常用大肠杆菌 (K-12株来源) 的基因型
常用大肠杆菌 (K-12株来源) 的基因型

3[bTbZb KRX[NLQWXUXP_ LJ[JUXP XWURWN4grrnI>>sss=rbibpb=cmk=cl
2/01
?twv~~ ?twv~* ?twv*v
பைடு நூலகம்
bm[c;@4@9@jd]^efhZ_l n M6-* GP M,9yxx TGQF trs o N GKPDOQDCu
X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X \ ` g iX
wr 6@KG@QGPs ;us 0OGQPFs /u 0u @KC :@JAOLLHs 3u qw*~*r 5I`]WUf]Sc A]`_[_Y5 ? HST`cSe`ci IS_fS] xKCu DCu xr ?LRKEs 9u +u @KC .@SGPs 9u =u qw*~yr LU[W_UW AAAs }}~t}~xu yr ?@KGPBFt8DOOLKs -us <GDGO@s 3u @KC 6DPPGKEs 3u qw*~{r EW_W BBs wvytww*u
批注本地保存成功开通会员云端永久保存去开通
. u*wvy{ .3012,|+/ xz~}
:QO@GK ,,z ,5xw q./yr ,6x{u{ ,62}wtw~uyxq ,=ywy -tI@ -|vv -3xy| .2w .2{ .2{l .8{vwyvp /.~|{z /.~}|} /9w|z} 2,wvw 26:w}z 36~y 36wvw 36wv{ 36wv| 36wv} 36wv~ 36wv* 36wwv 4~vx 4~vy 5/y*x 6-wv|w 6<ww~z 6<ww*y 7LS@,IRD 99w ;+8*v ;1w ;1x ;2x >5wt,IRD
常用大肠杆菌 (K-12株来源) 的基因型
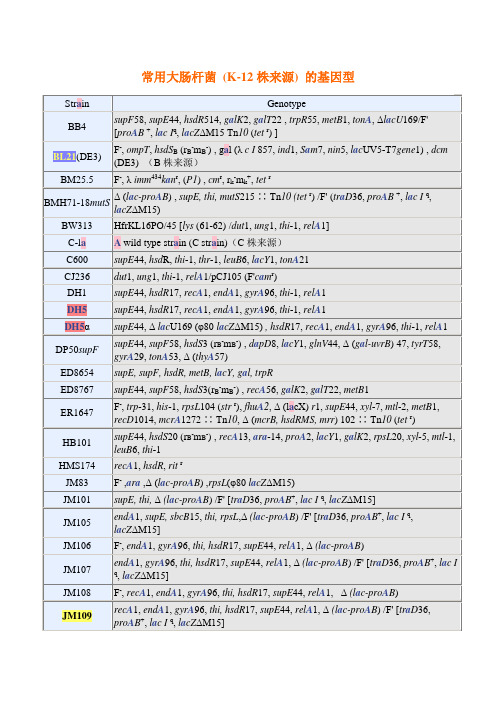
supE44,hsdR17,recA1,endA1,gyrA96,thi-1,relA1
DH5α
supE44, ΔlacU169 (φ80lacZΔM15), hsdR17,recA1,endA1,gyrA96,thi-1,relA1
DP50supF
supE44,supF58,hsdS3 (rB-mB-) ,dapD8,lacY1,glnV44, Δ (gal-uvrB) 47,tyrT58,gyrA29,tonA53, Δ (thyA57)
常用大肠杆菌(K-12株来源)的基因型
Strain
Genotype
BB4
supF58,supE44,hsdR514,galK2,galT22 ,trpR55,metB1,tonA, ΔlacU169/F' [proAB+,lac Iq,lacZΔM15 Tn10(tetr) ]
BL21(DE3)
F-,ompT,hsdSB(rB-mB-) , gal (λc I857,ind1,Sam7,nin5,lacUV5-T7gene1) ,dcm(DE3)(B株来源)
K803
F-,lacY1 or Δ (lac Iq-Y) 6 (lac-3) ,supE44,galK2,galT22,mcrA, rfbD,metB1,mcrB1,hsdR3
LE392
supE44,supF58,hsdR514,galK2,galT22,metB1,trpR55,lacY1
MC1061
hsdR, mcrB,araD139, Δ (araABCห้องสมุดไป่ตู้leu) 7679, ΔlacX74,galU, galK, rpsL, thi
MV1184
大肠杆菌表达系统

大肠杆菌表达系统总结随着分子生物学和蛋白组学的迅猛发展,外源基因表达的遗传操作技术日趋成熟。
表达系统是外源基因表达的核心,常用表达系统一般为模式生物,包括真核表达系统和原核表达系统,其中真核系统包括了哺乳动物细胞表达系统、植物体表达系统、昆虫杆状病毒表达载体系统以及酵母表达系统,原核表达系统则主要为大肠杆菌表达系统。
大肠杆菌是目前应用最广泛的原核表达系统,也是最早进行研究的外源基因表达系统,其遗传学背景清晰、生长快、较易实现高密度培养、成本低、产量高,相较于其它表达系统具有难以比拟的优越性,是商业生产中应用最广泛的表达系统,取得了巨大的科研价值和经济效益。
大肠杆菌表达系统目前广泛应用于表达生产多种蛋白质/多肽类药物和生物化学产品,包括:重组人胰岛素、a2b型干扰素、兰尼单抗、紫色杆菌素和牡丹皮葡萄糖苷等。
据统计,1986-2018年由美国FDA和欧洲EMA批准上市的重组蛋白类药物中有26%来自于大肠杆菌。
与此同时,目前通过大肠杆菌表达的基因工程疫苗也进入市场或处于临床实验阶段,如戊型肝炎疫苗、人乳头瘤病毒疫苗、流感A型疫苗等。
常见的大肠杆菌表达系统有BL21系列、JM109系列、 W3110系列和K802系列等,其中大肠杆菌 BL21( DE3)菌株是目前应用于重组蛋白表达研究最广泛的菌株之一,BL21(DE3)是由大肠杆菌B系列与K-12系列的衍生菌株通过 P1 转导等遗传突变获得的。
该类菌株通常为宿主蛋白酶缺失型,以保证外源蛋白在表达过程中不被降解,维持表达的稳定性。
大肠杆菌表达系统在商业生产中具有巨大的优越性和价值,但建立高效匹配的表达系统是实现商业价值的关键,包括宿主菌、外源基因、载体的选择与匹配。
宿主菌的选择是第一步,对表达活性和表达量影响很大,理想的宿主菌株是蛋白酶缺陷型,避免蛋白酶过多引起的产物不稳定,常见的蛋白酶缺陷型菌株为BL21系列菌株。
其次是外源基因,外源基因决定了是否可获得目的产物,原核基因可在大肠杆菌中直接表达,而真核基因不能再大肠杆菌中直接表达。
大肠杆菌基因工程菌常用类型

1、大肠杆菌DH5a菌株DH5a是世界上最常用的基因工程菌株之一。
由于DH5α是DNA酶缺陷型菌株,有利于基因克隆,保存质粒,但该菌株的蛋白酶没有缺陷,表达的蛋白容易被降解,因此通常不作为表达菌株。
E.coli DH5a在使用pUC系列质粒载体转化时,可与载体编码的β-半乳糖苷酶氨基端实现α-互补。
可用于蓝白斑筛选鉴别重组菌株。
基因型:F-,φ80dlacZΔM15,Δ(lacZYA-argF)U169,deoR,recA1,endA1,hsdR17(rk-,mk+),phoA,supE44,λ-,thi-1,gyrA96,relA12、大肠杆菌BL21(DE3) 菌株该菌株用于高效表达克隆于含有噬菌体T7启动子的表达载体(如pET系列)的基因。
T7噬菌体RNA聚合酶位于λ噬菌体DE3区,该区整合于BL21的染色体上。
该菌适合表达非毒性蛋白。
基因型:F-,ompT,hsdS(rBB-mB-),gal,dcm(DE3)3、大肠杆菌BL21(DE3) pLysS菌株该菌株含有质粒pLysS,因此具有氯霉素抗性。
PLysS含有表达T7溶菌酶的基因,能够降低目的基因的背景表达水平,但不干扰目的蛋白的表达。
该菌适合表达毒性蛋白和非毒性蛋白。
基因型:F-,ompT hsdS(rBB-mB-),gal,dcm(DE3,pLysS ,Camr4、大肠杆菌JM109菌株该菌株在使用pUC系列质粒载体进行DNA转化或用M13 phage载体进行转染时,由于载体DNA产生的LacZa多肽和JM09编码的LacZΔM15进行α-互补,从而显示β-半乳糖苷酶活性,由此很容易鉴别重组体菌株。
基因型:recA1,endA1,gyrA96,thi-1,hsdR17,supE44,relA1,Δ(lac-proAB)/F’[traD36,proAB+,lacIq,lacZΔM15]5、大肠杆菌TOP10菌株该菌株适用于高效的DNA克隆和质粒扩增,能保证高拷贝质粒的稳定遗传。
大肠杆菌基因组

