
胆固醇含量减低,脂肪酸链短,不饱和度高,相变温度低,PC/SM比例高,流动性大(胆固醇即可增强流动性,又可降低流动性),由此可得流动性与胆固醇含量、脂肪酸链长短、饱和程度、PC/SM比值有关。
研究膜蛋白流动性(侧向运动)的实验
荧光抗体免疫标记细胞融合:用抗鼠细胞质膜蛋白的荧光抗体(绿色荧光)和抗人细胞质膜蛋白的荧光抗体(红色荧光)分别标记小鼠和人的细胞表面,然后用灭活的仙台病毒介导两种细胞融合,10min后,不同颜色的荧光开始在融合细胞的表面扩散,40min后分辨不出融合细胞表面的绿色荧光和红色荧光区域,两种荧光均匀的分布在融合细胞表面。这一实验证明膜蛋白在脂膜上的运动。
带3蛋白
band3 protein
是红细胞质膜Cl-/HCO3-阴离子运输的载体蛋白,带3蛋白的N末端伸向细胞质膜基质面折叠成不连续的水不溶性的区域,为膜支架蛋白提供结合位点,多次跨膜蛋白,内在膜蛋白
细胞质膜的功能
1、为细胞的生命活动提供相对稳定的内环境,2、选择性的物质运输,3、提供细胞识别位点,4、为多种酶提供结合位点;5、介导细胞与细胞,细胞与胞外基质之间的连接;6、质膜参与形成具有不同功能的细胞表面特化结构;7、膜蛋白的异常与某些遗传病、恶性肿瘤、自身免疫病甚至神经退行性疾病有关,很多膜蛋白可作为疾病治疗的药物靶标
第4章 细胞质膜 (西安农林科技大学)

第四章细胞质膜•细胞质膜是围绕细胞最外层的一层由脂质和蛋白质组成的生物膜4.1 细胞膜的结构模型与基本成分膜。
•质膜提供给细胞一个相对稳定的内环境①。
在物质运输①③,能量转换③,信息传递④⑤,细胞连接和黏着⑤等方面有重要作用。
用•真核细胞内部存在由膜构建的各种细胞器,它们有与质膜结构类似的生物膜②。
Singer 和Nicolson 的流动镶嵌模型是膜生物学的核心理论。
膜的流动性(流动):膜脂处于流动状态,膜脂和膜蛋白均处于动态运动中,可侧向运动。
膜蛋白分布的不对称性(镶嵌):膜蛋白分布的不对称性(镶嵌)膜蛋白以多种方式镶嵌于脂双分子层内、贯穿其中或附着在脂层表面。
三明治模型和流动镶嵌模型。
电镜超薄切片显示的细胞膜生物膜模型:A,流动镶嵌;B,生物膜结构示意100的质膜凹陷区也富含•膜泡窖:50~100nm 的质膜凹陷区。
也富含胆固醇、鞘脂和糖基磷脂酰肌醇脂锚定膜蛋白。
含有窖蛋白。
膜泡窖:transient confinement 瞬时封闭,caveolae 膜泡窖,caveolin 窖蛋白,nanodomain 超微区,GPI 糖基磷脂酰肌醇,fluid lipid 流动脂。
脂筏形成机制:蛋白亚基聚集导致与其连接的相对稳定的膜脂微区域聚集。
ligand 配体,lectin 凝集素,oligomerization 低聚化,adapter 接头蛋白,scaffold 支架蛋白。
3. 生物膜可看成是蛋白质在双分子层中的二维溶液;但存在脂筏等结构功能单位;4. 生物膜在三维空间上可弯曲、折叠、延伸等改变,处于动态变化之中。
4.1.2膜脂:膜脂为两性分子兼具亲水性和疏水性。
■膜脂主要有三种:磷脂、糖脂和胆固醇。
●磷脂:甘油磷脂和鞘氨醇磷脂。
约占膜脂的50%以上。
神经系统含鞘磷脂较丰富。
主要特征:•一个个极性头、两个非极性尾(脂肪酸链)。
•脂肪酸碳链为偶数,16,18或20个碳原子。
•常含有不饱和脂肪酸(如油酸)。
第四章:细胞质膜

的脂双层中呈不均
匀分布,如:PC和
SM主要分布在外小
叶,PE和PS分布在
内小叶。
镰刀形红细胞与荧光标记的 annexinV共孵育的荧光显微镜照片
流式细胞仪检测到的PS暴露
于表面的红细胞的百分比
2,膜蛋白的不对称性 包括:整合蛋白插入的深度不同; 穿过蛋白质两侧的基团不同;
