15 蛋白质合成及转运 PPT课件
第十五章蛋白质的生物合成

第一位
(5ˊ)
U
U
C
A
G
遗传密码字典 第二位
C
A
G
第三位
(3ˊ)
U C A G
U C A G
U C A G
U C A G
6 4 组 密 码 子 中 , AUG 既 是 甲 硫氨酸的密码,又是起始密码; 有三组密码不编码任何氨基酸, 而是多肽链合成的终止密码子: UAG、UAA、UGA。
• 在原核生物和真核生物中,均存在另一 种携带蛋氨酸的tRNA,识别非起动部位 的蛋氨酸密码,AUG。
第十五章蛋白质的生物合成
核糖体的研究历史
1.早在本世纪30年代后期就发现细胞质和细胞核中都有 核酸存在,不过用1924年福尔根发明的染色法只能使细胞核 中的核酸染色。但两种核酸在260nm的吸收非常相似。
第十五章 蛋白质合成及转运
蛋白质的生物合成
中心法则指出,遗传信息的表达最终是合成出 具有特定氨基酸顺序的蛋白质,这种以mRNA上所 携带的遗传信息,到多肽链上所携带的遗传信息的传 递,就好象以一种语言翻译成另一种语言时的情形 相似,所以称以mRNA为模板的蛋白质合成过程为 翻译(translation)。
2.Crick, F.比较了核酸和氨基酸的大小和形状后,认为不可 能在空间上互补,因此预测:(1) 存在一类分子转换器,使信 息从核酸序列转换成氨基酸序列;(2) 这种分子很可能是核酸; (3) 它不论以何种方式进入蛋白质翻译系统的模板,都必须与 模板形成氢键(即配对);(4) 有20种分子转换器,每种氨基 酸一个;(5) 每种氨基酸必定还有一个对应的酶,催化与特定 的分子转换器结合。
2.1941年,细胞学家J.Brachet和T.Caspersor注意到细 胞质中的核酸与蛋白质的合成有密切的关系。
第五节蛋白质合成后的加工及转运(共73张PPT)

〔四〕、叶绿体的蛋白质转运
转运到基质的前体蛋白具有典型的N端序列。转运到叶绿 体内膜和类囊体膜的前体蛋白含有两个N端信号序列,第一个 被切除后,暴露出第二个信号序列,将蛋白导向内膜或 类囊体膜。
叶绿体的蛋白质定向 转运
〔五〕、进入到细胞核的蛋白质的 运转:
1、核孔的结构及作用; 2、核质蛋白上的入核信号;
e、转移通道的开启与关闭
膜上存在一个直径1.5nm的孔道,平时由Bip蛋白封闭。 当新生肽链达70个氨基酸左右的长度时,转移通道开启,信
号肽结合在通道上。合成蛋白通过内质网膜人腔,一旦合成 结束,Bip蛋白又将孔道封闭
转移通道的开启
f、蛋白质进入ER腔
信号肽的切除; 信号肽移到脂双层中,最终被降解;
Blobel因此项发现获1999年诺贝尔生理医学奖。
〔2〕、蛋白质定位的信号:
A、信号序列〔signal sequence〕:存在于蛋白质 一级结构上的线性序列,通常15-60个氨基酸残基, 可以指导新合成的蛋白质发生定向转移。有些信号序 列在完成蛋白质的定向转移后被信号肽酶〔signal peptidase〕切除.
③对所牵引的蛋白质没有特异性要求,非线粒体蛋白连接上此 类信号序列,也会被转运到线粒体。
前体蛋白信号序列特点
3、蛋白质输入线粒体的过程
〔1〕、进入外膜的蛋白:具有N端信号序列,其后还有疏 水性序列作为停止转移序列,然后蛋白质被TOM复合体安 装到外膜上,如线粒体的各类孔蛋白。
〔2〕、进入线粒体基质蛋白质:可以先通过TOM复合体进入 膜间隙,然后通过TIM复合体进入基质。也可以通过线粒体 内、外膜间的接触点,一步进入基质,在接触点上TOM与TIM 协同作用完成蛋白质向基质的输入。
①胞质环〔cytoplasmic ring〕,位于核孔复合体胞质一侧,环上有8 条纤维伸向胞质; ②核质环〔nuclear ring〕,位于核孔复合体பைடு நூலகம்质一侧,上面伸出8条 纤维,纤维端部与端环相连,构成笼子状的结构;
大连理工大学生物化学课件--蛋白质合成与转运

