第十三章 蛋白质的生物合成共78页文档
合集下载
第十三章蛋白质的生物合成(共91张PPT)

已发现少数例外,如动物细胞的线粒体、植物细胞 的叶绿体等。
密码的通用性进一步证明各种生物进化自同一祖 先。
4. 方向性(direction):
• 指阅读mRNA模板上的三联体密码时, 只能沿5’→3’方向进行。
5. 摆动性(wobble):
转运氨基酸的tRNA的反密码需要通过碱基互补与 mRNA上的遗传密码反平行配对结合,但反密码与 密码之间常常不严格遵守碱基配对规律,称为摆
阶段。
一、多肽链合成的起始阶段
(一)原核生物翻译起始复合物形成
• 包括以下几个步骤:
➢核蛋白体大小亚基分离;
➢mRNA在小亚基定位结合;
➢起始氨基酰-tRNA的结合;
➢核蛋白体大亚基结合。
1. 核蛋白体大、小亚基分离: IF-1和IF-3与小亚基结合,促进核蛋白体大、小亚
基拆离,为新一轮合成作准备。
• 成肽是由转肽酶(transpeptidase)催化的 肽键形成过程。
• 在转肽酶的催化下,将P位上的tRNA所携带 的甲酰蛋氨酰基或肽酰基转移到A位上的氨 基酰tRNA上,与其-氨基缩合形成肽键。
• 此步骤需Mg2+,K+。
成肽反应过程
3. 转位(translocation):
• 延长因子EF-G有转位酶(translocase)活性,可 促进核蛋白体向mRNA的3´侧移动相当于一个密码 的距离,同时使肽酰基tRNA从A位移到P位。
氨基酰tRNA合成酶催化的反应
第一步:活化反应
氨基酸 +ATP-E → 氨基酰-AMP-E + PPi
第二步:连接反应
氨基酰-AMP-E +
tRNA
↓
氨基酰-tRNA +
有高度特 异性。
密码的通用性进一步证明各种生物进化自同一祖 先。
4. 方向性(direction):
• 指阅读mRNA模板上的三联体密码时, 只能沿5’→3’方向进行。
5. 摆动性(wobble):
转运氨基酸的tRNA的反密码需要通过碱基互补与 mRNA上的遗传密码反平行配对结合,但反密码与 密码之间常常不严格遵守碱基配对规律,称为摆
阶段。
一、多肽链合成的起始阶段
(一)原核生物翻译起始复合物形成
• 包括以下几个步骤:
➢核蛋白体大小亚基分离;
➢mRNA在小亚基定位结合;
➢起始氨基酰-tRNA的结合;
➢核蛋白体大亚基结合。
1. 核蛋白体大、小亚基分离: IF-1和IF-3与小亚基结合,促进核蛋白体大、小亚
基拆离,为新一轮合成作准备。
• 成肽是由转肽酶(transpeptidase)催化的 肽键形成过程。
• 在转肽酶的催化下,将P位上的tRNA所携带 的甲酰蛋氨酰基或肽酰基转移到A位上的氨 基酰tRNA上,与其-氨基缩合形成肽键。
• 此步骤需Mg2+,K+。
成肽反应过程
3. 转位(translocation):
• 延长因子EF-G有转位酶(translocase)活性,可 促进核蛋白体向mRNA的3´侧移动相当于一个密码 的距离,同时使肽酰基tRNA从A位移到P位。
氨基酰tRNA合成酶催化的反应
第一步:活化反应
氨基酸 +ATP-E → 氨基酰-AMP-E + PPi
第二步:连接反应
氨基酰-AMP-E +
tRNA
↓
氨基酰-tRNA +
有高度特 异性。
第十三章蛋白质的生物合成

11111111111111111111
三、 转移RNA的功能
天然蛋白质的20种氨基酸都有自己特定的tRNA,而且 一种氨基酸常有数种tRNA,所以tRNA的种类可达数 10种。它们在ATP提供能量和酶的作用下,可分别与特 定的氨基酸结合。实验表明,tRNA必须具备倒L型的 三级结构才具有携带氨基酰的功能。
将微粒体用去污剂处理,可以使核糖体 从内质网上分离出来,离心后即可获得 纯化的核糖体。将核糖体与放射性标记 氨基酸、ATP、Mg2+和大白鼠肝脏的胞 浆上清液部分一起保温,就可以进行肽 链的合成。此后又发现许多其他的细胞, 如网织红细胞、大肠杆菌等均可使氨基 酸掺人核糖体。上述一系列实验结果清 楚地指出:核糖体是细胞内蛋白质合成 的部位。
上述遗传密码虽然是用大肠杆菌为实验材料所得的结果, 但是后来证明生物的遗传密码是通用的,即不论病毒、 原核生物还是真核生物都用同一套密码。但1980年底, 有的实验室报导酵母链孢霉和哺乳动物线粒体的遗传密 码有的不同于标准密码。如在线粒体中UGA不是终止 密码而是色氨酸的密码。AUA是甲硫氨酸的密码,而 不再是异亮氨酸的密码,CUA应是亮氨酸的密码,但是 在线粒体中却是苏氨酸的密码。由此看来,细胞核和亚 细胞的密码子略有不同。这到底是进化结果还是突变的 产物,尚没有统一的认识。
总之,利用酶法或化学法合成有特定序 列的均聚核苷酸,共聚核苷酸及核糖体 结合技术等方法,仅仅用了4年时间,于 1965年完全弄清了20种天然氨基酸的多 组密码子(表13-1)。