两侧表面边周蛋白的性质种类不同。
1,2:膜整合蛋白 3,4:脂锚定蛋白 5,6:外周蛋白
1、整合膜蛋白(integral membrane protein) 1)也称跨膜蛋白(transmembrane proteins),为两 性分子,非极性区(疏水区)插在脂双层分子之间, 极性区(亲水区)则朝向膜的表面,占70%~80%。 2)主体穿过细胞膜脂双层,分为单次跨膜、多次跨 膜和多亚基跨膜蛋白三种。 3)单次跨膜蛋白肽链穿过脂双层一次,有胞外、胞 质和跨膜三个结构域,跨膜域含有20~30个疏水性 氨基酸,以α螺旋构象穿越脂双层。穿膜的方式包 括:
结合,因此只要改变溶液的离
子强度甚至提高温度就可以从
膜上分离下来。
有时很难区分整合蛋白和外周蛋白,主要是因为一
个蛋白质可以由多个亚基构成,有的亚基为跨膜蛋
白,有的则结合在膜的外部。
附着
膜蛋白
膜蛋白
周边蛋白
周边蛋白
附着在其他膜蛋白上的周边蛋白
3、脂锚定蛋白(lipid-anchored protein)
滑入细胞质或在脂肪双层翻转,也可能参与环境相互
作用,细胞识别。
二、细胞膜的特性 (一)膜的不对称性决定膜功能的方向性 膜组分分布不对称性与膜功能的不对称性,使生 命活动高度有序。 1、膜脂的不对称性 1)膜脂内外层化学组成不对称 2)不同膜性细胞器中脂类成分组成不同
040细胞质膜:
①
②
③ ③④脂锚定膜蛋白 ③④脂锚定膜蛋白
⑤ ⑤⑥外周膜蛋白 ⑤⑥外周膜蛋白
①②膜整合蛋白 ①②膜整合蛋白
膜蛋白类型
外在膜蛋白 或外周膜蛋白 extrinsic or peripheral membrane protein
内在膜蛋白 或整合膜蛋白 intrinsic or integral membrane protein
血影蛋白 锚定蛋白 带3蛋白 蛋白 带4.1蛋白 蛋白 肌动蛋白 血型糖蛋白
1 膜脂分子的运动
第四章 细胞质膜
第二节 细胞膜基本特征
影响膜脂流动性的因素
胆固醇 胆固醇的含量增加会降低膜的流动性。 胆固醇的含量增加会降低膜的流动性。
脂肪酸链饱和度 脂肪酸链所含双键越多越不饱和,流动 脂肪酸链所含双键越多越不饱和, 性增加。 性增加。 脂肪酸链链长 卵磷脂/ 卵磷脂/鞘磷脂 长链脂肪酸相变温度高,膜流动性降低。 长链脂肪酸相变温度高,膜流动性降低。 比例高则膜流动性增加, 比例高则膜流动性增加,是因为鞘磷脂 粘度高于卵磷脂。 粘度高于卵磷脂。 温度、酸碱度、离子强度等。 温度、酸碱度、离子强度等。
第四章 细胞质膜
第一节 细胞膜结构膜
Phospholipids
第四章 细胞质膜
第一节 细胞膜结构膜
磷脂类型
-CH2-CH-N+3 I COOH 磷酯酰丝氨酸
-CH2-CH2-N+3 O C O CH2 O C O CH O H 2C O P O OCH2 CH2 CH3
磷脂酰乙醇胺: 磷脂酰乙醇胺: 脑磷脂
EF
PF
第四章 细胞质膜
第二节 细胞膜基本特征
1 膜脂的不对称性 同一种脂分子在膜的脂双层中呈不均匀分布, 同一种脂分子在膜的脂双层中呈不均匀分布,质膜的内 外两侧分布的磷脂的含量比例也不同。 外两侧分布的磷脂的含量比例也不同。
第4章 细胞质膜

脂锚定膜蛋白的3种类型
A:脂肪酸结合到膜蛋白 N 端的甘氨酸残基上 B:烃链结合到膜蛋白 C 端的半胱氨酸残基上 C:通过糖脂锚定在细胞质膜上
去垢剂
第 一
去垢剂是一端亲水一端疏水的双亲媒性分子, 是 分离与研究膜蛋白的常用试剂
节
细 胞 膜 的 结 构 模 型
膜糖:糖脂、糖蛋白
细胞质膜上的膜糖都位于质膜外表面, 内膜系统中的膜糖则面向细胞器腔面!
