11第十一章-细胞质遗传
细胞质遗传PPT课件

.
8
母性影响的表现与细胞质遗传相似,但不 是由于细胞质基因组所决定的,而是由于 核基因的产物在卵细胞中积累所决定的, 例:椎实螺外壳旋转方向的遗传 椎实螺外壳旋转方向由一对基因控制
右旋(S+)对左旋(S)为显性 母本基因型→卵细胞状态→后代表现型 F3出现3:1分离,故又叫做延迟遗传
.
9
S+S+ S+S+
P IjIj♀ ijij♂
绿色
条纹
F1
Ijij 全部绿色
F2 IjIj Ijij ijij
3绿:1白条
.
17
P ijij ♀ IjIj ♂
条纹
绿色
F1 Ijij Ijij Ijij
绿条白
♀ Ijij IjIj绿
IjIj IjIj IjIj Ijij Ijij Ijij
绿条 白绿条白
.
18
二、叶绿体遗传的分子基础
.
3
真核生物细胞质中的遗传物质主要存在于: 线粒体、质体、中心体等细胞器中。
在原核和某些真核生物的细胞质中,除细胞 器外,还有另一类称:附加体(episome)和共 生体(symbiont)的细胞质颗粒,如,大肠肝菌 的F因子
细胞质基因组: 所有细胞器和细胞质颗 粒中遗传物质的统称
.
4
二、细胞质遗传的特点
20
1、低等植物的叶绿体基因组:
⑴ ct DNA仅能编码叶绿体本身结构和组成的一 部分物质; 如各种RNA、核糖体(70S)蛋白质、光合
作用膜蛋白和RuBp羧化酶8个大亚基。
⑵ 特性: 与抗药性、温度敏感性和某些营养缺陷有关。
.
21
2. 高等植物的叶绿体基因组:
第十一章细胞质遗传

下能稳定地遗传下去,经过多次接种移植、 其遗传方式和表现型都不会改变。
②. 试验: 缓慢生长特性只能在通过母本传递给
后代(其核遗传正常,呈1∶1分离)。
③.分析:缓慢生长突变型 在幼嫩培养阶段时无细胞 色素氧化酶,酶是正常代 谢所必需的。
某个体的表现型并不由其自身的基因型直接决定, 而是由母本卵细胞状态决定,母本卵细胞的状态又由 母本的基因型决定。
正交 ♀(右旋)SS×♂(左旋)ss
↓异体受精
反交 ♀(左旋)ss×♂(右旋)SS
↓异体受精
F1 全部为右旋(Ss)
全为左旋(Ss)
↓自体受精
↓自体受精
F2 1SS∶2Ss∶1ss(均右旋) 1SS∶2Ss∶1ss(均右旋)
接受花粉的枝条 提供花粉的枝条 杂种表现
白色
白色
绿色 花斑 白色
白色
绿色
绿色 花斑
绿色
白色
花斑
绿色 花斑
白、绿、 花 斑
结果: 白→白 绿→绿 花斑→白、绿、花斑
枝条和叶色的遗传物质是 通过母本传递的。
原因:
花斑枝条: 绿细胞中含有正常的绿色质体(叶绿体); 白细胞中只含无叶绿体的白色质体(白色体); 绿白组织交界区域:某些细胞内既有叶绿体、又有白色体。 ∴紫茉莉的花斑现象是叶绿体的前体(质体)变异而引起的。 花粉中精细胞内不含质体,上述三种分别发育的卵细胞无论接受
1984年,Lonsdale测出玉米mtDNA: * 含有570kbp; * 并存在多个重复序列(主要有6种); * rRNA基因、tRNA基因和细胞色素C氧
化酶基因均已定位。
㈢、线粒体遗传信息的复制、转录和翻译系统:
第十一章 细胞质遗传《遗传学》

