第三章植物的光合作用(2)
植物的光合作用ppt课件

叶绿体是由叶绿体被膜、基质和类囊体三部分组成
1.叶绿体 被膜
2.基质及 内含物
3.类囊体
高等植物的类囊体垛叠成基粒,其意义有 二:
膜的垛叠意味着捕 获光能机构的高度 密集,更有效地收 集光能,加速光反 应;
膜系统是酶的排列 支架,膜垛叠就犹 如形成一条长的代 谢传递带,使代谢 顺利进行。
(二)类囊体膜上的蛋白复合体
(3) 营养元素
➢ 叶绿素的形成必须有一定的营养元素。
氮和镁是叶绿素的组成成分,铁、锰、铜、锌是叶绿素合 成过程中酶促反应的辅因子。缺少这些元素时就会引起缺 绿症。
➢ 因此,缺少这些元素时都会引起缺绿症,其中尤以氮的影 响最大,因而叶色的深浅可作为衡量植株体内氮素水平高 低的标志。
缺N
CK
萝卜缺N的植株老叶发黄 缺N老叶发黄枯死,新叶色淡,生长矮小,
类囊体膜上含有由多种亚基、多种成分组成的蛋白复合体, 主 要 有 四 类 , 即 光 系 统 Ⅰ ( PSI ) 、 光 系 统 Ⅱ ( PSⅡ ) 、 Cytb6/f复合体和ATP酶复合体(ATPase),它们参与了光能 吸收、传递与转化、电子传递、H+输送以及ATP合成等反应。 由于光合作用的光反应是在类囊体膜上进行的,所以称类囊 体膜为光合膜(photosynthetic membrane) 。
两者结构上的差别仅在于叶绿素a第二个吡咯环上的 一个甲基(-CH3)被醛基(-CHO)•所取代(•图)。
叶绿素的结构特点
叶绿素分子含有一个卟啉 环的“头部”和一个叶绿醇( 植醇,phytol)“尾巴”。卟 啉环由四个吡咯环以四个甲烯 基(-CH=)连接而成。
卟啉环的中央结合着一个 镁离子。镁离子带正电荷,而 与其相连的氮原子则带负电荷 ,因而具有极性,是亲水的。
植物的光合作用-2

所以其量子效率接近1 。
(三)光能的吸收与传递
1、光合作用单位
根据能否进行光化学反应,将叶绿体色素分为二 类:
一类是反应中心色素:它具有光化学活性,既能捕获光能, 又能将光能转换为电能(称为“陷阱”),少数特殊状态的 叶绿素a分子属于此类。 另一类是聚光色素:又称天线色素,它没有光化学活性, 只能进行光物理过程,把吸收的光能传递到反应中心色素, 绝大多数色素(包括大部分chla和全部的chlb、胡萝卜素、 叶黄素等)都属于此类。
一个是吸收短波红光(680nm)的光系统Ⅱ(PSⅡ), PSⅡ颗粒较大,位于类囊体膜的内侧。 另一个是吸收长波红光(700nm)的光系统I(PSⅠ), PSⅠ颗粒较小,在类囊体膜的外侧。 这两个光系统是以串联的方式协同作用的。
4、PSⅠ和PSⅡ的光化学反应
PSⅠ的原初电子受体是叶绿素分子(A0),PSⅡ的 原初电子受体是去镁叶绿素分子(Pheo),它们的次 级电子受体分别是铁硫中心和醌分子。 PSⅠ的原初反应: P700· 0 A
2、光合电子传递体的组成与功能
(1)PSⅡ复合体
A、PSII由3部分组成:
反应中心由2个交叉排列多 肽 D 1和D2组成,其中含有原 初电子供体(Z)、P680、原初电 子受体去镁叶绿素(Pheo)和质 体醌(QA和QB), D 1和D2之间 可能由Fe连接;
PSII反应中心结构模式图
PSII外围是由聚光色素蛋白复合体与细胞色素b559结合的2 条多肽;它们围绕P680,可更快地把吸收的光能传至PSⅡ反应 中心,所以被称为中心天线或“近侧天线”。 放氧复合体(锰聚合体)
卟啉环
第三节 光合作用(Photosynthesis)的机理
光合作用当然需要光,但不是任何步骤都需要光。 根据需光与否,光合作用将分为两个反应─光反应(light reaction)和暗反应(dark reaction)。
第三章 植物的光合作用(试卷及其参考答案2)

第三章植物的光合作用(试卷)姓名:曹晓青学号:同组人:溪树梅、李学梅、曹艳梅、陈以相、黄桂林一、选择题(每题1分,共10分)1、每个光合单位中含有几个叶绿素分子。
()A、100—200B、200—300C、250—3002、C3 途径是由哪位植物生理学家发现的?()A、MitchellB、HillC、Calvin3、类胡萝卜素对可见光的最大吸收峰在()A、蓝紫光区B、绿光区C、红光区4、PSI的光反应属于()A、长波光反应B、短波光反应C、中波光反应5、高等植物碳同化的二条途径中,能形成淀粉等产物的是()A、C4 途径B、CAM 途径C、卡尔文循环6、正常叶子中,叶绿素和类胡萝卜素的分子比例约为()A、2:1B、1:1C、3:17、光合作用碳反应发生的部位是()A、叶绿体膜B、叶绿体基质C、叶绿体基粒8、卡尔文循环中CO2 固定的最初产物是()A、三碳化合物B、四碳化合物C、五碳化合物9、光合产物中淀粉的形成和贮藏部位是()A、叶绿体基质B、叶绿体基粒C、细胞溶质10、光合作用吸收CO2 与呼吸作用释放的CO2 达到动态平衡时,外界的CO2浓度称为()A、CO2 饱和点B、O2 饱和点C、CO2 补偿点二、填空题(每空1分,共20分)1、光合作用的重要性主要体现在3 个方面:________ 、________ 和________ 。
2、在荧光现象中,叶绿素溶液在透射光下呈________ 色,在反射光下呈________ 色。
3、在光合电子传递中,最初的电子供体是________ ,最终电子受体是________。
4、类胡萝卜素吸收光谱最强吸收区在________ 。
5、光合单位由________ 和________ 两大部分构成。
6、卡尔文循环大致可分为3 个阶段,包括________ 、_______ 和________ 。
7、卡尔文循环的CO2 受体是________ 、形成的第一个产物是________ 、形成的第一个糖类是________ 。
植物生理学题库(含答案)第三章 植物的光合作用