遗传密码
64种密码子,有61种是氨基酸的密码。每种氨基酸可 以有1—6种密码。其中AUG不仅是甲硫氨酸的密码, 也是“起始”密码。UAA,UAG,UGA是终止密码, 也就是蛋白质合成的终止信号。
第十三蛋白质的生物合成

(E)
15
3.密码子AUG可能的功能,是 A.启动密码子 B.终止密码子 C.蛋氨酸密码子 D. Val密码子
(A,C)
4. 关于密码子,错误的叙述是
(A)
A. AUG表示蛋白质生物合成的启动
B.密码子AUG代表甲酰蛋氨酸
C.除AUG外,有时GUG是原核生酸
UUC (Phe)
亮氨酸
(Leu)
ACU ACC ACA ACG
苏氨酸
(Thr)
AUG 蛋氨酸/甲酰蛋氨酸
(Met)
12
3 通用性
从病毒、细菌到人 ,所有生物均使用同一套密码子 说明各种生物在进化上具有同源性 通用性是相对的,哺乳动物线粒体除外
线粒体中:UAG不代表终止信号而是代表色氨酸
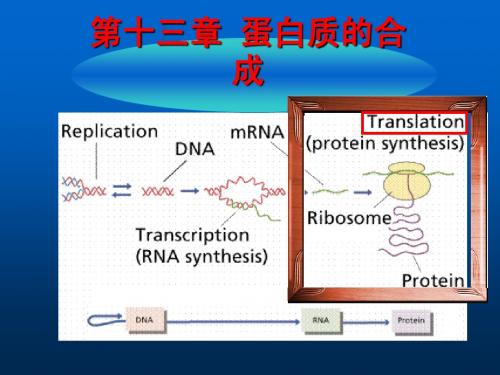
第12章 蛋白质的生物合成
(翻译) Protein Biosynthesis,Translation
1
DNA复制
RNA复制
转录
翻译
DNA
RNA
蛋白质
反转录
2
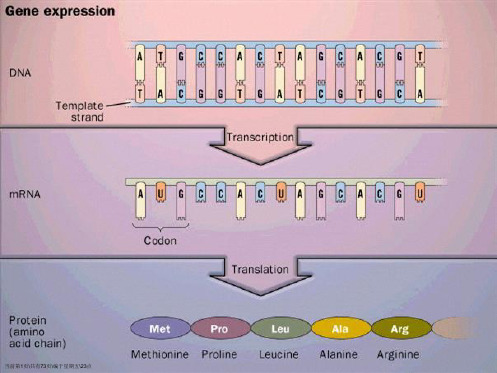
翻译(translation)
蛋白质生物合成即翻译过程,是以mRNA作为 模板、由氨基酸通过肽键结合,形成特定多肽 链的过程。这样,mRNA分子中的遗传信息被 具体地翻译成为蛋白质的氨基酸排列顺序
E. 密码子AUG代表蛋氨酸
16
二 氨基酸的“搬运工具”-tRNA
17
tRNA功能区
氨基酸臂:3’-CCA-OH直接与活化的氨基酸 结合,携带氨基酸
反密码环:识别密码子
1 3
18
tRNA转运特点
tRNA转运氨基酸具有特异性 一种tRNA对应一种氨基酸 一种氨基酸可有多种 tRNA转运
氨基酸结合于tRNA的3’末端(CCA-OH)而活化
第十三章+蛋白质生物合成+翻译

膜
外
分泌蛋白
信号肽酶
4
3
DP2
膜内
N
N 1
SRP
胞浆
信号肽
转运系统
正常:CTT→GAA
Glu
镰刀型贫血:CAT→GUA
Val
复制 、转录、翻译的区别
•
复制
转录
翻译
• 模板 DNA(双链) DNA(模板链) mRNA
•酶
DDDP
DDRP 氨基酰-tRNA合
•
成酶、转肽酶
• 底物 dNTP
NTP
AA
• 产物 子代DNA
RNA
肽链
• 引物 有
无
无
• 特点 半保留复制
不对称转录 核蛋白体循环
•
体外 (影响合成任何因素)
• 一、阻断剂
• 作用:抗菌、治疗肿瘤。
• 种类:
•
抗生素
核蛋白体
•
抗代谢药
竞争性抑制
•
信息传递各环节
•
毒素
信息传递各环节
•
生物活性物
信息传递各环节
二、分子病
定义:由于基因缺陷,使RNA与蛋白
质的合成异常,导致机体某
些结构与功能变异引起的疾病。
e.g :
DNA mRNA Hb链(第6位AA)
•
半不连续复制
模板方向 3 ’ 5 ’ 3’ 5’
5’
3’
产物方向 5’ 3’ 5’ 3’
N
C
• 某些抗生素对蛋白质合成的影响
• 干扰物
作用原理
作用点
• 四环素族 阻碍氨基酰-tRNA
小亚基
•
与小亚基结合;易 (30S、40S )
第十三蛋白质的生物合成
搬运工具—tRNA的反密码子与mRNA的密码子
当前第27页\共有73页\编于星期五\23点
(三) tRNA的功能
tRNA分子上与蛋白质合成有关的位点: 1. 3′端-CCA上的氨基酸接受位点。 2. 识别氨酰-tRNA合成酶的位点。 3. 核糖体识别位点,使延长中的肽链附着于核糖 体上。 4. 反密码子位点。 第一套密码系统(mRNA):决定蛋白质肽链氨基酸排 列顺序。 第二套密码系统(tRNA):决定tRNA与哪种氨基酸结合。
当前第28页\共有73页\编于星期五\23点
第四节 蛋白质生物合成的过程
(一). 氨基酸活化 (二). 肽链起始 (三). 肽链延长 (四). 肽链的终止和释放 (五). 肽链合成后的加工 (六). 蛋白质的分拣(sorting)
与定向输送 (七). 蛋白质生物合成的抑制剂
当前第29页\共有73页\编于星期五\23点
Differences with other tRNAs
细胞内有2种可携带Met的tRNA,它们都识别同样的AUG 密码子,但它们的一级结构和功能不同。
1.tRNAf Met 带上Met后能甲酰化,是起始tRNA,用于肽链合
成的起始。
2.tRNAmMet带上Met后不能甲酰化,用于肽链的内部,在 肽链延伸中起作用。
