抗体及基因重排
抗体

mIgM
mIgD
未成熟B细胞
成熟B细胞
活化或记忆B细胞
5)IgE;含量少/寿命断/热敏感/亲细胞/进化 最晚/含量最低/与超敏反应/寄生虫免疫有关
五、抗体的编码基因
抗体的胚系基因(germline genes)
重链:V基因片段(variable segment)
D基因片段(diversity segment) J基因片段(joining segment)
1、基本结构 2、辅助结构 3、结构域(Domain)
1、基本结构
1)四肽链通过链间二硫键组成H2L2
重链:450aa, Mr.5X104,五类(a、g、m、d、e) 轻链:214aa, Mr.2.5X104 ,两型(k、l) 2)分三个功能区: 可变区(Variable region, VH/VL, Fv):结合抗原 恒定区(Constant region, CH/CL):次级反应、抗原性 绞链区(Hinge region)CH1-CH2间, 30aa,2-5链间二硫键
3)蛋白酶可酶解
木瓜蛋白酶(papain):Fab、 Fc
胃蛋白酶(pepsin):
F(ab’)2 →2XFab’、pFc’
2、辅助结构
1)J链(Joining chain):
20KD、10%糖、Cys、二硫键连接IgH
2)分泌片(Secretory piece,SP):
70KD、6%糖、上皮细胞合成
抗体的特异性和类型的不同,称为异质性 (heterogeneity )
不同抗原表位刺激机体所产生的免疫球蛋白分子重链类别和轻链型别不 同
不同抗原表位刺激机体所产生的免疫球蛋白分子识别抗原的特异性不同
不同抗原表位诱导的同一类型的免疫球蛋白(IgG),其识别抗原的特 异性不同
抗体、基因重排及B细胞发育

抗体结构
膜 型 IgM
sIgM
J链
Iga Igb
Iga Igb
IgE 分 子 的 结 构
IgD 分 子 的 结 构
IgA dimer
分泌链 J链
抗体类别 重链类别 重链亚类 轻链类别 分子式
分子量(kDa) 固定补体能力 血清浓度 (mg/100ml) 血清半衰期(天) 穿过胎盘 肥大细胞和嗜碱 性细胞脱颗粒 裂解细菌能力 抗病毒能力
表位
抗原
抗体 CDR
Ag
“Ig-like Domain (Ig样结构域)”
Ig Superfamily
Ig分子超家族的成 员多数为细胞膜表 面分子。图中显示 的是Ig分子超家族 的一些主要成员及 其分子结构。
免疫球蛋白超家族
免疫球蛋白 (IgM)
-S-S-S-S-
Iga/b
-S-S-
-S-S-S-S-
50 40
30
CDR1
CDR2
20
10
抗体分子的多样性
CDR3
0 0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 210 220
抗体分子的抗原结合部位
HV4
NH2
CDR1 CDR3
CDR2
COOH
B 细胞表位
3’
L VH1 L VH201 L VHn
5’
DH1 DH DH JH Cµ Cd Cg 3 Cg 1 Cg2b Cg2a Ce Ca
Adaptive Immunity
Three Representatives MHC
Antibodies TCRs
抗体重排rearrangement

The Nobel Prize in Physiology or Medicine 1987 “For his discovery of the genetic principle for generation of antibody diversity"
Massachusetts Institute of Technology (MIT) Cambridge, MA, USA
因为微生物生长速度比脊椎动物快许多倍因此微生物产生突变体的速度可以很快如果免疫球蛋白仅是由胚系基因编码的那么脊椎动物就无法对付抗原飞速的变化体细胞多样性使得机体不受限制地产生抗体
淋巴细胞抗原识别受体的编码基因 及
受体多样性的产生
淋巴细胞抗原识别特点:
1. TB细胞都用其抗原识别受体识别抗原
B细胞:B cell receptor (BCR) T细胞:T cell receptor (TCR)
问题: 为什么人产生的抗体比他自身携带
的总的基因数还要多?
内容: 一、BCR重排的发现及BCR胚系基因结构 二、产生BCR多样性的机制 三、BCR基因表达的一些特点 四、TCR的基因结构和多样性的产生
一、 BCR重排的发现及其胚系基因结构
BCR或抗体由可变区和恒定区组成,其多样性是 由可变区决定的。
可变区多样性是怎样产生的?
抗体多样性的两种假说:
体细胞突变假说
B细胞发育过程中发生了突变,导致生成的BCR或者抗 体变成许多种
基因片段重排假说 ?
Dreyer和Bennett于1965年提出,即两个基因一条多 肽链理论。他们认为BCR由两类基因片段组成,一类编 码BCR的V区,一类编码BCR的C区。V区基因含有多个基 因片段,只有其中一个片段与C区形成抗体。
人类抗体重排机制的起源