第十一章细胞质遗传本章习题1、什么叫细胞质遗传?它有哪些特点?试举例说明之。
答:细胞质遗传指由细胞质内的遗传物质即细胞质基因所决定的遗传现象和规律,又称非染色体遗传、非孟德尔遗传、染色体外遗传、核外遗传、母体遗传。
细胞质遗传的特点:⑴. 遗传方式是非孟德尔式的;杂交后一般不表现一定比例的分离。
⑵. 正交和反交的遗传表现不同;F1通常只表现母体的性状,故又称母性遗传。
⑶. 通过连续回交能将母本的核基因几乎全部置换掉,但母本的细胞质基因及其所控制的性状仍不消失。
⑷. 由附加体或共生体决定的性状,其表现往往类似病毒的转导或感染。
举例:罗兹(Rhoades M. M.)报道玉米的第7染色体上有一个控制白色条纹的基因(ij),纯合的ijij植株叶片表现为白色和绿色相间的条纹。
以这种条纹株与正常绿色进行正反杂交,并将F1自交其结果如下:当以绿色株为母本时,F1全部表现正常绿色与非绿色为一对基因的差别,纯合隐性(ijij)个体表现白化或条纹,但以条纹株为母本时,F1却出现正常绿色、条纹和白化三类植株,并且没有一定的比例,如果将F1的条纹株与正常绿色株回交,后代仍然出现比例不定的三类植株,继续用正常绿色株做父本与条纹株回交,直至ij基因被全部取代,仍然没有发现父本对这个性状的影响,可见是叶绿体变异之后的细胞质遗传方式。
2、何谓母性影响?试举例说明它与母性遗传的区别。
答:由于母本基因型的影响,使子代表现母本性状的现象叫做母性影响,又叫前定作用。
母性影响所表现的遗传现象与母性遗传十分相似,但并不是由于细胞质基因组所决定的,而是由于核基因的产物在卵细胞中积累所决定的,故不属于母性遗传的范畴。
举例:如椎实螺外壳的旋转方向有左旋和右旋,这对相对性状是母性影响。
把这两种椎实螺进行正反交,F1外壳的旋转方向都与各自的母体相似,成为右旋或为左旋,但其F2却都有全部为右旋,到F3世代才出现右旋和左旋的分离。
这是由一对基因差别决定的,右旋(S+)对左旋(S)为显性,某个体的表现型并不由本身的基因型直接决定,而是由母体卵细胞的状态所决定,母本卵细胞的状态又由母本的基因型所决定。
第十一章 细胞质遗传

◆叶绿体基因组是存在于核基因组之外的另一 遗传系统, 遗传系统,它含有为数不多但作用不小的遗传 基因。 基因。 ◆但是,与核基因组相比,叶绿体基因组在遗 但是,与核基因组相比, 传上所起的作用是十分有限的: 传上所起的作用是十分有限的:
就叶绿体自身的结构和功能而言, 就叶绿体自身的结构和功能而言,叶绿体基因组 所提供的遗传信息仅仅是其中的一部分,对叶绿体十 所提供的遗传信息仅仅是其中的一部分, 分重要的叶绿素合成酶系、 分重要的叶绿素合成酶系、电子传递系统以及光合作 固定途径有关的许多酶类, 用中CO2 固定途径有关的许多酶类,都是由核基因编 用中 码的。 码的。
椎实螺卵裂方式 (a)右旋 (b)左旋 右旋 左旋
欧洲麦蛾( 欧洲麦蛾(Ephestia kuhuniella)色素的遗传 ) 欧洲麦蛾野生型幼虫的皮中含有色素, 欧洲麦蛾野生型幼虫的皮中含有色素,成虫复 眼为深褐色。这种色素是由犬尿氨酸所形成的, 眼为深褐色。这种色素是由犬尿氨酸所形成的, 由一对基因( )控制。 由一对基因(Aa)控制。
线粒体基因组大小变化较大, 哺乳动物的约 线粒体基因组大小变化较大,从哺乳动物的约 16kbp到高等植物的几百 的几百kbp(如玉米的为 如玉米的为570kbp) 到高等植物的几百 如玉米的为
(二)线粒体基因组的结构 ◆人、鼠和牛的mt 鼠和牛的 DNA显示相同的基本 显示相同的基本 遗传信息结构。 遗传信息结构。
第十一章 细胞质遗传
第1节 节 第2节 节 3节 第 3节 第4节 节 第5节 节 第6节 节 细胞质遗传的概念和特点 母性影响 叶绿体遗传 线粒体遗传 共生体和质粒的遗传 植物雄性不育的遗传
第1节 细胞质遗传的概念和特点 一 细胞质遗传的概念
◆由细胞质基因所决定的遗传现象和遗传规律 细胞质遗传(cytoplasmic inheritance), 叫做细胞质遗传 叫做细胞质遗传 , ◆细胞质遗传又称: 细胞质遗传又称 非染色体遗传(non-chromosomal inheritance) 非染色体遗传 非孟德尔遗传(non-Mendelian inheritance) 非孟德尔遗传 染色体外遗传(extra-chromosomal inheritance) 核外遗传(extra-nuclear inheritance) 核外遗传 母体遗传(maternal inheritance) 母体遗传
十一章细胞质遗传