第 一、生物膜的基本特征
二
节 膜的流动性
生 物
膜的不对称性:
膜
基 本
二、细胞膜的基本功能
特
征
与
功
能
一、膜的流动性
膜的流动性是是所有的生物膜的基本特征,是细胞 生长增殖等生命活动的必要条件
结
构
模
型
甘油磷脂
甘油磷脂的类型
磷脂酰乙醇胺(脑磷脂)——PE 磷酯酰丝氨酸——PS 磷脂酰胆碱(卵磷脂)——PC 磷脂酰肌醇——PI
甘油磷脂分子的主要特征
一个极性头部,两个非极性尾。心磷脂除外,有 4个非极性尾。
极性头部的大小影响脂双层的曲度 脂肪酸链碳原子数为偶数(多为16或18个碳原子) 除饱和脂肪酸外,常含有不饱和脂肪酸。
生
物 膜
• 膜骨架、鞭毛、纤毛、微绒毛和伪足等
基
本
特
征
与
功
能
膜骨架
第 二
是由质膜下与膜蛋白相连的由纤维蛋白组成的
节
网架结构;参与维持质膜的形状并协助质膜完
生 物
成多种生理功能。位于细胞质膜下约0.2μm厚
膜
基 的溶胶层。
细胞生物学第四章细胞质膜

最丰富和最具特性的脂。它有一个极性的头部 和一个疏水的尾部。
(internal membrane), 习惯上把细胞所有膜 结构统称为 生物膜 (biomembrane)。
细胞生物学第四章细胞质膜
虽然细胞很早就在光镜下被发现,但其是否有明确的边界结构,尚 未可知,直到电镜发明发现质膜的超微结构,但人们并不惊奇,因 为在此之前已侦知。
1、关于膜的化学组成的早期研究: 18世纪90年代,Overton 用植物的根毛作实验,
第四章、细胞质膜
知识要点: 1 、掌握几种膜分子结构模型学说,并评价之。 2 、了解膜结构的组成成分和组成方式。 3 、理解质膜流动性和不对称性两大特点。 4、了解红细胞膜骨架的组成和功能。
细胞生物学第四章细胞质膜
第一节、细胞膜结构模型与成分 一、细胞膜结构
膜(membrane)是细胞 的重要结构, 包括细 胞质膜 (plasma membrane)、内膜
高尔基体膜)、细胞类型(肌细胞、肝细胞)、生物类型(动物、植物 和原核生物)的不同而不同。
一般而言ห้องสมุดไป่ตู้膜脂占50%,蛋白质占40%,碳水化合物约1-10%
细胞生物学第四章细胞质膜
1、膜脂
膜脂都具有双亲性,这种性质使生物膜具有屏 障作用,大多数水溶性物质不能自由通过,只 允许亲脂性物质通过。
膜脂是生物膜的基本组成成分, 主要有三大类 型:磷脂、糖脂、胆固醇。
质膜的片层结构模型
细胞生物学第四章细胞质膜
5、单位膜模型(unit membrane model)
1959年,J.D.Robertson在电子显微镜下发现细胞膜是类似 铁轨结构,两条暗线被一条明亮的带隔开,显示暗—明— 暗三层,总厚度为7.5nm,中间层为3.5nm,内外两层各为 2nm。并推测:暗层是蛋白质,透明层是脂,并建议将这种 结构称为单位膜。
植物生理学第四章 细胞质膜

2、鞘脂
2.1、鞘磷脂(sphingomyelin,SM)在脑和神经细胞膜中特别 丰富,亦称神经醇磷脂,它是以鞘胺醇(sphingoine)为骨架, 与一条脂肪酸链组成疏水尾部,亲水头部也含胆碱与磷酸结合。 原核细胞和植物中没有鞘磷脂。
2.2、糖脂
存在于原核和真核细胞的质膜上,其含量约占膜脂总量的 5%以下,在神经细胞膜上糖脂含量较高,约占5-10%。糖 脂也是两性分子,结构与磷脂相似,由一个或多个糖残基代 替磷脂酰胆碱而与鞘氨醇的羟基结合。
膜脂的功能:
1)支撑,膜脂是细胞的骨架; 2)维持构象并为膜蛋白行使功能提供环境; 3)是部分酶行驶功能所必需的。
三、 膜蛋白
膜蛋白是膜功能的主要体现者。酵母核基因组编码的蛋白 质中约1/3为膜蛋白。
根据与膜脂的结合方式以及在膜中的位置的不同,膜蛋白 分 为 内 在 ( integral protein)、 外 周 蛋 白 ( peripheral protein)和脂锚定蛋白(lipid-anchored protein)。
个磷酸基团,胆碱、乙醇胺、丝氨酸或肌醇等分子籍磷酸基团
连接到脂分子上。
主要类型有:
磷脂酰胆碱(phosphatidyl choline,PC,旧称卵磷脂)
磷脂酰丝氨酸(phosphatidyl serine,PS)
磷脂酰乙醇胺(phosphatidyl ethanolamine ,PE,旧称脑磷脂) 磷脂酰肌醇(phosphatidyl inositol,PI)等。
第四章 细胞质膜
细胞质膜的结构模型与基本成分 生物膜基本特征与功能(重点/难点) 细胞质膜相关的膜骨架
