蛋白质分选的基本途径与类型ppt
合集下载
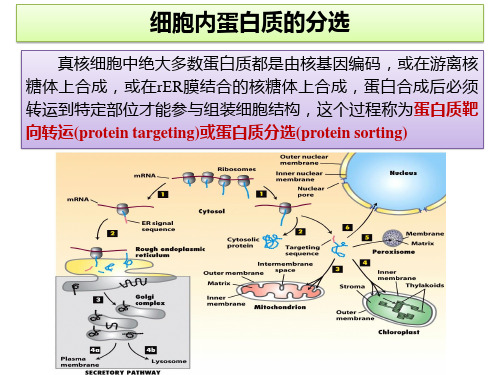
蛋白质的分选课件
• SRP受体(SPR receptor):又称停泊蛋白(docking protein,DP),72KD,异二聚体蛋白,属内质网膜整合蛋白, 与SRP特异结合。
• 一个亚基(640个Aa)暴露于细胞质,亲水;
• 另一亚基(300个Aa)嵌入膜内,疏水。
学习交流PPT
9
• 4.信号假说的实验验证及rER蛋白质合成过程确定
23
• 1 跨膜运输 2 门控运输 3 膜泡运输
蛋白质通过跨膜通道进入目的地。如细胞质 中合成的蛋白质通过内质网或线粒体上的转 位 因 子 ( translocator ) , 以 解 折 叠 的 线 性分子进入内质网腔或线粒体。
如核孔可以选择性的主动运输大分子物质 和RNP复合体,并且允许小分子物质自由进 出细胞核。
• • Blobel、Dobberstein和Walter用分离微粒体和非细胞体
系(cell-free system)实验证实:信号序列的存在,和分 泌蛋白合成过程。
• 实验方法:放射性氨基酸脉冲标记孵育、密度梯度离心获 得微粒体、去垢剂处理对照、蛋白酶降解等。
• RER合成蛋白与信号肽、SRP与DP间的关系实验
• COPI包被的构成:含7种蛋白亚基和1种ARF蛋白。
• 其中,ARF蛋白:GTP结合蛋白,类似Sar1p,调控COPI 包被蛋白复合物和网格蛋白有被小泡组装与去组装。
学习交流PPT
28
COP I 衣被小泡
学习交流PPT
29
• 3.网格蛋白有被小泡介导多条转运途径:
• • 高尔基体TGN(网格蛋白有被小泡发源地)→胞内体、质膜、
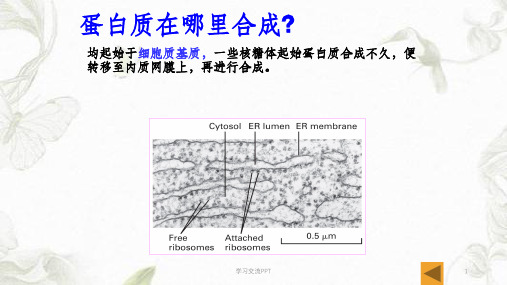
蛋白质在哪里合成?
