转录 ppt课件
合集下载
第十三章基因的转录、转录后加工及逆转录ppt课件
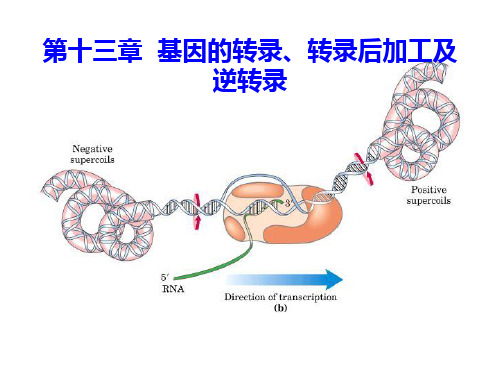
◆真核生物启动子 (1)DNA序列在转录起始点的5’端区(上游 区)(2)-25bp :TATA盒(Hogness box) (3)-90bp :GC盒 (4)-70bp :CAAT盒
-90
-70
GC
CAAT
RNA聚合酶Ⅱ催化的转录起始
RNA聚合酶Ⅱ催化各种前体mRNA的合 成
需要多种TF参与:TFⅡA-J
第一节 参与转录的酶
RNA聚合酶——依赖DNA的RNA聚合酶 (DNA-dependent RNA polymerase,DDRP)
以DNA为模板,催化2个游离的NTP 形成3’,5’-磷酸二酯键
一、原核生物RNA聚合酶
1、大肠埃希菌RNA聚合酶的组成 (1)全酶(holoenzyme)
由4种(5个)亚基α2ββ’σ组成 (2)核心酶(core enzyme)
作用位置
步骤1 步骤2
200~250
(3)真核生物mRNA转录后加工—剪接
内含子
外显子
DNA
hnRNA
●剪接所需条件
snRNA (U1-U6) + 蛋白质 (核内小分子核酸)
多种snRNP (核内小核蛋白颗粒)
多种snRNPs装配成
剪接体 (参与剪接过程)
4、RNA编辑(RNA editing)
二.真核生物RNA转录后的加工 1、rRNA转录后的加工
真核生物rRNA 的基因
(rDNA)
转录产物
成簇纵列串联排列
高度重复序列DNA
核质:(Ⅲ)--不需加 工
5s rRNA
核仁:(Ⅰ)--加工
5.8s rRNA 28s rRNA 18s rRNA
rDNA 内含子
基因间隔
遗传信息的转录PPT课件

G
-
13
A A T C AA T AG U UA G
G
-
14
A A T C AA T AG U UA G U
G
-
15
A A T C AA T AG U UA G UU
G
-
16
A A T C AA T AG U UA G UU A
G
-
17
A A T C AA T AG U UA G UU AU
-
18
A A T C AA T AG U UA G UU AUC
-
19
DNA的一条单链即模板链 A A T C AA T AG U UA G UU AUC
mRNA
-
20
mRNA通过核孔进入细胞质
细
胞
核
A AT C AA U UAGAU
T A
AU
G C
细
胞
质
-
21
1、转录与DNA复制有什么共同之处?
-
3
DNA,RNA 的主要区别
比较项目
DNA
RNA
基本单位
脱氧核苷酸
核糖核苷酸
五碳糖
脱氧核糖
核糖
含氮碱基 A T C G
AUCG
结构
多为双链结构 多为单链结构
主要存在部位 细胞核
细胞质
★RNA是由核苷酸连接而成,跟DNA一样能储存遗
★传R信N息A一。般是单____链,而且比 DNA 短,因此能够通过
A.胰腺细胞 B.肠黏膜细胞
C
C.成熟红细胞 D.白细胞
-
25
3、如图是DNA转录过程中的一个片段,其
核苷酸的种类有(C)
-
13
A A T C AA T AG U UA G
G
-
14
A A T C AA T AG U UA G U
G
-
15
A A T C AA T AG U UA G UU
G
-
16
A A T C AA T AG U UA G UU A
G
-
17
A A T C AA T AG U UA G UU AU
-
18
A A T C AA T AG U UA G UU AUC
-
19
DNA的一条单链即模板链 A A T C AA T AG U UA G UU AUC
mRNA
-
20
mRNA通过核孔进入细胞质
细
胞
核
A AT C AA U UAGAU
T A
AU
G C
细
胞
质
-
21
1、转录与DNA复制有什么共同之处?