人类抗体重排机制的起源
人类抗体重排机制的起源
人类抗体重排机制(简称V(D)J重排)是一种典型的遗传变异机制,它可以使B淋巴细胞发挥出多样性,从而使人体能够对多种抗原(如病毒、细菌、细胞外抗原)作出应答。
V(D)J重排机制起源于数十亿年前,当时地球上原始的生物正处在一个激烈的竞争当中,要想在这种环境中存活,就必须要不断的进化以保持适应性。
而V(D)J重排机制正是一种可以让细胞能够快速进化,从而提供生物体抵御外界病原体的重要武器之一。
V(D)J重排机制的具体机制是,人体的B淋巴细胞分子表面上都存在着一种特殊的受体——名为抗原受体(Antigen Receptor)的蛋白质,它由两种不同的结构片段组成——称为V(variable)和J(joining)的抗体受体基因片段,它们携带着不同的抗原识别特异性。
当B淋巴细胞受到病原体攻击时,V(D)J重排机制就会被激活,抗原受体基因片段会重新组合,从而创造出多种不同的受体,这些受体就可以识别到不同的外来抗原,从而使B淋巴细胞可以更快速、更有效率地对抗外来抗原。
不仅如此,V(D)J重排机制还可以提供人体抵御多种抗原的能力,因为在重排过程中,抗体基因片段可以经历不同的组合,从而创造出各种不同的抗原受体,使得人体拥有抵御多种抗原的能力。
此外,V(D)J重排机制还可以使抗体受体不断变异,以更好地适应环境的变化,从而使人体更容易抵抗多种病原体,从而保护人类的健康。
V(D)J重排机制的起源源于数十亿年前,它是一种可以使B淋巴细胞发挥多样性的重要机制,它可以使人体拥有抵御多种抗原的能力,也可以使抗体受体不断变异,以更好地适应环境的变化,从而保护人类的健康。
基因重排名词解释

基因重排名词解释
基因重排(Gene Rearrangement)是指在某些生物体的基因组中,由于基因片段的移动、插入、删除或倒位等现象,导致基因在DNA分子上的排列顺序发生改变。
基因重排是基因组进化中的一个重要机制,可以增加基因组的多样性和复杂度,同时也可能对生物体的适应性和健康产生影响。
下面是一些与基因重排相关的名词解释:
1. 基因片段(Gene Segment):指编码蛋白质的DNA序列中的一段区域,通常具有特定的功能和序列特征。
2. 重组酶(Recombinase):一类催化基因重排的酶,能够切割DNA链并促使基因片段的互相重组。
3. V(D)J重排(V(D)J Rearrangement):在脊椎动物免疫系统中,负责生成抗体和T细胞受体的基因由不同的V、D和J基因片段
组成,通过V(D)J重排来产生不同的抗原识别结构,以应对不同的病原体。
4. 染色体易位(Chromosome Translocation):指两个染色体之间的基因重排,通常发生在染色体的两端或中间,可能影响基因的表达和功能。
5. 基因分化(Gene Divergence):由于基因重排等机制,在不
同物种或个体之间,基因片段序列和组合会发生变化,导致基因的结构和功能差异增加,称为基因分化。
抗体基因重排原理

抗体基因重排原理咱先得知道啥是抗体。
抗体就像是咱们身体里的小卫士,专门去识别那些外来的坏家伙,像细菌啊、病毒啊之类的。
那这些抗体是怎么来的呢?这就和基因重排有大大的关系喽。
在咱们的细胞里啊,抗体基因可不是一开始就整整齐齐地排好队准备工作的。
它们就像一堆乱七八糟的小积木块。
对于抗体来说,有好几个不同的基因片段,就比如说有V(可变区)片段、D(多样性区,这个不是每个抗体都有的哦)片段还有J(连接区)片段。
这些片段就散落在基因的各个角落。
当一个B淋巴细胞(这可是产生抗体的小工厂细胞呢)开始准备制造抗体的时候,就像是一场超级大抽奖。
这个细胞会从众多的V片段里挑出一个,然后呢,如果有D片段的话,再从D片段里选一个,最后从J片段里挑一个。
这一挑啊,那可就是随机的,就像你从好多不同颜色的糖果里乱抓一把一样。
选好这些片段之后呢,它们就要凑在一起啦。
这就像是把不同的小零件组装成一个超级武器。
细胞里有专门的酶来把这些片段连接起来,这个过程就像是小工匠在精心打造一件独一无二的宝贝。
一旦这些片段连接好了,就形成了抗体的可变区。
这个可变区可不得了,它就像是一把专门为识别特定敌人打造的钥匙。
为啥要这么复杂地重排基因来制造抗体呢?这就是大自然的智慧啦。
因为外界的细菌和病毒那可是千奇百怪的,如果抗体都是固定不变的,那肯定应付不过来呀。
通过这种基因重排,就可以产生数不清的不同抗体。
就好比你有了无数把不同的钥匙,总能找到一把去打开那些入侵坏家伙的锁。
而且哦,这个基因重排还不是一次性的。
在B淋巴细胞的发育过程中,还可能会发生一些小变化,让抗体变得更加多样化。
这就像是给小卫士们不断升级装备一样。
你看,咱们身体里的这个小世界多神奇呀。
这些微观的小基因,就像一群小精灵,通过这种奇妙的重排方式,为咱们的健康打造出强大的防御体系。
每次想到这个,我都觉得大自然真的是一个超级有创意的大艺术家呢。
它用这种看似有点“调皮”的方式,让我们的身体有了应对各种危险的能力。
抗体(Antibody,Ab)基本结构和作用