大肠杆菌基因组
大肠杆菌(Escherichia coli)是一种常见的革兰氏阴性菌,广泛存在于自然界中,是人和动物肠道的正常菌群之一。
以下是大肠杆菌的基因组特点:
1. 基因组大小:大肠杆菌的基因组长度约为4.6-5.5百万个碱基对,包含了约4,000-5,500个基因。
2. 基因组结构:大肠杆菌的基因组呈圆形双链DNA分子,有一个单独的起始点和终止点。
3. GC含量:大肠杆菌的基因组GC含量约为50%,属于高GC菌株。
4. 基因功能:大肠杆菌的基因组包含了许多与代谢、运输、感应、合成等生命活动相关的基因,其中约1/3的基因没有已知的生物学功能。
5. 基因编码:大肠杆菌的基因组可以编码出各种蛋白质、RNA 和其他重要分子,如rRNA、tRNA、mRNA等。
6. 基因组变异:大肠杆菌基因组在不同的菌株之间存在着一定程度的变异,包括插入序列、转座子、基因重排等。
大肠杆菌的基因组研究对于了解其代谢、生长和适应环境能力等方面具有重要意义。
目前已经对大肠杆菌进行了多次基因组测序,并建立了完整的大肠杆菌K-12 MG1655基因组数据库,为大肠杆菌的进一步研究提供了强有力的工具。
细菌菌株基因型及基因符号说明

大肠杆菌基因型及遗传符号说明前言:实验室的一般大肠杆菌拥有4288条基因,每条基因的长度约为950bp,基因间的平均间隔为118bp(基因Ⅷ)。
E.coli基因组中还包含有许多插入序列,如λ-噬菌体片段和一些其他特殊组份的片段,这些插入的片段都是由基因的水平转移和基因重组而形成的,由此表明了基因组具有它的可塑造性。
利用大肠杆菌基因组的这种特性对其进行改造,使其中的某些基因发生突变或缺失,从而给大肠杆菌带来可以观察到的变化,这种能观察到的特征叫做大肠杆菌的表现型(Phenotype),把引起这种变化的基因构成叫做大肠杆菌的基因型(Genotype)。
具有不同基因型的菌株表现出不同的特性。
分子克隆中常用的大肠杆菌及其遗传标记按Demerec等1966年提出的命名原则,采用的菌株所有的基因都假定处于野生型状态,除非在基因型上另外注明。
大肠杆菌基因型的表示方法(Demerec, et, al. 1966):一、一般规则:1、根据基因产物或其作用产物的英文名称的第一个字母缩写成3个小写斜体字母来表示。
例如:DNA Adenine Methylase→dam。
2、不同的基因座,其中任何一个突变所产生的表型变化可能相同,其表示方法是在3个小写斜体字母后加上一个斜体大写字母来表示区别。
例如:Recombination→recA、recB、recC。
3、突变位点应通过在突变基因符号后加不同数字表示。
如supE44(sup基因座E的44位突变)。
如果不知道几个等位基因中哪一/几个发生了功能性突变,则用连字符“-”代替大写字母,如trp-31。
4、细菌的基因型中应该包含关于其携带的质粒或附加体的的信息。
这些符号包括菌株携带的质粒或附加体、质粒或附加体上的突变基因座和突变位点。
其基因符号应与基因座的表示符号明显区别,符号的第一个字母大写、不斜体并位于括号内;质粒或附加体上的突变基因座和突变位点的基因符号的表示方法与染色体上突变基因座、突变位点的符号相同。
大肠杆菌的基因型-概述说明以及解释

大肠杆菌的基因型-概述说明以及解释1.引言1.1 概述大肠杆菌是一种常见的革兰氏阴性杆菌,属于肠道菌群中的重要成员。
它在自然界和人体内广泛存在,并且具有广泛的基因型多样性。
这使得大肠杆菌成为了微生物遗传学和进化生物学领域的研究模型。
在大肠杆菌中,基因型是指该菌株拥有的基因组合和基因的分布情况。
大肠杆菌的基因型可以通过不同的方法进行分类和鉴定。
目前主要的分类方法包括单核苷酸多态性分析、基因片段分析和全基因组测序等。
通过这些方法,我们可以更全面地了解大肠杆菌的基因型组成和种群结构。
大肠杆菌的基因型在其功能和特点方面具有重要意义。
大肠杆菌是一种典型的益生菌,它在人体内具有多种有益作用,包括帮助消化吸收、维持肠道稳定性和参与免疫调节等。
不同基因型的大肠杆菌可能具有不同的功能特点,比如某些基因型可能携带耐药基因或致病因子,导致感染和疾病的发生。
因此,对大肠杆菌基因型的研究有助于我们深入了解其功能机制和生态适应能力。
总之,大肠杆菌作为一种常见的菌株,其基因型具有多样性和重要性。
通过研究大肠杆菌的基因型,我们可以深入探索其功能特点和生态适应能力,进一步促进微生物遗传学和进化生物学的研究。
未来,我们可以通过结合多样的研究方法和技术,进一步挖掘和解析大肠杆菌基因型的奥秘,并探索其在人体健康和疾病中的作用。
文章结构是指文章部分之间的逻辑关系和组织,它有助于读者理解文章的内容和思路。
本文的结构如下:1. 引言1.1 概述1.2 文章结构1.3 目的2. 正文2.1 大肠杆菌的基因型分类2.2 大肠杆菌基因型的功能和特点3. 结论3.1 大肠杆菌基因型的重要性3.2 未来研究的方向文章结构部分是为了描述本文的组织结构,它有助于读者了解文章的内容安排和逻辑关系。
在本文中,我们首先介绍引言部分,包括概述、文章结构和目的。
在概述中,我们简要介绍了大肠杆菌的基因型。
在文章结构中,我们明确了本文的结构和章节安排,帮助读者理解文章的整体框架。
细菌基因型及其基因符号1102

大肠杆菌基因型及遗传符号说明前言:实验室的一般大肠杆菌拥有4288条基因,每条基因的长度约为950bp,基因间的平均间隔为118bp(基因Ⅷ)。
E.coli基因组中还包含有许多插入序列,如λ-噬菌体片段和一些其他特殊组份的片段,这些插入的片段都是由基因的水平转移和基因重组而形成的,由此表明了基因组具有它的可塑造性。
利用大肠杆菌基因组的这种特性对其进行改造,使其中的某些基因发生突变或缺失,从而给大肠杆菌带来可以观察到的变化,这种能观察到的特征叫做大肠杆菌的表现型(Phenotype),把引起这种变化的基因构成叫做大肠杆菌的基因型(Genotype)。
具有不同基因型的菌株表现出不同的特性。
分子克隆中常用的大肠杆菌及其遗传标记按Demerec等1966年提出的命名原则,采用的菌株所有的基因都假定处于野生型状态,除非在基因型上另外注明。
大肠杆菌基因型的表示方法(Demerec, et, al. 1966):一、一般规则:1、根据基因产物或其作用产物的英文名称的第一个字母缩写成3个小写斜体字母来表示。
例如:DNA Adenine Methylase→dam。
2、不同的基因座,其中任何一个突变所产生的表型变化可能相同,其表示方法是在3个小写斜体字母后加上一个斜体大写字母来表示区别。
例如:Recombination→recA、recB、recC。
3、突变位点应通过在突变基因符号后加不同数字表示。
如supE44(sup基因座E的44位突变)。
如果不知道几个等位基因中哪一/几个发生了功能性突变,则用连字符“-”代替大写字母,如trp-31。
4、细菌的基因型中应该包含关于其携带的质粒或附加体的的信息。
这些符号包括菌株携带的质粒或附加体、质粒或附加体上的突变基因座和突变位点。
其基因符号应与基因座的表示符号明显区别,符号的第一个字母大写、不斜体并位于括号内;质粒或附加体上的突变基因座和突变位点的基因符号的表示方法与染色体上突变基因座、突变位点的符号相同。
大肠杆菌克隆菌株TOP10和DH5α的比较