-
3
DNA,RNA 的主要区别
比较项目
DNA
RNA
基本单位
脱氧核苷酸
核糖核苷酸
五碳糖
脱氧核糖
核糖
含氮碱基 A T C G
AUCG
结构
多为双链结构 多为单链结构
主要存在部位 细胞核
细胞质
★RNA是由核苷酸连接而成,跟DNA一样能储存遗
★传R信N息A一。般是单____链,而且比 DNA 短,因此能够通过
A.胰腺细胞 B.肠黏膜细胞
C
C.成熟红细胞 D.白细胞
-
25
3、如图是DNA转录过程中的一个片段,其
核苷酸的种类有(C)
遗传信息的转录和翻译ppt课件.pptx

DNA → DNA
细胞分裂间期 2个相同的DNA分子 边解旋边复制
半保留复制
DNA → mRNA
mRNA → 蛋白质
生物生长发育的过程中
RNA
蛋白质
边解旋边转录 DNA仍保留
1个mRNA分子可结合 多个核糖体
1.RNA的组成、结构与类型。 2.遗传信息的转录和翻译过程。 3.遗传信息、密码子与反密码子的作用 4.DNA复制、转录和翻译的比较
为什么RNA适于作DNA的信使?
RNA由基本单位——核苷酸连接而成,也 可以储存遗传信息;
RNA能通过核孔,从细胞核转移到细胞质。
RNA遵循“碱基互补配对原则”,以RNA为 媒介可将遗传信息传递到细胞质中。
DNA → DNA A —— T、C —— G T ——A、 G ——C
酶( RNA 聚合酶等) 和ATP
DNA → mRNA A —— U、C —— G T ——A、 G ——C
酶、ATP 和tRNA
mRNA → tRNA A —— U、C —— G U ——A、 G ——C
信息传递 时间 产物 特点
密码子
在mRNA上
直接控制蛋白质的氨基酸的排列 顺序
反密码子 在tRNA上
识别密码子
2、转录、翻译与DNA复制的比较
项目 场所
DNA复制 主要是细胞核
转录 主要是细胞核
翻译 细胞质和核糖体
模板
DNA的两条链
D种核糖核苷酸
20种氨基酸
其他条件
碱基配对 方式
酶( 解旋酶、DNA 聚合酶等)和ATP
1、遗传信息、密码子和反密码子 (1)遗传信息:指基因(或DNA)中控制遗传性状的脱氧核苷 酸顺序,它间接决定氨基酸的排列顺序。
第四章-细胞的复制与转录PPT课件
奇迹是怎样创造的?
•3
第四节 遗传物质的复制
replication of the genomic materials 一、DNA 复制过程及其特征 二、DNA 复制的有关酶类和蛋白质
•4
一、DNA 复制过程及其特征
• 半保留复制 • 复制从复制起始点开始 • 复制叉、双向复制和复制泡 • 复制叉的不对称性:前导链、后随链和
配校读
DNA连接酶
连接岗崎片段3‘-OH与5’-P •36
第四章 细胞核nucleus
第一节 核被膜 第二节 染色质和染色体 第三节 核仁 第四节 遗传物质的复制 第五节 遗传信息的表达
•37
分化
受精卵→胚胎干细胞 →→→ 各种细胞
分化:基因选择性表达的结果
红细胞:专一表达血红蛋白
分化
•38
一个人的两种细胞:源于同一个细胞(同一个受精卵),携 带完全相同的遗传信息却有着截然不同的形态和功能。
点向两个方向移动
•15
3.复制叉、双向复制和复制泡 (replication forks,
two-direction replication & replication bubbles)
复制叉: 已经打开的2条单链与未解开的双链间形成•1Y6 形。
复制从复制起始点 开始向两个方向推进, 形成两个反向的复制叉。
由RNA聚合酶转录
RNA聚合酶:催化这一合成反应,所以又叫转录酶。作用
是解开DNA双链,以其中的反基因链为模板,将4种三磷酸
核苷酸聚合成与模板互补的RNA链。
•51
RNA聚合酶催化的细菌基因的转录:聚合酶遇到启动子序列 就与之结合,打开双链开始转录,直至遇到终止子.•卡52 通
•3
第四节 遗传物质的复制
replication of the genomic materials 一、DNA 复制过程及其特征 二、DNA 复制的有关酶类和蛋白质