IgD在B细胞膜上出现,是B细胞成熟的 标志。这些B细胞都难以产生免疫耐受 性,而B细胞膜上只有IgM而无IgD时, 容易因相应抗原作用而形成免疫耐受。
五类免疫球蛋白特性比较
Ig种类
IgG
IgM
IgA/SIgA IgD IgE
IgA不能激活补体的经典途径, 但能激活补体的替代途径。不能作 用为调理素,但能凝集颗粒性抗原 和中和病毒。
SIgA
分泌型IgA是机体 粘膜防御感染的 重要因素。
在保护肠道、泌 尿生殖道、乳腺 和眼睛抵抗微生 物入侵方面起关 键作用。
4、IgE
IgE为单体分子; 分子量190kDa,在血清中含量最少,约为
IgM和SIgA结构示意图
(二) 功能区
用X射线衍射分析法发现,Ig多肽链是由若干折叠 成球形结构组成的一种立体构型。 每一球形结构是肽链的一个亚单位,约110个氨基 酸组成,具有一定的生理功能,故称功能区(或结构域)。 在功能区中氨基酸序列有高度同源性。 L链有VL、CL两个功能区; IgG、IgA、IgD的重链有VH、CH1、CH2、CH3共四个功 能区;IgM、IgE重链则有VH、CH1、CH2、CH3、CH4五个功 能区。
0.01~0.9%;
种系进化中出现最 晚;
由呼吸道和胃肠道 浆细胞产生;
IgE
人和动物血清中浓度 很低,但它有独特的 Fc区,能结合肥大细 胞和嗜碱性细胞,可 引起I型超敏反应。
IgE还与对蠕虫侵袭的 免疫应答有关。
IgE在56℃ 30min即被
5、IgD
血清中含量极少,为Ig总量的1%左右。 IgD为单体分子,血清中IgD的功能尚 不清楚。
免疫球蛋白的基因重排

免疫球蛋白的基因重排摘要:I.免疫球蛋白概述- 免疫球蛋白的概念- 免疫球蛋白的功能II.免疫球蛋白的基因重排- 基因重排的定义- 免疫球蛋白基因重排的过程- 免疫球蛋白基因重排的意义III.免疫球蛋白基因重排与疾病的关系- 免疫球蛋白基因重排与免疫疾病的关系- 免疫球蛋白基因重排与癌症的关系IV.研究免疫球蛋白基因重排的意义- 治疗免疫疾病- 预防癌症正文:免疫球蛋白,也称为抗体,是人体免疫系统的重要组成部分。
它能够识别并结合病原体,协助免疫系统清除入侵者。
然而,免疫球蛋白的功能不仅仅局限于防御,它还能调节免疫反应,参与免疫细胞的信号传导等。
免疫球蛋白的产生是由基因重排过程完成的。
基因重排是指在基因水平上,将不同基因片段进行重组,产生新的基因组合。
在免疫球蛋白的产生过程中,B 淋巴细胞通过基因重排,产生具有独特抗原结合位点的免疫球蛋白。
这一过程使得人体能够产生大量的抗体,应对各种病原体的入侵。
免疫球蛋白基因重排的意义重大。
首先,基因重排产生了大量的抗体,提高了免疫系统对病原体的识别和清除能力。
其次,基因重排使得免疫系统能够针对不同的病原体产生特异性的抗体,避免了不必要的免疫反应。
然而,免疫球蛋白基因重排过程也存在一定的风险。
基因重排的异常可能导致某些癌症的发生,如滤泡性淋巴瘤和套细胞淋巴瘤等。
近年来,随着研究的深入,人们逐渐认识到免疫球蛋白基因重排与许多疾病的发生和发展密切相关。
例如,免疫球蛋白基因重排异常可能导致免疫系统过度激活,从而引发自身免疫性疾病,如类风湿性关节炎、红斑狼疮等。
此外,免疫球蛋白基因重排异常还与癌症的发生密切相关。
因此,研究免疫球蛋白基因重排对于治疗免疫疾病和预防癌症具有重要意义。
通过深入研究免疫球蛋白基因重排的机制,科学家们可以寻找新的治疗方法,提高免疫系统的功能,预防疾病的发生。
抗体测序精确度—抗体多样性及分子基础

抗体多样性及分子基础抗体多样性来源抗体的多样性来源主要有4个,分别是:1、VH, DH 和JH三条链重组成有功能的重链,VL 和VJ (λor κ) 两条链组成有功能的轻链,轻链要么λ型,要么是κ型,二者不会同时出现在同一个抗体上。
2、VDJ基因拼接时,接头的多样性,这个多样性的主要有4种产生方式:1)D基因可以已任一方向在三个开放阅读框中的任何一个中翻译,以产生总共六个可能的肽片段。
2)在重排过程中会形成发卡结构,导致氮核苷酸的增加或者减少,从而增加多样性。
3)VDJ连接机制,可能添加或去除氮核苷酸作为VDJ连接过程的一部分。
有时,几个氨基酸残基的编码序列可能在重组过程中丢失。
4)可以通过末端脱氧核苷酸转移酶(TdT)的活性来添加或替换氮核苷酸,特别是在构成功能性V区的CDR-H3的VDJ连接的D片段的任一侧上。
据估计,这些氮核苷酸的变化可导致CDR-H3的多样性大于107,包括CDR长度范围从仅仅几个氨基酸残基到超过25个氨基酸残基。
3、体细胞高频突变。
4、类别转换和基因转换。
人类抗体基因组成配系细胞中含有如下抗体基因1、C基因片段(constant gene segment)其编码抗体轻链和重链C区;2、轻链V基因片段(variable gene segment)和J基因片段(joining segment),二者编码轻链可变区;3、重链V、D基因片段和C基因片段(diversity segment),他们编码重链可变区。
上述基因片段位于不同染色体上,呈不连续成簇分布。
如图1所示,人抗体基因中总共有170-176个IGH(Immunoglobulin heavy chain:免疫球蛋白重链)基因,其中有76-84个是有功能的,包括38-46个IGHV基因(重链V基因片段)、23个IGHD基因(重链D基因片段)和9个IGHC基因(重链C基因片段)。
编码k轻链的基因座位于2号染色体的短壁上,k基因座一共有82个基因,由76个IGKV(k 轻链V基因片段)基因和5个IGKJ(k轻链J基因片段)基因组成,大概只有31-36个IGKV 基因参与形成有功能的V k(k型抗体的重链)。
多克隆抗体 基因重排