又称质膜(plasma membrane),是指围绕 在细胞最外层,由脂质和蛋白质组成的生物膜。
细胞生物学-4第四章质膜

七、细胞质膜的功能
为细胞的生命活动提供相对稳定的内环境; 选择性的物质运输,包括代谢底物的输入与代谢
产物的排除,其中伴随着能量的传递; 提供细胞识别位点,并完成细胞内外信息跨膜传递; 为多种酶提供结合位点,使酶促反应高效而有序地进行; 介导细胞与细胞、细胞与基质之间的连接; 质膜参与形成具有不同功能的细胞表面特化结构。
脂质体的应用
研究膜脂与膜蛋白及其生物学性质; 脂质体中裹入DNA可用于基因转移; 在临床治疗中,脂质体作为药物或酶等载体
三、膜蛋白
基本类型 内在膜蛋白与膜脂结合的方式 外在膜蛋白与膜脂结合的方式 去垢剂(detergent)
基本类型
外在(外周)膜蛋白(extrinsic/peripheral membrane proteins )
膜的不对称性
细胞质膜各部分的名称 膜脂与糖脂的不对称性
糖脂仅存在于质膜的ES面,是完成其生理功能的结构基础 膜蛋白与糖蛋白的不对称性
膜蛋白的不对称性是指每种膜蛋白分子在细胞膜上都 具有明确的方向性; 糖蛋白糖残基均分布在质膜的ES面(GO+3HBH4 labeling); 膜蛋白的不对称性是生物膜完成复杂的在时间与空间上 有序的各种生理功能的保证。
糖脂:糖脂普遍存在于原核和真核细胞的质膜上(5%以下),神经细 胞糖脂含量较高;
胆固醇:胆固醇存在于真核细胞膜上(30%以下),细菌质膜不含有 胆固醇,但某些细菌的膜脂中含有甘油脂等中性脂类。
运动方式
沿膜平面的侧向运动(基本运动方式),其扩散 系数为10-8cm2/s;相当于2μm/s
脂分子围绕轴心的自旋运动; 脂分子尾部的摆动; 双层脂分子之间的翻转运动,发生频率还不到
第四章 细胞质膜 (Cell membrance)
第4章细胞质膜

脂筏最初可能在内质网或 高尔基体上形成,最终转运 到细胞膜上。
有些脂筏可在不同程度上 与膜下细胞骨架蛋白交联。
推测一个100nm大小的脂 筏可能载有600个蛋白分 子。
脂筏就像一个蛋白质停泊 的平台,与膜的信号转导、 物质跨膜运输、蛋白质分 选均有密切的关系。
膜的研究历史:
E. Gorter & F. Grendel 1925 用有机溶剂提取 了人类红细胞质膜的脂类成分,将其铺展在水 面,测出膜脂展开的面积二倍于细胞表面积, 因而推测细胞膜由双层脂分子组成。
三明治模型
J. Danielli & H. Davson 1935 发现质膜的表面 张力比油-水界面的张力低得多,推测膜中含 有蛋白质,从而提出了“蛋白质-脂类-蛋白质” 的三明治模型。认为质膜由双层脂类分子及其 内外表面附着的蛋白质构成的。
3.生物膜可看成是蛋白质在双层脂分子中的二维溶液。 然而膜蛋白与膜脂之间,膜蛋白与膜蛋白之间及其与 膜两侧其它生物大分子的复杂的相互作用,限制了膜 蛋白和膜脂的流动性。
细胞质膜的化学构成
质膜主要由膜脂和膜蛋白组成,另外还有少量糖,主 要以糖脂和糖蛋白的形式存在。
膜脂是膜的基本骨架,膜蛋白是膜功能的主要体现者。 一般,脂类约占50%,蛋白质约40%,糖类约2%-
1959年在上述基础上提出了修正模型,认为膜 上还具有贯穿脂双层的蛋白质通道,供亲水物 质通过。
单位膜模型
J. D. Robertson 1959 用超薄切片技术获得了清晰的 细胞膜照片,显示暗-明-暗三层结构,厚约7.5nm。 这就是所谓的“蛋白质-脂类-蛋白质”“单位膜”模 型。它由厚约3.5nm的双层脂分子和内外表面各厚约 2nm的蛋白质构成。
细胞生物学 第四章细胞质膜

蛋白与膜的结合方式 ①、②整合蛋白;③、④脂锚定蛋白;⑤、⑥外周蛋白
(一)内在蛋白(integral proteins)
内在蛋白又称为整合蛋白,以不同程度嵌入脂双层的内部 ,有的为全跨膜蛋白(tansmembrane proteins)。膜蛋白为
两性分子。它与膜结合非常紧密,只有用去垢剂(detergent)
5.血型糖蛋白(glycophorin ) 血型糖蛋白又称涎糖蛋白(sialo glycoprotein),因 它富含唾液酸。血型糖蛋白是第一个被测定氨 基酸序列的蛋白质,有几种类型,包括A、B、C、 D。血型糖蛋白B、C、D在红细胞膜中浓度较 低。血型糖蛋白A是一种单次跨膜糖蛋白, 由 131个氨基酸组成, 其亲水的氨基端露在膜的外 侧, 结合16个低聚糖侧链。血型糖蛋白的基本 功能可能是在它的唾液酸中含有大量负电荷,防 止了红细胞在循环过程中经过狭小血管时相互 聚集沉积在血管中。