•4
一、DNA 复制过程及其特征
• 半保留复制 • 复制从复制起始点开始 • 复制叉、双向复制和复制泡 • 复制叉的不对称性:前导链、后随链和
配校读
DNA连接酶
连接岗崎片段3‘-OH与5’-P •36
第四章 细胞核nucleus
第一节 核被膜 第二节 染色质和染色体 第三节 核仁 第四节 遗传物质的复制 第五节 遗传信息的表达
•37
分化
受精卵→胚胎干细胞 →→→ 各种细胞
分化:基因选择性表达的结果
红细胞:专一表达血红蛋白
分化
•38
一个人的两种细胞:源于同一个细胞(同一个受精卵),携 带完全相同的遗传信息却有着截然不同的形态和功能。
点向两个方向移动
•15
3.复制叉、双向复制和复制泡 (replication forks,
two-direction replication & replication bubbles)
复制叉: 已经打开的2条单链与未解开的双链间形成•1Y6 形。
复制从复制起始点 开始向两个方向推进, 形成两个反向的复制叉。
由RNA聚合酶转录
RNA聚合酶:催化这一合成反应,所以又叫转录酶。作用
是解开DNA双链,以其中的反基因链为模板,将4种三磷酸
核苷酸聚合成与模板互补的RNA链。
•51
RNA聚合酶催化的细菌基因的转录:聚合酶遇到启动子序列 就与之结合,打开双链开始转录,直至遇到终止子.•卡52 通
第五章转录ppt课件

一、E.coli RNA聚合酶
E.coli RNA聚合酶由5个亚基组成(5个 多肽链),即α2ββ′σ
αββ′σ四个亚基的分子量分别为36.5KDa、 150 KDa、160KDa和82 KDa,整个酶 分子的分子量为465KDa
分别是基因rpoA、rpoB、rpoC和rpoD的
产物
RNA聚合酶Ⅲ也存在于核质中,其功 能是合成tRNA和5S rRNA以及转录 Alu序列
在细胞质中也能发现一些RNA聚合酶 Ⅲ,它是从细胞核中渗漏出来的。
三种主要的RNA聚合酶的分子量都在 500 KDa左右(14S-15S),每种酶分子 含有两个大亚基和4~8个小亚基,每 个小亚基的分子量为10 KDa-90KDa
这些名称最早是依据它们从DEAE-纤维素柱上洗脱的 先后顺序而定出来的。后来发现不同生物的三种RNA聚 合酶的洗脱顺序并不相同,因而改用三种不同的RNA聚 合酶对于α-鹅膏蕈碱(α-amanitine)的敏感性不同来 进行区别。RNA聚合酶I基本不受α-鹅膏蕈碱的抑制, 在大于10-3 mol/L时才表现出轻微的抑制作用;RNA聚 合酶Ⅱ对于α-鹅膏蕈碱最为敏感,在10-9-10-8mol/L 浓度下就会被抑制;RNA聚合酶Ⅲ的敏感性介于RNA聚 合酶Ⅰ和Ⅱ之间,在10-5-10-4 mol/L时表现抑制作用。
与RNA聚合酶相结合的一个很小的蛋白质(MW= 10KDa),叫做ω亚基,其功能尚不清楚,有人认 为ω亚基对于RNA聚合酶的结构和功能没有太大的 影响。
原核生物的 RNA聚合酶
亚基 分子量
36512
150618
155613
70263
功能
决定哪些基因被转录 催化功能
转录ppt(共66张PPT)

RNA 聚合酶
Ⅰ
Ⅱ
Ⅲ
转录
产物
加工
45SrRNA
18SrRNA 28SrRNA
hnRNA tRNA 5SRNA
snRNA
mRNA
利福平
抑制原核生物的RNA聚合酶
鹅膏蕈碱
抑制真核生物的RNA聚合酶
利福平 主要用于治疗结核病、麻风病等
鹅膏蕈碱 鬼笔鹅膏
三、真核生物的转录产物为单顺
反子
与原核生物不同,真核生物一个转 录单位仅生成一个mRNA分子,经翻 译生成一条多肽链
(三)都形成3’, 5’-磷酸二酯键
二、复制与转录的不同点
模板
原料
酶
校读
复制 两条链 dNTP DNA聚合酶
有
转录 一条链
NTP RNA聚合酶
无
配对
产物
引物
复制
A=T GC
子代双
链DNA
需要
转录
A=U T=A
GC
mRNA tRNA rRNA
不需要
第一节 原核生物的转录