还需核糖体大小亚基参与,形成一个70S的真正起始 复合物。
当前第44页\共有73页\编于星期五\23点
inactive 70S ribosome
当前第45页\共有73页\编于星期五\23点
SD sequence
30S initiation complex
GDP + Pi
当前第46页\共有73页\编于星期五\23点
核酸代谢与蛋白质生物合成

第1页/共78页
第1节 核酸代谢
一、核苷酸的合成代谢 途径:从头合成、补救合成。
第2页/共78页
1.从头合成途径:是指细胞利用一碳单位、 磷酸核糖和氨基酸等基本原料,经过一系 列复杂的酶促反应,合成嘌呤或嘧啶核苷 酸的过程。
2.补救合成途径:是指细胞利用已有的嘌呤 碱或嘧啶碱以及它们的核苷形式,经过简 单的酶促反应,合成嘌呤或嘧啶核苷酸的 过程。
第21页/共78页
原核生物三种DNA聚合酶及其作用
5´→3´ 聚合作用
3´→5´ 核酸外切酶活
性
5´→3´ 核酸外切酶活
性
功能
DNA聚合酶 DNA聚合酶 DNA聚合酶
Ⅰ
Ⅱ
Ⅲ
有
有
有
有
有
有
有
无
无
校读
目前尚不完
损伤后修复 全清楚
第22页/共78页
复制过程中的 主要催化酶
真核生物DNA聚合酶 : 有α、β、γ、δ和ε等五种。
SSB:维持、保护单 链DNA模板。
引物酶:催化 合成RNA引物。
拓扑异构酶参与 松解DNA超螺旋, 在复制的整个过 程中都发挥作用。
第27页/共78页
延长阶段
第28页/共78页
模板方向:3´→5´ 合成的方向:5´→3´ 碱基配对:
A=T、G ≡ C
前导链与随从链
AG CTTAG CAAG C CATC G GA TC GAA
第3页/共78页
(一)嘌呤核苷酸的合成 1.嘌呤核苷酸的从头合成
嘌呤环的元素来源
第4页/共78页
第5页/共78页
Asp Mg2+ GTP
NAD+ H2O
NADH+
第1节 核酸代谢
一、核苷酸的合成代谢 途径:从头合成、补救合成。
第2页/共78页
1.从头合成途径:是指细胞利用一碳单位、 磷酸核糖和氨基酸等基本原料,经过一系 列复杂的酶促反应,合成嘌呤或嘧啶核苷 酸的过程。
2.补救合成途径:是指细胞利用已有的嘌呤 碱或嘧啶碱以及它们的核苷形式,经过简 单的酶促反应,合成嘌呤或嘧啶核苷酸的 过程。
第21页/共78页
原核生物三种DNA聚合酶及其作用
5´→3´ 聚合作用
3´→5´ 核酸外切酶活
性
5´→3´ 核酸外切酶活
性
功能
DNA聚合酶 DNA聚合酶 DNA聚合酶
Ⅰ
Ⅱ
Ⅲ
有
有
有
有
有
有
有
无
无
校读
目前尚不完
损伤后修复 全清楚
第22页/共78页
复制过程中的 主要催化酶
真核生物DNA聚合酶 : 有α、β、γ、δ和ε等五种。
SSB:维持、保护单 链DNA模板。
引物酶:催化 合成RNA引物。
拓扑异构酶参与 松解DNA超螺旋, 在复制的整个过 程中都发挥作用。
第27页/共78页
延长阶段
第28页/共78页
模板方向:3´→5´ 合成的方向:5´→3´ 碱基配对:
A=T、G ≡ C
前导链与随从链
AG CTTAG CAAG C CATC G GA TC GAA
第3页/共78页
(一)嘌呤核苷酸的合成 1.嘌呤核苷酸的从头合成
嘌呤环的元素来源
第4页/共78页
第5页/共78页
Asp Mg2+ GTP
NAD+ H2O
NADH+
蛋白质的生物合成(共106张PPT)

现 )和真菌中发现UAG可能是编码第22种氨基酸 pyrrolysine(吡咯赖氨酸)的密码子。
遗传密码动画
2022/9/17
12
遗传密码的特点
1.方向性(direction) 翻译时的阅读方向只能是5→3,即读
码从mRNA的起始密码子AUG开始,按 5→3的方向逐一阅读,直至终止密码子。
2022/9/17
为遗传密码(也称密码子)。 mRNA在核糖体小亚基就位;
-Ser-Lys-Leu-(PST序列) 顺反子(cistron):遗传学将编码一个多肽的遗传单位称为顺反子。 氨基酰-tRNA合成酶
• 开放阅读框架(open reading frame,ORF):从 开放阅读框区(open reading frame, ORF)
AMP-E或氨基酰-tRNA的酯键水解,再换上与密码子相对应
的氨基酸。
• 氨基酰-tRNA的表示方法:
• Ala-tRNAAla、Ser-tRNASer、Met-tRNAMet
氨基酸的活化形式:氨基酰-tRNA 氨基酸的活化部位:α-羧基 氨基酸与tRNA连接方式:酯键 氨基酸活化耗能:2个~P
2022/9/17
23
核糖体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基 21种 30S 16S 33种 40S 18S