多克隆抗体是由B淋巴细胞分泌的一种抗体分子,它们能够识别并结合多种抗原。
多克隆抗体的制备通常需要使用小鼠等动物进行免疫,然后从免疫动物的脾脏等组织中提取出B淋巴细胞,再通过一系列的细胞培养和筛选步骤来获得多克隆抗体。
基因重排是指在B淋巴细胞中发生的一种基因重组过程。
在B淋巴细胞的发育过程中,它们会经历一系列的基因重排,从而产生出具有不同抗原识别能力的抗体分子。
基因重排是通过重排基因的染色体位置来实现的,这一过程需要一系列的基因重组酶的参与。
在多克隆抗体的生产过程中,基因重排是非常重要的一个步骤,因为它能够产生出具有不同可变区的抗体分子,从而增加多克隆抗体的抗原识别能力和多样性。
抗体种类千千万,你知道原因么?

抗体种类千千万,你知道原因么?人类生活在复杂多变的环境中,每时每刻都会接触到各种各样的微生物,受到一些类似抗原物质的侵扰,从而使机体致病。
为了抵御这些外来侵扰,使自身得以继续生存,人体必须形成几十万、几百万甚至更多种相应的特异性抗体以抵抗外界的抗原物质,才能免遭其害,保护自己。
我们会从抗体的产生及多样性进行其原因的阐述与分析。
1.抗原的呈递抗原呈递细胞(antigen presenting cell, APC)的抗原呈递作用是一个涉及抗原摄取、处理与呈递的复杂过程[1]。
最主要的抗原呈递分子是主要组织相容性复合物(majorhistocompatibility complex, MHC)[2]。
MHC分为两个大类:MHC-I和MHC-II,它们呈递的抗原蛋白来源不同,降解抗原的方式不同,结合肽段长度也不同,可以分别将抗原呈递给细胞毒性T细胞 (CD8 T cell) 和辅助型T细胞 (CD4 T cell)[3,4]。
结合现以MHC-II呈递外源性蛋白为例简述抗原提成过程。
外源性抗原经APC吞噬或吞饮作用,被摄入胞内形成吞噬体,后者与溶酶体融合形成吞噬溶酶体。
抗原在吞噬溶酶体内酸性环境中被蛋白水解酶降解为小分子多肽,其中具有免疫原性的称为抗原肽[3,5]。
内质网中合成的MHC-II类分子进入高尔基体后,由分泌小泡携带,通过与吞噬溶酶体融合,使抗原肽与小泡内MHC-Ⅱ类分子结合形成抗原肽-MHCⅡ类分子复合物。
所形成的复合物可能在高尔基复合体参与下被转运到细胞膜表面,被T细胞受体(TCR)识别并呈递给TH细胞[6]。
供TCR识别的先决条件是两种细胞的直接接触并相互作用。
这种细胞间的相互作用涉及APC与TH表面多种分子。
除了TCR特异性地同时识别多肽-MHC-Ⅱ分子的复合物外,某些粘附分子也参与抗原呈递过程。
活化的TH细胞可分泌各种细胞因子,用于B细胞、Tc细胞等的激活过程,产生可清除抗原的特异性抗体。
rag小鼠 品系特征

RAG小鼠,全称为重组激活基因(Recombination-Activation Gene)小鼠,是一类通过基因工程方法构建的小鼠品系。
它们的特点是能够在体内或体外有效地重组抗体基因,从而产生特定类型的抗体。
RAG小鼠通常用于免疫学和分子生物学领域的研究,特别是在研究抗体基因的重组和表达、免疫系统发育和功能等方面具有重要价值。
RAG小鼠的品系特征主要体现在以下几个方面:
1. 抗体基因重排能力:RAG小鼠具有重组激活基因,能够促进抗体基因的重排,产生多样化的抗体。
这使得它们在研究抗体结构和功能、以及免疫应答机制方面具有优势。
2. 免疫缺陷:某些RAG小鼠品系可能因为基因改造而具有免疫缺陷,这使得它们在研究免疫系统的调控和病原体感染机制时更为便利。
3. 遗传背景:RAG小鼠的遗传背景通常是C57BL/6或BALB/c,这两种品系因其稳定的遗传背景和良好的繁殖性能,被广泛用于实验研究。
4. 基因敲除能力:通过Cre-loxP或其他基因敲除技术,RAG小鼠可以用于创建特定基因被敲除的模型,这对于研究基因功能和疾病机理非常有用。
5. 自发突变率:由于RAG小鼠的基因改造性质,它们的自发突变率可能较高,这为研究基因变异和突变效应提供了资源。
6. 实验应用:RAG小鼠在科研中的应用范围广泛,包括但不限于抗体工程、基因敲除、免疫学研究、肿瘤学研究等领域。
在使用RAG小鼠进行实验研究时,研究人员需要考虑到这些品系的特定特征,以及可能对实验结果产生的影响。
此外,由于RAG小鼠的基因改造性质,研究人员在实验设计和数据分析时需要特别注意基因敲除或插入可能带来的非特异性影响。
抗体的结构与功能