植物生理学题库(含答案)第三章植物的光合作用一、名词解释1、爱默生效应:如果在长波红光(大于685nm)照射时,再加上波长较短的红光(650nm),则量子产额大增,比分别单独用两种波长的光照射时的总和还要高。
2、光合作用:绿色植物吸收阳光的能量,同化CO2和H2O,制造有机物质,并释放O2的过程。
3、荧光现象:指叶绿素溶液在透射光下呈绿色,在反射光下呈红色,这种现象就叫荧光现象。
4、磷光现象:当去掉光源后,叶绿素溶液还能继续辐射出极微弱的红光,它是由三线态回到基态时所产生的光。
这种发光现象称为磷光现象。
5、光反应:光合作用的全部过程包括光反应和暗反应两个阶段,叶绿素直接依赖于光能所进行的一系列反应,称光反应,其主要产物是分子态氧,同时生成用于二氧化碳还原的同化力,即ATP和NADPH。
6、碳反应:是光合作用的组成部分,它是不需要光就能进行的一系列酶促反应。
7、光合链:亦称光合电子传递链、Z—链、Z图式。
它包括质体醌、细胞色素等。
当然还包括光系统I和光系统II的反应中心,其作用是传递将水在光氧化时所产生的电子,最终传送给NADP+。
8、光合磷酸化:指叶绿体在光下把有机磷和ADP转为A TP,并形成高能磷酸键的过程。
9、光呼吸:植物的绿色细胞依赖光照,吸收O2和放出CO2的过程。
10、景天科酸代谢:植物体在晚上的有机酸含量十分高,而糖类含量下降;白天则相反,有机酸下降,而糖分增多,这种有机物酸合成日变化的代谢类型,称为景天科酸代谢。
11、光合速率:指光照条件下,植物在单位时间单位叶面积吸收CO2的量(或释放O2的量)12、光补偿点:指同一叶子在同一时间内,光合过程中吸收的CO2和呼吸过程中放出的CO2等量时的光照强度。
13、光饱和现象:光合作用是一个光化学现象,其光合速率随着光照强度的增加而加快,这种趋势在一定范围的内呈正相关的。
但是超过一定范围后光合速率的增加逐渐变慢,当达到某一光照强度时,植物的光合速率就不会继续增加,这种现象被称为光饱和现象。
《植物的光合作用》PPT课件

整理课件
15
观察在照光的叶绿体中淀粉粒会增长 光合作用的另一个产物是有机物
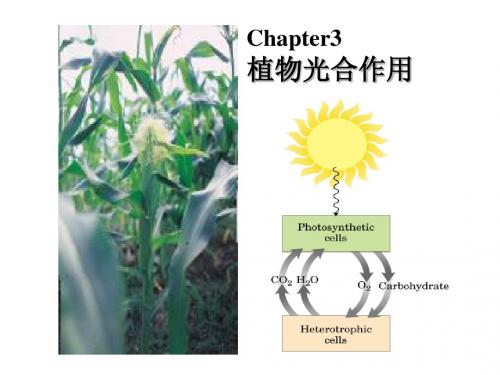
光能
CO2+H2O 绿色植物 (CH2O)+O2 细菌的光合作用
十九世纪的三十年代 C B Van Niel
某些细菌 醋酸 琥珀酸 H2S
CO2+2H2S
(CH2O)+H2O+2S
比较 植物释放的氧来自水,而不是二氧化碳
叶绿醇 是叶绿素分子的亲脂部分,是长链 亲脂“尾巴”,伸入类囊体内
“头部”是金属卟啉环,Mg偏正电荷,N原 子偏带负电荷,呈极性,具亲水性(可和蛋白质结 合),排列在类囊体脂类的表面.
整理课件
22
㈡叶绿素的化学性质 ⑴不溶于水 而溶于有机溶剂
用水配85%丙酮提取叶绿素
⑵皂化作用
C32H30ON4Mg COOCH3 +2KOH
h 普朗克常数 1.58×10-34卡.秒
c 光速 3×1010㎝/秒
COOC20H39 C32H30ON4Mg COOK
+CH3OH
+C20H39OH
COOK
皂化叶绿素 叶醇 整理课件
甲醇
23
⑶形成去镁叶绿素
phMg+2H+ 褐色
H
Ph
+Mg++
H
H
ph H + Cu++(Zn++)
绿色
phCu(Zn)+2H+
整理课件
24
三,叶绿素的光学性质 ⑴吸收光谱
波长在600-660nm的红光 波长在430-450nm蓝紫光 绿光吸收最少
植物的光合作用教案