大亚基 36种 50S 23S 5S
49种
28S 60S 5.8S
5S
核蛋白体
70S
80S
2022/9/17
24
30S小亚基:有mRNA结合位点 50S大亚基: E位:排出位(Exit site)
• 氨基酰-tRNA合成酶:存在于胞液中,催化氨基酸的 活化。
遗传密码动画
2022/9/17
12
遗传密码的特点
1.方向性(direction) 翻译时的阅读方向只能是5→3,即读
码从mRNA的起始密码子AUG开始,按 5→3的方向逐一阅读,直至终止密码子。
2022/9/17
为遗传密码(也称密码子)。 mRNA在核糖体小亚基就位;
-Ser-Lys-Leu-(PST序列) 顺反子(cistron):遗传学将编码一个多肽的遗传单位称为顺反子。 氨基酰-tRNA合成酶
• 开放阅读框架(open reading frame,ORF):从 开放阅读框区(open reading frame, ORF)
AMP-E或氨基酰-tRNA的酯键水解,再换上与密码子相对应
的氨基酸。
• 氨基酰-tRNA的表示方法:
• Ala-tRNAAla、Ser-tRNASer、Met-tRNAMet
氨基酸的活化形式:氨基酰-tRNA 氨基酸的活化部位:α-羧基 氨基酸与tRNA连接方式:酯键 氨基酸活化耗能:2个~P
2022/9/17
23
核糖体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基 21种 30S 16S 33种 40S 18S
大亚基 36种 50S 23S 5S
49种
28S 60S 5.8S
5S
核蛋白体
70S
80S
2022/9/17
24
30S小亚基:有mRNA结合位点 50S大亚基: E位:排出位(Exit site)
• 氨基酰-tRNA合成酶:存在于胞液中,催化氨基酸的 活化。
蛋白质的合成
起始因子 IF-1 原核 生物 EIF-2 EIF-3 eIF-2 eIF-2B,eIF-3 IF-4A 真核 生物 eIF--4B eIF-4E eIF-4G eIF-5 eIF-6 生物功能 占据A位防止结合其他tRNA 促进起始tRNA与小亚基结合 促进大小亚基分离,提高P位对结合起始tRNA敏感性 促进起始tRNA与小亚基结合 最先结合小亚基促进大小亚基分离 eIF-4F复合物成分,有解螺旋酶活性,促进mRNA结 合小亚基 结合mRNA,促进mRNA扫描定位起始AUG eIF-4F复合物成分,结合mRNA5'帽子 eIF-4F复合物成分,结合eIF-4E和PAB 促进各种起始因子从小亚基解离,进而结合大亚基 促进核蛋白体分离成大小亚基
e IF-4A
真核 生物 eIF--4B eIF-4E eIF-4G eIF-5 eIF-6
(一)原核生物翻译起始复合物形成
1. 核蛋白体大小亚基分离; 2. mRNA在小亚基定位结合; 3. 起始氨基酰-tRNA的结合;
4. 核蛋白体大亚基结合。
1. 核蛋白体大小亚基分离
IF-1 IF-3
一个碱基突变
二、tRNA :
在氨基酸tRNA合成酶催化下,特定的tRNA可与相应的 氨基酸结合,生成氨基酸tRNA,从而携带氨基酸参与蛋 白质的生物合成。 tRNA反密码环中部的三个核苷酸构成 三联体,可以识别mRNA上相应的密码,此三联体就称 为反密码(anticoden)。 反密码对密码的识别,通常也是根据碱基互补原则,即 A—U,G—C配对。但反密码的第一个核苷酸与第三核苷 酸之间的配对,并不严格遵循碱基互补原则。如反密码第 一个核苷酸为Ⅰ,则可与A、U或C配对,如为U,则可与 A或G配对,这种配对称为不稳定配对。(P187表)
e IF-4A
真核 生物 eIF--4B eIF-4E eIF-4G eIF-5 eIF-6
(一)原核生物翻译起始复合物形成
1. 核蛋白体大小亚基分离; 2. mRNA在小亚基定位结合; 3. 起始氨基酰-tRNA的结合;
4. 核蛋白体大亚基结合。
1. 核蛋白体大小亚基分离
IF-1 IF-3
一个碱基突变
二、tRNA :
在氨基酸tRNA合成酶催化下,特定的tRNA可与相应的 氨基酸结合,生成氨基酸tRNA,从而携带氨基酸参与蛋 白质的生物合成。 tRNA反密码环中部的三个核苷酸构成 三联体,可以识别mRNA上相应的密码,此三联体就称 为反密码(anticoden)。 反密码对密码的识别,通常也是根据碱基互补原则,即 A—U,G—C配对。但反密码的第一个核苷酸与第三核苷 酸之间的配对,并不严格遵循碱基互补原则。如反密码第 一个核苷酸为Ⅰ,则可与A、U或C配对,如为U,则可与 A或G配对,这种配对称为不稳定配对。(P187表)