二、蛋白质生物合成过程
• • • • 蛋白质生物合成过程包括三大步骤: ①氨基酸的活化与搬运; ②活化氨基酸在核蛋白体上的缩合; ③多肽链合成后的加工修饰。
核糖体主要存在于粗面ER
核糖体存在的场所 (1)粗面内质网(主要) 一个细菌细胞内约有20000个核糖体 • (2)细胞溶液 • 真核细胞内可达106个 (3)线粒体和叶绿体 • 在未成熟的蟾蜍卵细胞内则高达1012个
(二)肽链延长阶段:
1.进位:与mRNA下一个密码相对应 的氨基酰tRNA进入核蛋白体的受位(A 位),需GTP,Mg2+,和EF参与。 2.成肽:在转肽酶的催化下,将给位 上的tRNA所携带的甲酰蛋氨酰基或肽 酰基转移到受位上的氨基酰tRNA上, 与 其 α- 氨 基 缩 合 形 成 肽 键 。 此 步 骤 需 Mg2+,K+。给位上已失去蛋氨酰基或肽 酰基的tRNA从核蛋白上脱落。
8、供能物质和无机离子
• 多肽链合成时,需ATP、GTP作为供能物质,并需 Mg2+、K+参与。
• 氨基酸活化时需消耗2分子高能磷酸键,肽键形成 时又消耗2分子高能磷酸键,故缩合一分子氨基酸 残基需消耗4分子高能磷酸键。
氨酰- tRNA合成酶
氨基酸 + tRNA + ATP
无机磷酸酶
氨酰- tRNA + AMP + 2Pi
一、参与蛋白质生物合成的物质
• 生物体内的各种蛋白质都是利用生物体内的氨基酸 为原料自行合成的。参与蛋白质生物合成的各种因 素构成了蛋白质合成体系,该体系包括: ① mRNA:作为蛋白质生物合成的模板,决定多肽链 中氨基酸的排列顺序; ② tRNA:搬运氨基酸的工具; ③ 核蛋白体:蛋白体生物合成的场所; ④ 酶及其他蛋白质因子;氨酰-tRNA合成酶; ⑤ 供能物质及无机离子。ATP和GTP, Mg2+、K+ ;
蛋白质的转运与信号传导课件.ppt
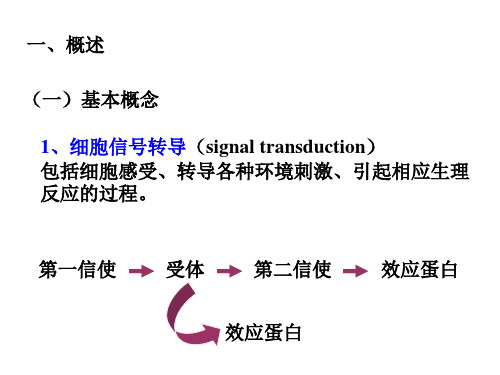
三、反应
同时,植物细胞信号转导系统在某些方面还保留了低等 原核细胞的信号转导机制,例如植物激素乙烯受体ETR1 与细菌双组份信号转导系统之间具有极大的相似性。
二、植物细胞信号转导过程
植物细胞的信号转导过程可以简单概括为: 刺激与感受——信号转导——反应三个重要的环节。
(一)刺激与感受 受体
(二)信号转导
此过程相当复杂,主要包括胞外信号的跨膜转换、细 胞内第二信使系统和信号的级联放大以及蛋白质的可 逆磷酸化。
1、信号跨膜转换
(1)通过离子通道连接受体跨膜转换信号
离子通道(ion channel)是存在于膜上可以跨膜转运 离子的一类蛋白质。
(2)酶促信号直接跨膜转换
该过程的跨膜信号转换主要由酶连受体来完成。
cell
胞外环境信号:指机械刺激、磁场、辐射、温度、风、 光、CO2、O2、土壤性质、重力、病原因子、水分、营养 元素、伤害等影响植物生长发育的重要外界环境因子。
胞间信号:指植物体自身合成的、能从产生之处运到别 处,并对其他细胞作为刺激信号的细胞间通讯分子,通 常包括植物激素、气体信号分子NO以及多肽、糖类、细 胞代谢物、甾体、细胞壁片段等。
3、第二信使(second messenger)
又称次级信使,是指细胞感受胞外环境信号和胞间信 号后产生的胞内信号分子,从而将细胞外信息转换为 细胞内信息。
一般公认的细胞内第二信使有钙离子(Ca2+)、肌 醇三磷酸(inositol 1,4,5-trisphosphate,IP3)、二酰 甘油(1,2-Diacylglycerol,DG)、环腺苷酸 (cAMP)、环鸟苷酸(cGMP)等。
•高特异性 •高的亲合力 •高饱和性 •可逆性 •通过磷酸化(与动物细胞信号传 导的区别)
蛋白质的生物合成及转运