50%致死 胚胎致死
♀♀
♂♂
图 20- 果蝇眼色的母体影响
置换。
• 由附加体或共生体决定的性状,其表现往
往类似病毒的转导或感染。
第三节 叶绿体遗传
1)1908,Carl Corrans,非孟德尔遗传发现 者,紫茉莉花斑枝条
(一)叶绿体系统的核外遗传
• 1909年由Carl Corrans发现(irabilis jalapa)
表 20-8 紫茉莉花斑植物杂交的结果
表 20-5 酵母线粒体中合成的酶亚基
酶
分子量
亚基
(Kda) 线粒体中合成
细胞质中合成
ATPase
340 ATPase 6、8、9 ATP 1、2、3、4、7
(对寡霉素敏
(FO 膜因子)
(F1 ATPase)
感)
细胞色素 C 氧 137
CO 1、2、3
CO 4、5、6、7
化酶
细胞色素 bc1 160 细胞色素 b 脱辅
LSC:long single copy sequence
IR: inverted repeats
LSC
Fig. 19- The chloroplast genome is a circle, divided by two inverted repeats into the short single copy sequence and long single copy sequence (GENES Ⅵ Fig24.3 )
酵母小菌落(peptite)突变
• 1940,Boris Ephrnssi • 在正常的细胞群体中有0.1%~1%的
细胞会自发变成小菌落。在插入剂 ( 如 : 溴 化 乙 锭 , EB ) 存 在 时 100% 的细胞都变成小菌落。
14遗传学第11章-细胞质遗传

§11 细胞质遗传Chapter Eleven Cytoplasmic Inheritance 核遗传(细胞核遗传)指染色体基因组所控制的遗传现象和遗传规律。
细胞质遗传指由细胞质遗传物质引起的遗传现象,又称非染色体遗传、非孟德尔遗传、染色体外遗传、核外遗传、母性遗传等。
细胞质基因组是所有细胞器和细胞质颗粒中遗传物质的统称。
在真核生物的有性过程中:卵细胞有细胞核、大量的细胞质和细胞器,为子代提供核基因和全部或绝大部分的细胞质基因。
精细胞只有细胞核,细胞质或细胞器极少或没有,为子代只能提供核基因,不能或极少提供细胞质基因。
因此,一切受细胞质基因所决定的性状,其遗传信息一般只能通过卵细胞传给子代,而不能通过精细胞遗传给子代。
代表两种细胞核代表两种细胞质代表两种质体A B B A细胞质遗传的特点1 正反交的遗传表现不同2 连续回交母本的性状不消失3 由附加体和共生体决定的性状表现类似病毒的转导或感染§11.1 母性影响§11.2 叶绿体遗传§11.3 线粒体遗传§11.4 植物细胞质雄性不育§11.1 母性影响1 概念正反交的结果不同,子代的表型受母亲细胞核基因型的影响而和母亲表型一样的现象叫母性影响。
2 种类短暂的母性影响:麦粉蛾持久的母性影响:椎实螺短暂的母性影响:麦粉蛾(Ephestia phestia)野生型:幼虫皮肤有色,成虫复眼深褐色(犬尿素)突变型:幼虫皮肤无色,成虫复眼红色(缺乏犬尿素)⏹图13-1持久的母性影响:椎实螺(Limnaea)⏹图11-2椎实螺外壳旋向的遗传椎实螺外壳旋向的遗传受母亲基因型制约正交反交椎实螺的受精卵是螺旋式卵裂,成体外壳的旋向决定于最初两次卵裂中纺锤体的方向。
1、2为第一次卵裂3、4为第二次卵裂1和3是左旋2和4是右旋母性影响与细胞质遗传的区别母性影响性状表现依赖于母方细胞核基因的作用,而这些基因是以经典方式传递的,它的特点是父方的显性基因延迟一代表现和分离。
11第十一章-细胞质遗传PPT课件