植物的光合作用教案一、教学内容本节课我们将学习《植物的光合作用》,该部分内容位于教材第三章第二节。
详细内容包括植物光合作用的基本概念、反应式、影响因素、生态环境意义等。
二、教学目标1. 理解并掌握植物光合作用的基本原理,能够正确书写光合作用反应式。
2. 了解影响植物光合作用的因素,认识到光合作用在生态环境中的重要作用。
3. 培养学生的实验操作能力和观察分析能力,激发对生物学研究的兴趣。
三、教学难点与重点重点:植物光合作用的基本原理和反应式。
难点:影响植物光合作用的因素及其在生态环境中的意义。
四、教具与学具准备教具:多媒体课件、黑板、粉笔、植物光合作用实验装置。
学具:笔记本、笔、实验报告单。
五、教学过程1. 实践情景引入(5分钟)利用多媒体展示绿色植物的生长过程,引导学生思考:为什么绿色植物能生长得如此茂盛?2. 知识讲解(15分钟)通过讲解,使学生了解植物光合作用的基本原理、反应式以及影响光合作用的因素。
3. 例题讲解(15分钟)分析一道关于植物光合作用的例题,让学生巩固所学知识。
4. 随堂练习(10分钟)发放随堂练习题,检查学生对本节课知识的掌握情况。
5. 实验演示(10分钟)演示植物光合作用实验,引导学生观察实验现象,并分析原因。
6. 小组讨论(10分钟)学生分小组讨论影响植物光合作用的因素,并汇报讨论成果。
六、板书设计1. 植物的光合作用基本原理反应式影响因素生态环境意义七、作业设计1. 作业题目:(1)简述植物光合作用的基本原理。
(2)写出植物光合作用的反应式。
(3)举例说明影响植物光合作用的因素。
2. 答案:(1)植物光合作用是绿色植物在光的作用下,将二氧化碳和水合成有机物,同时释放氧气的过程。
(2)6CO2 + 6H2O + 光能→ C6H12O6 + 6O2(3)影响植物光合作用的因素有光照强度、温度、水分、二氧化碳浓度等。
八、课后反思及拓展延伸1. 课后反思:本节课学生是否掌握了植物光合作用的基本知识,实验操作是否熟练,讨论环节是否积极参与。
光合作用 (2)
基质 主要成分是可溶性蛋白质及
其它代谢活跃物质。羧化酶约占可
溶性蛋白质的50﹪,还DNA、
RNA、核糖体、淀粉体、嗜锇颗粒
(叶绿体的脂类仓库)等。
二、光合色素的结构与性质 光合色素主要有三类:叶绿素、类胡 萝卜素、藻胆素。它们存在于类囊体上。 前两类为高等植物的叶绿体色素。 1、叶绿素(chlorophyll,chl) 主要有Chla和Chlb,不溶于水,易溶 于乙醇、丙酮等有机溶剂。
类胡萝卜素的最大吸收峰在蓝紫光区。不 吸收长波光
(三)荧光现象和磷光现象
荧光现象:叶绿素溶液在透射光下呈 绿色,而在反射光下呈红色的现象。
Chl + hν chl* 激发态
基态 光子能量
蓝 光
红 光
荧光(fluorescence): CHL从第一
-
单线态回到基态所发射的光。
磷光(phosphorescence):CHL从 第一三单线态回到基态所发射的光。 叶绿素的荧光和磷光现象说明叶 绿素能被光所激发,而叶绿素的激发 是将光能转变为化学能的第一步。
吸收光谱:叶绿素对不同波长光吸收后 形成的光谱。
叶绿素在红光区(640~660nm)和蓝紫 光区( 430~450nm)有最强吸收。叶绿素 对绿光吸收最少,故叶绿素溶液呈绿色。
类胡箩卜素在蓝紫光区有最强的吸收。
chla与chlb吸收光谱的区别: ▽ chla在红光区的吸收带偏向长波方向, 吸收带较宽,吸收峰较高。在蓝紫光区吸收 带偏向短波方向,吸收带较窄,吸收峰较低。 对蓝紫光的吸收为对红光的吸收的1.3倍。 ▽ chlb在红光区的吸收带偏向短波方向, 吸收带较窄,吸收峰较低。在蓝紫光区吸收 带偏向长波方向,吸收带较宽,吸收峰较高。 对蓝紫光的吸收为对红光的吸收的3倍,说明 chlb吸收短波蓝紫光的能力较chla 强。
第三章 植物的光合作用

2.意义 (1)将无机物转变成有机物 CO2 + 4H+ —— (CH2O) (2)随着物质的转变,将光能转变为化学能。
光能电能活跃的化学能稳定的化学能 hv eATP、NADPH CH2O
(3)释放氧,净化空气;并产生臭氧,滤去太 阳光中对生物有强烈破坏作用的紫外光。
第三章 植物的光合作用
430 nm波长的蓝光量子的能量为4.5710-19J
(二)吸收光谱(absorption spectrum) 太阳光到达地表面的波长大约是 300 nm —— 2600 nm 可见光的波长是 390 nm —— 770 nm 连续光谱: 光束通过三棱镜后可把白光分为七色 连续光谱。
吸收光谱:
第三、四、五节 光合作用过程
光合作用机制的研究
• 在研究外界条件影响时发现:弱光下增加光强能提高 光合速率,但当光强增加到一定值时,光合速率便不 再随光强的增加而提高;此时提高温度或CO2浓度才 能增强光合速率。由此推理,光合作用至少有两个步 骤:其一需要光,另一个则与温度相关。 希尔反应的发现和水氧化钟模型的提出。 藻类闪光实验:在光能相同时,一种用连续光照;另一 用闪光照射,中间间隔一暗期。发现后者的光合速率 比前者的要高。表明,光合作用不是任何步骤都需要 光。根据需光与否,将光合作用分为两个反应—光反 应和暗反应。
(1)叶绿素a在红光区的吸收带偏向长光波方 向,而在兰紫光区则偏向短光波方向.
(2)叶绿素a在红光区的吸收带宽些,兰紫光 区窄些,叶绿素b与其相反.
2.类胡罗卜素的吸收光谱 1)吸收光谱 —— 兰紫光区 2)重要功能 —— 吸收光能并向叶绿素传递 ——可进行光保护 吸收兰紫光形成激发态的类胡罗卜素 热耗散返回基态淬灭激发态的叶绿素 避免吸收的多余能量对光合系统的伤害
植物的光合作用