大肠杆菌克隆菌株TOP10和DH5α的比较大肠杆菌克隆菌株是一种经过基因工程改造的细菌,可以用于转化和扩增外源DNA,如质粒、文库或表达载体。
不同的克隆菌株具有不同的特性,如转化效率、抗性、突变、表达水平等。
本文将比较两种常见的大肠杆菌克隆菌株:TOP10和DH5α,分析它们的优缺点和价格。
TOP10的优缺点TOP10是一种来源于大肠杆菌K12的菌株,具有mcr/mrr突变,可以克隆甲基化的DNA,如真核基因组DNA。
它的基因型如下:•Top10:F- mcrA Δ(mrr-hsdRMS-mcrBC) φ80lacZΔM15 ΔlacX74 recA1 araD139 Δ(ara-leu)7697 galU galK rpsL (StrR) endA1 nupGTOP10的优点主要有以下几个方面:•高转化效率:TOP10具有高达10^9 cfu/µg DNA的转化效率,这意味着它可以获得更多的重组菌株,提高克隆的成功率。
•低突变率:TOP10具有recA1突变,可以抑制同源重组,保持质粒的序列完整性。
•高稳定性:TOP10具有endA1突变,可以阻断内切核酸酶的表达,减少质粒DNA的降解。
它还可以在不含抗生素的培养基中维持质粒的复制。
•广泛的耐药性:TOP10具有多种抗性基因,如rpsL (StrR)、nupG等,可以使用多种抗生素进行筛选。
•高数据一致性:TOP10具有高度的基因组稳定性和质粒保留性,可以保证实验的可重复性和可靠性。
TOP10的缺点主要有以下几个方面:•高成本:TOP10的价格相对较高,可能超出一些实验室的预算。
•低表达水平:TOP10不适合用于蛋白表达,因为它缺乏一些必要的表达因子,如DE3溶原菌、T7 RNA聚合酶等。
•低兼容性:TOP10不适合用于一些特殊的载体,如含有重复序列的慢病毒载体和其它逆转录病毒载体,因为它可能会发生质粒的重组或缺失。
DH5α的优缺点DH5α是一种由Messing在1975年从大肠杆菌K12菌株中构建而成的菌株,具有mcr/mrr突变,可以克隆甲基化的DNA,如真核基因组DNA。
大肠杆菌基因型及遗传符号说明

大肠杆菌基因型及遗传符号说明前言:实验室的一般大肠杆菌拥有4288条基因,每条基因的长度约为950bp,基因间的平均间隔为118bp(基因Ⅷ)。
E.coli基因组中还包含有许多插入序列,如λ-噬菌体片段和一些其他特殊组份的片段,这些插入的片段都是由基因的水平转移和基因重组而形成的,由此表明了基因组具有它的可塑造性。
利用大肠杆菌基因组的这种特性对其进行改造,使其中的某些基因发生突变或缺失,从而给大肠杆菌带来可以观察到的变化,这种能观察到的特征叫做大肠杆菌的表现型(Phenotype),把引起这种变化的基因构成叫做大肠杆菌的基因型(Genotype)。
具有不同基因型的菌株表现出不同的特性。
分子克隆中常用的大肠杆菌及其遗传标记按Demerec等1966年提出的命名原则,采用的菌株所有的基因都假定处于野生型状态,除非在基因型上另外注明。
大肠杆菌基因型的表示方法(Demerec, et, al. 1966):一、一般规则:1、根据基因产物或其作用产物的英文名称的第一个字母缩写成3个小写斜体字母来表示。
例如:DNA Adenine Methylase→dam。
2、不同的基因座,其中任何一个突变所产生的表型变化可能相同,其表示方法是在3个小写斜体字母后加上一个斜体大写字母来表示区别。
例如:Recombination→recA、recB、recC。
3、突变位点应通过在突变基因符号后加不同数字表示。
如supE44(sup基因座E的44位突变)。
如果不知道几个等位基因中哪一/几个发生了功能性突变,则用连字符“-”代替大写字母,如trp-31。
4、细菌的基因型中应该包含关于其携带的质粒或附加体的的信息。
这些符号包括菌株携带的质粒或附加体、质粒或附加体上的突变基因座和突变位点。
其基因符号应与基因座的表示符号明显区别,符号的第一个字母大写、不斜体并位于括号内;质粒或附加体上的突变基因座和突变位点的基因符号的表示方法与染色体上突变基因座、突变位点的符号相同。
2020大肠杆菌的分型方法及其研究进展

2020大肠杆菌的分型方法及其研究进展肠埃希氏菌是一种条件性致病菌,致病性的大肠埃希氏菌具有高度的传染性,会严重危害健康。
快速准确地测定大肠埃希氏菌的污染来源对有效缩小疫情影响范围极有帮助,从而避免对人类健康和经济贸易造成重大损失。
建立简便高效的分型方法是微生物溯源的关键,常见的大肠埃希氏菌分型方法可分为表型分型和分子分型,这些分型方法各有优劣,具有不同的适用范围。
大肠埃希氏菌(Escherichia coli,E. coli)又称大肠杆菌,属革兰氏阴性菌,于1885年首次被发现[1],是人和动物体内的正常寄居菌。
大肠埃希氏菌是一种条件致病菌,在正常情况下不致病,然而一些特殊血清型菌株可导致人或动物腹泻、腹痛甚至会产血性粪便,重症病例会并发溶血性的尿毒综合征以及血小板减少性紫癜[2-3]。
根据大肠埃希氏菌对人类的致病机理不同,可将其分为5种类型:.肠致病性大肠埃希氏菌(Enteropathogenic escherichia coli,EPEC).肠产毒性大肠埃希氏菌(Enterotoxigenic escherichia coli,ETEC).肠侵袭性大肠埃希氏菌(Enteroinvasive escherichia coli,EIEC).肠出血性大肠埃希氏菌(Enterohemorrhagic escherichia coli,EHEC) .肠集聚性大肠埃希氏菌(Enteroaggregative escherichia coli,EAEC) 大肠埃希氏菌的肠道传染具有比较广泛的特性,而食品在生产、包装及运输过程中极易感染此菌,进而引发传染性疾病[5]。
2018年6月,大肠埃希氏菌O157:H7污染生菜事件的暴发,影响了美国36个州,事件造成96人住院和5人死亡[6]。
2019年4月,美国的10个州暴发牛肉感染大肠埃希氏菌O103事件,导致177人感染,其中21人住院,涉事公司紧急召回了53 200磅牛肉[7]。
MG1655(de3)感受态细胞使用方法

MG1655(DE3) 感受态细胞MG1655(DE3) Chemically Competent CellHuayueyang9982 北京华越洋生物MG1655(DE3) 感受态细胞10x100ul备注:以上包装均含有Compcell Control Plasmid pUC19(0.1ng/ul)5ul(质量控制用)。
储存:-70℃保存六个月。
MG1655(DE3) 感受态细胞介绍:MG1655(DE3)感受态细胞是采用特殊工艺处理得到的感受态细胞,可用于DNA 的化学转化。
基因型为:K12 F入-ilvG-rfb-50 rph-1(DE3)MG1655(DE3) 感受态细胞特点:MG1655(DE3)菌株来源于W1485,是K12 的衍生菌株,是一种经过较少改造、比较接近于“野生型”的大肠杆菌工程菌株。
MG1655(DE3)外观形态标准,既可做为扩增大肠杆菌野生型基因的横板使用也可作为蛋白表达的宿主菌株使用;T7 RNA 聚合酶位于入噬菌体DE3 区,该区域被整合于MG1655 的染色体上,该菌株不含核酸酶endAl 突变。
操作步骤:MG1655(DE3) 感受态细胞以下操作均按无菌条件的标准进行:转化:取感受态细胞置于冰浴中(解冻1-2分钟),加入目的DNA.轻轻混匀,在冰浴中放置30 分钟。
注意:所使用DNA 体积不要超过感受态细胞悬液体积的1/10.■热激:将离心管置于42℃水浴中放置60-90 秒,然后快速将管转移到冰浴中,使细胞冷却2-3分钟,该过程不要摇动离心管。
■复苏:向每个离心管中加入500ul 无菌的SOC 或LB培养基(不含抗生素),混匀后置于37℃180rpm摇床振荡培养45-60 分钟,目的是使质粒上相关的抗性标记基因表达,使菌体复苏。
口涂板:根据实验要求(质粒,重组连接产物转化),吸取适量体积已转化的感受态细胞加到含相应抗生素的SOC 或LB 固体琼脂培养基上,将细胞均匀涂开。
大肠杆菌的基本知识概述