抗体的产生:由B细胞在抗原刺激下产生 抗体的种类:IgG、IgM、Ig、IgE、IgD等 抗体的调节:由T细胞和B细胞相互作用调节 抗体的寿命:短寿命抗体和长寿命抗体 抗体的效应:中和作用、调理作用、补体结合作用等
自身免疫性疾病: 如系统性红斑狼疮、 类风湿性关节炎等
过敏性疾病:如 哮喘、过敏性鼻 炎等
抗体在生物技术中的应用:用于基因工程、细胞工程、酶工程等生物技术的研究与开发
抗体药物的研发:通过基因工程技术 将抗体基因导入到细菌、酵母或哺乳 动物细胞中进行大规模生产
抗体药物的前景:随着基因工程技 术的不断发展抗体药物的研发和应 用将更加广泛和深入
添加标题
添加标题
添加标题
添加标题
抗体药物的应用:用于治疗癌症、 自身免疫性疾病、感染性疾病等
抗体与自身免疫疾病相关可能 导致自身免疫性疾病
参与免疫调节:抗体可以调节免疫 反应防止免疫系统过度反应
参与细胞分化和增殖:抗体可以影 响细胞的分化和增殖影响细胞的生 长和发育
添加标题
添加标题
添加标题
添加标题
参与细胞信号传导:抗体可以与细 胞表面的受体结合传递信号影响细 胞功能
参与细胞凋亡:抗体可以诱导细胞 凋亡清除受损或异常细胞维持机体 健康
抗体的产生和调节
抗原刺激:抗原进入机体刺激免疫系统产生抗体 B细胞活化:抗原刺激B细胞使其活化并开始产生抗体 抗体生成:B细胞通过基因重排和表达生成特异性抗体 抗体分泌:抗体通过B细胞的分泌途径释放到血液中 抗体调节:免疫系统通过反馈机制调节抗体的产生和分泌
亲和力成熟:抗体与抗原结合的能力逐渐增强的过程 类别转换:抗体从一种类别转变为另一种类别的过程 免疫球蛋白重链和轻链的基因重排:抗体产生的基础 抗体的亲和力成熟和类别转换的机制:免疫系统对病原体的识别和反应
三、基因重排的分子机制
三、基因重排的分子机制
早在50年代,人们就认识到抗体分子的每一条链都是由高度多变的V区和相对不变的C区组成的,V区赋予抗体分子对抗原的特异性。
抗体分子V区的多样性和C区的稳定性显然是矛盾的。
Dreyer和Bennett 于1965年首次提出假设,认为每条抗体链实际上至少由两个基因所编码,其中一个是恒定的,一个是可变的。
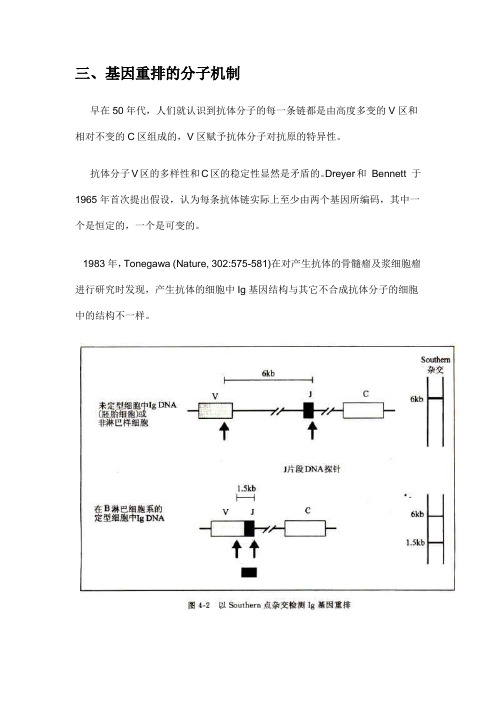
1983年,Tonegawa (Nature, 302:575-581)在对产生抗体的骨髓瘤及浆细胞瘤进行研究时发现,产生抗体的细胞中Ig基因结构与其它不合成抗体分子的细胞中的结构不一样。
在所有物种中,胚系Ig基因的构成基本上相同。
Ig重链和轻链(λ和κ链)基因座都由多个编码V区和C区蛋白质的基因组成,并被非编码的DNA所分隔。
抗体分子由4条(两对)多肽链组成,包括两条相同的轻链(L-chain)和两条相同的重链(H-chain)。
轻链和重链在相对分子质量上有较大差别,前者约2.3x104,后者则介于5.3x104-7.0x104之间。
所有Ig分子都含有两类轻链中的一类,即κ型或λ型。
Κ型和λ型轻链的恒定区和可变区的氨基酸序列是不同的。
在小鼠中,95%的抗体轻链是κ型,而人类抗体轻链中,κ型和λ型各占50%左右。
免疫球蛋白重链基因DNA重排以后,大量间隔序列被切除,使位于J-Cμ之间的增强子序列得以发挥作用,增强基因转录。
基因组重排在细胞增殖和疾病中的作用