第三章植物的光合作用一填空1.绿色植物和光合细菌都能利用光能将合成有机物,它们都属于光养生物。
从广义上讲,所谓光合作用,是指光养生物利用把合成有机物的过程。
(CO2,光能,CO2)2.光合作用本质上是一个氧化还原过程。
其中是氧化剂,是还原剂,作为CO2还原的氢的供体。
(CO2,H2O)3.1940年S.Ruben等发现当标记物为H218O时,植物光合作用释放的O2是,而标记物为C18O2时,在短期内释放的O2则是。
这清楚地指出光合作用中释放的O2来自于。
(18O2,O2,H2O)4.1939年Robert.Hill发现在分离的叶绿体悬浮液中加入适当的电子受体,如铁氰化钾或草酸铁等,照光时可使水分解而释放氧气,这一现象称为,其中的电子受体被称为。
(希尔反应,希尔氧化剂)5.1954年美国科学家D.I.Arnon等在给叶绿体照光时发现,当向体系中供给无机磷、ADP和NADP时,体系中就会有和两种高能物质的产生。
同时发现,只要供给了这两种高能物质,即使在黑暗中,叶绿体也可将转变为糖。
所以这两种高能物质被称为“”。
(ATP,NADPH,CO2,同化力) 6.20世纪初人们研究光强、温度和CO2浓度对光合作用影响时发现,在弱光下增加光强能提高光合速率,但当光强增加到一定值时,再增加光强则不再提高光合速率。
这时要提高温度或CO2浓度才能提高光合速率。
用藻类进行闪光试验,发现在光能量相同的前提下闪光照射的光合效率是连续光下的200%~400%。
这些实验表明光合作用可以分为需光的和不需光的两个阶段。
(光反应,暗反应) 7.由于ATP和NADPH是光能转化的产物,具有在黑暗中使光合作用将CO2转变为有机物的能力,所以被称为“ ”。
光反应的实质在于产生“”去推动暗反应的进行,而暗反应的实质在于利用“”将转化为有机碳(CH2O)。
(同化力,同化力,同化力,CO2)8.量子产额的倒数称为,即光合作用中释放1分子氧和还原1分子二氧化碳所需吸收的。
植物生理第三章

(二)原初反应的过程 —光能的吸收、传递和光化学反应
1.天线色素接受光能,以诱导共振方式 将能量传递到光合反应中心。
能量传递效率: Chla,b几乎100%传给反应中心色素, 类胡萝卜素 约90%传给反应中心色素。
42
2.光合反应中心发生光化学反应
hυ ┋ D P A → D P* A → D P+ A- → D+ P A①特殊叶绿素a ②高能电子脱离,
9
外被膜
被膜 (envelop)
内被膜 选择透性
叶绿体
(Chloroplast)
膜—光合色素、光合链——原初反应、 电子传递和光合磷酸化(光合膜 ) (thylacoid) 类囊体 腔—光合放O2 间质(stroma)——光合碳循环酶(Rubisco), CO2固定(同化); DNA,RNA,核糖体70S——部 分遗传自主
醛基(CHO)
14
1.叶绿素的结构:
②双羧酸尾部:
1个羧基在副环(V)上 以酯键与甲基结合 --甲基酯化; 另一个羧基(丙酸) 在IV环上与植醇 (叶绿醇)结合- -植醇基酯化; 非极性,亲脂,插 入类囊体的疏水区, 起定位作用。
15
2.叶绿素的作用:
收集和传递光能 (大部分Chl a和全部Chl b) 将光能转换为电能(少数特殊Chl a)
2.类胡萝卜素
强吸收区: 400-500 (蓝紫); 不吸收区: 500以上
25
(二)光能的吸收和释放
物质吸收光子,其原子中的e 重新排列,分子从基态(最 低、最稳定)跃迁到激发态 (高能、不稳定)
Chl+ hγ= Chl* 处于激发态的分子,趋 于释放能量回到基态
26
潘炽瑞植物生理学习题答案.3

第三章植物的光合作用一、英译中(Translate)1、heterophyte2.、autophyte3、photosynthesis4、chloroplast5.、thylakoid6.、Photosynthetic membrane7、chlorophyll8、carotenoid9、carotene10、xanthophyll11、absorption spectrum12、etiolation13、light reaction14、carbon reaction15、primary reaction16、photosynthetic unit17、Emerson effect18、electron transport19、photosynthetic chain20、photophosphorylation21、coupling factor22、chemiosmotic hypothesis23、the Calvin cycle24、reductive pentose phosphate pathway25、phosphoenol pyruvate 26、photorespiration27、dark respiration28、peroxisome29、photosynthetic product30、Photosynthetic rate31、light compensation32、light saturation33、shade plant34、photoinhibition35、greenhouse effect36、solar constant37、thylakoid lumen38、Rubisco39、antenna pigment40、light –harvesting pigment41、reaction center42、photosystem I43、oxygen-evolving complex44、water splitting45、water oxidizing clock46、core complex47、assimilatory power48、CO2 assimilation49、fluorescence二、中译英(Translate)1、异养植物2、自养植物3、光合作用4、叶绿体5、类囊体6、光合膜7、叶绿素8、类胡萝卜素9、胡萝卜素10、叶黄素11、吸收光谱12、黄化现象13、光反应14、碳反应15、原初反应16、光合单位17、爱默生效应18、电子传递19、光合链20、光合磷酸化21、偶联因子23、化学渗透假说23、卡尔文循环24、还原戊糖磷酸途径125、磷酸烯醇式丙酮酸26、光呼吸27、暗呼吸28、过氧化物酶体29、光合产物30、光合速率31、光补偿点32、光饱和现象33、阴生植物34、光抑制35、温室效应36、太阳常数37、类囊体腔38、CO2补偿点39、天线色素40、聚光色素41、反应中心42、光系统I43、放氧复合体44、水裂解45、水氧化钟46、核心复合物47、同化力48、CO2同化49、荧光三、名词解释(Explain the glossary)1、爱默生效应18. absorption spectrum2、光合作用19. assimilatory power3、荧光现象20. ATP synthase(ATPase or CFo-CF1)4、磷光现象21. C3 plants5、光反应22. C4 cycle6、碳反应23. C4 plants7、光合链24. Calvin cycle8、光合磷25. CAM plants9、光呼吸酸化26. Cytochrome b6f complex10、景天科酸代谢27. mesophyll11、光合速率28. photosynthesis12、光补偿点29. photosynthetic electron transport13、光饱和现象30. photosystem14、光抑制31. photosystem I(PSI)15、光能利用率32. photosystem II(PSII)16、光合单位33. rubisco17、CO2补偿点34. chloroplast四、是非题(True or false)()1、叶绿体是单层膜的细胞器。
第三章植物的光合作用