才能从膜上洗涤下来,常用SDS和Triton-X100。
内在蛋白的跨膜结构域形成亲水通道有两种形式,一是由多
个α螺旋组成亲水通道;二是由β折叠组成亲水通道。
内在蛋白与脂膜的结合方式:
膜蛋白的跨膜结构域与脂双层分子的疏水核心的相互作用。
跨膜结构域两端带正电荷的aa残基与磷脂分子带负电的
极 性头形成离子键,或带负电的氨基酸残基通过Ca2+、Mg2+等 阳离子与带负电的磷脂极性头相互作用。 膜蛋白在细胞质基质一侧的半胱氨酸残基上共价结合脂肪 酸分子,插入脂双层之间, 还有少数蛋白与糖脂共价结合。
细胞融合技术观察蛋白质运动
光脱色恢复技术(FRAP)
4.膜流动性的意义
质膜的流动性是保证其正常功能的必要条件。例如 跨膜物质运输、细胞信息传递、细胞识别、细胞免疫 、细胞分化以及激素的作用等等都与膜的流动性密切 相关。当膜的流动性低于一定的阈值时,许多酶的活 动和跨膜运输将停止,反之如果流动性过高,又会造
第4章 细胞质膜
膜的不对称性的意义
二、膜脂——生物膜的基本组成成分
成分:膜脂主要包括磷脂、糖脂和胆固醇三种 类型。
膜脂的运动方式
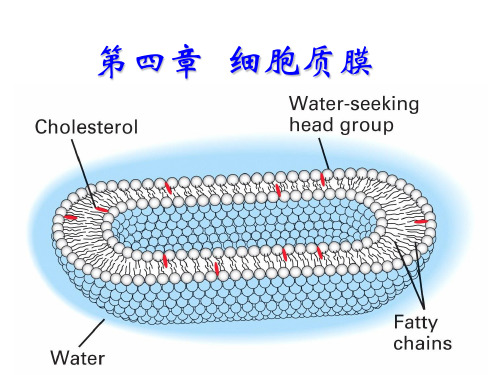
脂质体(liposome)
成 分
磷脂:膜脂的基本成分(50%以上)
分为二类: 甘油磷脂和鞘磷脂
主要特征:①具有一个极性头和两个非极性的尾(脂肪酸链) (心磷脂除外);②脂肪酸碳链碳原子为偶数,多数碳链由
流动镶嵌模型
该模型强调了膜的流动性,蛋白质和膜脂均可侧向运动,膜蛋白分布的 不对称性。有的镶嵌在脂双分子层表面,有的则全部或部分嵌入其内, 有的横跨整个脂类区
液态镶嵌模型比较合理地解释了膜中所发生的生
理现象,特别是它以动态的观点分析膜中各种化学组 分的相互关系,受到了人们的广泛关注。 该模型不足之处是忽视了蛋白质分子流动性的控 制作用,忽视了膜的各部分流动性的不均匀性。 通过进一步的研究,有学者提出强调生物膜的膜
结构式
模型
空间结构
运 动 方 式
沿膜平面的侧向运动(基本运动
方式),其扩散系数为10-8cm2/s;
脂分子围绕轴心的自旋运动; 脂分子尾部的摆动; 双层脂分子之间的翻转运动,发生频率还不到脂分子侧向交 换频率的10-10。但在内质网膜上,新合成的磷脂分子翻转运
动发生频率很高。
某些文献介绍有6种运动方式
不足之处:
(1)单位膜为一种静态单一的结构,无法说明膜的动态结构
的变化, (2)各种膜的功能不一样,而单位膜模型显示不出来,
(3)各种不同细胞和同一细胞中的不同部分膜的厚度实际上
并不都是7.5nm。 60年代以后,应用一系列新技术,证明膜中的蛋白质和脂
类分子主要以疏水键相结合。电镜冰冻蚀刻技术证明:脂类
第四章 细胞质膜

•在缺少胆固醇培养基中,不能合第成四胆章固细醇胞质的膜突变细胞株很快发生自溶。
(二)、膜脂的运动方式
• 沿膜平面的侧向运动 • 围绕轴心的自旋运动 • 尾部的摆动
• 伸缩震荡运动 • 在脂双层上的翻转运动(一般极
少发生) • 旋转异构化
第四章 细胞质膜
影响膜脂流动性的因素
• 胆固醇 • 脂肪酸链的饱和度 • 脂肪酸链的链长 • 卵磷脂/鞘磷脂:该比例高则膜流动性增加,
HC CH 2
H 2C
O
O
OH H
H
OH
H OH
H
OH H
O
OH OH
O P OO
HC
CH 2
O
C OC O
C OC O
C OCO
C OC O
R1
R2
PE
R1
R2
PS
R1
R2
第四章 细胞质膜 PC
R1
R2
PI
Diphosphatidylglycerol
OH
O CH2 C CH2 O CH2 O C R1 O H O
第四章 细胞质膜
Phospholipids in Plasma Membrane
+ NH 3
CH 2 CH 2 O O P O-
第四章细胞质膜

2. 围绕轴心的自旋运动:膜脂分子围绕与膜平面垂直的轴进行快速旋转。 3.脂分子尾部的摆动:膜脂分子围绕与膜平面垂直的轴进行左右摆动。 4.双层脂分子之间的翻转运动:膜脂分子从脂双层的一层翻转到另一
层 。是在翻转酶(flippase)催化下完成发生频率不到脂分子侧向交换频率的 10-10。但在内质网膜上,新合成的磷脂分子翻转运动发生频率很高。
2.脂质体的应用