右旋
左旋
-
6
右旋
左旋
-
7
旋向取决于受精卵最初 几次卵裂纺锤体的方向
左旋
右旋
-
8
▪ 锥实螺旋壳方向的遗传属于核遗传范畴 ▪ 由于受精前母体卵细胞状态决定子代性状 表现的母性影响也称为前定作用
▪ 其孟德尔分离比例(3:1)在F3 代才表现 也称为延迟遗传
-
9
二. 面粉蛾的眼色
任何两个基因,一起被保留或丢失的
频率越高,连锁越紧密。
-
34
在酵母中一个由多种抗性(aptR barR cobR)的菌系被用来诱导 500个小菌落,其中人一对抗性标记中丢失了一种抗性的小菌落数 列于下表,请利用这些结果画一线粒体基因图。
基因对 apt bar apt cob bar cob
第一抗性基因丢失
暂时的母性影响,缺乏A基因,不能制造色素, 随个体发育色素逐渐消耗,成虫时已成为红色。
-
10
第三节 叶绿体和线粒体遗传
一. 叶绿体的遗传表现
1. 紫茉莉的花斑叶色遗传 2. 玉米叶片的埃型条纹遗传 3. 衣藻的核外遗传 4. 衣藻叶绿体基因作图
-
11
1. 紫茉莉花斑的遗传
亲本枝条
♀卵细胞×♂花粉 后代表型
↓ KK+卡 稳定放毒
Kk无卡 敏感
↓ KK无卡
敏感
kk+卡 不稳定放毒
kk无卡 敏感
Kk+卡 放毒
↓ KK+卡 稳定放毒
Kk卡 放毒
↓ KK+卡 稳定放毒
kk+卡 不稳定放毒
kk+卡 不稳定放毒
第五节 植物雄性不育性
遗传学第11章 细胞质遗传

