2019年色氨酸操纵子.ppt
原核表达调控与色氨酸操纵子优秀课件

1、不依赖于ρ因子的终止子(强终止子)
• 结构特点:RNA具有一个发夹结构(富含GC) 和随后polyU片段。
• 作用机制:RNA pol + 发夹结构 → 转录暂停→ 后随杂合分子不稳定的poly(U-dA) → RNA容易 从模板上脱落→RNA-DNA-RNA pol 解聚 → 转 录终止
-10
+1
+10
stream
start point
downstream
一、转录的起始
• 关键:全酶能以很高的亲和性结合在启 动子promoter ;
• 启动子:RNA聚合酶识别、结合并起始 转录的一段DNA序列。
转录的起始
• 核心酶在σ因子帮助下特异性结合到DNA上; • RNA pol与启动子-35 box 结合,形成封闭型起
RNA polymerase
holoenzyme and
DNA
a promoter
Incorporating the first few Nt
二、原核生物转录的延伸
• RNA pol沿着模板链移动,RNA链不断延伸, 并保持三元复合物的结构;
• 转录泡中的DNA螺旋前开、后合,RNA延长, 不断脱离DNA;
• RNA的转录包括promotion,elongation, termination 三过程;
• 从promoter到terminator称为transcriptional unit; • 原核生物中的转录单位多为 polycistron ; • 转录起始点记为+1,其上游记为负值,下游记为正值。
原核表达调控与色氨酸操纵子

开发高效原核表达系统
继续优化原核表达系统,提高外源基因在原核细 胞中的表达效率和稳定性,降低生产成本,推动 基因工程药物和疫苗的广泛应用。
深入解析代谢途径调控
深入研究色氨酸等代谢途径的调控机制,揭示代 谢物与基因表达的相互作用关系,为代谢工程和 合成生物学提供新的理论和方法。
04
研究方法和实验结果
介绍本研究所采用的研究方法、实验设计和实验结果 ,以及对实验数据的分析和解读。
05
研究意义和展望
总结本研究的意义和贡献,以及对未来研究方向的展 望和建议。
02
原核表达调控概述
原核生物基因表达特点
80%
转录与翻译偶联
原核生物的转录和翻译过程在时 间和空间上紧密偶联,转录未完 成时翻译已经开始。
催化RNA合成的酶,其活性受到多种因子的调控。
其他调控因子
03
如小分子代谢物、环境因素(温度、pH值)等也能影响原核生
物的基因表达。
03
色氨酸操纵子结构与功能
色氨酸操纵子结构组成
阻遏蛋白
结合到操纵基因上,阻止RNA聚合酶的结合 和转录的起始。
结构基因
编码色氨酸生物合成所需的酶。
操纵基因
与阻遏蛋白结合的区域,控制转录的起始。
转化与筛选
将重组质粒转化入宿主细胞, 通过选择性培养基筛选阳性克 隆。
诱导表达
在含有合适诱导剂的培养基中 培养阳性克隆,诱导目标蛋白 的表达。
表达产物检测
通过SDS-PAGE、Western blot等方法检测目标蛋白的表 达情况。
数据收集和处理方法
色氨酸操纵子
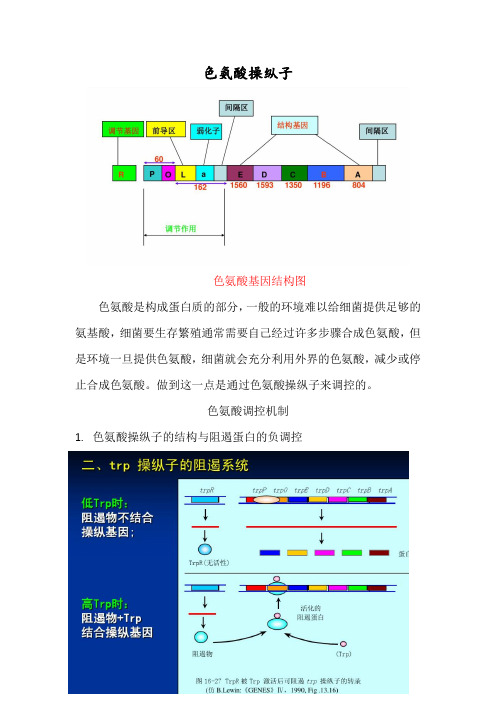
色氨酸操纵子
色氨酸基因结构图