一、转录模板 二、RNA聚合酶 三、模板和酶的辨认结合
第 11 章
RNA的生物合成(转录)
DNA DNA DNA
RNA
蛋白质
中心法则
目录
前 言 复制与转录的异同 第1节 原核生物的转录
第2节 真核生物的转录特点
第3节 真核生物转录后加工
前言 复制与转录的异同
一、相同点 二、不同点
一、复制与转录的相同点
(一)服从碱基配对规则
(二)合成方向都是5’-3’
DNA-dependent RNA polymerase (二)合成方向都是5’-3’ (四)腺嘌呤脱氨成为次黄嘌呤 σ亚基从三元起始复合物上脱落后,核心酶的构象随之发生改变,并沿着模板链的3’→5’方向滑行,进入延长阶段 第1节 原核生物的转录 原核生物的RNA聚合酶仅1种,合成全部RNA 主要用于治疗结核病、麻风病等 真核生物rRNA转录后加工 α2 β β’ ω σ称为全酶
最新转录(transcription)是指以DNA为模板ppt课件

第二节 启动子
启 动 子 ( promoter ) 是 指 DNA 分 子 上 被 RNA 聚 合酶识别并结合形成起始转录复合物的区域,它 还包括一些调节蛋白因子的结合位点。无论是原 核生物还是真核生物,启动子是控制转录起始的 序列,并决定着某一基因的表达强度。与RNA聚 合酶亲和力高的启动子,其起始频率和效率均高。 启动子的DNA序列本身就可以提供特定的信号, 而转录区域的DNA序列要转变成RNA或蛋白质 后才能体现它所贮存的信息。原核生物和真核生 物的启动子的结构还有些差异(见图5-2)。
(4)-10区和-35区间的距离 -35区和-10 区之间的距离在90%的原核生物启动子为16到 18bp。少数情况下,该距离可少至15bp,多至 20bp。该区域内的碱基序列并不重要,但碱基 的数目即该距离的长短却是至关重要的。适宜的 距离可以为RNA聚合酶提供合适的空间结构,便 于转录的起始。
2、σ因子
σ因子在RNA聚合酶识别启动子的过程中起 关键作用,大肠杆菌至少有4种σ因子,σ70负 责 识 别 一 般 的 启 动 子 , σ32 识 别 热 激 ( heat shock)蛋白质基因启动子,σ38负责识别碳 源饥饿条件下的基因启动子,σ54识别与氮代 谢有关的基因启动子。
表5–1 大肠杆菌的σ因子及其特性
转录(transcription)是指以 DNA为模板
转录(transcription)是指以DNA为模板,在依赖 于 DNA 的 RNA 聚 合 酶 (DNA-dependent RNA polymerase)的催化下,以4种rNTP(ATP、CTP、 GTP和UTP)为原料,沿5′→3′方向合成RNA 的过程。RNA是DNA将遗传信息传递给蛋白质的 中心环节。在有些RNA病毒中,RNA也可以指导 合成RNA。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
启动子的一致序列是综合统计了多种基因的启动子序
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
3 转录
3.1 转录的要点 3.2 原核生物的转录 3.3 真核生物的转录 3.4 RNA 的剪辑与加工
ppt课件
1
中心法则
R everse tra n scrip tio n D N A syn th esis
ppt课件
2
3.1 转录的纲要
ppt课件
3
关键词
•转录(transcription) :拷贝出一条与DNA链序列完全相同 的RNA单链的过程。
•有义链 sense strand (编码链coding strand)与mRNA序列 相同的那条DNA链。
•反义链 antisense strand (模板链Template strand) 有义链的 互补链,根据碱基配对原则指导mRNA合成的DNA链。
ppt课件
4
关键词
•启动子(Promoter) RNA聚合 酶识别和结合并启动转录的 DNA序列.