●长度大小约80~600kb。最大的来自天竺葵 长度大小约 。最大的来自天竺葵cpDNA, , 只有85kb。 有 217kb,而一种绿藻的 ,而一种绿藻的cpDNA只有 只有 。 ●拷贝数:以显花植物为例,其每个叶肉细胞中约有 拷贝数:以显花植物为例 其每个叶肉细胞中约有 100 个叶绿体 每个叶绿体中又含有 个叶绿体,每个叶绿体中又含有 每个叶绿体中又含有100 多个质体基因组 拷贝。大麦:60个叶绿体 叶绿细胞,200个cpDNA分子 拷贝。大麦: 个叶绿体/叶绿细胞, 个 分子/ 个叶绿体 叶绿细胞 分子 叶绿体,> 分子/叶绿细胞 叶绿体,>10000个cpDNA分子 叶绿细胞;拟南芥:核 ,> 个 分子 叶绿细胞;拟南芥: 基因组、线粒体基因组和叶绿体基因组的比例 基因组、线粒体基因组和叶绿体基因组的比例=1:13:280
(2)叶绿体的序列结构与基因组成 ) ○反向重复序列(IRA和IRB) 反向重复序列( ○大单拷贝区(large single-copy region,LSC) 大单拷贝区( , ) ○小单拷贝区(small single-copy region,SSC) 小单拷贝区( , ) 含有87-183个基因,其中一半以上与蛋白质合 个基因, 含有 个基因 成有关。 成有关。
与紫茉莉花斑叶色一样, 与紫茉莉花斑叶色一样,天竺葵花斑叶性状也是通过细 胞质遗传的,但由于其精子中带有少量的细胞质, 胞质遗传的,但由于其精子中带有少量的细胞质,少数叶绿 精子中带有少量的细胞质 体则可以通过精子向后代传递,因此二者又是有区别的。 体则可以通过精子向后代传递,因此二者又是有区别的。 紫茉莉、 紫茉莉、天竺葵镶嵌花斑遗传的比较 紫 茉 莉 ♀ 亲 代 ♂ ♀ ♂ ♀ 天 竺 葵 ♂ ♀ ♂
绿色 × 花斑
第十一章 细胞质遗传第十一章 细胞质遗传

第十一章细胞质遗传第一节细胞质遗传的概念和特点一、细胞质遗传的概念由细胞质内的基因即细胞质基因所决定的遗传现象和遗传规律叫做细胞质遗传,有时又称非染色体遗传、非孟德尔遗传、染色体外遗传、核外遗传、母体遗传等。
真核生物的细胞质中的遗传物质主要存在于线粒体、质体、中心体等细胞器中。
通常把上述所有细胞器和细胞质颗粒中的遗传物质,统称为细胞质基因组。
二、细胞质遗传的特点细胞学的研究表明,在真核生物的有性繁殖过程中,卵细胞内除细胞核外,还有大量的细胞质及其所含的各种细胞器;精子内除细胞核外,没有或极少有细胞质,因而也就没有或极少有各种细胞器(图11-1)。
细胞质遗传的特点是:1、遗传方式是非孟德尔式的;杂交后代—般不表现一定比例的分离;2、正交和反交的遗传表现不同;F1通常只表现母本的性状,故细胞质遗传又称为母性遗传;3、通过连续回交能将母本的核基因几乎全部置换掉,但母本的细胞质基因及其所控制的性状仍不消失;4、由附加体或共生体决定的性状,其表现往往类似病毒的转导或感染。
第二节母性影响一、概念:母性影响:由核基因的产物积累在卵细胞中的物质所引起的,子代表现母本性状的遗传现象。
∴母性影响不属于胞质遗传的范畴,十分相似而已。
二、特点:下一代表现型受上一代母体基因的影响。
三、实例:椎实螺的外壳旋转方向的遗传。
椎实螺是一种♀、♂同体的软体动物,每一个体又能同时产生卵子和精子,但一般通过异体受精进行繁殖。
∴椎实螺即可进行异体杂交、又可单独进行个体的自体受精。
椎实螺外壳的旋转方向有左旋和右旋之分,属于一对相对性状。
第六节植物雄性不育的遗传植物雄性不育的主要特征是雄蕊发育不正常,不能产生有正常功能的花粉,但是它的雌蕊发育正常,能接受正常花粉而受精结实。
一、雄性不育的类别及其遗传特点可遗传的雄性不育性可分为核不育型和质核不育型等多种类型。
(一)核不育型由核内染色体上基因所决定的雄性不育类型,简称核不育型。
多属自然发生的变异。
第十一章 细胞质遗传