色氨酸是构成蛋白质的部分,一般的环境难以给细菌提供足够的氨基酸,细菌要生存繁殖通常需要自己经过许多步骤合成色氨酸,但是环境一旦提供色氨酸,细菌就会充分利用外界的色氨酸,减少或停止合成色氨酸。
做到这一点是通过色氨酸操纵子来调控的。
色氨酸调控机制
1.色氨酸操纵子的结构与阻遏蛋白的负调控
如图所示:在调控色氨酸合成的结构基因上游有一个操纵基因trpR ●在低色氨酸浓度时,trpR控制的阻遏蛋白无活性,下游的结构基
因可正常转录翻译。
●在高色氨酸浓度时,trpR控制的阻遏蛋白具有活性。
能与trpO特
异性结合,阻遏结构基因的转录。
从而阻遏体内的色氨酸合成。
2.衰减子的作用
当色氨酸达到一定程度,但没有高到能够活化阻遏蛋白使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,靠着衰减子来调控。
如图所示:在高色氨酸时,trp mRNA在第一个trp E基因开始转录之前即停止生长。
低色氨酸时,mRNA正常转录。
这是因为在色氨酸操纵元trp O与第一个结构基因trp E 之间有一段前导序列。
高色氨酸时转录就会停止在这里。
如图所示:
在低浓度色氨酸条件下,2-3形成发卡结构,不含有U区域,不会形成终止子结构,不会停止转录,继续转录翻译形成色氨酸在高浓度色氨酸条件下,3-4会形成发卡结构,含有U区域,形成终止子结构,停止转录,阻遏色氨酸的合成。
第2节 色氨酸操纵子

内容提要: 色氨酸操纵子的结构 色氨酸操纵子的阻遏系统 色氨酸操纵子的弱化机制
一、色氨酸操纵子的结构
调控基因
结构基因
trpR
催化分枝酸转变为色氨酸的酶
分支酸 → 邻氨基苯甲酸 → 磷酸核糖基 → CDRP → 吲哚甘油-磷酸 → 色氨酸 邻氨基苯甲酸
邻氨基苯甲酸合成酶
RNA聚合酶 结构基因
5’
前导肽
23
核1 糖体
2 43
4
UUUU…U…UUU……
trp 密码子 序列3、4不能形成衰减子结构
2.当色氨酸浓度低时
High Trp Low Trp
弱化机制
高Trp时: Trp-tRNATrp 存在
核糖体通过片段1(2个Trp密码子) 封闭片段2
片段3,4形成发夹结构 类似于不依赖ρ因子的转录终止序列
Leader peptide
夹结构 / 富含 C G
U 的单链末端 C G
Aaaaaa C G
Met Lys Aly Ile Phe Val Leu Lys Gly Trp Trp Arg Thr Ser
A
GC
CG
A
CG
UU
AA
图 16-28 trp 操纵子含有 5 个结构基因和 1 个控制区。控制区由启动子、操纵基因、前导顺序和衰减子 构成。前导区编码 14 个氨基酸,其中有 2 个是色氨酸。(仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.38)
四、原核生物转录的整体调控模式
由成群的操纵子组成的基因转录调控网络称为调 节子。通过组成调节子调控网络,对若干操纵子 及若干蛋白质的合成进行协同调控,从而达到整 体调控的目的。
色氨酸操纵子与负控阻遏系统

被称为适应性表达基因,或被称为奢侈基因
基因的表达调控方式
➢ 基因水平的调控
➢ 转录水平的调控
➢ 转录产物加工的调控
➢ 翻译水平的调控以及翻译后的加工等
原核基因表达调控分类