均起始于细胞质基质,一些核糖体起始蛋白质合成不久,便 转移至内质网膜上,再进行合成。
2023年高中生物竞赛课件:细胞内蛋白质的分选
ER单次跨膜蛋白的合成
新生肽链跨膜取向
新生跨膜蛋白的肽链中没 有N端信号肽,只有内部 信号序列(internal signal sequence)。内部信号序列 的转位方向主要取决于其 侧翼氨基酸残基电荷的分 布。一般而言,带正电荷 的氨基酸残基一侧朝向细 胞质基质一侧
一、信号假说与蛋白质分选信号
Ⅳ型:G蛋白偶联受体、葡 萄糖转运蛋白、电压门 Ca2+通道、CFTR(Cl-)通道
图6-4 内质网膜整合蛋白的拓扑学类型
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
ER单次跨膜 新生跨膜蛋白肽链中既有N端信号肽,又 蛋白的合成 有停止转移信号
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
1、分泌性蛋白在游离核糖体上起始合成 2、多肽链延伸至80个氨基酸残基时,N端内质网信号序列暴露,与 SRP结合,肽链延伸暂停,防止新生肽链N端损伤和成熟前折叠
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
3、SRP与ER膜上SRP受体结合,核 糖体新生肽复合物附着到ER膜,两 分子GTP分别与SRP-p54亚基和SRP 受体α亚基结合,相互作用加强
5、腔面信号肽酶切除信号肽并使之快速降解 6-8、肽链继续延伸,并在ER腔内折叠,移位子关闭
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
➢ 开始转移序列(start transfer sequence) 作为内质网的信号序列,指导新合成的多肽向内
新生肽链跨膜取向
新生跨膜蛋白的肽链中没 有N端信号肽,只有内部 信号序列(internal signal sequence)。内部信号序列 的转位方向主要取决于其 侧翼氨基酸残基电荷的分 布。一般而言,带正电荷 的氨基酸残基一侧朝向细 胞质基质一侧
一、信号假说与蛋白质分选信号
Ⅳ型:G蛋白偶联受体、葡 萄糖转运蛋白、电压门 Ca2+通道、CFTR(Cl-)通道
图6-4 内质网膜整合蛋白的拓扑学类型
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
ER单次跨膜 新生跨膜蛋白肽链中既有N端信号肽,又 蛋白的合成 有停止转移信号
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
1、分泌性蛋白在游离核糖体上起始合成 2、多肽链延伸至80个氨基酸残基时,N端内质网信号序列暴露,与 SRP结合,肽链延伸暂停,防止新生肽链N端损伤和成熟前折叠
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
3、SRP与ER膜上SRP受体结合,核 糖体新生肽复合物附着到ER膜,两 分子GTP分别与SRP-p54亚基和SRP 受体α亚基结合,相互作用加强
5、腔面信号肽酶切除信号肽并使之快速降解 6-8、肽链继续延伸,并在ER腔内折叠,移位子关闭
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
➢ 开始转移序列(start transfer sequence) 作为内质网的信号序列,指导新合成的多肽向内
蛋白质分选的基本途径与类型

离子通道和转运蛋白的调节
改变离子通道和转运蛋白的活性,调节物质进出细胞的速率。
03
蛋白质的囊蛋白质分选的 重要途径之一,通过囊泡将蛋白 质从一处转运到另一处,实现蛋 白质的定位和功能。
02
囊泡运输涉及到多种细胞器之间 的相互联系和蛋白质的跨膜转运 ,对于维持细胞结构和功能具有 重要意义。