随着RNA 链的延伸,转录泡与核心酶一起沿着DNA模板链同步移
动。释放出来的σ因子可以重新与核心酶结合,启动新一轮DNA
的转录,称为RNA在转录泡前面解链, 同时在转录泡后面重新形成双链,拓扑异构酶 能够在转录泡的前方解除因解链形成的正超螺 旋,在转录泡的后方解除因解链形成的负超螺 旋。
ppt课件
16
启动子有三个部分
1) -10区, 又称 Pribnow Box 保守序列为T80A95T 45A60A50T96 。在RNA聚合酶的作用下,富含
AT的Pribnow框内的DNA双螺旋首先发生解链,与RNA聚合酶形成 开放性启动子复合物。(下标表示碱基在不同基因的启动子中出现
的概率 )
与模板DNA的结合部位。
ppt课件
10
sigma 亚基只在转录的起始阶段起作用
Sigma因子 是酶的别构效应物, 提高RNA聚合酶对启动子的亲和 力,降低与模板DNA上非特异位 点的结合力。
大肠杆菌中有多个因子, 每个 因子能识别不同的启动子。
ppt课件
11
E.coli的σ因子由基因rpo D编码,其主要作用是特异
ppt课件
23
3.2.4 转录延伸
RNA链延伸的过程
一旦RNA pol合成了大约10nt左右的RNA链,延伸便开始了。这
时转录物的长度足以让RNA取代σ因子的位置,使RNA pol的核心
酶与σ因子解离。核心酶因此而可以离开启动子,沿模板链移动
了,这一过程称启动子清空(promoter clearance)。在σ因子被释
ppt课件
6
3.2 原核生物的转录
3.2.1 RNA 聚合酶 3.2.2 启动子的组成 3.2.3 转录的起始和延伸 3.2.4 大肠杆菌中两种终止子
ppt课件
7
3.2.1 RNA聚合酶
RNA聚合酶核心酶: 2
RNA聚合酶全酶: 2
RNA聚合酶各亚基的 功能(见左图)
ppt课件
8
ppt课件
9
细菌RNA pol的核心酶含两个相同的α亚基(由rpoA
放以后,延伸因子Nus-A蛋白加入进来,转录即进入延伸阶段。
失去σ因子的核心酶通过封闭的钳子握住DNA,以更快的速度沿
着DNA模板链移动,使延伸反应可以持续进行。RNA pol的结构
变化使延伸中的RNA 链从RNA/DNA杂交双链中脱离,单链DNA重
新与互补链配对,转录泡的大小维持在17 bp左右。也就是说,
•封闭复合物: 全酶 ·DNA 双链体
•开放复合物: 全酶 ·开链 DNA 双链体
• 三元复合物 (Ternary complex): 全酶 ·开 链 DNA 双链体·新生 RNA (up to 9 nucleotides)
•当新生 RNA达到8-9个核苷酸长度时 , 因子 从RNA聚合酶上脱离下来,形成延伸三元复 合物,包括核心酶·DNA ·新生 RNA.