根据调控机制: 负转录调控 调节基因编码阻遏蛋白,阻止结构基因转 录 分为负控诱导,负控阻遏 正转录调控 调节基因编码激活蛋白,促进结构基因转 录。 分为正控诱导,正控阻遏
鼠伤寒沙门氏菌中已陆续发现不少操纵子都有弱化
致转录终止。当色氨酸浓度较低时,TRAP失活,转录可以继
现象。 弱化子(attenuator)是指原核生物操纵子中能显著 减弱甚至终止转录作用的一段核苷酸序列,该区域
续,结构基因得以表达。另外枯草杆菌对未负荷色氨酸的 tRNATrp也很敏感,后者大量堆积,会诱导合成抗TRAP 蛋白 (anti -PRAP,AT)。AT与Trp激活的PRAP结合,可以取消其 转录终止活性。trpG表达也受PRAP调控,活化的TRAP与和
空 白 演 示 水平可提高6倍。研究发现,当mRNA开始合成后,
除非培养基中完全不含色氨酸,否则转录总是在这 个区域终止,产生一个仅有140个核苷酸的RNA分
养基中Trp 浓度很低时,负载有Trp 的tRNATrp也就少,这样 翻译通过两个相邻色氨酸密码子的速度就会很慢,当4区被转 录完成时,核糖体滞留1区,这时的前导区结构是2 - 3配对, 不形成3 - 4配对的终止结构,所以转录可继续进行。反之,核
不受葡萄糖或cAMP-CAP的调控。
空白演示
在此输入您的封面副标题
弱化子
弱化作用
是在研究大肠杆菌的色氨酸操纵子表达弱化现象 中发现的。在trp mRNA 5,端trp正基因的起始密 码前有一个长162 bp的DNA序列称为前导区,其
第六章-色氨酸(trp)操纵子3

The trp operator is a palindromic
DNA sequence
trp操纵子转录的调控是通过Trp阻遏物实现的,它结 合于trp操纵基因序列 ,但Trp阻遏物的DNA结合活性 直接受色氨酸调控,色氨酸结合Trp阻遏物,并起着 一个效应分子的作用(也称之辅阻遏物)。
在有高浓度色氨酸存在时,Trp阻遏物-色氨酸复合物 形成一个同源二聚体,并且紧密结合于trp操纵基因 序列,因此可以阻止转录。然而当色氨酸水平低时, 缺少色氨酸的Trp阻遏物以一种非活性形式存在,不 能结合DNA。在这样的条件下,trp操纵子被RNA聚 合酶转录,同时色氨酸生物合成途径被激活。
而当培养基中色氨酸浓度高时,核糖体可顺利 通过两个相邻的色氨酸密码子,在4区被转录 之前,核糖体就到达2区,这样使2-3不能配对, 3-4区可以自由配对形成茎-环状终止子结构, 转录停止,trp操纵子中的结构基因被关闭而 不再合成色氨酸(图6-24)。所以,弱化子对 RNA聚合酶的影响依赖于前导肽翻译中核糖体 所处的位置。
转录弱化作用
转录的弱化理论认为mRNA转录的终止是通过前导 肽基因的翻译来调节的。因为在前导肽基因中有两 个相邻的色氨酸密码子,所以这个前导肽的翻译必 定对tRNATrp的浓度敏感。
当培养基中色氨酸的浓度很低时,负载有色氨酸的 tRNATrp也就少,这样翻译通过两个相邻色氨酸密 码子的速度就会很慢,当4区被转录完成时,核糖 体 才 进 行 到 1 区 ( 或 停 留 在 两 个 相 邻 的 trp 密 码 子 处),这时的前导区结构是2-3配对,不形成3-4配 对的终止结构,所以转录可继续进行,直到将trp 操纵子中的结构基因全部转录。