第十三章 蛋白质的生物合成

二、翻译起始(原核)
1.翻译的起始—原核生物
(1)fMet - tRNAfMet 的形成 大肠杆菌蛋白质合成的第一个氨基酸都 是甲酰化的甲硫氨酸,即 N-甲酰甲硫氨酸 (fMet)。 fMet - tRNAfMet 能特异地识别起始密码 子AUG。
(2)翻译起始信号的辨认
mRNA上的起始密码子AUG通常离mRNA5’-末端 约20~30个碱基,在这段前导顺序中,具有一段 特殊顺序AGGAGGU,位于起始密码子AUG之前的固 定位置上,称为S.D序列(Shine-Dalgano顺序)。
四、辅助因子(P343)
在蛋白质合成体系中,还有溶解在胞质中的蛋 白质,在蛋白质合成的不同阶段起作用,分别有:
起始因子(IF):原核生物中有IF1、IF2、IF3
延长因子(EF):原核生物中有EF-Tu、EF-Ts、EF-G 终止因子(RF):原核生物中有RF1、RF2、RF3 其它因子:ATP、GTP、Mg2+
核蛋白体的大、小亚基分别有不同的功能: 1.小亚基:可与mRNA、GTP和起动tRNA结合。 2.大亚基: (1)具有两个不同的tRNA结合点。A位(右)— 受位或氨酰 (2)具有转肽酶活性:将给位上的肽酰基转移给受位上的氨基 (3)具有GTPase活性,水解GTP,获得能量。 (4)具有起始因子、延伸因子及释放因子的结合部位。
。所以说遗传密码基本通用,但非绝对通用。
(5)起始密码子和终止密码子
在64个密码子中,有3个密码子不编码任何氨基酸, 从而成为肽链合成的终止信号,称为终止密码子或无 义密码子,它们是UAA、UAG、UGA。其余的61个密码子均
编码不同的氨基酸,其中AUG既是Met的密码子,又是肽链 合成的起始信号,称为起始密码子。
东北师范大学生物化学第十三章蛋白质的生物合成

第十三章 蛋白质的生物合成
遗传信息的传递
一 蛋白质的合成体系
蛋白质合成的原料是氨基酸,其合成过程非常复杂。
真核生物细胞合成蛋白质需要70多种核糖体蛋白,20 多种 活化氨基酸的酶,10多种辅酶和其他蛋白质因子参加,同时 还要100多种附加的酶类参与蛋白质合成后的修饰、40种 tRNA、4种rRNA,总计300多种不同的大分子参与多肽的 合成。
反密码子
氨酰tRNA合成酶
该酶具有很高的专一性 不仅对其活化的氨基酸专一,而且对 一定的tRNA专一(第二套遗传密码) 校正作用: 水解非正确组合的氨基酸和tRNA之间的共价连 接,对于翻译的正确性是十分重要的
(二)蛋白质的生物合成过程(原核生物) 1 肽链合成的起始
2 肽链的延伸
3 肽链的终止和释放
密码子与反密码子结合时具有一定摆动性
摆动性(变偶性)
密码子的第三位碱基配对不严 格 摆动性的存在合理的解释了密 码子的兼并性,基因突变的危 害降到最低
U A G U C A U C
G
I
3 氨酰tRNA合成酶结合位点
4 核糖体识别位点
二 蛋白质生物合成的分子机制
(一)氨基酸的活化
氨酰tRNA合成酶 消耗2摩尔的ATP 氨基酸的活化是羧基端的活化
白喉毒素: 真核细胞蛋白质合 成的抑制剂
抑制EF2的活性
青霉素
青霉素是本世纪20年代未的一项划时代的科学发现 1928年,英国的细菌学家亚历山大· 弗莱明(英国圣 玛丽学院)发现,发表在1929年英国皇家《实验病 理季刊》 1938年,英国牛津大学病理学家弗洛里和德国生物化 学家钱恩从期刊的资料中找到了有关青霉素的文献 1940年,分离纯化成功青霉素进入临床试验阶段
遗传信息的传递
一 蛋白质的合成体系
蛋白质合成的原料是氨基酸,其合成过程非常复杂。
真核生物细胞合成蛋白质需要70多种核糖体蛋白,20 多种 活化氨基酸的酶,10多种辅酶和其他蛋白质因子参加,同时 还要100多种附加的酶类参与蛋白质合成后的修饰、40种 tRNA、4种rRNA,总计300多种不同的大分子参与多肽的 合成。
反密码子
氨酰tRNA合成酶
该酶具有很高的专一性 不仅对其活化的氨基酸专一,而且对 一定的tRNA专一(第二套遗传密码) 校正作用: 水解非正确组合的氨基酸和tRNA之间的共价连 接,对于翻译的正确性是十分重要的
(二)蛋白质的生物合成过程(原核生物) 1 肽链合成的起始
2 肽链的延伸
3 肽链的终止和释放
密码子与反密码子结合时具有一定摆动性
摆动性(变偶性)
密码子的第三位碱基配对不严 格 摆动性的存在合理的解释了密 码子的兼并性,基因突变的危 害降到最低
U A G U C A U C
G
I
3 氨酰tRNA合成酶结合位点
4 核糖体识别位点
二 蛋白质生物合成的分子机制
(一)氨基酸的活化
氨酰tRNA合成酶 消耗2摩尔的ATP 氨基酸的活化是羧基端的活化
白喉毒素: 真核细胞蛋白质合 成的抑制剂
抑制EF2的活性
青霉素