能进行光合作用的细菌称之为光合细菌 。光合细 菌包括蓝细菌、紫细菌和绿细菌等。其中蓝细菌的 光合过程与真核生物相似,紫细菌和绿细菌则不能 分解水而需利用有机物或还原的硫化物等作为还原 剂。
例如:紫色硫细菌(purple-Sulfur bacteria)和绿色硫 细菌 (green-sulfur bacteria)利用H2S为氢供体,在 光下同化CO2:
色,这种现象叫荧
光现象,发出的光
叫荧光.
磷光现象: 当荧光出现后,立即中断光源,色素分子仍 能持续短时间的“余辉”,这种现象,叫磷
光现象,发出的光叫磷光。
荧光现象与磷光现象产生的原因 (一) 激发态的形成
通常色素分子是处于能量的最低状态-基态(ground state) 。 色素分子吸收了一个光子后,会引起原子结构内电子的重新 排列。
醇
(三)藻胆素
仅存在于红藻和蓝藻中,主 要有藻红蛋白、藻蓝蛋白和 别藻蓝蛋白三类,前者呈红 色,后两者呈蓝色。它们的 生色团与蛋白以共价键牢固 地结合。 藻胆素分子中的四个吡咯环 形成直链共轭体系,不含镁 也没有叶绿醇链。藻胆素也 有收集光能的功能。 由于类胡萝卜素和藻胆素吸 收的光能能够传递给叶绿素 用于光合作用,因此它们被 称为光合作用的辅助色素。
镁卟啉
亲水的“头部”
颜色来源
叶醇基(双萜)
亲脂的尾部
(二) 类胡萝卜素
种类:胡萝卜素和 叶黄素
紫罗兰酮环
溶解性:有机溶剂
颜色:胡萝卜素呈橙 黄色,叶黄素呈黄色 功能:收集光能,防 护多余光照伤害叶绿 素
结构式:C40H56,常 见的为β-胡萝卜素, 叶黄素是胡萝卜素衍 生的醇类,C40H56O2
2. 发射荧光与磷光 激发态的叶绿素分子回至基态时,以光子形式释放 能量。 处在第一单线态的叶绿素分子回至基态时所发出的 光称为荧光(fluorescence)。
植物生理学003 植物的光合作用2

2020/10/27
2020/10/27
➢ 铁氧还蛋白(Fd)和铁氧还蛋白-NADP+还原酶(FNR 、Fp)都是存在类囊体膜表面的蛋白质。
➢ Fd是通过它的2铁-2硫活性中心中的铁离子的氧化还 原传递电子的。
➢ FNR(Fp) 含1分子的黄素腺嘌呤二核苷酸(FAD),依 靠核黄素的氧化还原来传递H+。FNR(Fp)是光合电 子传递链的末端氧化酶,接收Fd传来的电子和基质 中的H+,还原NADP+为NADPH,反应式可用下 式表示:
网络式地进行。
➢ 质体醌是双电子、双质子传
Cytb6/f
递体,对类囊体膜内外建立
质子梯度起着重要的作用。
2020/10/27
质蓝素
➢ 质蓝素(PC)是位于类囊体膜内侧表面的含铜的蛋白 质,氧化时呈蓝色。它是介于Cyt b6/f复合体与 PSⅠ之间的电子传递成员。通过蛋白质中铜离子的 氧化还原变化来传递电子。
第三节 光合作用的机理
通过小球藻闪光实验发现,同等光强相同时间下,连续照射, 光合效率 小,放氧少;间歇照射(每次闪光0.0034S,间歇 0.0166S,每秒50次), 光合效率高,放氧多——说明光合作用需要光照,但并不是任何过程都 需要光照。
因此提出光反应(light reaction)和暗反应(dark reaction)的概念。 光反应是必须在光下才能进行的、由光所引起的光化学反应;暗反应是 在暗处(也可在光下)进行的,由若干酶所催化的化学反应。光反应是 在类囊体(光合膜)上进行的,而暗反应是在叶绿体的基质中进行的。
3、光能的吸收与传递
光照到叶绿体上时,类囊体膜上的叶绿体色素吸收光能而激发,由于色素分子 排列紧密及其特殊的共轭体系,激发能可在相同色素分子间(2nm)以激子传递 (exciton transfer)或相同及不同色素分子间(>2nm)共振传递(resonance transfer)的方式传递,最后传给反应中心的中心色素(chla)。 激发能的传递速度及效率很高,一个红光量子可传过几百个chla分子(30ps/个), 效率达90~100%。
七年级生物上册3-4-1植物的光合作用(第2课时)教案(新版)苏科版