trpE基因是第一个被翻译的基因,和trpE紧邻的是启 动子区和操纵区。另外,前导区和弱化子区分别定名为 trpL和trpa(不是trpA)。
16.3.3 色氨酸操纵子

基础生物化学Basic Biochemistry色氨酸(Trp)操纵子色氨酸操纵子是用来编码生成色氨酸的重要元件之一。
研究表明当有足够的Trp时,操纵子自动关闭,细菌直接利用外界的Trp。
缺乏Trp时,Trp操纵子被打开,5个结构基因表达,产生3个酶催化分支酸合成为Trp。
1、阻遏蛋白的负调控合成Trp的酶,需要5 个基因E、D、C、B和A的共同编码。
这5个基因的表达受上游启动子p和操纵基因o的调控。
在远离p-o-结构基因群的位置有一个调控基因TrpR,它能低水平的表达阻遏蛋白R’。
R’并无活性,当提供足够的Trp时,Trp与R’结合使其构象改变而成为有活性形式R,R与O特异性结合,阻遏结构基因的转录。
2、Trp操纵子的衰减调控⏹前导序列编码了一个14个氨基酸的前导肽;前导肽的第10、11位是相邻的两个Trp密码子。
⏹先导序列后半段含有4个彼此互补的区域(1、2、3、4),在被转录生成mRNA时相互间能形成发夹结构。
原核生物转录和翻译几乎同时进行,当转录起始后,RNA聚合酶沿DNA转录合成mRNA,同时核糖体结合在mRNA上开始翻译。
UUUU (34)UUUU 3’34核糖体前导肽前导mRNA当色氨酸浓度高时转录衰减机制125’trp 密码子衰减子结构就是终止子可使转录前导DNAUUUU 3’RNA 聚合酶终止UUUU (34)2423UUUU ……核糖体前导肽前导mRNA15’trp 密码子结构基因前导DNA RNA 聚合酶当色氨酸浓度低时Trp 合成酶系相关结构基因被转录序列3、4不能形成衰减子结构乳糖操纵子和色氨酸操纵子的比较Lac操纵子负责营养碳源的分解,只有当需要消耗乳糖时,才通过诱导物使阻遏蛋白失活而开放,是可诱导的负调控基因;此外还存在CAP的正调控。
trp操纵子负责Trp的合成,平时开放,调节基因的产物使其关闭,是可阻遏的负调控;此外还存在翻译与转录耦联的衰减子调控手段。
第三节 色氨酸操纵子

制作:王庆容
一、结构及调控模式 1.结构 2.粗调节
3.细微调控
二、弱化子与前导肽 1.弱化子attenuator :
位于trp mRNA 5‘端trpE基因的起始密码前,调 节已启动转录的trp操纵子是否继续下去的特定区域。
2.前导肽Leader peptide:
当trp浓度较高时, trp操纵子转录产生一个 140bp的RNA分子的前导序列,它编码的一个14个 AA的短肽。
3.mRNA 前导区的序列分析 4.转录弱化作用 三、trp操纵子弱化机制的实验依据 四.小结
R
P
O
L
a
E
D
C
B
A
RNA pol. mRNA
R'
R
P
O
L
a
E
D
C
B
A
mRNA
R'
色氨酸
色氨酸合酶 α和β亚基
色氨酸操纵子各组份详图
组 成
R P O
L a
间 隔 序 列
E
D
C
B
A
碱 基 数 功 能
未知 辅 阻 遏 蛋 白
60
162
1560
邻氨 基苯 甲酸 合酶
1593
ห้องสมุดไป่ตู้1350
1196
804
粗 开 关
微调控 前导肽
邻氨基苯 吲哚甘 甲酸磷酸 油磷酸 核糖转移 合酶 酶
色氨酸合酶α 和β亚基
R
RNA pol.