基因组重排在细胞增殖和疾病中的作用基因组重排是指染色体内部的一些片段在不同染色体之间或同一染色体上发生了移动、插入或删除等事件,导致基因组的结构发生改变。
这种结构变化是细胞增殖和发育的重要因素之一,在疾病的发生和发展中也起着重要作用。
一、基因组重排在细胞增殖中的作用1.增加基因组多样性基因组重排可以在基因组中创造新的基因型,从而使新兴物种具有某种适应性和竞争的优势。
例如在哺乳动物的免疫系统中,基因组重排使得B细胞和T细胞能够产生成千上万种抗体和T细胞受体,以抵御不同的病原体,保障身体免疫系统的稳定和健康。
2.帮助细胞适应环境基因组重排可以导致基因的拆分、移动和复制,从而调节细胞中的信号转导途径和基因表达水平,使细胞能够适应环境的变化。
肿瘤细胞中的基因组重排事件就是这样一种现象,它可以在肿瘤细胞生长和扩散过程中发挥重要作用。
3.促进基因家族的扩张和演化基因组重排还可以促进某些基因家族的扩张和演化。
例如在智人的基因组中,ALU元件是重要的基因重排产物,它可以在基因组中扩张并产生新的功能序列,在人类智力和行为等方面扮演重要角色。
二、基因组重排在疾病中的作用1.发生突变或失活基因组重排可以导致染色体中部分区域的基因突变或失活,从而导致某些疾病的发生。
例如在新生儿中,缺少染色体的一部分或染色体上的某个基因被删除可能会导致儿童发生先天性缺陷的风险增加。
2.导致癌症发生基因组重排是肿瘤细胞中的常见现象,特别是恶性肿瘤。
它可以导致肿瘤抑制基因的缺失、癌症相关基因的激活等突变事件,而这些变化在一定程度上促进或加速肿瘤细胞的增殖和扩散。
3.影响药物代谢基因组重排可以影响药物代谢时酶的表达或功能,从而导致药物代谢产物积累或者药物作用失灵。
例如,在一些亚洲人种中,CYP2D6重复竞争和插入等基因重排事件会增加这一基因的拷贝数,大幅度地影响肝脏中药物代谢酶的功能和药物代谢产物的积累。
4.改变基因表达水平基因组重排还可以改变基因的表达模式和水平,从而有可能产生疾病。
抗体小知识

科技名词定义中文名称:抗体英文名称:antibody;Ab定义1:能与相应抗原(表位)特异性结合的具有免疫功能的球蛋白。
应用学科:免疫学(一级学科);免疫系统(二级学科);免疫分子(三级学科)定义2:在人和动物体内,由于抗原或半抗原入侵刺激机体而在细胞中产生的免疫球蛋白。
能可逆、非共价、特异地与相应抗原结合,形成抗原-抗体复合体。
应用学科:生物化学与分子生物学(一级学科);总论(二级学科)定义3:机体内B细胞在抗原刺激下所产生的具特异性免疫功能的球蛋白。
应用学科:水产学(一级学科);水产生物病害及防治(二级学科)定义4:具有抗原结合部位,能与抗原分子上相应表位发生特异性结合的具有免疫功能的球蛋白。
应用学科:细胞生物学(一级学科);细胞免疫(二级学科)本内容由全国科学技术名词审定委员会审定公布求助编辑百科名片Y型抗体结构示意图抗体(antibody)指机体的免疫系统在抗原刺激下,由B淋巴细胞或记忆细胞增殖分化成的浆细胞所产生的、可与相应抗原发生特异性结合的免疫球蛋白。
主要分布在血清中,也分布于组织液及外分泌液中。
目录概念多克隆抗体的制备单克隆抗体的制备概述生物活性抗体结构抗体基因重排单克隆抗体抗体的多样性抗体的功能抗体规律抗体的分类按作用对象按理化性质和生物学功能按可见反应按抗体来源世界著名抗体公司Santa公司Abcam公司Abgent公司AbMARTProteintech GroupMBLCell Signaling Technology(CST) Zymed Laboratories公司台湾AbnovaChemiconPierce公司BDPeproTechCaymanAssay DesignDynal Biotech ASA.DSL公司罗氏公司(Roche)BioLegend公司eBioscience公司BioVision. Inc.MABTech电影基本信息上映日期剧情简介概念多克隆抗体的制备单克隆抗体的制备概述生物活性抗体结构抗体基因重排单克隆抗体抗体的多样性抗体的功能抗体规律抗体的分类按作用对象按理化性质和生物学功能按可见反应按抗体来源世界著名抗体公司Santa公司Abcam公司Abgent公司AbMARTProteintech GroupMBLCell Signaling Technology(CST) Zymed Laboratories公司台湾AbnovaChemiconPierce公司BDPeproTechCaymanAssay DesignDynal Biotech ASA.DSL公司罗氏公司(Roche)BioLegend公司eBioscience公司BioVision. Inc.MABTech电影基本信息上映日期剧情简介展开编辑本段概念最初有人用电泳证明血清中抗体活性在γ球蛋白部分,故曾把抗体统称为丙种(γ)球蛋白。
抗体重排