⼤肠杆菌的基本知识概述2019-08-31⼤肠杆菌是重要的模式⽣物,在⾼中⽣物学中有.多处知识涉及到,如原核⽣物的基本结构与分裂、T2’噬菌体侵染细菌实验、基因⼯程常见运载体、⼤肠杆菌的鉴定、微⽣物的酶合成调节等。
这些知识涉及点多,分布零散,缺乏系统性,下⾯就⼤肠杆菌的基本知识作简要概述。
1 认识⼤肠杆菌的基本历程⼤肠杆菌是⼤肠埃希⽒菌的俗称,属肠杆菌科埃希⽒菌属,1885年埃舍利希⽒⾸次发现。
在相当长的时间内,⼈们⼀直把它当作正常肠道菌群的组成部分,认为是⾮致病菌。
直到20世纪中叶,才认识到⼀些特殊⾎清型的⼤肠杆菌对⼈和动物有病原性,尤其对婴⼉和幼禽,常引起严重腹泻和败⾎症。
根据不同的⽣物学特性,将致病性⼤肠杆菌分为5类:致病性⼤肠杆菌(EPEC)、肠产毒性⼤肠杆菌(ETEC)、肠出⾎性⼤肠杆菌(EHEC)、肠侵袭性⼤肠杆菌(EIEC)、肠黏附性⼤肠杆菌(EAEC)。
2 ⼤肠杆菌的基本结构2.1 细胞壁⼤肠杆菌的细胞壁厚约11µm,分外膜和肽聚糖层。
外膜是⼤肠杆菌细胞壁的主要成分,占细胞壁⼲重80%,位于肽聚糖层的外侧,主要由磷脂、蛋⽩质和脂多糖组成。
脂多糖是⾰兰⽒阴性细菌的内毒素,也是⾰兰⽒阴性细菌细胞壁的特有成分,主要与抗原性、致病性及对噬菌体的敏感性有关。
肽聚糖层由1―2层⽹状的肽聚糖组成,占细胞壁⼲重的10%,是细菌特有的成分,由聚糖链、短肽和肽桥三部分组成。
由,脂蛋⽩将外膜和肽聚糖层连接起来,从⽽使⼤肠杆菌的细胞壁形成⼀个整体结构。
2.2 细胞膜⼤肠杆菌的细胞膜与其他⽣物细胞膜的结构相似,但上⾯的蛋⽩质含量⾼、种类多,具有选择透性,可控制营养物质进出细胞。
⼤肠杆菌的细胞膜含有丰富的酶系,是⼤肠杆菌的能量转化的场所,参与细胞壁的合成。
2.3 细胞质⼤肠杆菌的细胞质含有糖原颗粒、核糖体和质粒等结构。
2.3.1 核糖体⼤肠杆菌的核糖体包含两个亚基,即50S亚基(23S rRNA、5S rRNA、34种蛋⽩质)和.30S亚基[16SrRNA、21种蛋⽩质(S1~S21)]。
大肠杆菌

大肠杆菌百科名片肠埃希氏菌(E. coli)通常称为大肠杆菌,是Escherich在1885年发现的,在相当长的一段时间内,一直被当作正常肠道菌群的组成部分,认为是非致病菌。
直到20世纪中叶,才认识到一些特殊血清型的大肠杆菌对人和动物有病原性,尤其对婴儿和幼畜(禽),常引起严重腹泻和败血症,它是一种普通的原核生物,根据不同的生物学特性将致病性大肠杆菌分为5类:致病性大肠杆菌(EPEC)、肠产毒性大肠杆菌(ETEC)、肠侵袭性大肠杆菌(EIEC)、肠出血性大肠杆菌(EHEC)、肠黏附性大肠杆菌(EAEC)。
大肠杆菌属于细菌。
中文名称:大肠杆菌外文名称:学名:Escherichia coli (T.Escherich 1885) 界:细菌界门:变形菌门(Bacteria) 纲:γ-变形菌纲(Proteobacteria) 目:肠杆菌目(Enterobacteriales) 科:肠杆菌科(Enterobacteriaceae) 属:埃希氏菌属(Escherichia) 种:大肠杆菌种(E. coli)[编辑本段]常见种类介绍大肠杆菌大肠杆菌0 157:H7血清型属肠出血性大肠杆菌,自1982年在美国首先发现以来,包括我国等许多国家都有报道,且日见增加。
日本近年来因食物污染该菌导致的数起大暴发,格外引人注目。
在美国和加拿大通常分离的肠道致病菌中,目前它已排在第二或第三位。
大肠杆菌O 157:H7引起肠出血性腹泻,约2%~7%的病人会发展成溶血性尿毒综合征,儿童与老人最容易出现后一种情况。
致病性大肠杆菌通过污染饮水、食品、娱乐水体引起疾病暴发流行,病情严重者,可危急生命。
大肠杆菌(Escherichia coli,E.coli)革兰氏阴性短杆菌,大小0.5×1~3微米。
周身鞭毛,能运动,无芽孢。
能发酵多种糖类产酸、产气,是人和动物肠道中的正常栖居菌,婴儿出生后即随哺乳进入肠道,与人终身相伴,其代谢活动能抑制肠道内分解蛋白质的微生物生长,减少蛋白质分解产物对人体的危害,还能合成维生素B和K,以及有杀菌作用的大肠杆菌素。
大肠杆菌基因型及遗传符号说明系列一DXY

大肠杆菌基因型及遗传符号说明系列一点击次数:982 作者:佚名发表于:2009-09-27 00:00转载请注明来自丁香园来源:丁香园实验室的一般大肠杆菌拥有4288条基因,每条基因的长度约为950bp,基因间的平均间隔为118bp (基因Ⅷ)。
E.coli基因组中还包含有许多插入序列,如λ-噬菌体片段和一些其他特殊组份的片段,这些插入的片段都是由基因的水平转移和基因重组而形成的,由此表明了基因组具有它的可塑造性。
利用大肠杆菌基因组的这种特性对其进行改造,使其中的某些基因发生突变或缺失,从而给大肠杆菌带来可以观察到的变化,这种能观察到的特征叫做大肠杆菌的表现型(Phenotype),把引起这种变化的基因构成叫做大肠杆菌的基因型(Genotype)。
具有不同基因型的菌株表现出不同的特性。
分子克隆中常用的大肠杆菌及其遗传标记按Demerec等1966年提出的命名原则,采用的菌株所有的基因都假定处于野生型状态,除非在基因型上另外注明。
大肠杆菌基因型的表示方法(Demerec, et, al. 1966):一、一般规则:1、根据基因产物或其作用产物的英文名称的第一个字母缩写成3个小写斜体字母来表示。
例如:D NA Adenine Methylase→dam。
2、不同的基因座,其中任何一个突变所产生的表型变化可能相同,其表示方法是在3个小写斜体字母后加上一个斜体大写字母来表示区别。
例如:Recombination→recA、recB、recC。
3、突变位点应通过在突变基因符号后加不同数字表示。
如supE44(sup基因座E的44位突变)。
如果不知道几个等位基因中哪一/几个发生了功能性突变,则用连字符“ -”代替大写字母,如trp-31。
4、细菌的基因型中应该包含关于其携带的质粒或附加体的的信息。
这些符号包括菌株携带的质粒或附加体、质粒或附加体上的突变基因座和突变位点。
其基因符号应与基因座的表示符号明显区别,符号的第一个字母大写、不斜体并位于括号内;质粒或附加体上的突变基因座和突变位点的基因符号的表示方法与染色体上突变基因座、突变位点的符号相同。
大肠埃希氏菌K-12糖代谢的外膜蛋白质组学研究的开题报告