间隔区 调节基因 前导区 弱化子 结构基因 间隔区
60 R P O L a 162 E 1560 D 1593 C 1350 B 1196 A 804
大肠杆菌的色氨酸操纵子

1.色氨酸操纵子结构
(1)结构基因
E
D
编码5种酶,在催化分支酸转变为
C
B
L-色氨酸的过程中发挥作用
A
L
转录产物是先导mRNA
(2)调控元件
启动子(P)
操纵基因(O)
2.色氨酸操纵子表达的调控 阻遏物对色氨酸操纵子的负调控
衰减作用对色氨酸操纵子的调控 (1)阻遏物对色氨酸操纵子的负调控 ①阻遏物:同二聚体蛋白质 ②阻遏物本身不能与操纵基因O结合,必须 和色氨酸结合才能与O结合,而阻遏结构基 因的表达 ③色氨酸是一种共阻遏物
③cAMP通过和一种称为分解物基因激活物蛋 白质(CAP)形成cAMP-CAP复合物。
④cAMP-CAP复合物与乳糖操纵子的调控元件 结合后,促发结构基因的转录,因此CAP 是一种转录起始的正调控物。
⑤cAMP-CAP对转录的激活可能是通过直接与 RNA聚合酶作用,或其与启动子的结合来 改变启动子的构象,易于形成开放的RNA 聚合酶起始复合物
(3)组装成转录起始复合体的模型过程 a.TEIID(包括TBP和TAFs)同启动子结合。 b.RNA聚合酶II全酶再与TFIID结合形成起 始复合体。
SRBs
polII
RNA聚合酶
TFIIF
TFIIB
II全酶
TFIIE
激活蛋白
TFIID
TBP TAFs
以RNA聚合酶II全酶为单位组装成的RNA转录起始复合体
4.乳糖操纵子的复合调控 (1)乳糖阻遏蛋白参与负调控 (2)环化AMP受体蛋白(CAP)参与下的正调控 ①分解物阻遏作用:
E.coli在含有葡萄糖的培养基生长时,一些
分解代谢酶(半乳糖苷酶等)的水平很低,这种 葡萄糖对其他酶的抑制效应称为分解物阻遏作用。 与cAMP有关。 ②当细菌处于缺乏葡萄糖的培养环境中,cAMP浓 度升高,而在含有葡萄糖、半乳糖的培养环境中 加入cAMP,对乳糖代谢有关的基因的阻遏即被消 除。
细胞、分子与基因_ 细胞与分子3_18 色氨酸操纵子_

前导序列mRNA的特点:
前导区L可转录生成一段长为162碱基的前 导mRNA、 其中含有4个富含G 、C碱基的 特殊序列
前导序列mRNA的特点:
前导肽编码区:前导序列mRNA 第27 ~ 68碱基,包 含序列1 ,编码14aa组成的前导肽,其中第10、11 位密码子为两个连续的色氨酸密码子;
1
2
3
4
UUUU……
trp 密码子 终止密码子
14aa前导肽编码UU区U:U包…含…序列1 第10、11密码子为trp密U码U子UU…… 衰减子结构
是否形成衰减子结构,取决于前 导肽合成时核糖体所处的位置,
与色氨酸浓度有关
UUUU……
三、转录衰减机制
前导DNA
前导mRNA
1 5’
核糖体
RNA聚合酶
弱化子能较快地通过抗终子的方法来增加Trp结 构基因的表达,迅速提高内源色氨酸的浓度。
四、阻遏调控与
转录衰减的协调
细菌通过弱化作用弥补阻遏作用的不足, 因为阻遏调控是一个粗调开关,主管转 录是否启动;弱化作用是细微调控,决 定已经启动的转录是否进行下去。在细 菌细胞内这两种作用相辅相成,体现着 生物体内周密的调控作用。
色氨酸操纵子的结构 色氨酸操纵子的阻遏调控 色氨酸操纵子的转录衰减 阻遏调控与转录衰减的协调 衰减子的生物学意义
一、色氨酸操纵子结构
P
162 bp
L
结构基因
trpR
PO
a trpE trpD trpC trpB trpA
123~150
mRNA
辅阻遏蛋白
邻氨基苯 邻氨基苯甲 吲哚甘油 色氨酸合酶 甲酸合酶 酸磷酸核糖 磷酸合酶 α和β亚基
18.2.2原核基因表达调控——色氨酸操纵子

18.2.2 原核基因表达调控—色氨酸操纵子Regulation of Gene Expression inProkaryote ---trp operon目录操纵子有两种类型诱导操纵子:即诱导基因,这些基因因环境中某些物质的出现而被活化。
许多负责糖分解代谢的基因属于这种类型,如乳糖操纵子。