囊泡运输的调控机制
信号分子调控
某些信号分子可以与囊泡上的受 体结合,调控囊泡的转运方向和 目的地。
能量依赖性调控
囊泡运输需要消耗能量,如ATP 水解产生的能量,以驱动囊泡的 转运过程。
蛋白激酶与磷酸化
调控
某些蛋白激酶可以调控囊泡运输 相关蛋白的磷酸化状态,从而影 响囊泡的转运过程。
04
蛋白质的膜泡运输
囊泡运输的途径
蛋白质从粗面内质网(RER)到高尔基体的运输
在蛋白质合成过程中,新生蛋白质通过RER进行合成,然后通过囊泡转运到高尔基体进行 加工和分类。
跨膜运输
囊泡可以穿过细胞膜,将蛋白质从一个细胞器转运到另一个细胞器,如从高尔基体转运到 溶酶体或转运到细胞膜。
胞吐作用
当囊泡与细胞膜融合时,其内容物会被释放到细胞外,如神经递质的释放。
药物研发过程中,针对影响蛋白质分选的靶点进行设计, 可以实现对特定蛋白质的调控,从而达到治疗疾病的目的 。
THANKS FOR WATCHING
感谢您的观看
01
蛋白质通过内质网-高尔基体途径进行膜泡运输,该途径包括顺 面高尔基体、反面高尔基体和溶酶体等细胞器。
02
蛋白质还可以通过核膜-内质网途径进行膜泡运输,该途径涉及
核孔复合体和内质网等细胞器。
此外,还有其他的膜泡运输途径,如线粒体膜泡运输和叶绿体
改变离子通道和转运蛋白的活性,调节物质进出细胞的速率。
03
蛋白质的囊蛋白质分选的 重要途径之一,通过囊泡将蛋白 质从一处转运到另一处,实现蛋 白质的定位和功能。
02
囊泡运输涉及到多种细胞器之间 的相互联系和蛋白质的跨膜转运 ,对于维持细胞结构和功能具有 重要意义。
囊泡运输的调控机制
信号分子调控
某些信号分子可以与囊泡上的受 体结合,调控囊泡的转运方向和 目的地。
能量依赖性调控
囊泡运输需要消耗能量,如ATP 水解产生的能量,以驱动囊泡的 转运过程。
蛋白激酶与磷酸化
调控
某些蛋白激酶可以调控囊泡运输 相关蛋白的磷酸化状态,从而影 响囊泡的转运过程。
04
蛋白质的膜泡运输
囊泡运输的途径
蛋白质从粗面内质网(RER)到高尔基体的运输
在蛋白质合成过程中,新生蛋白质通过RER进行合成,然后通过囊泡转运到高尔基体进行 加工和分类。
跨膜运输
囊泡可以穿过细胞膜,将蛋白质从一个细胞器转运到另一个细胞器,如从高尔基体转运到 溶酶体或转运到细胞膜。
胞吐作用
当囊泡与细胞膜融合时,其内容物会被释放到细胞外,如神经递质的释放。
药物研发过程中,针对影响蛋白质分选的靶点进行设计, 可以实现对特定蛋白质的调控,从而达到治疗疾病的目的 。
THANKS FOR WATCHING
感谢您的观看
01
蛋白质通过内质网-高尔基体途径进行膜泡运输,该途径包括顺 面高尔基体、反面高尔基体和溶酶体等细胞器。
02
蛋白质还可以通过核膜-内质网途径进行膜泡运输,该途径涉及
核孔复合体和内质网等细胞器。
此外,还有其他的膜泡运输途径,如线粒体膜泡运输和叶绿体
细胞内蛋白质的分选和运输-细胞生物学-课件1-10
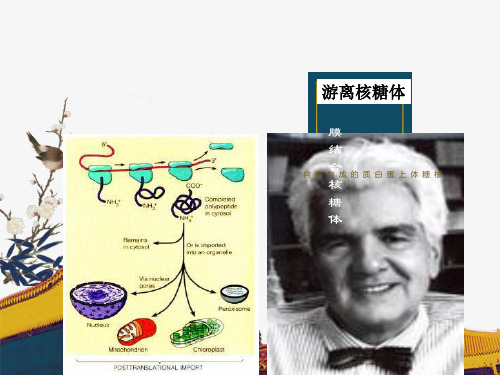
受体介导的内吞作用 (receptor-mediated endocytosis)
细胞摄入胆固醇的方法 (LDL颗粒)-受体介导的胞吞
有被小泡
01
02
遗传性高胆固醇血症原因之一
受体缺陷:不能形成有被小泡
溶酶体的形成与胞吞密切相关 早期内体 晚期内体 溶酶体 高尔基体
从细胞内到细胞表面的蛋白质分选和运输
门控运输为通过核孔复合体进出核的运输,是一个信号识别 (核输入、输出受体分别识别核输入、输出信号)和载体介导 的主动运输过程。具有选择性、双向性、耗能的特点。蛋白在 核质间的转运受严格的控制。
线粒体、内质网、过氧化物酶体三种细胞器的蛋白质输入皆 为穿膜运输。需要定位信号、信号识别的受体、蛋白质转运子 等多种结构的协同完成。
胞吞)将胞外大分子装入胞吞小泡,经内体到达溶酶体,并在 那里被消化降解。