大肠埃希氏菌K-12糖代谢的外膜蛋白质组学研究的开题报告一、研究背景和意义大肠杆菌(Escherichia coli)是一种广泛存在于自然界中的革兰氏阴性菌。
它存在于大部分生物体内,包括人体肠道中。
大肠杆菌K-12在比较基因组学的研究中是一个重要的模式菌株。
大肠杆菌K-12是已知最好的微生物模型之一,广泛用于生物化学、遗传学、生物物理学和分子生物学的研究中。
因此,大肠杆菌K-12的研究对于我们理解生命基本过程和发展新型治疗方法具有重要意义。
糖代谢是维持生命活动的重要过程之一,大肠杆菌K-12在糖代谢过程中起着重要的作用。
外膜蛋白质是外露在大肠杆菌K-12胞外的一类重要蛋白质,它们在细菌细胞质膜的合成、通透性等方面起到至关重要的作用。
因此,揭示大肠杆菌K-12糖代谢相关的外膜蛋白质谱组学特征,有助于深入了解大肠杆菌K-12的糖代谢过程及其调节机制,为开发针对大肠杆菌K-12的治疗方法提供理论依据。
二、研究内容和方法本研究旨在通过外膜蛋白质组学的方法,揭示大肠杆菌K-12的糖代谢相关外膜蛋白质的谱组学特征,具体研究内容包括以下几个方面:1. 确定大肠杆菌K-12在糖代谢过程中相关外膜蛋白质的表达情况。
2. 应用蛋白质组学技术,进行大肠杆菌K-12糖代谢相关的外膜蛋白质的筛选和鉴定。
3. 进行外膜蛋白质的定量和定位分析,以确定不同条件下外膜蛋白质的表达量变化以及它们在细菌胞外的分布情况。
4. 对蛋白质组学数据进行生物信息学分析,全面解析糖代谢相关外膜蛋白质的功能、亚细胞定位和代谢途径。
本研究将结合DNA微阵列、蛋白质组学技术、定量PCR等方法,筛选大肠杆菌K-12在糖代谢过程中的相关外膜蛋白质、分析它们的表达情况和分布特征、揭示它们在大肠杆菌K-12代谢途径中的功能及其调节机制。
三、研究期望和创新点本研究旨在揭示大肠杆菌K-12糖代谢过程中涉及的外膜蛋白质的表达情况和分布特征,为深入理解大肠杆菌K-12糖代谢过程提供全面的信息。
国家批准的菌株号

国家批准的菌株号国家批准的菌株号是指经过中国科学院微生物文化集中分发管理中心或其他相关部门正式批准并登记的微生物菌株编号。
该编号通常由两部分组成:前缀部分(以字母或数字表示菌株类型或来源)和后缀部分(以数字表示该菌株在编号体系中的序号)。
菌株是指从自然界或人工选育中获得的、具有明显特征和遗传基础的微生物单体。
菌株在微生物学研究中具有重要的作用,可用于基础科学研究、生产与应用、环境监测等领域。
国家批准的菌株号作为微生物菌株的唯一标识符,具有以下几个重要特点:1.准确性:国家批准的菌株号是由专业机构或相关部门负责管理和维护的,保证了菌株号的准确性和稳定性。
2.可追溯性:菌株号能够追溯到具体的微生物文化集中分发管理中心或相关部门,确保了菌株来源和性质的可靠性。
3.规范性:菌株号采用标准化命名法,为微生物菌株命名提供了参考和规范,避免了同一菌株出现重复命名的情况。
4.信息共享性:国家批准的菌株号提供了科学家之间信息共享的平台,为微生物学研究提供了便利。
目前,国家批准的菌株号已经广泛应用于微生物学研究的不同领域,如基础研究、生产与应用、环境监测等。
本文将介绍若干具有代表性的国家批准的菌株号。
1. 铜绿假单胞菌 ATCC 13985铜绿假单胞菌菌株 ATCC 13985 是一株可引起人和动物感染的病原菌。
该菌株编号由“ATCC”代表该菌株来自于美国 ATCC(American Type Culture Collection)菌株库,编号为“13985”。
2. 大肠杆菌 K-12 MG16553. 白腐霉 Trichoderma reesei QM6a白腐霉菌株 Trichoderma reesei QM6a 是一种被广泛应用于生产纤维素酶的工业菌株。
该菌株编号由 JCM(Japan Collection of Microorganisms)代表该菌株来自日本 JCM 菌株库,编号为“JCM 6165”。
4. 肝炎病毒 HBV AD38肝炎病毒菌株 HBV AD38 是一种常见的乙型肝炎病毒菌株。
大肠杆菌基因型解读