抗体多样性的产生—基因重排基因重排的分子机制早在50年代,人们就认识到抗体分子的每一条链都是由高度多变的V区和相对不变的C区组成的,V区赋予抗体分子对抗原的特异性。
抗体分子V区的多样性和C区的稳定性显然是矛盾的。
Dreyer 和Bennett 于1965年首次提出假设,认为每条抗体链实际上至少由两个基因所编码,其中一个是恒定的,一个是可变的。
1983年,Tonegawa (Nature, 302:575-581)在对产生抗体的骨髓瘤及浆细胞瘤进行研究时发现,产生抗体的细胞中Ig基因结构与其它不合成抗体分子的细胞中的结构不一样。
在所有物种中,胚系Ig基因的构成基本上相同。
Ig重链和轻链(λ和κ链)基因座都由多个编码V区和C区蛋白质的基因组成,并被非编码的DNA所分隔。
抗体分子由4条(两对)多肽链组成,包括两条相同的轻链(L-chain)和两条相同的重链(H-chain)。
轻链和重链在相对分子质量上有较大差别,前者约2.3x104,后者则介于5.3x104-7.0x104之间。
所有Ig 分子都含有两类轻链中的一类,即κ型或λ型。
Κ型和λ型轻链的恒定区和可变区的氨基酸序列是不同的。
在小鼠中,95%的抗体轻链是κ型,而人类抗体轻链中,κ型和λ型各占50%左右。
免疫球蛋白重链基因DNA重排以后,大量间隔序列被切除,使位于J-Cμ之间的增强子序列得以发挥作用,增强基因转录。
IgH基因重排基因重排与抗体多样性1、正常淋巴细胞在发育中是多克隆性质, 但恶性肿瘤表现为单克隆性基因重排。
如:t (14 18) (q32 q21) 是滤泡性淋巴瘤( FL) 中的一个常见的染色体易位,该易位导致bcl22 与IgH重排。
所以, 通过基因重排检测不仅可以鉴别淋巴组织是肿瘤性增生还是反应性增生,而且使准确判断细胞起源, 完善淋巴瘤的分型成为可能。
另外,多发性骨髓瘤(multiple myeloma, MM) 是一种以产生单克隆免疫球蛋白为特点的异常浆细胞恶性增殖性疾病。
抗体及基因重排