研究生物膜的特性研究膜脂与膜蛋白及
其生物学性质;
转基因脂质体中裹入DNA可用于基因转移;
制备药物在临床治疗中,脂质体作为药物或
酶等载体。
三、膜蛋白
占核基因组编码蛋白质的30%。 1、基本类型 (1)外在(外周)膜蛋白(extrinsic/peripheral membrane protein ):水溶性蛋白,靠离子键或其它弱键与膜内表面的 蛋白质分子或脂分子极性头部非共价结合,改变溶液的离子 强度、提高温度就可从膜上分离下来。 (2)内在(整合)膜蛋白(intrinsic/ integral membrane protein):整合蛋白为跨膜蛋白,水不溶性蛋白。是两性分 子,形成跨膜螺旋,跨膜结构域为1至多个疏水的α螺旋, 与膜结合紧密,需用去垢剂使膜崩解后才能从膜上洗涤下来。
小窝蛋白
糖基磷脂酰肌醇锚定
细胞膜穴样内陷
过渡限制区
微结构域
二、膜脂——生物膜的基本组成成分
膜脂的成分:膜脂主要包括磷脂、糖脂 和胆固醇三种。
膜脂的运动方式 脂质体(liposome)
(一)膜脂成分
1.磷脂: 膜脂的基本成分(50%以上),包括甘油磷脂和鞘磷脂二 类。 主要特征:①具有一个极性头部和两个非极性的尾部(心磷 脂除外);②脂肪酸碳链碳原子为偶数,多数碳链由16、18 或20个组成;③含饱和脂肪酸(如软脂酸)及不饱和脂肪酸 (如油酸);
第四章 细胞质膜

第四章细胞质膜第一节细胞质膜的结构模型与基本成分一细胞质膜的结构模型发现过程:1925年,E.Gorter和F.Grendel通过对红细胞质膜的研究揭示了质膜是由双层脂分子构成的。
之后,Davson和Danielli推测出质膜中含有蛋白质成分,并提出三明治式模型—蛋白质-脂质-蛋白质1959年,Robertson提出单位膜模型,并大胆推测所有的生物膜都是由蛋白质-脂质-蛋白质的单位膜构成。
这一推测得到了X射线分析结果的支持Singer和Nicolson在1972年提出生物膜的流动镶嵌模型,该模型得到了各种实验结果的支持。
1975年Uuwin和Henderson首次报道了质膜蛋白菌紫红质的三维结构,他是一个跨膜蛋白镶嵌在膜上。
流动镶嵌模型强调:1,膜的流动性,即膜蛋白和膜脂都可以流动;2.膜蛋白分布的不对称性,有的在膜表面,有的镶嵌,有的跨膜。
Simons在1988年提出了脂筏模型。
脂筏模型—在生物膜上胆固醇,鞘磷脂等富集区域形成相对有序的脂相,如同漂浮在脂双层上的脂筏一样载着执行某些特定生物学功能的各种膜蛋白。
生物膜结构归纳总结:1.具有极性头部和非极性尾部的磷脂分子在水相中具有自发形成封闭的膜系统的性质,磷脂分子以疏水性的尾部相对,极性头部朝向水相形成脂双分子层,每层磷脂分子称为一层小叶。
2.蛋白质分子以不同的方式镶嵌在脂双层分子中或结合在其表面,蛋白质的类型,蛋白质分布的不对称性及其与脂分子的协同作用赋予生物膜各自的特性与功能。
3.生物膜可看成是蛋白质在双层脂分子之中的二维溶液。
膜蛋白与膜脂之间,膜蛋白与膜蛋白之间及其与膜两侧其他生物大分子的复杂的相互作用,在不同成度上限制了膜蛋白和膜脂的流动性。
4.在细胞生长和分裂整个生命活动中,生物膜在三维空间上可出现弯曲,折叠,延申等改变,处于不断的动态变化中。
二、膜脂(一)成分1.甘油磷脂构成了膜脂的基本成分,占整个膜脂的一半以上。
是3-磷酸甘油的衍生物.主要在内质网合成主要特征:1,具有一个与磷酸基团相结合的极性头部和两个非极性的尾部,但是存在于线粒体和某些细菌质膜上的心磷脂除外,它有4个尾部。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
4.1 膜的流动性和不对称性 • 膜脂的流动性和不对称分布 • 胆固醇嵌于磷脂分子之间既限 制了磷脂分子的运动,又降低 了磷脂分子的相变温度。
磷脂中的不饱和脂肪酸增加了增加了膜的流动性
膜脂分子的运动
顺式双键脂肪酸链降低了膜脂分子排列的有序性,增 加膜脂流动性,使膜脂双分子层更难被低温所凝固。
哺乳动物质膜中几种主要磷脂和 糖脂电荷特征及在红细胞膜中的 不对称分布。卵磷脂(PC,极性头 部大)、鞘磷脂(SM)、主要分布 于双分子层的细胞外一侧。脑磷 脂(PE,极性头部小)、磷脂酰丝氨 酸(PS)、磷脂酰肌醇(PI)主要分布 于质膜内侧。如磷脂酰丝氨酸在 细胞衰老时或凝血时才由内侧翻 向外侧,被吞噬细胞识别后被吞 噬去除。胆固醇(CI)。糖脂均位 于细胞膜外小叶。磷脂不对称性 可能与合成部位有关。膜脂不对 称分布是膜行使生理功能所必需 的。