第十二章蛋白质的生物合成及转运蛋白质的生物合成在细胞代谢中占有十分重要的地位。
目前已经完全清楚,贮存遗传信息的DNA并不是蛋白质合成的直接模板,DNA上的遗传信息需要通过转录传递给mRNA。
mRNA才是蛋白质合成的直接模板。
mRNA是由4种核苷酸构成的多核苷酸,而蛋白质是由20种左右的氨基酸构成的多肽,它们之间遗传信息的传递与从一种语言翻译成另一种语言时的情形相似。
所以人们称以mRNA为模板合成蛋白质的过程为翻译或转译(translation)。
翻译的过程十分复杂,几乎涉及到细胞内所有种类的RNA和几十种蛋白质因子。
蛋白质合成的场所是核糖体,合成的原料是氨基酸,反应所需能量由A TP和GTP提供。
蛋白质合成的早期研究工作都是用大肠杆菌的无细胞体系进行的,所以对大肠杆菌的蛋白质合成机理了解最多。
真核细胞蛋白质合成的机理与大肠杆菌的有许多相似之处。
第一节遗传密码任何一种天然多肽都有其特定的严格的氨基酸序列。
有机界拥有1010~1011种不同的蛋白质,构成数目这么庞大的不同的多肽的单体却只有20种氨基酸。
氨基酸在多肽中的不同排列次序是蛋白质多样性的基础。
目前已经清楚,多肽上氨基酸的排列次序最终是由DNA上核苷酸的排列次序决定的,而直接决定多肽上氨基酸次序的却是mRNA。
不论是DNA还是mRNA,基本上都由4种核苷酸构成。
这4种核苷酸如何编制成遗传密码,遗传密码又如何被翻译成20种氨基酸组成的多肽,这就是蛋白质生物合成中的遗传密码的翻译问题。
一、密码单位用数学方法推算,如果mRNA分子中的一种碱基编码一种氨基酸,那么4种碱基只能决定4种氨基酸,而蛋白质分子中的氨基酸有20种,所以显然是不行的。
如果由mRNA 分子中每2个相邻的碱基编码一种氨基酸,也只能编码42=16种氨基酸,仍然不够。
如果采用每3个相邻的碱基为一个氨基酸编码,则43=64,可以满足20种氨基酸编码的需要。
所以这种编码方式的可能性最大。
应用生物化学和遗传学的研究技术,已经充分证明了是293三个碱基编码一个氨基酸。
蛋白质的合成与运输课件