阻遏操纵子:即阻遏基因,一般情况下处于表达状态,但当其产物大量出现时即关闭,合成氨基酸的操纵子属于这一类型,如色氨酸操纵子。
目录目录Trp Trp 高时Trp 低时mRNAtrpE trpD trpC trpB trpAO P trpR 调节区结构基因RNA聚合酶RNA 聚合酶色氨酸操纵子通过阻遏蛋白调控基因表达色氨酸操纵子目录色氨酸操纵子(trp operon )的调控模式:阻遏蛋白的调控(粗调)和转录衰减(微调)。
转录衰减(attenuation )是转录-翻译的偶联调控目录调节区结构基因trpR O P 前导序列UUUU……前导mRNA 1234终止密码子序列1有独立的起始和终止密码子,可翻译成为有14个氨基酸残基的前导肽,它的第10位和第11位都是色氨酸残基。
trp 密码子trpE trpD trpC trpB trpA 目录UUUU……UUUU……调节区结构基因trpR O P 前导序列衰减子区域UUUU……前导mRNA 1234衰减子结构形成发夹结构能力强弱:序列1/2>序列2/3>序列3/4UUUU……trpE trpD trpC trpB trpA目录UUUU……342423UUUU……核糖体前导肽mRNA15’trp 密码子结构基因DNA RNA 聚合酶1.当色氨酸浓度低时Trp 合成酶系相关结构基因被转录序列3、4不能形成衰减子结构目录UUUU……34UUUU 3’34核糖体前导肽mRNA 2.当色氨酸浓度高时转录衰减机制125’trp 密码子衰减子结构就是终止子可使转录DNAUUUU 3’RNA 聚合酶终止目录原核生物这种在色氨酸浓度高时,通过阻遏作用和转录衰减机制共同关闭基因表达的方式,保证了营养物质和能量的合理利用。
色氨酸操纵子、弱化子动画

转录衰减机制
前导DNA
RNA聚合酶
前导mRNA
4
UUUU 3’
衰减子结构 就是终止子 可使转录 终止
5’
1
核糖体
3 2
3
UUUU 3’ UUUU……4ຫໍສະໝຸດ trp 密码子前导肽
1.当色氨酸浓度高时
前导DNA
Trp合成酶系相关 结构基因被转录
RNA聚合酶 结构基因
前导mRNA
2 3 2 4
5’
核糖体 1
3
UUUU…… UUUU……
4
trp 密码子 前导肽
序列3、4不能形成衰减子结构 2.当色氨酸浓度低时
三、其他转录调节机制
(一)转录衰减
调节区
trpR
结构基因
RNA聚合酶 O P RNA聚合酶
Trp 低时
mRNA
Trp 高时 Trp
色氨酸操纵子
调节区
trpR
结构基因
P
O 前导序列 衰减子区域
前导mRNA
1
2
3
4
UUUU……
trp 密码子
终止密码子
14aa前导肽编码区: 包含序列1 UUUU…… 衰减子结构 UUUU…… 第10、11密码子为trp密码子 UUUU…… 形成发夹结构能力强弱: 序列1/2>序列2/3>序列3/4
色氨酸操纵子ppt

大家有疑问的,可以询问和交流
可以互相讨论下,但要小声点
2、前导序列:在trp mRNA5‘端trpE基因得起始密码 前一个长162bp得mRNA片段。
调节区
trpR
PO
前导序列
前导mRNA
1
2
结构基因
衰减子区域
3
4
UUUU……
trp 密码子 终止密码子
1
2 第141a0a、前1导1密肽码编子码UU为区Ut:Ur包…p密含…U码序U子U列U1……衰减子结构
色氨酸得调节
转录衰减
调节区
trpR RNA聚P合酶O
RNA聚合酶
Trp 低时
结构基因
mRNA
Trp 高时
Trp
色氨酸操纵子
三、trp 操纵子得弱化机制
衰减子(attenuator)/弱化子 前导序列(leader sequence)
1、衰减子/弱化子:DNA中可导致转录过早终止得一 段核甘酸序列(123-150区)。
RNA聚合酶 结构基因
5’
前导肽
23
核1 糖体
2 43
4
UUUU…U…UUU……
trp 密码子 序列3、4不能形成衰减子结构
2、当色氨酸浓度低 时
High Trp Low Trp
弱化机制
高Trp时: Trp-tRNATrp 存在