去垢剂
两种去垢剂结构:两性 分子,拥有疏水端(绿 色)和亲水端(蓝色)
用非离子型去垢剂温和解离蛋白,非 离子型去垢剂可能是由于分子大电荷 小不易进入蛋白内部
去垢剂在水中形成微团粒, 类似磷脂
温和型去垢剂在膜蛋白解离、纯 化及膜-蛋白体系重建中的应用: Na+-K+ ATPase的分离和功能
4
生物膜的基本特征
+
磷脂磷脂模式图(卵磷脂)
+
H3C C CH2 COO-
NH3 CH2 H
NH3
+
CH3 CH3 N CH2 CH2
OH H H OH H H OH O P OO H2C O C HC O O C O CH2 OH OH OH H
OH O O CH2 C H CH2 O O O CH2 O C R3
H2C O C O
CH2 O P OO O
O
O P OO H2C O C HC O O C O CH2 H2C O C HC O
O O P OO CH2
CH2 O C R1 O
R2 C O CH CH2
O P OH O P OH R4 C O CH O O CH2
HC O
CH2
O C O
O C O
心磷脂(存在于线粒体内膜和一些细菌质膜中)
磷脂在水中的存在形式
磷脂在水中形成 的脂质体显微结 构
•
• •
糖脂:普遍存在于真核、原核生物质膜中,约占膜脂总量5%以下。神 经细胞膜含量高,约占5-10%。 不含磷酸,由一个或多个糖残基与鞘氨醇的羟基结合。 糖脂结构变化复杂,神经节苷脂是神经元膜中的特征性成分。
两种糖脂结构: 半乳糖脑苷脂 (A)由于亲水头部不带电荷被称 为中性糖脂。神经节苷脂(B)头 部亲水区含有带负电荷的唾液 酸(C)。细菌及植物中几乎所有 磷脂和糖脂均来自甘油酯,在 动物细胞中糖脂大多来自鞘氨 醇。 NANA,唾液酸; Gal, 半乳糖; Glc,葡萄糖; GalNAc,N-乙酰氨基半乳糖。
和跨 非膜 极 性螺 侧旋 链的 分氨 布基 酸 极 性 侧 链
原基 典 性及 型 的二 的 细硫 跨 胞键 膜 内位 蛋 侧于 白 不细 : 形胞 成外 右 二侧 手 硫, 螺 键巯 旋 。基 跨 在膜 还, 糖
α
α
几种以β折叠跨膜的蛋白:不同数量的 β折叠链形成不同大小的β桶行使不同功能。 (1) The E. coli OmpA protein (8 β strands), which serves as a receptor for a bacterial virus. (2) The E. coli OMPLA protein (12 β strands), is a lipase(脂酶)that hydrolyses(水解)lipid molecules. The amino acids that catalyze the enzymatic reaction (shown in red) protrude(突出) from the outside surface of the barrel. (3) A porin(膜孔 蛋白,位于壁外膜) from the bacterium Rhodobacter capsulatus(一种光合细菌), which forms waterfilled pores across the outer membrane (16 β strands). The diameter of the channel is restricted by loops (shown in blue) that protrude into the channel. (4) The E. coli FepA protein (22 β strands), which transports iron ions. The inside of the barrel is completely filled by a globular (球状) protein domain (shown in blue) that contains an iron-binding site. This domain is thought to change its conformation(构象) to transport the bound iron, but the molecular details of the changes are not known.