胞吞途径是指细胞通过胞吞作用(吞噬、吞饮、受体介导的
室间,以及从高尔基体到细胞表面或溶酶体的蛋白运输过程。 其中既包括前向小泡运输,也有逆向小泡运输。
生物合成-分泌途径是指从内质网到高尔基体,高尔基体各区
思考题
一、解释名词
门控运输 穿膜运输 小泡运输 翻译后转运 共翻译转运 信号肽(斑) 靶向运输 生物合成-分泌途径 胞吞途径 受调分泌 固有分泌 吞噬作用 受体介导的内吞
细胞器之间通过运输小泡进行的蛋白质运输称为
第四节 细胞内蛋白质的小泡运输
伴随膜的运动 耗能 能运输膜蛋白、 膜脂、可溶性蛋白 要运输的蛋白质 衣被蛋白 运输小泡的形成
三种类型的有被小泡介导不同的运输途径
网格蛋白有被小泡:质膜-溶酶体,高尔基体-晚期内体
COPI有被小泡:高尔基体膜囊间,高尔基体-内质网
《蛋白质的分选》课件

细胞生物学技术
利用细胞生物学技术,如荧光标记、免疫荧光染色等,观察蛋白质在细胞内的定位和动 态变化,揭示蛋白质分选的细胞生物学过程。
蛋白质分选的潜在治疗策略
靶向治疗
针对异常表达的蛋白质或蛋白质分选相关基 因进行靶向治疗,以纠正异常的蛋白质分选 过程,治疗相关疾病。
基因治疗
通过基因工程技术,对相关基因进行修饰或敲除, 以改变蛋白质的表达和分选,达到治疗目的。
膜泡运输
01
指蛋白质在细胞质基质中形成膜泡,然后通过胞吐作用将膜泡
释放到细胞外或细胞内的其他部位。
膜泡运输的类型
02
包括内吞作用、外排作用和胞饮作用等,每种类型都有其特定
的运输途径和作用。
膜泡运输的机制
03
涉及多种蛋白质和细胞器的协同作用,如网格蛋白、细胞骨架
和溶酶体等。
蛋白质的细胞质运输途径
细胞质运输
针对蛋白质分选过程的治疗策略可能 对癌症治疗具有重要意义。
蛋白质分选与神经退行性疾病的关系
神经退行性疾病是指神经元或 神经胶质细胞逐渐退化并导致 功能障碍的一类疾病,如帕金
森病、阿尔茨海默病等。
某些神经退行性疾病可能与蛋 白质聚集物的形成有关,这些 聚集物可能干扰蛋白质的正常 分选和功能。
蛋白质分选异常可能导致神经 元死亡和神经退行性疾病的发
3
临床应用转化
将蛋白质分选的研究成果转化为临床应用,开发 新的治疗策略和方法,为疾病治疗提供更多选择 。
THANKS
THANK YOU FOR YOUR WATCHING
05
蛋白质分选的研究前景与展望
蛋白质分选的研究方法与技术
蛋白质组学技术
利用蛋白质组学技术,如质谱分析、蛋白质免疫印迹等,对蛋白质进行定性和定量分析 ,深入了解蛋白质分选的机制和过程。
利用细胞生物学技术,如荧光标记、免疫荧光染色等,观察蛋白质在细胞内的定位和动 态变化,揭示蛋白质分选的细胞生物学过程。
蛋白质分选的潜在治疗策略
靶向治疗
针对异常表达的蛋白质或蛋白质分选相关基 因进行靶向治疗,以纠正异常的蛋白质分选 过程,治疗相关疾病。
基因治疗
通过基因工程技术,对相关基因进行修饰或敲除, 以改变蛋白质的表达和分选,达到治疗目的。
膜泡运输
01
指蛋白质在细胞质基质中形成膜泡,然后通过胞吐作用将膜泡
释放到细胞外或细胞内的其他部位。
膜泡运输的类型
02
包括内吞作用、外排作用和胞饮作用等,每种类型都有其特定
的运输途径和作用。
膜泡运输的机制
03
涉及多种蛋白质和细胞器的协同作用,如网格蛋白、细胞骨架
和溶酶体等。
蛋白质的细胞质运输途径
细胞质运输
针对蛋白质分选过程的治疗策略可能 对癌症治疗具有重要意义。
蛋白质分选与神经退行性疾病的关系
神经退行性疾病是指神经元或 神经胶质细胞逐渐退化并导致 功能障碍的一类疾病,如帕金
森病、阿尔茨海默病等。
某些神经退行性疾病可能与蛋 白质聚集物的形成有关,这些 聚集物可能干扰蛋白质的正常 分选和功能。
蛋白质分选异常可能导致神经 元死亡和神经退行性疾病的发
3
临床应用转化
将蛋白质分选的研究成果转化为临床应用,开发 新的治疗策略和方法,为疾病治疗提供更多选择 。
THANKS
THANK YOU FOR YOUR WATCHING