第1节植物的光合作用(第2课时)一、教学目标(一)知识目标(1)在进行探究植物光合作用场所的活动后,理解光合作用的场所是叶绿体、主要器官是叶,从光合作用的场所这一角度了解绿色植物才能进行光合作用。
(2)理解“光合作用”这一核心概念的公式和含义。
(二)能力目标(1)学习从观察植物图片发现并提出光合作用场所的问题,尝试对众多的问题进行分析整合、寻找问题的关键。
(2)由制定探究计划中对实验材料的巧妙选择,学习如何控制唯一变量、排除其它因素的干扰。
(3)学习不断反思和总结自己所做的探究活动及所得到的结论,在反思与总结中加深对叶的结构和光合作用场所的认知。
(三)情感态度与价值观目标(1)通过自己的探究活动深刻认识到绿色植物才能进行光合作用,认同绿色植物在生物圈中不可替代的重要性。
(2)通过“课外延伸”活动,引导学生学以致用,利用所学探究技能探究生活的实际问题。
二、教学重难点重点:掌握光合作用的概念,理解探究实验设计。
难点:有关光合作用的实验设计和操作。
三、课时安排2课时四、教学过程(一)情境导入绿色植物在生物圈中,是重要的生产者,因为它们能进行光合作用。
经过前面的探究学习我们已经知道:光合作用能将二氧化碳和水合成淀粉等有机物,并放出氧气;光合作用的条件是光照。
可是,光合作用的场所在哪里呢?学生:复习光合作用的合成淀粉等有机物、条件是光照。
(二)学习新课一、了解“恩吉尔曼实验”现阶段,我们对“光合作用”的认识,还是对前人研究的继承和发展。
所以,首先让我们一起去了解德国科学家恩吉尔曼所做的相关研究。
(播放恩吉尔曼实验动画)1880年,恩吉尔曼用水绵和好氧细菌进行了光合作用的实验。
(1)实验中,把载有水绵和好氧细菌的临时装片,放在没有空气并且黑暗的环境中,排除环境中光线和氧的影响,然后用极细的光束照射水绵。
通过显微镜观察,发现好氧细菌只集中在叶绿体受光部位的周围;(2)将上述临时装片完全暴露在阳光下,则观察到好氧细菌集中在叶绿体所有受光部位的周围。
植物的光合作用 no2c

• 但也不是光照超强越好,在一定范围内,光合速 率随着光照强度的增加而加快,当光照达到强度 时,光合速率达到最大值,以后,光合速率不再 随光照强度的增加而加快,这种现象称为: 光饱和现象 light saturation。 • 使光合速率达到最大值的最小光强,称为: 光饱和点 light saturation point。 • 出现光饱和现象的原因可能有两个: ① 光合色素和光反应来不及利用更多的光能,每 个叶绿素分子每秒只能接受10个光量子; ② 暗反应慢于光反应,不能充分利用光反应产生 的同化力。
Rate of photosynthesis
Effect of [O2] on C3 vs C4 Photosynthesis
110 100 90 80 70 60 50 40 30 20 10 0 0 20 40 60 Percent Oxygen Concentration
C4 - corn
Rate of PS (%)
二、影响光合作用的内部因素
• 影响光合作用的内部因素很多,非常复杂,所有光合过程 所涉及到的因素,都会影响到光合作用,但从大的环节看, 主要有几个方面: 1.叶绿素含量 chlorophyll content • 叶绿素含量高,吸收光量子的能力强,光反应强,可为 CO2固定提供较多的同化力。 2. Rubisco 和PEPC的活性 (activity) • Rubisco 和PEPC含量高,CO2固定和同化的效率高,光 合作用的速度加快。 3.叶片气孔的数量和大小 number and size of stomata • 叶片的气孔是CO2进入叶片内部的通道,气孔的多少和大 小将影响CO2的供应,从而影响光合作用。
3.CO2
• CO2是光合作用的原料,在一定范围内,随CO2浓度升高, 光合速率增大。光合作用也存在CO2饱和现象 CO2 saturation。CO2饱和点约为1000ppm,CO2饱和的原因 可能是由于气孔关闭。 • 在CO2饱和点以下,随CO2浓度降低,光合速率降低,当 CO2降到某一浓度时,光合吸收的CO2数量与呼吸释放的 CO2数量相等,(也就是表观光合速率为零),这时的 CO2浓度称为CO2补偿点 CO2 compensation point。 • 不同类型植物CO2补偿点不同,C4植物由于CO2呼吸极低, CO2补偿点很低,小于10ppm,C3植物CO2补偿点较高 30~70ppm。(μl· -1) L • CO2补偿点代表了植物利用低浓度CO2的能力。
植物生理学第3章 植物的光合作用(二)

3.3 光合作用机理
反应中心
• 光能转换色素分子+原初电子受体+原初电 子供体。
• 基本成分为蛋白质和脂类。 • 光能转换色素分子为叶绿素a分子,这个叶
绿素a分子与脂蛋白结合排列在片层结构上。 • 色素电子传递。 • 高等植物最终电子供体是水,最终电子受
体为NADP。
7.10 Basic concept of energy transfer during photosynthesis
Cytb6-f复合体
• 组成:有4个多肽组成,其 中三个含Fe,(分别含cytb6, cytf,和 Fe-S)。
• 功能:参与电子传递和质子 传递(PQ穿梭)
4) ATP合成酶复合体
• 组成:
CF1: α:β:γ:δ:ε 5 种多肽。
组成比例为 α:β:γ:δ: ε= 3:3:3:3:1。
CF0由I、II、III和IV四个 亚基组成。组成比例为 I: II:III:IV= 1:1:12:1。
种波长的光波促进光合效率的现象称 为 Emerson enhancement effect.
3.3 光合作用机理
Emerson效应: Emerson(1957)发现在远红光条件下如补充红光(波 长大于650nm),则量子产量大增,且比这两种波长光单独照射的量 子产量还多,这种现象称为Emerson效应。
光系统II(PSII)
• 三部分组成: A. D1&D2:
• 中心色素分子:P680 • 原初电子受体:pheo • 原初电子供体:Z(Tyr) • QA,QB等传递体 LHCII: CP43 & CP47, B559 OEC or MSP: • 33 kDa, 23 kDa & 16
光合作用的奥秘初中生物之植物的光合作用 (2)