⼤肠杆菌基因型解读E. coli genotypesContents[hide]1 Nomenclature & Abbreviations2 Methylation Issues in E. coli3 Commonly used strains3.1 AG13.2 AB11573.3 BL213.4 BL21(AI)3.5 BL21(DE3)3.6 BL21 (DE3) pLysS3.7 BNN933.8 BNN973.9 BW26434, CGSC Strain # 76583.10 C6003.11 C600 hflA150 (Y1073, BNN102)3.12 CSH503.13 D12103.14 DB3.13.15 DH13.16 DH5α3.17 DH10B (Invitrogen)3.18 DH12S (Invitrogen)3.19 DM1 (Invitrogen)3.20 E. cloni(r) 5alpha (Lucigen)3.21 E. cloni(r) 10G (Lucigen)3.22 E. cloni(r) 10GF' (Lucigen)3.23 E. coli K12 ER2738 (NEB)3.24 ER2566 (NEB)3.25 ER2267 (NEB)3.26 HB1013.27 HMS174(DE3)3.28 High-Control(tm) BL21(DE3) (Lucigen) 3.29 High-Control(tm) 10G (Lucigen)3.30 IJ11263.31 IJ11273.32 JM833.33 JM1013.34 JM1033.35 JM1053.36 JM1063.37 JM1073.38 JM1083.39 JM1093.40 JM109(DE3)3.41 JM1103.42 JM2.3003.43 LE3923.44 Mach13.45 MC10613.46 MC41003.47 MG16553.48 OmniMAX23.49 OverExpress(tm)C41(DE3) (Lucigen)3.50 OverExpress(tm)C41(DE3)pLysS (Lucigen) 3.51 OverExpress(tm)C43(DE3) (Lucigen)3.52 OverExpress(tm)C43(DE3)pLysS (Lucigen) 3.53 Rosetta(DE3)pLysS3.54 Rosetta-gami(DE3)pLysS3.55 RR13.56 RV3083.57 SOLR (Stratagene)3.58 SS320 (Lucigen)3.59 STBL2 (Invitrogen)3.60 STBL3 (Invitrogen)3.61 STBL43.62 SURE (Stratagene)3.63 SURE2 (Stratagene)3.64 TG1 (Lucigen)3.65 TOP10 (Invitrogen)3.66 Top10F' (Invitrogen)3.67 W31103.68 XL1-Blue (Stratagene)3.69 XL1-Blue MRF' (Stratagene)3.70 XL2-Blue (Stratagene)3.71 XL2-Blue MRF' (Stratagene)3.72 XL1-Red (Stratagene)3.73 XL10-Gold (Stratagene)3.74 XL10-Gold KanR (Stratagene)4 Other genotype information sources5 ReferencesNomenclature & AbbreviationsA listed gene name means that gene carries a loss of function mutation, a Δ preceding a gene name means the gene is deleted. If a gene is not listed, it is not known to be mutated. Prophages present in wt K-12 strains (F, λ, e14, rac) are listed only if absent. E. coliB strains are naturally lon- and dcm-.F- = Does not carry the F plasmidF+ = Carries the F plasmid. The cell is able to mate with F- through conjugation.F'[ ] = Carries an F plasmid that has host chromosomal genes on it from a previous recombination event. This cell can also mate with F- through conjugation. Chromosomal genes carried in the F plasmid are listed in brackets.r B/K+/- = The (B/K) defines the strain lineage. The +/- indicates whether the strain has or hasn't got the restriction system. m B/K+/- = The (B/K) defines the strain lineage. The +/- indicates whether the strain has or hasn't got the modification (methylation) system.hsdS = Both restriction and methylation of certain sequences is deleted from the strain. If you transform DNA from such a strain into a wild type strain, it will be degraded.hsdR = For efficient transformation of cloned unmethylated DNA from PCR amplificationsINV( ) = chromosomal inversion between locations indicatedahpC = mutation to alkyl hydroperoxide reductase conferring disulfide reductase activityara-14 = cannot metabolize arabinosearaD = mutation in L-ribulose-phosphate 4-epimerase blocks arabinose metabolismcycA = mutation in alanine transporter; cannot use alanine as a carbon sourcedapD = mutation in succinyl diaminopimelate aminotransferase leads to succinate or (lysine + methionine) requirementΔ( ) = chromosomal deletion of genes between the listed genes (may include unlisted genes!)dam = adenine methylation at GATC sequences exist; high recombination efficiency; DNA repair turned ondcm = cytosine methylation at second C of CCWGG sites exist. dam & dcm are the default properties and always elided, while dam- or dcm- should be declare explicitlydeoR = regulatory gene that allows constitutive expression of deoxyribose synthesis genes; permits uptake of largeplasmids. See Hanahan D, US Patent 4,851,348. ***This has been called intoquestion, as the DH10B genome sequence revealed that it is deoR+. See Durfee08, PMID 18245285.dnaJ = one of the chaparonins inactivated; stabilizes some mutant proteinsdut1 = dUTPase activity abolished, leading to increased dUTP concentrations, allowing uracil instead of thymine incorporation in DNA. Stable U incorporation requires ung gene mutation as well. endA1 = For cleaner preparations of DNA and better results in downstream applications due to the elimination of non-specific digestion by Endonuclease I(e14) = excisable prophage like element containing mcrA gene; present in K-12 but missing in many other strainsgalE = mutations are associated with high competence, increased resistance to phage P1 infection, and 2-deoxygalactose resistance. galE mutations block the production of UDP-galactose, resulting in truncation of LPS glycans to the minimal, "inner core". The exceptional competence ofDH10B/TOP10 is thought to be a result of a reduced interference from LPS in the binding and/or uptake of transforming DNA. galE15 is a point mutation resulting in a Ser123 -> Phe conversion near the enzyme's active site. See van Die, et al. PMID 6373734, Hanahan, et al. PMID 1943786, and EcoSal ISBN 1555811647. --Dcekiert 16:56, 23 January 2008 (CST)galk = mutants cannot metabolize galactose and are resistant to 2-deoxygalactose. galK16 is anIS2 insertion ~170bp downstream of the galK start codon. See EcoSal ISBN 1555811647. --Dcekiert 16:56, 23 January 2008 (CST)galU = mutants cannot metabolize galactosegor = mutation in glutathione reductase; enhances disulphide bond formationglnV = suppression of amber (UAG) stop codons by insertion of glutamine; required for some phage growthgyrA96 = mutation in DNA gyrase; conveys nalidixic acid resistancegyrA462 = mutation in DNA gyrase; conveys resistance to ccdB colicin gene producthflA150 = protease mutation stabilizing phage cII protein; high frequency of lysogenization by λΔ(lac)X74 = Deletion of the entire lac operon as well as some flanking DNA (complete deletion is Δcod-mhpF; seeMol.Micro., 6:1335, and J.Bact., 179:2573)lacI q or lacI Q = overproduction of the lac repressor protein; -35 site in promoter upstream of lacI is mutated from GCGCAA to GTGCAAlacI Q1 = overproduction of the lac repressor protein; contains a 15 bp deletion to create optimal -35site in promoter upstream of lacIlacY = deficient in lactose transport; deletion of lactose permease (M protein)lacZΔM15 = partial deletion of the lacZ gene that allows α complementation of the β-galactosidase gene; required forblue/white selection on XGal plates. Deletes the amino portion of lacZ (aa 11-41). leuB = requires leucineΔlon = deletion of the lon proteasemalA = cannot metabolize maltosemcrA = Mutation eliminating restriction of DNA methylated at the sequence C m CGG (possiblym CG). Carried on the e14 prophage (q.v.)mcrB = Mutation eliminating restriction of DNA methylated at the sequence R m CmetB = requires methioninemetC = requires methioninemrr = Mutation eliminating restriction of DNA methylated at the sequence C m AG or G m ACmtlA = cannot metabilize mannitol(Mu) = Mu prophage present. Muδ means the phage is defective.mutS - mutation inhibits DNA repair of mismatches in unmethylated newly synthesized strands nupG = same as deoR ompT = mutation in outer membrane protein protease VII, reducing proteolysis of expressed proteins(P1) = Cell carries a P1 prophage. Cells express the P1 restriction system.