ppt课件
25
ppt课件
26
延伸的暂停和阻滞
在延伸阶段,RNA pol每催化1个新的磷酸二酯键 形成,就面临3种选择,其一是继续延伸合成新的磷 酸二酯键,其二是倒退切除新掺入的核苷酸,其三是 延伸复合物解离,完全停止转录。 (1) 暂停 若 在转录过程中,转录产物形成了特定的二级结构,如 发夹结构,或NTP暂时短缺,均有可能造成转录暂停。 暂停有可能使原核生物转录和翻译同步,也有可能对 转录调节蛋白发挥作用有帮助,还有可能是转录倒退 或完全终止的前奏。RNA合成的重新启动需要GreA和 GreB蛋白来解除暂停状态,在RNA pol倒退后,GreA 和GreB切除3'-端几个核苷酸,以便让RNA的3'-OH能 重新回到活性中心。
ppt课件
19
(1) RNA pol全酶与双链DNA的非特异性结合
核心酶与σ因子结合成全酶,RNA pol全 酶与非特异性DNA序列具有一定的亲和性, 但亲和性较低。然而,一旦聚合酶与DNA结 合,即可沿DNA滑动扫描,直到发现启动子 序列。RNA pol与启动子的结合则是特异性 的,具有很高的亲和性。
性地识别转录的起始位点启动子。启动子(promoter) 是RNA聚合酶识别、结合并起始转录的一段特异性 的DNA序列,一般位于转录起始位点(+1)的上游, 可以同RNA聚合酶特异性结合,但本身的序列不被 转录。核心酶虽可与DNA结合,但不能区别启动子 和一般的DNA序列,或者说不能正确识别启动子的 特异序列。σ因子与核心酶结合后,全酶对启动子的 特异性结合能力是对其他DNA序列结合能力的107倍。 其原因是,核心酶对DNA序列结合的亲和力建立在 碱性蛋白和酸性核酸之间静电吸附作用力之上,这 种结合是没有特异性的松散结合,而且被结合的 DNA仍然保持双螺旋状态。σ因子能大大降低RNA pol与一般DNA序列的结合常数和停留时间,同时又 增加RNA pol与启动子的结合常数和停留时间,使 RNA pol能正确的识别启动子的特异序列,并与之结 合。
•转录起始点 (startsite) 与新生 RNA链第一个核苷酸相对应 DNA链上的碱基,通常为一个 嘌呤
•终止子(terminator) DNA上引起 RNA聚合酶终止转录的序列.
ppt课件 Transcription unit 5
RNA的酶促合成
rNTP n rNTP
DNA template RNA-P, Mg2
ppt课件
21
(3) 封闭复合物转化成开放复合物
σ因子使DNA部分解链,形成大小为12 bp~17 bp的转录泡,使 DNA模板链进入活性 中心,封闭复合物转变成开放复合物(open complex)。一开始,转录泡覆盖–10~–1,但 它很快以一种依赖于Mg 2+的方式从–12延伸 到+2。开放复合物十分稳定,其半寿期在几 个小时以上,此时的聚合酶与启动子的相互 作用既有静电引力,又有氢键。足印法测定 表明,在此阶段聚合酶复盖–55~+20区域。
ppt课件
20
(2) RNA pol与启动子形成封闭复合物
一旦RNA pol遇到–35区,便形成封闭复 合物(closed complex)。在此阶段,DNA并没 有解链,聚合酶主要以静电引力与DNA结合。 这种复合物并不十分稳定,半衰期为15min~ 20min。足印法分析表明,在此阶段聚合酶复 盖–55到+5区域。
ppt课件
22
(4) RNA合成的起始
在前两个与模板链互补的NTP从次级通道进入聚 合酶的活性中心以后,由活性中心催化第一个NTP的 3'-OH亲核进攻第二个NTP的5'-磷酸基,形成第一个 磷酸二酯键。第一个掺入的核苷酸总是嘌呤核苷酸, 这是因为聚合酶的第一个NTP结合位点优先结合嘌呤 核苷三磷酸,而第二个NTP结合位点与4种NTP结合的 亲和力相同。此外,新合成的初级转录物一般含有 5'-三磷酸,这与成熟的RNA分子不同。一旦有了第 一个磷酸二酯键,RNA-DNA-RNA pol的三元复合物 (the ternary complex)就形成了。RNA pol全酶催 化形成6个~10个磷酸二酯键以后,与核心酶结合 的σ因子即释放出来,从此转录进入延伸阶段。
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
3 转录
3.1 转录的要点 3.2 原核生物的转录 3.3 真核生物的转录 3.4 RNA 的剪辑与加工
ppt课件
1
中心法则
R everse tra n scrip tio n D N A syn th esis
ppt课件
2
3.1 转录的纲要
ppt课件
3
关键词
•转录(transcription) :拷贝出一条与DNA链序列完全相同 的RNA单链的过程。
•有义链 sense strand (编码链coding strand)与mRNA序列 相同的那条DNA链。
•反义链 antisense strand (模板链Template strand) 有义链的 互补链,根据碱基配对原则指导mRNA合成的DNA链。
ppt课件
4
关键词
•启动子(Promoter) RNA聚合 酶识别和结合并启动转录的 DNA序列.