光合作用主要发生在叶绿体中,需要光、水、二氧化碳和光合色素等基本条件。
光合作用可以分为光反应和暗反应两个阶段,光反应释放氧气并合成ATP和NADPH,暗反应则 合成有机物。
光合作用的过程
吸收光能:植物吸收太阳光能,转化为化学能 水的光解:水分子在光的作用下分解成氧气和氢离子 碳的固定:二氧化碳与五碳化合物结合生成两个三碳化合物 能量转化:将光能转化为化学能,储存在有机物中
温度对光合作 用过程中的气 体交换:温度 会影响植物的 气孔开度,进 而影响光合作 用过程中的气 体交换。
二氧化碳浓度对光合作用的影响
二氧化碳是光合 作用的主要原料, 直接影响光合作 用的速率
高浓度的二氧化 碳可以促进植物 气孔开放,增加 光合作用的原料 供给
低浓度的二氧化 碳可能导致气孔 关闭,影响光合 作用的进行
光合作用的产物
水:光合作用中释放出的氧气是光解水产生的
葡萄糖:光合作用中生成的有机物主要是葡萄糖 能量:光合作用中储存的能量来源于光能 合成蛋白质和脂肪:光合作用中还可以合成一些蛋白质和脂肪 等有机物
04
光合作用的影响因素
光照强度对光合作用的影响
光照强度是影响光合作用的重要因素之一,光照强度越高,光合作用速率越快。
意义:光合作用是植物生长和发育的基础,也是生态系统稳定和持续发展的关键。
光合作用的条件
光照:光合作用需要光 照作为能量来源
水:光合作用需要水作 为原料
二氧化碳:光合作用需 要二氧化碳作为另一个 重要原料
叶绿体:光合作用主要 在叶绿体中进行,叶绿 体是绿色色素和酶的复 合体,能够吸收光能并 转化为化学能
生物治理:光合作用能够吸收 二氧化碳并释放氧气,可以用 于治理环境污染和缓解全球气 候变暖等问题。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
光合作用中各种能量转变情况
•
能量转变 光能
贮能物质 量子 转变过程
电能
电子
活跃的化学能
ATP、NADPH2 光合磷酸化
稳定的化学能
碳水化合物等 碳同化 101-102 叶绿体间质
原初反应
电子传递
时间跨度(秒)10-15-10-9 10-10-10-4 100-101 反应部位
PSⅠ、PSⅡ颗粒 类囊体膜 类囊体
第3节 光合作用的过程
H 2O
光解 光能 吸收
色素分子
O2
2C3
酶
[H]
ATP
还
固
CO2
供能
多种酶 定 C5
原
(CH2O)
酶
酶
ADP+Pi
暗反应阶段
CO2的固定: CO2+C5 C3化合物还原:2 C3
6
光反应阶段
水的光解:H2O
光解
2[H]+1/2 O2
酶
酶
2C3
光合磷酸化:ADP+Pi+能量
ATP
PSⅠ复合体存在类囊体非堆叠的部分,PSⅡ复合体存在堆叠 部分,而Cyt b6/f 比较均匀地分布在膜中,因而推测PC通 过在类囊体腔内扩散移动来传递电子。
5.PSⅠ复合体
PSⅠ的生理功能是吸收光能,进行光化学反应,产生强的 还原剂,用于还原NADP+,实现PC到NADP+的电子传递。
高等植物的PSⅠ由反应中心和LHCⅠ等组成。
ATP和NADPH
概念
光合磷酸化: 光下在叶绿体(或载色体) 中发生的由ADP与Pi合 成ATP的反应。
(一) 光合磷酸化的类型
1.非环式光合磷酸化
与非环式电子传递偶联产生ATP的反应。 2NADP++3ADP+3Pi 8hυ叶绿体 2NADPH+3ATP+O2+2H++6H2O
在进行非环式光合磷酸化的反应中,体系除生成ATP外,同时 还有NADPH的产生和氧的释放。 含有基粒片层的放氧生物所特有,在光合磷酸化中占主要地位。
铁氧还蛋白(Fd)和铁氧还蛋白-NADP+还原酶(FNR)都是存在类囊 体膜表面的蛋白质。 Fd是通过它的2铁-2硫活性中心中的铁离子的氧化还原传递电子 的。 FNR含1分子的黄素腺嘌呤二核苷酸(FAD),依靠核黄素的氧化还 原来传递H+。FNR是光合电子传递链的末端氧化酶,接收Fd传 来的电子和基质中的H+,还原NADP+为NADPH,反应式可用下 式表示:
光合膜上的电子与质子传递概况
红线表示电子传递,黑线表示质子传递,蓝线质子越膜运输
三、光合磷酸化
光合磷酸化的发现
1954,Arnon等发现向菠菜叶绿体加入ADP、Pi和NADP+, 在光下不加入CO2时,在体系中有ATP和NADPH产生。
破碎
黄色上清液 绿色上清液
含暗反应的酶
完整叶绿体
离心
ADP、 Pi和 NADP+
第四节 电子传递和光合磷酸化
原初反应的结果:
产生了高能电子推动着光 合膜上的电子传递。 电子传递的结果: 引 起 水 的 裂 解 放 氧 以 及 NADP+的还原; 建立了跨膜的质子动力势, 启动了光合磷酸化,形成 ATP。 这样就把电能转化为活跃的化学能。
一、光系统
人物:爱默生 事件:测定不同波长光的光合效率实验 结果: • 大于685nm的远红光照射,光合速率下降。-红降 (1943年) • 远红光与650nm的红光同时照射,光合速率增加,大于 两者单独照射的和。-双光增益效应(爱默生效应) (1957年)
2.环式光合磷酸化
与环式电子传递偶联产生ATP的反应。
ADP + Pi
光 叶绿体
ATP+ H2O
非光合放氧生物光能转换的唯一形式,主要在基质片层 内进行。它在光合演化上较为原始,在高等植物中可能 起着补充ATP不足的作用。
(二)光合磷酸化的机理
1.光合磷酸化与电子传递的关系--偶联