三 雄性不育性的利用
三 雄性不育性的利用
1. 真核生物在受精过程中, 真核生物在受精过程中,
不仅为子代提供
核基因,也为子代提供大部分细胞质基因;而 核基因,也为子代提供大部分细胞质基因; 则仅能为子代提供核基因, 则仅能为子代提供核基因,因此细胞质遗传与细胞 核遗传表现不同。 核遗传表现不同。 2. 玉米 系属于孢子体不育,假如某株基因型为Rr, 玉米T系属于孢子体不育,假如某株基因型为 , 系属于孢子体不育 产生的花粉有两种:一种含有 ,一种含有r, 产生的花粉有两种:一种含有R,一种含有 ,那 么基因为R的花粉育性为 么基因为 的花粉育性为 育性为 。 ,基因为r的花粉 基因为 的花粉
第6节 植物雄性不育的遗传
◆植物雄性不育(male sterility)的主要特征 是雄蕊发育不正常,但是它的雌蕊发育正常。 是雄蕊发育不正常,但是它的雌蕊发育正常。 免去人工去雄, 免去人工去雄,节约人力 保证种子纯度
第十一章 细胞质遗传(F)

第十一章细胞质遗传由细胞核内染色体上的基因即核基因所决定的遗传现象和遗传规律称为细胞核遗传或核遗传(nuclear inheritance)。
前面所介绍的遗传现象和规律都是由核基因所决定的。
早期遗传学曾经把染色体看作基因或遗传信息的唯一载体。
随着遗传学的发展逐渐证实,尽管核基因在遗传上占有重要和主导地位,但是细胞质不但是核基因发生作用的场所,而且存在着决定某些性状的遗传基因。
早在1909年柯伦斯(C.E.Correns)就报道了紫茉莉(Mirabilis jalapa)花斑叶色的遗传不符合孟德尔定律的遗传现象,但未引起重视。
以后在其他高等植物中也陆续报道了类似的核外遗传现象。
20世纪40年代初,有关红色面包霉、酵母和一些原生生物如草履虫、衣藻中核外遗传现象也被发现,人们推测细胞质中可能存在遗传物质。
但直到1963~1964年才获得了在线粒体和叶绿体中存在DNA的直接证据。
从此,核外遗传的研究逐渐成为遗传学中的重要领域之一。
这个领域的深入研究,对于正确认识核质关系,全面地理解生物遗传现象和人工创造新的生物类型具有重要意义。
第一节细胞质遗传的概念和特点一、细胞质遗传的概念由细胞质内的基因即细胞质基因所决定的遗传现象和遗传规律叫做细胞质遗传(cytoplasmic inheritance)。
研究发现,真核生物的细胞质中存在着一些具有一定形态结构和功能的细胞器,如线粒体、质体、核糖体等。
这些细胞器在细胞内执行一定的代谢功能,是细胞生存不可缺少的组成部分。
在原核生物和某些真核生物的细胞质中,除了细胞器外,还有另一类称为附加体(episome)和共生体(symbiont)的细胞质颗粒,它们是细胞的非固定成分,也能影响细胞的代谢活动,但它们并不是细胞生存必不可少的组成部分。
例如,果蝇的 (sigma)粒子、大肠杆菌的F因子以及草履虫的卡巴粒(Kappa particle)等,这些成分一般都游离在染色体之外,有些颗粒如F因子还能与染色体整合在一起,并进行同步复制。
遗传学 第11章细胞质遗传 PPT课件
ijij 有发动质体白化变异的作用,这个由隐性核基因所造成的质体变异具有不可 逆的性质,质体变异一经发生,便能以细胞质遗传的方式而稳定传递。 反交试验: P 条纹 × 绿色 ijij ↓ IjIj F1 绿色 Ijij 白化 Ijij 条纹 Ijij
绿色 IjIj 连续回交 ┌──┬──┬──┬──┬──┐ 绿色 条纹 白化 绿色 条纹 白化 IjIj IjIj IjIj Ijij Ijij Ijij (绿色、条纹、白化三类植株无比例) 连续回交 (轮回亲本为绿株),当ij基因被取代后,仍然未发现父本对这一性 状产生影响。 第11章 源自细胞质遗传
§1.细胞质遗传的概念和特点
一、细胞质遗传的概念
核遗传(nuclear inheritance): 以细胞核基因为基础遗传现象。 细胞质遗传(cytoplasmic inheritance): 以细胞质基因为基础的遗传现象。 细胞质基因主要在各种细胞器上,如质体、线粒体、 中心体、共生体,及游离体、附加体等,这些都在 细胞质内,所以称为细胞质遗传,又称核外遗传、 非染色体遗传、非孟德尔式遗传。
在遗传时,母本产生的卵细胞或胚囊既有细胞核又有细胞质,而父本 产生的精细胞不带细胞质或很少细胞质,后代的细胞质是母本提供的,
所以细胞质基因是通过母本来传递的,因而又称:母体遗传、母性遗传。
二、细胞质遗传的特征
1.正交和反交表现不相同 如紫罗兰,胚胎表皮的颜色 有: 深蓝色和黄色两个变种 ♀深蓝×黄色♂→F1 深蓝色 ♀黄色×深蓝♂→F1 黄色
§3 、线粒体的遗传
一、线粒体遗传的表现: ㈠、红色面包霉缓慢生长突变型的遗传: 两种接合型均可产生原子囊果(卵细胞)和分生孢子(精细胞)。 ①. 缓慢生长突变型:野生型形成正常菌落,有一种突变型的生长速度缓慢, 菌落相当于正常型的1/2~1/3,叫做缓慢生长型. ②. 试验: 缓慢生长特性只能在通过母本传递给后代(其核遗传正常,呈1∶1分离)。 红色面包霉在培养基中培养可以形成菌落,将这两种杂交,后代和母本相同。 ♀野生型 × 突变型♂ → F1 野生型 正常菌落 ♀突变型 × 野生型♂ → F1 突变型 小菌落 ③.分析:缓慢生长突变型在幼嫩培养阶段时无细胞色素氧化酶,酶是正常 代谢所必需的。 由于细胞色素氧化酶的产生与线粒体有关,且观察到缓慢生长突变型中的线 粒体结构不正常。 ∴有关基因存在于线粒体之中,由母本传递。
第11章细胞质遗传