随着RNA 链的延伸,转录泡与核心酶一起沿着DNA模板链同步移
动。释放出来的σ因子可以重新与核心酶结合,启动新一轮DNA
的转录,称为RNA在转录泡前面解链, 同时在转录泡后面重新形成双链,拓扑异构酶 能够在转录泡的前方解除因解链形成的正超螺 旋,在转录泡的后方解除因解链形成的负超螺 旋。
ppt课件
16
启动子有三个部分
1) -10区, 又称 Pribnow Box 保守序列为T80A95T 45A60A50T96 。在RNA聚合酶的作用下,富含
AT的Pribnow框内的DNA双螺旋首先发生解链,与RNA聚合酶形成 开放性启动子复合物。(下标表示碱基在不同基因的启动子中出现
的概率 )
与模板DNA的结合部位。
ppt课件
10
sigma 亚基只在转录的起始阶段起作用
Sigma因子 是酶的别构效应物, 提高RNA聚合酶对启动子的亲和 力,降低与模板DNA上非特异位 点的结合力。
大肠杆菌中有多个因子, 每个 因子能识别不同的启动子。
ppt课件
11
E.coli的σ因子由基因rpo D编码,其主要作用是特异
ppt课件
23
3.2.4 转录延伸
RNA链延伸的过程
一旦RNA pol合成了大约10nt左右的RNA链,延伸便开始了。这
时转录物的长度足以让RNA取代σ因子的位置,使RNA pol的核心
酶与σ因子解离。核心酶因此而可以离开启动子,沿模板链移动
了,这一过程称启动子清空(promoter clearance)。在σ因子被释
ppt课件
6
3.2 原核生物的转录
3.2.1 RNA 聚合酶 3.2.2 启动子的组成 3.2.3 转录的起始和延伸 3.2.4 大肠杆菌中两种终止子
ppt课件
7
3.2.1 RNA聚合酶
RNA聚合酶核心酶: 2
RNA聚合酶全酶: 2
RNA聚合酶各亚基的 功能(见左图)
ppt课件
8
ppt课件
9
细菌RNA pol的核心酶含两个相同的α亚基(由rpoA
放以后,延伸因子Nus-A蛋白加入进来,转录即进入延伸阶段。
失去σ因子的核心酶通过封闭的钳子握住DNA,以更快的速度沿
着DNA模板链移动,使延伸反应可以持续进行。RNA pol的结构
变化使延伸中的RNA 链从RNA/DNA杂交双链中脱离,单链DNA重
新与互补链配对,转录泡的大小维持在17 bp左右。也就是说,
•封闭复合物: 全酶 ·DNA 双链体
•开放复合物: 全酶 ·开链 DNA 双链体
• 三元复合物 (Ternary complex): 全酶 ·开 链 DNA 双链体·新生 RNA (up to 9 nucleotides)
•当新生 RNA达到8-9个核苷酸长度时 , 因子 从RNA聚合酶上脱离下来,形成延伸三元复 合物,包括核心酶·DNA ·新生 RNA.