05
蛋白质分选的研究前景与展望
蛋白质分选的研究方法与技术
蛋白质组学技术
利用蛋白质组学技术,如质谱分析、蛋白质免疫印迹等,对蛋白质进行定性和定量分析 ,深入了解蛋白质分选的机制和过程。
蛋白质分选的基本途径与类型

NLS序列可存在于亲核蛋白的不同部位,在指导 完成核输入后并不被切除;
NLS只是亲核蛋白入核的一个必要条件而非充分 条件 。
转录产物RNA的核输出
真核细胞中转录后的RNA通常需加工、修饰成为 成熟的RNA分子后才能被转运出核。 (1)RNA聚合酶I转录的rRNA分子:以RNP的形 式离开细胞核,需要能量; (2)RNA聚合酶III转录的5s rRNA与 tRNA的核 输出由蛋白质介导;
(3)RNA 聚合酶II转录的hn RNA,在核内进行 5′端加帽和3′端附加多聚A序列以及剪接等加工 过 程 , 然 后 形 成 成 熟 的 mRNA 出 核 , 5′ 端 的 m7GpppG“帽子”结构对mRNA的出核转运是 必要的;
细胞核中既有正调控信号保证mRNA的出核转运, 也有负调控信号防止mRNA的前体被错误地运输, 后者与剪接体(spliceosome)有关。
2)叶绿体蛋白质的运送及装配
转运肽(transit peptides):细胞质中合成的叶 绿体前体蛋白,在N端也含有一个额外的氨基酸 序列。
目前研究的较多的是类囊体膜和内囊体腔中蛋 白质的运送过程。如补光色素蛋白或称叶绿素 a/b结合蛋白前体,其转运肽含有35个氨基酸残 基,引导其穿过叶绿体进入基质,在基质中由特 异的蛋白酶加工切去转运肽成为成熟的蛋白质。
分子伴侣(molecular chaperones):细胞中的一 类蛋白质,可以识别正在合成的多肽或部分折叠 的多肽并与多肽的某些部位相结合,从而帮助这 些多肽转运、折叠或装配,但是其本身并不参与 最终产物的形成。
大部分属于热休克蛋白(hsp)进化上很保 守,无专一性。
跨膜蛋白运输机制 布朗棘轮模型(Brownian rachet model):Simon 线粒体基质Hsp70(mHsp70):转运发动机
NLS只是亲核蛋白入核的一个必要条件而非充分 条件 。
转录产物RNA的核输出
真核细胞中转录后的RNA通常需加工、修饰成为 成熟的RNA分子后才能被转运出核。 (1)RNA聚合酶I转录的rRNA分子:以RNP的形 式离开细胞核,需要能量; (2)RNA聚合酶III转录的5s rRNA与 tRNA的核 输出由蛋白质介导;
(3)RNA 聚合酶II转录的hn RNA,在核内进行 5′端加帽和3′端附加多聚A序列以及剪接等加工 过 程 , 然 后 形 成 成 熟 的 mRNA 出 核 , 5′ 端 的 m7GpppG“帽子”结构对mRNA的出核转运是 必要的;
细胞核中既有正调控信号保证mRNA的出核转运, 也有负调控信号防止mRNA的前体被错误地运输, 后者与剪接体(spliceosome)有关。
2)叶绿体蛋白质的运送及装配
转运肽(transit peptides):细胞质中合成的叶 绿体前体蛋白,在N端也含有一个额外的氨基酸 序列。
目前研究的较多的是类囊体膜和内囊体腔中蛋 白质的运送过程。如补光色素蛋白或称叶绿素 a/b结合蛋白前体,其转运肽含有35个氨基酸残 基,引导其穿过叶绿体进入基质,在基质中由特 异的蛋白酶加工切去转运肽成为成熟的蛋白质。
分子伴侣(molecular chaperones):细胞中的一 类蛋白质,可以识别正在合成的多肽或部分折叠 的多肽并与多肽的某些部位相结合,从而帮助这 些多肽转运、折叠或装配,但是其本身并不参与 最终产物的形成。
大部分属于热休克蛋白(hsp)进化上很保 守,无专一性。
跨膜蛋白运输机制 布朗棘轮模型(Brownian rachet model):Simon 线粒体基质Hsp70(mHsp70):转运发动机
细胞内蛋白质的分选和运输课件

信号假说
如何实现膜泡在细胞器之间的靶向运输?