核糖体通过片段1(2个Trp密码子) 封闭片段2
片段3,4形成发夹结构 类似于不依赖ρ因子得转录终止序列
色氨酸操纵子
一、色氨酸操纵子得结构
调控基因
结构基因
trpR
催化从分支酸到合成色氨酸所需得酶
分支酸 → N-氨基苯甲酸 →→→→→→吲哚-3-甘油-磷酸 → 色氨酸
色氨酸操纵子

色氨酸操纵子色氨酸是构成蛋白质的组分,一般的环境难以给细菌提供足够的色氨酸,细菌要生存繁殖通常需要自己经过许多步骤合成色氨酸,但是一旦环境能够提供色氨酸时,细菌就会充分利用外界的色氨酸、减少或停止合成色氨酸,以减轻自己的负担。
细菌这种对色氨酸利用的调节是通过色氨酸操纵子(trp operon)来实现的。
一、色氨酸操纵子的结构与阻遏蛋白的负性调控色氨酸操纵子的结构与乳糖操纵子相似,结构基因由合成色氨酸所需要酶类的基因E、D、C、B、A等头尾相接串连排列组成,结构基因上游为启动子P trp 和操纵序列O,不过其调控基因trpR的位置远离P-O-结构基因群,在其自身的启动子作用下,以组成性方式低水平表达其编码分子量为47KD的调控蛋白R。
点击后看大图色氨酸操纵子是属于一种负性调控的、可阻遏的操纵子。
以组成性方式低水平表达的阻遏蛋白R并不具有与O结合的活性,只有当环境能提供足够浓度的色氨酸时,R与色氨酸结合后构象变化,才能够与操纵序列O特异性亲和结合,阻遏结构基因的转录。
因此这类操纵子通常是开放转录的,有效应物(色氨酸为阻遏剂)作用时则关闭转录。
细菌不少生物合成系统的操纵子都属于这种类型,其调控可使细菌处在生存繁殖最经济最节省的状态。
二、衰减子及其作用实验观察表明:当色氨酸达到一定浓度、但还没有高到能够活化R使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,而且产生的酶量与色氨酸浓度呈负相关。
仔细研究发现这种调控现象受转录衰减(attenuation)机制的调节。
在色氨酸操纵子P trp-O与第一个结构基因trpE之间有一段162bp的前导序列构成衰减子区域(attenuator region),研究证明当色氨酸有一定浓度时,RNA 聚合酶的转录会终止在这里。
这段序列能够编码14个氨基酸的短肽,其中有2个色氨酸相连,在此编码区前有核糖体识别结合位点(RBS)序列,提示这段短序列在转录后是能被翻译的。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
蛋白 TrpR(无活性)
高Trp时: 阻遏物+Trp 结合操纵基因
阻遏物
活化的 阻遏蛋白
(Trp)
图 16-27 TrpR 被 Trp 激活后可阻遏 trp 操纵子的转录 (仿 B.Lewin:《GENES》Ⅳ,1990, Fig .13.16)
6
色氨酸的调节
转录衰减
调节区
trpR RNA聚P合酶O
2
3
UUUU……
形成发夹结构能力强弱:
3
4
序列1/2>序列2/3>序列3/4
14
前导DNA
转录衰减机制
前导mRNA
1 5’
核糖体
RNA聚合酶
UUUU 3’
衰减子结构
就是终止子
4 可使转录 终止
3
2
34
UUUU 3U’UUU……
前导肽
trp 密码子
1.当色氨酸浓度高时
15
前导DNA 前导mRNA
Trp合成酶系相关 结构基因被转录
10
邻氨基苯 吲哚甘油 色氨酸合成酶 甲酸合成酶 磷酸合成酶
TrpE terpD trpC trpB trpA t
t’
启动子 操纵基因
前导序列 衰减子
pppN26AUGAAAGCAAUUUUCGUACUGAAGGUUGGUGGCGCACUUCCUGAN43A UUUUUUUU 富含 G-C 的发 G C
4
特点:
(1) trpR和trpABCDE不连锁;
(2) 操纵基因(操作子)和启动子部分重叠 (3) 有衰减子(attenuator)/弱化子 (4) 启动子和结构基因不直接相连,二者被
前导序列(Leader)所隔开
5
二、trp 操纵子的阻遏系统
低Trp时: 阻遏物不结合 操纵基因;
trpR