重中素白细 要两蛋,菌 作个白多视 用极接个紫 。性受螺红 氨光旋质 基能中是 酸后央多 变是跨 构亲膜 在向水 胞性螺 次外质旋 第泵子结 传出通构 递 道膜 中,。整 起通在合 道色蛋 Asp H+ H+
绿色红假单胞菌 Rhodopseudomonas viridis 光反应中心复合体结构(x射线晶 体分析):The complex consists of four subunits, L, M, H, and a cytochrome(细胞 色素). The L and M subunits form the core of the reaction center, and each contains five a helices that span the lipid bilayer. The locations of the various electron carrier 荚膜红细菌 (Rhodobacter capsulatus,一种光合细菌)的膜孔蛋白三聚体 coenzymes are shown in black. (J. 结构:16条反向平行的β链形成内部亲水的β筒,三个β筒组成三聚体在细 Richardson and J. Deisenhofer Nature 胞壁外膜上运送亲水性物质,红色肽段位于中央控制扩散孔径。
根据与脂分子的结合方式分为: 3.2 膜蛋白 – 整合蛋白(integral protein) – 外周蛋白(peripheral protein) – 脂锚定蛋白(lipid-anchored protein)。
整合蛋白①②,外周蛋白⑤⑥,脂锚定蛋白③④。多数跨膜蛋白可能通过一个或多个α螺旋跨过脂双分子层,有的 通过β折叠跨膜, 它们中的有些蛋白还共价连接有插入脂双分子层单层中的脂肪酸链。脂锚定蛋白通过与蛋白共 价连接的脂肪酸链或异戊二烯链,插入细胞质一侧的脂单分子层中或通过低聚糖连接到细胞外侧脂单分子层中的磷 脂(磷脂酰肌醇)上。外周蛋白通过非共价相互作用方式与其他膜蛋白或带电荷的膜脂相连并附着在膜上。
第四章 细胞质膜
1 2 3 细胞质膜研究历史 生物膜结构模型 生物膜化学组成
4
5 6
生物膜特征
生物膜功能概述 膜骨架
•
细胞质膜是围绕细胞最 外层的一层由脂质和蛋 白质组成的生物膜。 质膜提供给细胞一个相 对稳定的内环境①。在 物质运输①③,能量转 换③ ,信息传递④⑤, 细胞连接和黏着⑤等方 面有重要作用。 真核细胞内部存在由膜 构建的各种细胞器,它 们有与质膜结构类似的 生物膜② 。
流动镶嵌模型和膜的流动性(其中成熟的膜蛋白在两层脂 分子间翻转不会发生)
3 生物膜的化学组成
3.1 膜脂:膜脂为两性分子兼具亲水性和疏水性。 • 膜脂主要有三种:磷脂、糖脂和固醇。 • 磷脂:甘油磷脂和鞘氨醇磷脂。约占膜脂的 50%以上。神经系统含鞘磷脂较丰富。 • 磷脂主要特征: • 一个极性头、两个非极性尾(脂肪酸链)。 • 脂肪酸碳链为偶数,16,18或20个碳原子。 • 常含有不饱和脂肪酸(如油酸)。
整合蛋白 • 靠跨膜结构域与膜中间疏水区相互作用 • 疏水结构域两端的带电荷的氨基酸(碱性或 酸性)与磷脂形成离子键整合于膜上。 • 通过半胱氨酸连接脂锚定于膜上。 • 在形成α螺旋或β折叠片时非极性氨基酸侧 链位于外部与膜脂疏水区相互亲和,而极性 侧链位于结构域内侧可形成特异或非特异极 性分子跨膜通道。 • 整合蛋白与膜的结合紧密,只有用去垢剂破 坏膜结构后才能从膜上洗涤下来。离子型去 垢剂常使蛋白结构改变而失活。 外周蛋白 • 靠非共价键作用方式结合于膜外周其他蛋白 或膜脂上。 • 通过改变溶液的离子强度或改变温度即可使 外周蛋白从膜上解离下来,而膜可不受破坏。 蛋白一般不失活。
•
•
固醇存在真核细胞和少数原核细胞质膜(如支原体)上,约占膜脂的1/3, 胆固醇存在于动物细胞膜中,植物细胞质膜含豆固醇等植物甾醇,真菌细 胞质膜含麦角固醇。大多数细菌质膜不含胆固醇,但含有甘油酯等中性酯。 在缺少胆固醇培养基中,不能合成胆固醇的突变细胞株很快发生自溶。
胆固醇结构
胆固醇在磷脂双分子层中嵌入单层磷脂分子间 缝隙中增加膜的稳定性,降低水溶性物质的通 透性。
R1
R2
R1
R2
R1
R2
R1
R2
PE
PS
PC
PI
鞘氨醇磷脂
四种主要磷脂
• 脂质体( 脂质体是一种人工膜。由磷脂在水中搅动后形成。 liposome) • 双层或单层脂分子球体,直径25~1000nm。 • 人工脂质体可用于: 1. 转基因 2. 制备的药物 3. 研究生物膜的特性