小泡如何形成?小泡内的货物如何分 选和装运?小泡如何实现靶向运输?
小泡的靶向运输
Paired sets of SNARE proteins mediate fusion of vesicles with target membranes.
Rab proteins and SNAREs help direct transport vesicles to their target membranes
Inner Membrane Intermembrane space
Outer Membrane
Additional targeting sequences specify other locations
Example: ATP Synthase subunit on inner membrane
Yeast cells expressing mutant Rab protein accumulate secretory vesicles that are unable to fuse with the plasma membrane.
Figure 15-21 Essential Cell Biology (© Garland Science 2010)
non-insulin dependent, normal insulin production,
lack of response to insulin
膜蛋白的上膜也可以是受调的
Hydropathy profiles of integral membrane proteins
All have hydrophobic domains
如何实现膜泡在细胞器之间的靶向运输?
小泡如何形成?小泡内的货物如何分 选和装运?小泡如何实现靶向运输?
小泡的靶向运输
Paired sets of SNARE proteins mediate fusion of vesicles with target membranes.
Rab proteins and SNAREs help direct transport vesicles to their target membranes
Inner Membrane Intermembrane space
Outer Membrane
Additional targeting sequences specify other locations
Example: ATP Synthase subunit on inner membrane
Yeast cells expressing mutant Rab protein accumulate secretory vesicles that are unable to fuse with the plasma membrane.
Figure 15-21 Essential Cell Biology (© Garland Science 2010)
non-insulin dependent, normal insulin production,
lack of response to insulin
膜蛋白的上膜也可以是受调的
Hydropathy profiles of integral membrane proteins
All have hydrophobic domains
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
转录产物RNA的核输出
真核细胞中转录后的RNA通常需加工、修饰成为
成熟的RNA分子后才能被转运出核。
(1)RNA聚合酶I转录的rRNA分子:以RNP的形
式离开细胞核,需要能量;
(2)RNA聚合酶III转录的5s rRNA与 tRNA的核
输出由蛋白质介导;
(3)RNA 聚合酶II转录的hn RNA,在核内进行 5′端加帽和 3′端附加多聚 A 序列以及剪接等加工 过 程 , 然 后 形 成 成 熟 的 mRNA 出 核 , 5′ 端 的 m7GpppG“ 帽子”结构对 mRNA 的出核转运是 必要的;
线粒体和叶绿体蛋白质的运转与装配
1)线粒体蛋白质的转运与装配 导肽(leader peptide):N端引导蛋白质转运的一 段氨基酸序列。20~80个氨基酸序列。特点: (1)含有丰富的碱性氨基酸,特别是Arg氨基酸; (4)可形成亲水性和疏水性的α螺旋结构;