青霉素是本世纪20年代未的一项划时代的科学发现 1928年,英国的细菌学家亚历山大· 弗莱明(英国圣 玛丽学院)发现,发表在1929年英国皇家《实验病 理季刊》 1938年,英国牛津大学病理学家弗洛里和德国生物化 学家钱恩从期刊的资料中找到了有关青霉素的文献 1940年,分离纯化成功青霉素进入临床试验阶段
基础生物化学 第十三章-蛋白质的分解与合成

(1)再合成氨基酸1
COO(CH2)2 C=O COO+NH3+
NAD(P)H+ H+ NAD(P)H+
COO(CH2)2 CH NH3 COO+
+H2O
谷氨酸脱氢酶
α-酮戊二酸
谷氨酸
(1)再合成氨基酸2
上述反应是多数有机体直接利用NH3 合成谷氨酸的主要途径,而且该反应在 其它所有氨基酸合成中都有意义。因为 谷氨酸的氨基可以转到任何一种α-酮 酸上,从而形成各种相应的氨基酸。
2.转氨基作用2(谷丙转氨酶1)
谷丙转氨酶(GPT)是催化谷氨酸
与丙酮酸之间转氨作用的酶,以肝脏
中活力最大。当肝细胞损伤时,酶就
释放到血液内,于是血液内酶的活力
明显地增加,以此来推断肝功能的正
常与否,
2.转氨基作用2(谷丙转氨酶2)
COO(CH2)2 CH NH3+ COO-
CH3
C=O COO-
当体内不需要将α-酮酸再合成氨基酸,并且
体内的能量供给又极充足时,α-酮酸可以转 变为糖及脂肪。
动物实验已证明,如用氨基酸饲养患人工糖
尿病的犬,大多数氨基酸可使尿中葡萄糖含量 增加,少数几种可使葡萄糖及酮体的含量同时 增加。而亮氨基酸只能使酮体的含量增加。
(3)转变成糖及脂肪2
生糖氨基酸:在体内可以转变成糖的氨基酸
糖、脂肪、氨基酸、与核酸代谢的联系(南大P475)
糖(糖原、葡萄糖) 丝氨酸 色、甘、丙、苏、 半胱、丝、羟脯 磷酸丙糖 丙酮酸
CO2
磷酸甘油 脂肪酸
脂肪
乙酰COA 柠檬酸
CO2
天门冬 酰氨
天门冬 氨酸 苯丙、酪
草酰乙酸
第十三章--蛋白质的生物合成ppt课件

真核细胞mRNA的结构特点
m7G-5´ppp-N-3 ´ p
顺反子
5´ “帽子”
Poly(A)尾巴的功能 ① 是mRNA由细胞核进入细胞质
所必需的形式 ② 它大大提高了mRNA在细胞质
中的稳定性
PolyA 3´
帽子结构功能
① 使mRNA免遭核酸酶的破坏 ② 使mRNA能与核糖体小亚基结合并开始
合成蛋白质 ③ 被蛋白质合成的起始因子所识别,从
1、肽链末端的修饰:
大肠杆菌合成后脱甲酰化,N-端70%的Met 被切除,真核生物Met全部被切除。
2、信号序列的切除 3、二硫键的形成 4、部分肽段的切除 5、个别氨基酸的修饰 6、糖基侧链的添加 7、辅基的加入
实例:胰岛素原的加工
Tu\Ts循环
多多核糖体与核糖体循环
核糖体循环
多核糖体
3ˊmRNA 5ˊ
而促进蛋白质的合成。
AAAAAAA-OH
t RNA
tRNA(transfer ribonucleic asid)在蛋白质合成中处于关键地位,它 不但为每个三联体密码子译成氨基酸提供接合体,还为准确无误地将所需 氨基酸运送到核糖体上提供运送载体。
1、tRNA的结构特征——三叶草型二级结构 2、tRNA的功能 (1)被特定的氨酰- tRNA合成酶识别,使 tRNA接受正确的活化氨基酸。
rRNA和多种蛋白因子参与。在此过程中mRNA为合
成的模板,tRNA为运输氨基酸工具,rRNA和蛋白
质构成核糖体,是合成蛋白质的场所,蛋白质合成
的方向为N—C端。
思考
遗传信息传递的 中心法则
生物的遗传信息以密码的形式储 存在DNA分子上,表现为特定的核苷 酸排列顺序。在细胞分裂的过程中, 通过DNA复制把亲代细胞所含的遗传 信息忠实地传递给两个子代细胞。在 子代细胞的生长发育过程中,这些遗 传 信 息 通 过 转 录 传 递 给 RNA, 再 由 RNA通过翻译转变成相应的蛋白质多 肽链上的氨基酸排列顺序,由蛋白质 执行各种各样的生物学功能,使后代 表现出与亲代相似的遗传特征。后来 人们又发现,在宿主细胞中一些RNA 病毒能以自己的RNA为模板复制出新 的病毒RNA,还有一些RNA病毒能以 其RNA为模板合成DNA,称为逆转录 这是中心法则的补充。
十三蛋白质的生物合成

编辑ppt
转位(移位)
转位酶
5′→3′
5′
AUG
3′
AUG
EFG,GTP
A
OH 21
21
OH
AUG
转位
AUG
进位
3 12
AUG
N→C延
成肽
编辑2p1pt
3
3
21
成肽
四、肽链合成终止
• 需终止因子RF、RR和IF3参与。终止信号 出现,释放因子(release factor,RF, RR) 与其结合。RF有三种RF1,RF2,RF3
编辑ppt
密码子的特点
(1)连续性:两个密码子之间无任何核苷酸加以 隔开和重叠,如插入/删除碱基,可发生移 码突变或框移