trpP trpO trpE trpD trpC trpB trpA
RNA聚合酶停止转录,产生衰减子转录产物 转录、翻译偶联,产生前导肽
19
低Trp时: Trp-tRNATrp 没有供应
核糖体翻译停止在片段1 (2个Trp密码子)
片段2,3 形成发夹结构
转录不终止
RNA聚合酶继续转录
20
细菌通过弱化作用弥补阻遏作用的不足,因为阻 遏作用只能使转录不起始,对于已经起始的转录, 只能通过弱化作用使之中途停下来。阻遏作用的信 号是细胞内色氨酸的多少;弱化作用的信号则是细 胞内载有色氨酸的tRNA的多少。它通过前导肽的翻 译来控制转录的进行,在细菌细胞内这两种作用相 辅相成,体现着生物体内周密的调控作用。
吲哚-3-甘油 磷酸合成酶
色氨酸合成酶
β链
α链
60,000 O L 间隔区 trpE
60,000 trpD
4 5,000 50,000 29,000
trpC
trpB
trpA
P
P:启动子;O:操作子; L:前导序列; E.coli trpO 的结构及其产物所催化的色氨酸合成反应
3
乳糖操纵子的基本结构与调控原理
11
2、前导序列:在trp mRNA5‘端trpE基因的起始密码 前一个长162bp的mRNA片段。
12
13
调节区
trpR
PO
前导序列
前导mRNA
1
2
结构基因
衰减子区域
3
4
UUUU……
trp 密码子 终止密码子
1
2 第141a0a、前1导1密肽码编子码UU为区Ut:Ur包…p密含…U码序U子U列U1……衰减子结构
RNA聚合酶 结构基因
5’
前导肽
23
核1 糖体
2 43
4
UUUU…U…UUU……
trp 密码子 序列3、4不能形成衰减子结构
2.当色氨酸浓度低时
16
High Trp Low Trp
17
弱化机制
18
高Trp时: Trp-tRNATrp 存在
核糖体通过片段1(2个Trp密码子) 封闭片段2
片段3,4形成发夹结构 类似于不依赖ρ因子的转录终止序列
21
色氨酸操纵子的调控机制
阻遏调控 弱化(衰减调控) ●前导序列L含有四段特殊序列,序列1前有一段前
导肽。其10-11位是连续的色氨酸; ●序列2-3或3-4可形成茎环结构,3-4之间的茎环结
构是一个转录终止子结构(衰减子) ●Trp缺乏时,核糖体的翻译停滞在11-12位色氨酸密
码处,序列2-3形成茎环结构,3-4就不能形成终止 子,结构基因转录继续进行。反之,停止。
RNA聚合酶
Trp 低时
Trp 高时 Trp
结构基因
mRNA
色氨酸操纵子
7
三、trp 操纵子的弱化机制 衰减子(attenuator)/弱化子 前导序列(leader sequence)
8
1、衰减子/弱化子:DNA中可导致转录过早终止的 一段核甘酸序列(123-150区)。
123~150
9
研究引起终止的mRNA碱基序列,发现该区mRNA通过自 我配对可以形成茎-环结构,有典型的终止子特点。
Leader peptide
夹结构 / 富含 C G
U 的单链末端 C G
Aaaaaa C G
Met Lys Ala Ile Phe Val Leu Lys Gly Trp Trp Arg Thr Ser
A
GC
CG
A
CG
UU
AA
图 16-28 trp 操纵子含有 5 个结构基因和 1 个控制区。控制区由启动子、操纵基因、前导顺序和衰减子构 成。前导区编码 14 个氨基酸,其中有 2 个是色氨酸。(仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.38)
22
重点
Байду номын сангаас 色氨酸操纵子的阻遏调控机理 色氨酸操纵子的衰减(弱化)调控机理
23
第四节 色氨酸操纵子(Trp operon)
内容提要: 色氨酸操纵子的结构 色氨酸操纵子的阻遏调控机制 色氨酸操纵子的弱化调控机制
1
一、色氨酸操纵子的结构
调控基因
结构基因
trpR
催化从分支酸到合成色氨酸所需的酶
2
分支酸 → N-氨基苯甲酸 →→→→→→吲哚-3-甘油-磷酸 → 色氨酸
邻氨基苯甲酸合成酶