(P2) = Cell carries a P2 prophage. Allows selection against Red+ Gam+ λ(φ80) = Cell carries the lambdoid prophage φ80. A defective version of this phage carryinglacZM15 deletion (as well as wild-type lacI, lacYA, and flanking sequences) is present in some strains. The φ80 attachment site is just adjacent to tonB.pLysS = contains pLysS plasmid carrying chloramphenicol resistance and phage T7 lysozyme, effective at attenuating activity of T7 RNA polymerase, for better inhibition of expression under non-induced conditions. The sequence can be found here.proA/B = requires prolinerecA1 = For reduced occurrence of unwanted recombination in cloned DNA; cells UV sensitive, deficient in DNA repair recA13 = as for recA1, but inserts less stable.recBCD = Exonuclease V; mutation in RecB or RecC reduces general recombination by a factor of 100; impaired DNA repair; UV sensitive, easier propagation of inverted repeatsrecJ Exonuclease involved in alternate recombinationrelA = relaxed phenotype; permits RNA synthesis in absence of protein synthesisrha = blocked rhamose metabolismrnc = encodes RnaseIII (rnc-14 is a common null mutant)rne = encodes RnaseE (rne-3071 is a common temperature sensitive mutant)rpsL = mutation in ribosomal protein S12 conveying streptomycin resistance; also called strA sbcBC = ExoI activity abolished; usually present in recBC strains; recombination proficient, stable inverted repeatssr1 = cannot metabolize sorbitolsupE = glnVsupF = tyrTthi = requires thiaminethyA = requires thymidineTn10 = transposon normally carrying Tetracycline resistanceTn5 = transposon normally carrying Kanamycin resistancetonA = Mutation in outer membrane protein conveying resistance to phage T1 and phage T5 traD = Mutation eliminating transfer factor; prevents transfer of F plasmidtrxB = mutation in thioredoxin reductase; enhances disulphide bond formation in the cytoplasm tsx = outer membrane protein mutation conveying resistance to phage T6 and colicin KtyrT = suppression of amber (UAG) stop codons by insertion of tyrosine; needed for some phage infection such as λgt11. ung1 = allows uracil to exist in plasmid DNAxyl-5 = blocked xylose metabolismSm R = Streptomycin resistanceMethylation Issues in E. coliType I methylation systems:E. coli K-12 restricts DNA which is not protected by adenine methylation at sitesAA*C[N6]GTGC or GCA*C[N6]GTT, encoded by the hsdRMS genes(EcoKI). Deletions in these genes removes either the restriction or methylation or both of these functions.E. coli B derivative strains contain an hsdRMS system (EcoBI) restricting and protectiing thesequence TGA*[N8]TGCT or AGCA*[N8]TCA.The mcrA gene (carried on the e14 prophage) restricts DNA which is methylated in C m CWGG or m CG sequences (methylation by the dcm gene product).The mcrBC genes restrict R m C sequences.The mrr gene product restricts adenine methylated sequences at CAG or GAC sites.E. coli methylates the adenine in GATC (and the corresponding A on the opposite strand) with thedam gene product.M.EcoKII methylates the first A at the palindromic site ATGCAT (as well as the corresponding A on the opposite strand), see (Kossykh VG (2004) J. Bact 186: 2061-2067 PMID 15028690) Note that this article has been retracted; the retraction appears to center on textual plagarism, notexperimental results. The homology to AvaIII is real. I think I believe it. tk 20:28, 9 December 2005 (EST). Rich Roberts reports: "We have tried ourselves to detect activity with this gene product and cannot detect any methyltransferase activity. In our case we used antibodies able to detect N6-methyladenine or N4 methylcytosine in DNA. The ones we have are very sensitive and should have been able to detect 5 methyl groups in the whole E. coli chromosome. Nothing was detected in an over expressing strain."For additional information see E. coli restriction-modification system and the NEB technicalinformation on methylation.Commonly used strainsAG1endA1 recA1 gyrA96 thi-1 relA1 glnV44 hsdR17(r K- m K+)AB1157thr-1, araC14, leuB6(Am), Δ(gpt-proA)62, lacY1, tsx-33, qsr'-0, glnV44(AS), galK2(Oc), LAM-, Rac-0, hisG4(Oc), rfbC1, mgl-51, rpoS396(Am), rpsL31(strR), kdgK51, xylA5, mtl-1, argE3(Oc), thi-1Bachmann BJ: Derivation and genotypes of some mutant derivatives of Escherichia coli K-12.Escherichia coli and Salmonella typhimurium. Cellular and Molecular Biology (Edited by: F C Neidhardt J L Ingraham KB Low B Magasanik M Schaechter H E Umbarger). Washington, D.C., American Society for Microbiology 1987, 2:1190-1219. See CGSC#1157BL21E. coli B F- dcm ompT hsdS(r B- m B-) gal [malB+]K-12(λS)The "malB region" was transduced in from the K-12 strain W3110 to make the strain Mal+λS. See Studier et al. (2009) J. Mol. Biol. 394(4), 653 for a discussion of the extent of the transfer.Stratagene E. coli Genotype StrainsBL21(AI)F– ompT gal dcm lon hsdS B(r B- m B-) araB::T7RNAP-tetAan E. coli B strain carrying the T7 RNA polymerase gene in the araB locus of the araBAD operon q.Transformed plasmids containing T7 promoter driven expression are repressed until L-arabinose induction of T7 RNA polymerase.Derived from BL21.See the product page for more information.Brian Caliendo (Voigt lab) reported trouble getting the Datsenko and Wanner (2000) plasmid pCP20 to transform into this strain, when other strains transformed fine. Cause is unknown.BL21(DE3)F– ompT gal dcm lon hsdS B(r B- m B-) λ(DE3 [lacI lacUV5-T7 gene 1 ind1 sam7 nin5])an E. coli B strain with DE3, a λ prophage carrying the T7 RNA polymerase gene and lacI qTransformed plasmids containing T7 promoter driven expression are repressed until IPTG induction of T7 RNA polymerase from a lac promoter.Derived from B834 (Wood, 1966) by transducing to Met+.See the original Studier paper or the summary in Methods in Enzymology for more details.Whole genome sequence available [1]BL21 (DE3) pLysSF- ompT gal dcm lon hsdS B(r B- m B-) λ(DE3) pLysS(cm R)pLysS plasmid chloramphenicol resistant; grow with chloramphenicol to retain plasmidChloramphenicol resistantThe pLysS plasmid encodes T7 phage lysozyme, an inhibitor for T7 polymerase which reduces and almost eliminates expression from transformed T7 promoter containing plasmids when not induced.see Moffatt87 for details of pLysS and pLysE plasmidsBNN93F- tonA21 thi-1 thr-1 leuB6 lacY1 glnV44 rfbC1 fhuA1 mcrB e14-(mcrA-) hsdR(r K-m K+) λ-Some C600 strains are really BNN93BNN97BNN93 (λgt11)A λgt11 lysogen producing phage at 42CBW26434, CGSC Strain # 7658Δ(araD-araB)567, Δ(lacA-lacZ)514(::kan), lacIp-4000(lacI q), λ-, rpoS396(Am)?, rph-1, Δ(rhaD-rhaB)568, hsdR514This information is from a printout sent by the E. coli Genetic Stock Center with the strain.B.L. Wanner strainrph-1 is a 1bp deletion that results in a frameshift over last 15 codons and has a polar effect on pyrE leading to suboptimal pyrimidine levels on minimal medium. (Jensen 1993 J Bact. 175:3401)Δ(araD-araB)567 was formerly called ΔaraBAD AH33 by Datsenko and WannerAm = amber(UAG) mutationReference: Datsenko and Wanner, 2000, PNAS, 97:6640NOTE:This promoter driving the expression of lacI was sequenced in this strain using a primer in mhpR (upstream of lacI) and a primer in the opposite orientation in lacI. The lac promoter was found to be identical to wildtype. Thus, the -35 sequence was GCGCAA not GTGCAA as expected with lacI q.Therefore this strain (or at least the version obtained from the E. coli Genetic Stock Center) does NOT appear to be lacI q. According to Barry Wanner, this is an unexpected result. -Reshma 13:19, 5 May 2005 (EDT)"We have now confirmed that BW25113, BW25141, and BW26434 are all lacI+, and not lacI q. We thank you for alerting us to the error with respect to BW26434. Apparently, the lacI region was restored to wild-type in a predecessor of BW25113." (from Barry Wanner November 18, 2005) The genotype has been corrected at the CGSCC600F- tonA21 thi-1 thr-1 leuB6 lacY1 glnV44 rfbC1 fhuA1 λ-There are strains circulating with both e14+(mcrA+) and e14-(mcrA-)General purpose hostSee CGSC#3004References: Appleyard, R.K. (1954) Genetics 39, 440; Hanahan, D. (1983) J. Mol. Biol. 166, 577.C600 hflA150 (Y1073, BNN102)F- thi-1 thr-1 leuB6 lacY1 tonA21 glnV44 λ- hflA150(chr::Tn10)host for repressing plaques of λgt10 when establishing cDNA librariesReference Young R.A. and Davis, R. (1983) Proc. Natl. Acad. Sci. USA 80, 1194.Tetracycline resistance from the Tn10 insertionCSH50F- λ- ara Δ(lac-pro) rpsL thi fimE::IS1See CGSC#8085References: Miller, J.H. 1972. Expts.in Molec.Genetics, CSH 0:14-0; Blomfeld et al., J.Bact. 173: 5298-5307, 1991.D1210HB101 lacI q lacY+DB3.1F- gyrA462 endA1 glnV44 Δ(sr1-recA) mcrB mrr hsdS20(r B-, m B-) ara14 galK2 lacY1 proA2rpsL20(Sm r) xyl5 Δleu mtl1useful for propagating plasmids containing the ccdB operon.gyrA462 enables ccdB containing plasmid propagationstreptomycin resistantappears to NOT contain lacI (based on a colony PCR) --Austin Che 16:16, 18 June 2007 (EDT)1. Bernard-JMolBiol-1992 pmid=13243242. Miki-JMolBiol-1992 pmid=1316444。
escherichia coli k12名词解释微生物