5′…. UACGGACAUCUG….3′ 酪甘 组 蛋
插入 缺失
5′….UACCGGACAUCUG….3′ 酪 精 苏 半胱
5′…. UACGACAUCUG….3′ 酪 天 异亮 编辑ppt
编辑ppt
一、氨基酸的活化与转运
氨基酸活化→活化氨基酸的搬运→活化氨 基酸与核蛋白体结合 1.参与活化转运的酶
氨基酰-tRNA合成酶:特异性强,催化特定 的氨基酸与特异的tRNA结合,每种氨基酸 有特异的合成酶催化,此种特异性保证了 遗传信息准确翻译
编辑ppt
氨基酰tRNA的生成--- 氨基酸的活化
编辑ppt
密码子的特点
(四)摆动性:一种氨基酸可有多个密码子
反密码子与mRNA的第三个核苷酸配对时,不 严格遵从碱基配对原则,可出现U-G,I-C,I-A,此种 配对为不稳定配对,又称摇摆性。一般前两个碱 基决定其专一性,第三位碱基可有变异
mRNA5′ ACG
转位(移位)
转位酶
5′→3′
5′
AUG
3′
AUG
EFG,GTP
A
OH 21
21
OH
AUG
转位
AUG
进位
3 12
AUG
N→C延
成肽
编辑2p1pt
3
3
21
成肽
四、肽链合成终止
• 需终止因子RF、RR和IF3参与。终止信号 出现,释放因子(release factor,RF, RR) 与其结合。RF有三种RF1,RF2,RF3
编辑ppt
密码子的特点
(1)连续性:两个密码子之间无任何核苷酸加以 隔开和重叠,如插入/删除碱基,可发生移 码突变或框移
5′…. UACGGACAUCUG….3′ 酪甘 组 蛋
插入 缺失
5′….UACCGGACAUCUG….3′ 酪 精 苏 半胱
5′…. UACGACAUCUG….3′ 酪 天 异亮 编辑ppt
编辑ppt
一、氨基酸的活化与转运
氨基酸活化→活化氨基酸的搬运→活化氨 基酸与核蛋白体结合 1.参与活化转运的酶
氨基酰-tRNA合成酶:特异性强,催化特定 的氨基酸与特异的tRNA结合,每种氨基酸 有特异的合成酶催化,此种特异性保证了 遗传信息准确翻译
编辑ppt
氨基酰tRNA的生成--- 氨基酸的活化
编辑ppt
密码子的特点
(四)摆动性:一种氨基酸可有多个密码子
反密码子与mRNA的第三个核苷酸配对时,不 严格遵从碱基配对原则,可出现U-G,I-C,I-A,此种 配对为不稳定配对,又称摇摆性。一般前两个碱 基决定其专一性,第三位碱基可有变异
mRNA5′ ACG
《蛋白质合成》PPT课件

5
5′-末端 碱基
U C
A
G
中间 碱 基
U
苯丙氨酸 苯丙氨酸
亮氨酸 亮氨酸 亮氨酸 亮氨酸 亮氨酸 亮氨酸
异亮氨酸 异亮氨酸 异亮氨酸 甲硫氨酸、起始 甲酰甲硫氨酸
缬氨酸 缬氨酸 缬氨酸 缬氨酸
C
A
丝氨酸 丝氨酸 丝氨酸 丝氨酸
脯氨酸 脯氨酸 脯氨酸 脯氨酸
苏氨酸 苏氨酸 苏氨酸 苏氨酸
丙氨酸 丙氨酸 丙氨酸 丙氨酸
T2
GTP和 Mg2+
肽链的延长分四步进行:
1、进位
2、转肽
3、脱落
4、移位
精选课件ppt
24
1、进位
fMet-
下一个氨酰- tRNA结合
到70s核糖体的A位点上。
A A
~~~~~U~AC~~~~~~~
AA
新进入的氨酰- tRNA上的
AUG
反密码子必须与A位点上 mRNA的密码子互补。
AA Tu-GTP
专一性 主要决 定于第 一、二 碱基。
精选课件ppt
9
5)通用性
近于完全通用。
哺乳动物线粒体中,UGA不是终止密码子,而是色氨 酸的密码;AUA是甲硫氨酸的密码,而不是异亮氨酸 的密码,CUA是亮氨酸的密码,线粒体中却是苏氨酸 的密码。
精选课件ppt
10
(二)核糖体
核糖体是细胞质中球状颗粒,游离存在或结合在 内质网上。 1、核糖体是蛋白质合成的部位 2、核糖体的组成
这一过程需要延长因子Tu、
OOO
Ts参与,消耗一分子GTP
Pi OOO
Tu -GTP
fMet-
Tu -GDP AA
Ts GDP
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2.人工合成多核苷酸和
无细胞体系蛋白质合成
1961年,M.W.Nirenberg等人,大肠杆菌的无 细胞体系中外加poly(U)模板、20种标记的 氨基酸,经保温后得到了多聚phe-phe-phe, 于是推测UUU编码phe。利用同样的方法得 到 CCC 编 码 pro , GGG 编 码 gly , AAA 编 码 lys。
如果利用poly(UC),则得到多聚Ser-LeuSer-Leu,推测UCU编码Ser,CUC编码Leu.
到1965年就全部破译了43 = 64组密码子。
M.W.Nirenberg等,1968年获诺贝尔生理医学 奖.