ppt课件
25
ppt课件
26
延伸的暂停和阻滞
在延伸阶段,RNA pol每催化1个新的磷酸二酯键 形成,就面临3种选择,其一是继续延伸合成新的磷 酸二酯键,其二是倒退切除新掺入的核苷酸,其三是 延伸复合物解离,完全停止转录。 (1) 暂停 若 在转录过程中,转录产物形成了特定的二级结构,如 发夹结构,或NTP暂时短缺,均有可能造成转录暂停。 暂停有可能使原核生物转录和翻译同步,也有可能对 转录调节蛋白发挥作用有帮助,还有可能是转录倒退 或完全终止的前奏。RNA合成的重新启动需要GreA和 GreB蛋白来解除暂停状态,在RNA pol倒退后,GreA 和GreB切除3'-端几个核苷酸,以便让RNA的3'-OH能 重新回到活性中心。
ppt课件
19
(1) RNA pol全酶与双链DNA的非特异性结合
核心酶与σ因子结合成全酶,RNA pol全 酶与非特异性DNA序列具有一定的亲和性, 但亲和性较低。然而,一旦聚合酶与DNA结 合,即可沿DNA滑动扫描,直到发现启动子 序列。RNA pol与启动子的结合则是特异性 的,具有很高的亲和性。
性地识别转录的起始位点启动子。启动子(promoter) 是RNA聚合酶识别、结合并起始转录的一段特异性 的DNA序列,一般位于转录起始位点(+1)的上游, 可以同RNA聚合酶特异性结合,但本身的序列不被 转录。核心酶虽可与DNA结合,但不能区别启动子 和一般的DNA序列,或者说不能正确识别启动子的 特异序列。σ因子与核心酶结合后,全酶对启动子的 特异性结合能力是对其他DNA序列结合能力的107倍。 其原因是,核心酶对DNA序列结合的亲和力建立在 碱性蛋白和酸性核酸之间静电吸附作用力之上,这 种结合是没有特异性的松散结合,而且被结合的 DNA仍然保持双螺旋状态。σ因子能大大降低RNA pol与一般DNA序列的结合常数和停留时间,同时又 增加RNA pol与启动子的结合常数和停留时间,使 RNA pol能正确的识别启动子的特异序列,并与之结 合。
•转录起始点 (startsite) 与新生 RNA链第一个核苷酸相对应 DNA链上的碱基,通常为一个 嘌呤
•终止子(terminator) DNA上引起 RNA聚合酶终止转录的序列.
ppt课件 Transcription unit 5
RNA的酶促合成
rNTP n rNTP
DNA template RNA-P, Mg2
ppt课件
21
(3) 封闭复合物转化成开放复合物
σ因子使DNA部分解链,形成大小为12 bp~17 bp的转录泡,使 DNA模板链进入活性 中心,封闭复合物转变成开放复合物(open complex)。一开始,转录泡覆盖–10~–1,但 它很快以一种依赖于Mg 2+的方式从–12延伸 到+2。开放复合物十分稳定,其半寿期在几 个小时以上,此时的聚合酶与启动子的相互 作用既有静电引力,又有氢键。足印法测定 表明,在此阶段聚合酶复盖–55~+20区域。
ppt课件
20
(2) RNA pol与启动子形成封闭复合物
一旦RNA pol遇到–35区,便形成封闭复 合物(closed complex)。在此阶段,DNA并没 有解链,聚合酶主要以静电引力与DNA结合。 这种复合物并不十分稳定,半衰期为15min~ 20min。足印法分析表明,在此阶段聚合酶复 盖–55到+5区域。
ppt课件
22
(4) RNA合成的起始
在前两个与模板链互补的NTP从次级通道进入聚 合酶的活性中心以后,由活性中心催化第一个NTP的 3'-OH亲核进攻第二个NTP的5'-磷酸基,形成第一个 磷酸二酯键。第一个掺入的核苷酸总是嘌呤核苷酸, 这是因为聚合酶的第一个NTP结合位点优先结合嘌呤 核苷三磷酸,而第二个NTP结合位点与4种NTP结合的 亲和力相同。此外,新合成的初级转录物一般含有 5'-三磷酸,这与成熟的RNA分子不同。一旦有了第 一个磷酸二酯键,RNA-DNA-RNA pol的三元复合物 (the ternary complex)就形成了。RNA pol全酶催 化形成6个~10个磷酸二酯键以后,与核心酶结合 的σ因子即释放出来,从此转录进入延伸阶段。