光合磷酸化作用都与电子传递相偶联: 如在叶绿体体系中加入电子传递抑制剂,光合磷酸化就会停止; 在偶联磷酸化时,电子传递则会加快,所以在体系中加入磷 酸化底物会促进电子的传递和氧的释放。 (解偶联:发生电子传递而不伴随磷酸化作用)
1、两个光系统
PSⅡ: PSⅡ复合体存在堆叠部分 • 反应中心色素P680 • 聚光复合体 • 放氧复合体
PSⅠ: PSⅠ复合体存在类囊体非堆叠的部分 • 反应中心色素P700 • 聚光复合体 • 电子受体
2、电子传递和质子传递:
(电能转化为不稳定的化学能)
• 光合电子传递链:(光合链)是指在类囊体膜上,由PSⅠ, PSⅡ和其它电子传递体相互衔接,所构成的电子传递体系。
[H], 酶 (CH2O) ATP
ADP+Pi
光合作用的过程和能量转变
光合作用的实质是将光能转变成化学能。 根据能量转变的性质,将光合作用分为三个阶段: 原初反应:即光能的吸收、传递和转换过程光能的吸 收、传递和转换成电能;
电子传递和光合磷酸化:电能转变为活跃化学能;
碳同化:活跃的化学能转变为稳定的化学能。
2.光合磷酸化的机理
中间产物学说 变构学说 化学渗透学说 许多实验都证实了这一学说的正确性。
化学渗透学说 (chemiosmotic theory)
由英国的米切尔(Mitchell 1961)提出, 该学说假设能量转换和偶联机构具有以下特点: ①由磷脂和蛋白构成的膜对离子和质子的透过具有选择性 ②具有氧化还原电位的电子传递体不匀称地嵌合在膜上 ③膜上有偶联电子传递的质子转移系统 ④膜上有转移质子0 A1
FX,FA,FB的Fe-S中心
铁-硫中心的模型 左:4铁-4硫型; 右:2铁-2硫型
FX、FA、FB是PSⅠ中3个铁 硫蛋白,都具有4铁-4硫中心结 构,其中4个硫与蛋白质的4个 半胱氨酸残基连接,它们主要 依4铁-4硫中心中的铁离子的氧 化还原来传递电子。
6.铁氧还蛋白和铁氧还蛋白-NADP+还原酶
2、原初反应过程
a.吸收:聚光色素(天线色素)
没有光化学活性,只有收集光能的作 用。包括大部分Chla、全部Chlb和全 部类胡萝卜素
b.传递:方式是诱导共振
方向从高能向低能,从短波色素向长波色 素。
类胡萝卜素
Chlb
Chla
c.转换:反应中心色素
具有光化学活性,既是光能的捕捉 器,又是光能的转换器。由少数特 殊状态的Chla组成。 反应中心:D—原初电子供体 电子受体 光 DPA DP*A 光能 P—反应中心色素 e DP+A电能 e D+PA?? A—原初
Cyt b6/f 复合体主要催化PQH2的氧化和PC的还原,并把质子 从类囊体膜外间质中跨膜转移到膜内腔中。因此Cyt b6/f 复 合体又称PQH2·PC氧还酶。 PQH2+2PC(Cu2+)
Cyt b6/f
PQ +2PC(Cu+)+ 2H+
4.质蓝素
质蓝素(PC)是位于类囊体膜内侧表面的含铜的蛋白质,氧化时 呈蓝色。它是介于Cytb6/f复合体与PSⅠ之间的电子传递成 员。通过蛋白质中铜离子的氧化还原变化来传递电子。
在解释光合磷酸化机理时,该学说强调:光合电子传递链的电子传 递会伴随膜内外两侧产生质子动力(proton motive force,pmf),并 由质子动力推动ATP的合成。
化学渗透学说 (chemiosmotic theory)
Peter Mitchell (1920 - 1992)
The 1978 Nobel Prize laureate in Chemistry
2Fd还原+NADP++ H+
FNR
2Fd氧化 + NADPH
7.光合膜上的电子与H+的传递
光合膜上的电子与质子传递
图中经非环式电子传递途径传递4个e-产生2个NADPH和3个ATP是根据光合作用总方程 式推算出的。在光反应中吸收8个光量子(PSⅠ与PSⅡ各吸收4个),传递4个e-能分解 2个H2O,释放1 个O2,同时使类囊体膜腔增加8个H+,又因为吸收8个光量子能同化1 个CO2,而在暗反应中同化1个CO2需消耗3个ATP和2个NADPH,也即传递4个e-, 可还原 2个NADPH,经ATP酶流出8个H+要合成3个ATP。
组成中心天线的CP47和CP43是指分子量分别为47 000、
43 000并与叶绿素结合的聚光色素蛋白复合体,它们
围绕P680,比LHCⅡ更快地把吸收的光能传至PSⅡ反 应中心,所以被称为中心天线或“近侧天线”。
PSⅡ反应中心的核心部分是分子量分别为32 000和34 000的D1和D2两条多肽。 反应中心的次级电子供体Z、中心色素P680、原初电 子受体Pheo、次级电子受体QA、QB等都结合在D1和D2上。 与D1 结合的质体醌定名为QB ,与D2 结合的质体醌定名 为QA。 QA 是单电子体传递体,每次反应只接受一个电子生成 半醌。它的电子再传递至QB。 QB 是双电子传递体,QB 可两次从QA 接受电子以及从周 围介质中接受2个H+ 而还原成氢醌(QH2 ) 。这样生成的氢 醌可以与醌库的PQ交换,生成PQH2。
光合作用是光反应和暗反应的综合过程。
1、原初反应定义
是指从光合色素分子被光激发到引起第一个光化学反应 为止的过程。
光化学-有电子得失 原初反应特点:
它包括:
光物理-光能的吸收、传递
• 速度非常快,10-12s-10-9s内完成;
• • 与温度无关 (77K,液氮温度)(2K,液氦温度); 量子效率接近1
PSⅡ复合体与多个Cyt b6/f
复合体发生联系,使得类囊 体膜上的电子传递成网络式