三 选择性的门控转运(gated transport) 指在细胞质基质中合成的蛋白质通过核孔复合体 选择性的完成核输入或从细胞核返回细胞质基质; 核孔复合体主要由蛋白质构成,其总相对分子 质量约为125×106,推测可能含有100余种不 同的多肽,共1 000多个蛋白质分子。
1)核孔复合体成分
gp210:结构性跨膜蛋白 p62:功能性的核孔复合体蛋白,具有两个功能 结构域; 疏水性N端区:可能在核孔复合体功能活动中直 接参与核质交换; C端区:可能通过与其它核孔复合体蛋白相互作 用,从而将p62分子稳定到核孔复合体上,为其N 端进行核质交换活动提供支持;
蛋白质分选的基本途径与类型
刘媛媛
蛋白质的分选大体可分为两条途径: (1)后转运:游离核糖体上合成的蛋白质如用于 催化代谢的酶类、核蛋白、线粒体和叶绿体蛋白 质等。 (2)共转运:在粗面内质网(ER)合成的蛋白质 如膜的整合蛋白、胞外分泌蛋白、构成细胞器中 的可溶性驻留蛋白等。
蛋白质分选的类型或机制的角度,可以分为四类:
细胞核中既有正调控信号保证 mRNA 的出核转运,
也有负调控信号防止mRNA的前体被错误地运输,
后者与剪接体(spliceosome)有关。 3’端在后。
mRNA的出核转运过程是有极性的,其5’端在前,
核输出信号 (Nuclear Export Signal , NES) :
RNA分子的出核转运需要蛋白分子的帮助,这些
核定位信号 (nuclear localization signal)特点
NLS是存在于亲核蛋白内的一些短的氨基酸序列 片段,富含碱性氨基酸残基,如 Lys 、 Arg ,此 外还常含有Pro; NLS 的氨基酸残基片段可以是一段连续的序列 (T抗原),也可以分成两段,两段之间间隔约 10个氨基酸残基(核质蛋白); NLS序列可存在于亲核蛋白的不同部位,在指导 完成核输入后并不被切除; NLS只是亲核蛋白入核的一个必要条件而非充分 条件 。
蛋白因子本身含有出核信号。
入核转运与出核转运之间有某种联系,它们可
能需要某些共同的因子。
B
A
二 膜泡运输(vesicular transport) 蛋白质通过不同类型的转运小泡从其粗面内质 网合成部位转运至高尔基体进而分选至细胞的 不同部位,其中涉及各种不同的定向转运,及 膜泡出芽与融合的过程。 目前发现三种不同类型的有被小泡具有不 同的物质运输作用:网格蛋白有被小泡, COPⅡ有被小泡,COPⅠ有被小泡。
2)叶绿体蛋白质的运送及装配 转运肽(transit peptides):细胞质中合成的叶 绿体前体蛋白,在N端也含有一个额外的氨基酸 序列。 目前研究的较多的是类囊体膜和内囊体腔中蛋 白质的运送过程。如补光色素蛋白或称叶绿素 a/b结合蛋白前体,其转运肽含有35个氨基酸残 基,引导其穿过叶绿体进入基质,在基质中由特 异的蛋白酶加工切去转运肽成为成熟的蛋白质。
分子伴侣(molecular chaperones):细胞中的一 类蛋白质,可以识别正在合成的多肽或部分折叠 的多肽并与多肽的某些部位相结合,从而帮助这 些多肽转运、折叠或装配,但是其本身并不参与 最终产物的形成。 大部分属于热休克蛋白(hsp)进化上很保 守,无专一性。
跨膜蛋白运输机制 布朗棘轮模型(Brownian rachet model):Simon 线粒体基质Hsp70(mHsp70):转运发动机
(1)蛋白质的跨膜转运 (transmembrane transport) (2)膜泡运输(vesicular transport ) (3)选择性的门控转运(gated transport ) (4)细胞质机制中的蛋白质的转运
一 蛋白质的跨膜转运(transmembrane transport) 主要是指细胞质基质中合成的蛋白质转运到内 质网、线粒体、质体(包括叶绿体)和过氧化物 酶体等细胞器的一种分选方式。
2)核孔复合体功能 一种特殊的跨膜运输蛋白复合体,是一个双 功能、双向性的亲水性核质交换通道。 双功能:有两种运输方式被动运输与主动运输; 双向性:介导蛋白质的入核转运,介导RNA 、 核糖核蛋白颗粒(RNP)的出核转运。
3)亲和蛋白质入核转运机制
(1)亲核蛋白(karyophilic protein) 在细胞质内合成后,需要或能够进入细胞核内 发挥功能的 一类蛋白质; ( 2 )核定位信号 (nuclear localization signal , NLS) 亲和蛋白的含有的特殊的氨基酸序列,具 有定向定位作用的一段序列; (3)胞质蛋白因子:importinα /β 、Ran等。