分子生物学研究方法
1 2
基因克隆和表达
通过基因工程技术,将目标基因克隆到表达载体 中,并在宿主细胞中表达,从而获得大量的目标 蛋白质。
基因敲除和敲入
利用基因编辑技术,对特定基因进行敲除或敲入 操作,以研究蛋白质合成和运输的调控机制。
3
转录组学分析
利用高通量测序技术,对细胞或组织中全部基因 的表达水平进行分析,从而了解蛋白质合成的转 录调控机制。
பைடு நூலகம்
05
蛋白质合成与运输的调控
基因表达调控
转录水平调控
通过调节基因转录的起始 和效率,控制蛋白质合成 的数量和种类。
转录因子
转录因子与DNA结合,调 控特定基因的表达。
表观遗传学
DNA甲基化、组蛋白修饰 等表观遗传学机制影响基 因表达。
翻译后修饰调控
磷酸化
磷酸化是蛋白质最常见的翻译后 修饰,通过改变蛋白质活性来调
蛋白质的胞内运
核糖体合成
蛋白质在核糖体上合成后,通过信号肽引 导进入内质网。
内质网加工
蛋白质在内质网中经过折叠、组装和糖基 化等加工。
高尔基体转运
经过内质网加工的蛋白质通过囊泡转运至 高尔基体进一步修饰和分拣。
蛋白质的分泌运
囊泡形成
蛋白质在高尔基体形成囊泡,通过胞吐作用释放到细胞外。
胞吐过程
囊泡与细胞膜融合,释放出内容物,完成蛋白质的分泌运输 。
THANKS
常见的翻译后修饰包括磷酸化、乙酰化、糖基化等,这些修饰可以影响蛋白质的结 构和功能,使其具有更广泛的生物活性。
翻译后修饰对于蛋白质的功能和稳定性具有重要作用,是蛋白质合成过程中的重要 环节。
04
蛋白质的运输
蛋白质的跨膜运
蛋白质合成PPT课件

.
12
6.1.2 核糖体—蛋白质制造厂
1.核糖体结构及种类 • 由大小二亚基组成 • 给位(P位,肽位): 起始时, tRNAimet结合于核糖体的肽位
延长成肽后,肽链转到此位。 • 受位(A位,氨基酰位): 延长成肽时,氨基酰tRNA就加入此位。
.
13
核糖体种类(胞质中)
• 游离的核糖体---合成细胞固有蛋白 • 与粗面内质网结合的核糖体---合成带有信
位
• 30S小亚基与mRNA识
别、结合
• IF1、IF3协助
• fmet-tRNAfmet -IF2-
3.fmet-tRNAfmet的结 合
GTP 通过其反密码与 mRNA上的起始密码
AUG相配对
.
31
SD序列(shine-Dalgarno序列):---原核生物 1.位于起始密码上游25个核苷酸, 2.序列富含嘌呤(如AGGA /GAGG)的一段序 列。 3.能和原核生物16s rRNA相应的富含嘧啶序 列
号肽的分泌性蛋白质
.
14
2. 核糖体的装配
.
15
.
16
• 多核糖体 • 一条mRNA链上同时具有许多个核糖 体
(每隔80核苷酸有一个核糖体)
• 一条mRNA可同时合成多条多肽链
.
17
3 .核糖体的活性位点
(1)mRNA结合位点 在16SrRNA的3`端有一段 顺序同多数原核生物的mRNA(AUG上游3-9个碱 基)的核糖体结合位点有互补关系,以便使 mRNA结合在小亚基上。
起“接合器”作用
mRNA摆动配对
.
6
3. tRNA的种类
• (1)起始tRNA与延伸tRNA
细胞内蛋白质的分选和运输课件

如何实现膜泡在细胞器之间的靶向运输?
小泡如何形成?小泡内的货物如何分 选和装运?小泡如何实现靶向运输?
小泡的靶向运输
Paired sets of SNARE proteins mediate fusion of vesicles with target membranes.
Rab proteins and SNAREs help direct transport vesicles to their target membranes
Inner Membrane Intermembrane space
Outer Membrane
Additional targeting sequences specify other locations
Example: ATP Synthase subunit on inner membrane
Yeast cells expressing mutant Rab protein accumulate secretory vesicles that are unable to fuse with the plasma membrane.
Figure 15-21 Essential Cell Biology (© Garland Science 2010)
non-insulin dependent, normal insulin production,
lack of response to insulin
膜蛋白的上膜也可以是受调的
Hydropathy profiles of integral membrane proteins
All have hydrophobic domains
第五节蛋白质合成后的加工及转运课件