第一位
(5ˊ
遗传密码字典
第二位
C
A
G
第三位
(3ˊ)
U C A G
U C A G
二、t RNA
tRNA在蛋白质合成中处于关键地位,它不 但为每个三联体密码子译成氨基酸提供接合体, 还为准确无误地将所需氨基酸运送到核糖体上 提供运送载体。
中心法则总结了生物体内遗传信息的流动规律,揭示遗传的分 子基础,不仅使人们对细胞的生长、发育、遗传、变异等生 命现象有了更深刻的认识,而且以这方面的理论和技术为基 础发展了基因工程,给人类的生产和生活带来了深刻的革命 。
遗 DNA
传
信
息
流
mRNA
动
示
核糖体
意
图
tRNA
第一节 RNA在蛋白质生物 合成中的重要功能
mRNA
mRNA(messenger RNA)是蛋白质生物合成过程 中直接指令氨基酸掺入的模板,是遗传信息的载体。
原核生物和真核生物mRNA的比较
原核细胞mRNA的结构特点
SD区
顺反子
5´
AGGAGGU
顺反子
顺反子
3´
先导区
插入顺序
插入顺序
末端顺序
帽子结构功能
① 半衰期短 ② 许多原核生物mRNA以多顺反子形式存在 ③ AUG作为起始密码;AUG上游7~12个核苷酸处有一被称为SD序列
tRNA分子上三个特定的碱基组成一个反 密码子,位于反密码子环上。
密码子与反密码子的 阅读方向均为5‘ 3’, 两者反向平行配对。3’ 5’
5’
CCG
I
GGC
CCA-OH 3’
4、密码子是近于完全通用的。
这是如火如荼的转基因的前提。 目前只发现线粒体和叶绿体内有列外情况。
64组密码子中,AUG既是编码蛋氨酸的 密码,又是起始密码;有三组密码不编 码任何氨基酸,而是多肽链合成的终止 密码子:UAG、UAA、UGA。
B、可以使DNA上的碱基组成有较达的变化余 地,而仍然保持多肽上氨基酸序列不变(意 思基本同上)。
3、密码的摆动性:
密码子中第三位碱基与反密码子第一位 碱基的配对有时不一定完全遵循A-U、 G-C的原则,Crick把这种情况称为摇 摆性,有人也称摆动配对或不稳定配对。
反密码子第一位的G可以与密码子第三位 的C、U配对,U可以与A、G配对,I可 以和密码子的U、C、A配对,这使得 该类反密码子的阅读能力更强。
生物的遗传信息以密码的形式储存在DNA分子上,表现为特定 的核苷酸排列顺序。在细胞分裂的过程中,通过DNA复制把 亲代细胞所含的遗传信息忠实地传递给两个子代细胞。在子 代细胞的生长发育过程中,这些遗传信息通过转录传递给 RNA,再由RNA通过翻译转变成相应的蛋白质多肽链上的氨 基酸排列顺序,由蛋白质执行各种各样的生物学功能,使后 代表现出与亲代相似的遗传特征。后来人们又发现,在宿主 细胞中一些RNA病毒能以自己的RNA为模板复制出新的病毒 RNA,还有一些RNA病毒能以其RNA为模板合成DNA,称 为逆转录这是中心法则的补充。
翻译过程十分复杂,需要mRNA、 tRNA、rRNA和多种蛋白因子参 与。在此过程中mRNA为合成的 模板,tRNA为运输氨基酸工具, rRNA 和 蛋 白 质 构 成 核 糖 体 , 是 合成蛋白质的场所,蛋白质合成 的方向为N—C端。
遗传信息传递的中心法则
复制
DNA
转录
逆转录
RNA 复制
翻译
蛋白质
的保守区, 16S rRNA3’- 端反向互补而使mRNA与核糖体结合。
真核细胞mRNA的结构特点
m7G-5´ppp-N-3 ´ p
顺反子
Poly(A)尾巴的功能 ① 是mRNA由细胞核进入细胞质所
必需的形式 ② 它大大提高了mRNA在细胞质中
的稳定性
5´ “帽子”
PolyA 3´
帽子结构功能
① 使mRNA免遭核酸酶的破坏 ② 使mRNA能与核糖体小亚基结合并开始
5‘
3‘
起始密
码子
如果在阅读框中插入或删除一个碱基 就会使其后的读码发生移位性错误 (称为移码)。
2.密码的简并性
由一种以上密码子编码同一个氨基酸的现象称 为简并,对应于同一氨基酸的一组密码子称 为同义密码子(Synonymous codon)。
简并性的生物学意义?
A、可以降低由于遗传密码突变造成的灾难性 后果
一、mRNA和遗传密码
原核生物mRNA的结构:4个特点
mRNA的半衰期很短,很不稳定,一旦完成其使 命后很快就被水解掉。 mRNA的分子大小差 异大。mRNA的合成是在DNA指导下的RNA 聚合酶的催化下先合成mRNA的前体,即核内 不均一RNA(hnRNA)。 hnRNA在细胞核 内加工成mRNA,然后转移到细胞质中。
合成蛋白质 ③ 被蛋白质合成的起始因子所识别,从
而促进蛋白质的合成。
AAAAAAA-OH
(一)遗传密码的破译
1.三联体密码与阅读框 遗传密码: DNA(或mRNA)中的核苷
酸序列与蛋白质中氨基酸序列之间的对 应关系称为遗传密码。
密码子(codon):mRNA上每3个相邻 的核苷酸编码蛋白质多肽链中的一个氨 基酸,这三个核苷酸就称为一个密码子 或三联体密码。
U C A G
U C A G
3.三核苷酸诱导安酰tRNA对核糖体的特异结合
技术要点:人工合成三核苷酸为模板+核糖体+AA-tRNA 保温
硝酸纤维滤膜过滤
分析留在滤膜上的核糖体-AAtRNA
确定与核糖体结合的AA
(二)遗传密码的性质
1、密码是无标点符号的且相邻密码子互不重叠。 编码一个肽链的所有密码子是一个接着一个地 线形排列,密码子之间既不重叠也不间隔,从 起始密码子到终止密码子(不包括终止子)构 成一个完整的读码框架,又称开放阅读框架 (ORF)。方向:从mRNA的5’到3’。