• IGH 95/51
23/23 9/6 11/9
• IGK 90/5005/5 1/1• IGL 60/310
4/4 7/7
2020/2/10
58
二、基因重排
VDJ两端为重排信号序列(7核苷酸序列) 12—23原则:带12-bp-RSS只与23bp-RSS结合
2020/2/10
59
基因片段两端RSS互相靠拢, 它们间DNA环被剪除,D,J连接
• 目前对IgD的生物学功能尚不明确 ,可能与超敏反应或自身免疫病有 关系。
•
2020/2/10
51
11.3 抗体分子的 Fc受体
2020/2/10
52
2020/2/10
53
2020/2/10
54
11.4 Ig分子超家族
IgSF类似IgV或C去折叠机构, 氨基酸与Ig一定同源性的分子。
2020/2/10
2020/2/10
42
• 由于IgM具有较多的补体结合位点,因 此具有较IgG更强大的激活补体经典途 径的能力,是抗感染、特别是抗早期 感染的主要抗体,当人体缺乏IgM可能 发生致死性败血症。
• 类风湿因子、冷凝集素、天然血型抗 体均为IgM类。
2020/2/10
43
三、IgA
2020/2/10
2020/2/10
8
轻链 轻链免疫原性的不同,分为κ、λ两型。
同一个体内可存在分别带有为κ和λ链 的Ig分子,同一天然Ig分子中不可能同时 有κ和λ链,两条轻链的型别必然相同。
9
二、可变区和恒定区
• 1.可变区 Ig近氨 基端轻链的1/2和 重链的1/4
(γ、α、δ)或 1/5(μ、ε).
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Figure 3-3
? 木瓜蛋白酶(papain)的水解部位位于Ig铰 链区重链间二硫键近N端处,水解后可得到 2个相同的抗原结合片段(fragment of antigen binding,Fab)和1个可结晶的片段 (fragment crystallizable,Fc)。
? Fab段具有抗体活性的部分,可单价与抗原 结合;
11.2 五类Ig的特性和功能 一、IgG
? IgG的功能和结构在五类Ig中研究得最为充 分。人类Ig共有4个亚类,即IgG1、IgG2、 IgG3和IgG4。
? IgG主要由脾脏和淋巴结中浆细胞合成,是 血清中含量最多(约占血清Ig总量的75%) 、半衰期最长(约23天)的Ig;
? 机体在出生后3个月就开始产生IgG,3~5 岁达到成人水平。
? 白喉或破伤风抗毒素经胃蛋白酶消化后精 制的制剂,由于去除了重链的Fc段可减少超 敏反应的发生。
Fi 1、J链 ? 连接链(joining chain,J链)是由浆细胞合
成的一条富含半胱氨酸的多肽链,可将单 体Ig连接为多聚体并使之稳定。如连接分泌 型IgA(secretory IgA,SIgA)成双体,连 接IgM成为五聚体。
IgG是五类Ig中 唯一可主动穿 过胎盘的Ig, 对防止新生儿 感染有重要作 用。
? 2.CL和CH1: 某些同种异 型的遗传标 记存在于该 区。
? 3.IgG的CH2和 IgM的CH3: 有补体C1q的结 合位点(CR1), 与补体经典途 径的激活有关 。
? IgG的CH2可通 过胎盘。
4.CH3或CH4:可与多种细胞表面 的相应的Fc受体结合,产生不同的 免疫效应。
溶细胞、细菌,调理作用,ADCC作用, 中和毒素,致超敏反应
,即重链的链间二 硫键连接处附近, 有一个可转动的铰 链区(hinge region )。
? 含较多的脯氨酸残 基,富于柔性,可 赋予较大的自由活 动度。该区由于是 一段较伸展的肽链 ,对蛋白酶敏感, 易被木瓜蛋白酶、 胃蛋白酶水解。
三、水解片段
? 在一定条件下,可通 过蛋白酶将Ig消化成 不同的片段,从而研 究Ig的结构和功能, 最常用的蛋白酶是木 瓜蛋白酶和胃蛋白酶 。
五、高变区(抗体分子立体结构)
? 重链和轻链V区( VH和VL)中各有 3个区域的氨基酸 组成和排列顺序变 化最为明显,称为 高变区或超变区( hypervariable region,HVR)。
★六、功能区的功能
? 1.VH、VL:是Ig分子特异性识别和结合抗 原的部位。
? 由于超变区的氨基酸组成和排列随所结合 的抗原特异性不同而不同,故其氨基酸的 种类和排列顺序千变万化,可形成很多能 与不同特异性抗原表位结合的抗体。
2.分泌片
? 分泌片(secretory piece,SP)又称分泌成 分(secretory componnent,SC),是由粘 膜上皮细胞合成含糖的肽链,以非共价形 式与IgA二聚体结合,使其成为分泌型IgA( SIgA)。其功能是保护SIgA免受外分泌液中 蛋白水解酶的水解作用,并能介导SIgA转运 ,是其分泌至粘膜表面,发挥粘膜免疫作 用。
抗体及基因重排
(Antibody,Ab)
遵义医学院 孙万邦
抗体( antibody,Ab)是B淋巴细 胞接受抗原刺激后增殖分化为浆细 胞,由浆细胞合成、分泌能与相应 抗原特异性结合的免疫球蛋白
(immunoglobulin,Ig)。
抗体主要存在于血清中,也存在于 各种体液中。
11.1 Ab(Ig)的基本结构及多样性
? Ig分子的基本结 构是由两条相同 的重链和两条相 同的轻链通过二 硫键连接而成的 四肽链结构,称 为Ig的单体。
? 一、重链和轻链 ? 1.重链(heavy chain ,H链) Ig 的重链由450~550 个氨基酸残基组成, 分子量约为50~75kD。
重链 根据重链免疫原性的不同,可将重链分为
? Fc段不能与抗原结合,保留了Ig重链的免疫 原性及其相应功能区的生物学活性。
Figure 3-3 part 1 of 2
胃蛋白酶水解片段
? 用胃蛋白酶(pepsin)可使Ig在重链间二硫 键近C端处断开,获得1个仍由二硫键连接 的F(ab′)2片段和若干小分子多肽碎片 pFc′。F(ab′)2是由两个Fab及铰链区 组成,可同时结合两个抗原表位。pFc′无 生物活性。
μ、γ、α、δ和ε五类(class), 与轻链结合为IgM、IgG、IgA、IgD和IgE。
某些又可亚类。γ分为四个亚类(γ1-4) ,α分为2个亚类(α1-21-2)。
2.轻链(light chain,L链) Ig的轻链约含 214个氨基酸, 约为重链的1/2, 分子量约25kD。
轻链 轻链免疫原性的不同,分为κ、λ两型。
? IgG抗病毒和细菌感染,中和毒素的重要抗 体,多数抗菌、抗病毒和抗毒素的抗体均 为IgG类。
? IgG的Fc段可与多种细胞表面的Fcγ R结合 ,发挥调理作用及ADCC效应。IgG的某些亚 类有亲细胞性,与I型超敏反应发生有关; 还可与葡萄球菌A蛋白(staphlococcus protein A,SPA)结合。某些自身抗体, 也多属IgG类。
同一个体内可存在分别带有为κ和λ链 的Ig分子,同一天然Ig分子中不可能同时 有κ和λ链,两条轻链的型别必然相同。
二、可变区和恒定区
? 1.可变区 Ig近氨 基端轻链的1/2和 重链的1/4
(γ、α、δ)或 1/5(μ、ε).
? 可变区
? 氨基酸排列顺序 在不同抗体分子 间变化较大称为 可变区( variable region ,V区)。
2.恒定区
? Ig分子近C端,轻链 的1/2和重链的3/4 (或4/5)区域中氨 基酸的数量、种类 、排列顺序及含糖 量均较为恒定,称 为恒定区( constant region,C 区)。
重链和轻链的恒 定区分别称为 CH和CL。 Ig的C区与抗体 的生物学功能相 关。
二、铰链区
? 铰链区 ? 在CH1和CH2之间