在蛋白质合成过程中,信号识别颗粒与新生蛋白质结合,引导蛋白 质向内质网定位。
作用
确保新生蛋白质正确地转运到内质网,进行进一步的加工和修饰。
跨膜运
跨膜运输
是指蛋白质通过生物膜的运输过 程,包括通过细胞膜、线粒体膜 、叶绿体膜等。
运输方式
包括主动运输和被动运输,其中 主动运输需要消耗能量,而被动 运输则不需要。
多肽链在核糖体上合成的同时,通过 信号肽的引导,进入内质网腔或跨膜 转运至高尔基体、溶酶体和细胞骨架 。
03
膜泡运输
通过形成囊泡的方式,将已经折叠好 的蛋白质从一个膜结构转运到另一个 膜结构。例如,从内质网到高尔基体 ,或从高尔基体到溶酶体。
PART 04
蛋白质合成后的加工和转 运的调节
蛋白质合成后的加工的调节
蛋白质的乙酰化
在蛋白质合成后,某些赖氨酸残基可被乙酰化,从而调节 蛋白质的稳定性。这一过程由乙酰转移酶催化。
蛋白质转运的调节
01
核孔复合体的调节
核孔复合体是细胞核膜上的转运孔道,可选择性地将蛋白质从细胞质转
运到细胞核内或从细胞核转运到细胞质。核孔复合体的转运活性受到多
种因素的调节。
02
囊泡转运的调节
2023-2026
ONE
KEEP VIEW
蛋白质合成后的加工 及转运课件
REPORTING
CATALOGUE
目 录
• 蛋白质合成后的加工 • 蛋白质的转运 • 蛋白质分选的信号和途径 • 蛋白质合成后的加工和转运的调节 • 蛋白质合成后的加工及转运异常与疾病的关系
PART 01
蛋白质合成后的加工
蛋白质二硫键的形成
在蛋白质合成后,某些氨基酸残基需要经过氧化形成二硫 键,以稳定蛋白质的高级结构。这一过程由特异的二硫键 异构酶催化。
第三章蛋白质的转运加工与ppt文档

1. 信号序列(斑块)(signal sequece/patch)
内质网(endoplasmic reticulum, ER)信号序列:存在于所有进 入分泌途径的蛋白质前体中。一般位于肽链 N-terminus ,引导 新生肽链从细胞质进入内质网
基质(matrix)信号序列:引导新合成的蛋白质通过跨膜转运 从细胞质进入细胞器基质
➢ 由两个亚基组成:
β subunit 为膜蛋白,含 300 个氨基酸残基
α subunit 是膜周边蛋白,含 640 个氨基酸残基,负载着 GDP,并且有 GTP 酶活性
➢ SRP receptor 功能: 与 SRP 结合并起始肽链向内质网膜转 运;使肽链的延长继续进行
SRP 与信号序列及 SRP 受体结合
进入内质网腔或膜的蛋白质除了一部分留在内质网 外,其他的形成转运小泡(transport visicle)被运输 到各个细胞器或分泌到细胞外(分泌途径)
分泌途径(secretory pathway)是指通过翻译 同步转运、小泡介导的方式把蛋白质分泌到 细胞外
蛋白质合成及转运优秀课件.ppt

肽链合成起始
指mRNA和起始氨基酰-tRNA分别 与核蛋白体结合而形成翻译起始复合物 (translational initiation complex)。
参与起始过程的蛋白质因子称起始 因子(initiation factor,IF)。
蛋白质合成及转运优秀课件
S-D序列: Shine和Dalgarno
tRNA分子具有4个位点:
• 3’CCA-OH氨基酸接受位点 • 识别氨酰-tRNA合成酶位点 • 核糖体识别位点 • 反密码子位点
蛋白质合成及转运优秀课件
(三)核糖体是蛋白质合成的工厂
核蛋白体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基
大亚基
核蛋白 体
21种 30S 16S 33种 34种 50S 23S 49种
蛋白质合成及转运优秀课件
真核生物翻译起始复合物形成
• 核蛋白体大小亚基分离; • 起始氨基酰-tRNA结合; • mRNA在核蛋白体小亚基就位; • 核蛋白体大亚基结合。
蛋白质合成及转运优秀课件
真核生物翻译起始因子
起始因子
生物功能
eIF-2
促进起始tRNA与小亚基结合
eIF-2B, eIF-3 促进大小亚基分离
——由同一种tRNA合成酶合成
起始因子识别tRNAiMet 延伸因子识别tRNAMet
蛋白质合成及转运优秀课件
原核生物中的甲酰Met
fMet - tRNAiMet
蛋白质合成及转运优秀课件
(五)翻译起始于mRNA与核糖体的结合 原核生物借助SD序列
(六)蛋白因子帮助合成的起始
蛋白质合成及转运优秀课件