escherichia coli k12名词解释微生物
Escherichia coli K12(大肠杆菌K12)是一种常用的微生物,属于Escherichia coli(大肠杆菌)的种。
它是一种革兰氏阴性菌,属于肠杆菌科。
由于其相对简单的基因组和易于培养的特性,大肠杆菌K12常被用作基因工程和分子生物学研究的基础工具。
在生物工程领域,大肠杆菌K12是第一个完成全基因组测序的微生物,也是第一个被广泛用于基因克隆和表达的微生物。
它的基因组相对较小,只有约4.6万个碱基对,这使得研究者可以相对容易地分离、克隆和操作其基因。
大肠杆菌K12被广泛用于蛋白质表达、基因敲除、转录分析和其他基因工程技术的研究中。
它的基因组中包含许多用于基因克隆和表达的标记基因,如lacZ、ompA、trp等,这些标记基因可以被用于基因表达的检测和调控。
此外,大肠杆菌K12还被用于生产各种生物制品,如抗生素、酶和其他蛋白质药物。
由于其容易培养和生产效率高的特性,大肠杆菌K12已成为生物工程领域的重要微生物。
以上内容仅供参考,建议查阅关于Escherichia coli K12的专业文献或咨询相关领域的专家以获取更准确和全面的信息。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
. u*wvy{ .3012,|+/ xz~}
:QO@GK ,,z ,5xw q./yr ,6x{u{ ,62}wtw~uyxq ,=ywy -tI@ -|vv -3xy| .2w .2{ .2{l .8{vwyvp /.~|{z /.~}|} /9w|z} 2,wvw 26:w}z 36~y 36wvw 36wv{ 36wv| 36wv} 36wv~ 36wv* 36wwv 4~vx 4~vy 5/y*x 6-wv|w 6<ww~z 6<ww*y 7LS@,IRD 99w ;+8*v ;1w ;1x ;2x >5wt,IRD
3[bTbZb KRX[NLQWXUXP_ LJ[JUXP XWURWN4grrnI>>sss=rbibpb=cmk=cl
2/01Βιβλιοθήκη ?twv~~ ?twv~* ?twv*v
bm[c;@4@9@jd]^efhZ_l n M6-* GP M,9yxx TGQF trs o N GKPDOQDCu
X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X X \ ` g iX
hw}}|
,-*+
1DKLQUMD
dfaDDG; dfaCCC; ZdVKD@C; YS]GA; YS]MAA ; ecaKDD; ^We@@; e`_?; k]SUN@EH>O7 `ac`?@ :; ]SU ob; ]SURkV@D[l;: 8eWe p9 a Ots `^aMs ZdVLK 8pK<^K<9 ; YS] 8n U o GDF; [_V@; LS^F; _[_D; ]SU\]D<[FYW_W@9 ; VU^ 8MNB9 q,lakr O<; n [^^CBC\S_c; 8J;9 ; U^p; ci<^i:; eWe p k 8]SU8ac`?@9 ; dfaC; eZ[; ^feLA@D 5 [l;: 8eWe p9 >O7 8ecSBBE; ac`?@ :; ]SU o b; ]SURkV@D9 QfpTU@EYX>CD `]id 8E@<EA9 >Vfe@; f_Y@; eZ[<@; cW]?@a J shjd rtne qrpbhl 8L qrpbhl9 dfaCCC; ZdVK; eZ[<@; eZc<@; ]Wf@E; ]SUQ@; e`_?A@ Vfe@; f_Y@; eZ[<@; cW]?@>nLS@?D 8O7US^p9 dfaCCC; ZdVK@F; cWU?@; W_V?@; Yic?HE; eZ[<@; cW]?@ dfaCCC; ZdVK@F; cWU?@; W_V?@; Yic?HE; eZ[<@; cW]?@ O<; mG?d>]SURkV@D; k8]SURQ?8ScYD9 \@EH; VW`K; cWU?@; W_V?@; ZdVK@F8cT<^T:9; aZ`?; dfaCCC; n<; eZ[<@; Yic?HE; cW]?@ dfaCCC; dfaDDG; ZdVL= 8cK<^K<9 ; VSaBG; ]SUQ@; Y]_OCC; k 8YS]8fgc@9 CF; eicMDG; Yic?AH; e`_?DB; k 8eZi?DF9 dfaC7 dfaD7 ZdVK7 ^We@7 ]SUQ7 YS]7 ecaK dfaCCC; dfaDDG; ZdVL= 8cK<^K<9 ; cWU?DE; YS]GA; YS]MAA; ^We@@ O<; eca<B@; Z[d<@; cadH@?C 8dec p9; XZf?<; k 8jbc^9 c@; dfaCCC; hi]<F; ^e]<A; ^We@@; cWUB@?@C; ^Uc?@AFA Y [l;:; k 8^Uc@; ZdVKIL7 ^cc9 @?A Y [l;: 8eWe p9 dfaCCC; k 8^UcA8^cc9; cWU?@B; ScS8@C7 ac`?A; ]SUQ@; YS]GA; cadHA?; hi]8D; ^e]8@; ]Wf@E; eZ[8@ O<; cWU?@; ZdVK; K[X p O<; ScS; k 8]SU8ac`?@9 ; cadH 8mG? ]SURkV@D9 dfaC; eZ[; k 8]SU8ac`?@9 >O7 `ecSBBE; ac`?@:; ]SU z b; ]SURkV@Da W_V?@7 dfaC7 dTU@@D7 eZ[7 cadH7 k 8]SU8ac`?@9 9O7 `ecSBBE7 ac`?@:7 ]SU o b7 ]SURkV@Da O<7 W_V?@7 Yic?HE7 eZ[7 ZdVK@F7 dfaCCC7 cW]?@7 k 8]SU8ac`?@9 W_V?@7 Yic?HE7 eZ[7 ZdVK@F7 dfaCCC7 cW]?@7 k 8]SU8ac`?@9 >O7 `ecSBBE; ac`?@:; ]SU o o; ]SURkV@Da O<7 cWU?@; W_V?@7 Yic?HE7 eZ[7 ZdVK@F7 dfaCCC7 cW]?@7 k 8]SU8ac`?@9 cWU?@7 W_V?@7 Yic?HE7 eZ[<@7 ZdVK@F8cT<^T:97 W@C< 8^Uc?<9; dfaCCC7 cW]?@7 k 8]SU8ac`?@9 > O7 `ecSBBE; ac`?@:; ]SU o o; ]SURkV@Da VS^7 VU^7 dfaCCC7 ZdVK@F7 eZ[7 ]Wf7 cadH@7 ]SUQ7 YS]G7 YS]M7 ScS7 e`_?7 eZc7 edh7 k 8]SU8ac`?@9 >O7`ecSBBE; ac`?@:; ]SU o b; ]SURkV@Da O<; ]SUQ@ mp k 8]SU o b8Q9 E 8]SU<B9 ; dfaCCC; YS]GA; YS]MAA; ^Uc?7 cXTB7 ^We@@; ^Uc@@7 ZdVKA O<; ]SUQ@ mp k 8]SU o b8Q9 E 8]SU<B9 ; dfaCCC; YS]GA; YS]MAA; ^Uc?7 cXTB7 ^We@@; ^Uc@@7 ZdVKB dfaCCC7 dfaDDG7 ZdVKD@C7 YS]GA7 YS]MAA7 ^We@@7 ecaKDD7 ]SUQ@ ZdVK7 ^Uc@7 ScSB@BH7 k 8ScS?@A8]Wf9 FEFH7 k]SUPFC7 YS]N7 YS]G7 cadH7 eZ[ ScS7 k 8]SU8ac`?@9 7 cadH7 eZ[ 8mG? ]SURkV@D9 7 k 8dc]8cWU?9 B?E Y [l;: 8eWe p9 >O7 `ecSBBE7 ac`?@:7 ]SU o b7 ]SURkV@Da k 8]SU8ac`?@9 7 cadH7 eZ[7 W_V?@; dTU@@D; ZdVKC; k 8dc]8cWU?9 B?E Y [l;: 8eWe p9 >O7 `ecSBBE7 ac`?@:7 ]SU o b7 ]SURkV@Da W_V?@; ZdVK@F; 8cT8^T:9 ; dfaCCC; eZ[<@; Yic?HE; cW]?@; ]SU; cWUJ@>O7; `ac`?@67 ]SU o bRkV@D; [l;: 8eWe p9 a dfaCCC; ZdVLA?; ScS<@C; ac`?A; ]SUQ@; YS]GA; cadHA?; hi]<D; ^e]<@ dfaCCC; dfaDDG; ZdVK7 ac`7 ]Wf@7 eZ[<@; cadH7 ]SUQ@; e`_?@; cWUB@H?B Y ^[_[8eWe dfaC; ZdVkD; eZ[; k 8]SU8ac`?@9 >O7 `ecSBBE; ac`?@:; ]SU o o; ]SURkV@Da dfaC; ZdVkD; eZ[; k 8]SU8ac`?@9 k 8dc]8cWU?9 B?E Y [l;: 8eWe p9 >O7 `ecSBBE; ac`?@:; ]SU o o; ]SURkV@Da dfaCCC; ZdVLA? 8cK<^K<9 ; cWU?@B; ScS<@C; ac`?A; ]SUQ@; YS]GA; cadHA?; hi]<D; ^e]<@; eZ[<@; ecaKEAC ZdVK@F; dfaCCC; cWU?@; W_V?@; Yic?CE; eZ[; cW]?@; ]SU>O7 `ac`?@:;]SU o b; ]SURkV@D Y [l;: 8eWe p9 a O<; eZf?DB; VSaBG; ^[_?@; ^[_@A; cXT<A; Y]_OCC 8dfaCCC9 ; k 8YS]<fgc@9 CF; eic?@CA; Yic?AH; `^d<A; ^WeAED; `d^<@ 8eeW <@9; O<; k]SUN@EH; dfaC; dfaD; ZdVK; ^We@; ecaK; XZf?A@; ac`A Y [l>>nVLHn O<; k]SUN@EH; ]`_<@??; ScSB@BH; cadH; ZX]?@D? `UZc Y [l;:>nVLHna O<; k]SUN@EH; ]`_<@??; ScSB@BH; cadH; dfaD; ^Uc?; ecaAAA Y [l;:>nVLHn k 8T[`F<SdV9 AH; UiU?@; UiU@A; ZdVKA q-lakr