S(rfrf) ×N或S(RfRf) → S(Rfrf) 可育 恢复系 S(rfrf) ×N或S(Rfrf) → S(Rfrf)可育 + S(rfrf) 不育 半恢复系
不育系: S(r r) 保持系: N(r r) 恢复系: N(RR)
S(RR)
• 表型 •M • MN •N
基因型 基因 人数 MM M 2235 MN M.N 4460 NN N 2579
Kk+卡 放毒
↓ KK+卡 稳定放毒 kk+卡 不稳定放毒
Kk卡 放毒
↓ KK+卡 稳定放毒 kk+卡 不稳定放毒
细胞质与细胞核遗传的异同
• 相同点
1、均按半保留方式遗传 2、表达方式一样 3、均能发生突变且稳定遗传 4、其诱导因素也一样
细胞质与细胞核遗传的异同
• 不同点:
质DNA 核DNA
突变频率较大具有较强的定向突变 ……小,难于定向突变 正反交不一样 正反交一样 Cell质常表现母体影响 不表现母体影响 基因定位难 根据杂交进行基因定位 基因分离无规律 基因分离有规律 细胞间分部不均匀 cell分部均匀
• P:正交 ♂Aa ×aa♀
反交♂Aa × aa♀ 1/2 Aa 1/2 aa
பைடு நூலகம்
• F1:1/2 Aa 1/2 aa
幼虫(有) 幼虫(无) 成虫(褐) 成虫(红)
幼虫(有) 幼虫(有) 成虫(褐) 成虫(红)
二. 面粉蛾的眼色
暂时的母性影响,缺乏A基因,不能制造色素, 随个体发育色素逐渐消耗,成虫时已成为红色。
第十一章细胞质遗传
• 细胞质遗传性质与特点 • 母体遗传 • 线粒体遗传 • 共生体遗传 • 植物雄性不育 • 基因型频率与基因频率
第11章 细胞质遗传 习题

第11章细胞质遗传习题一、名词解释:1.细胞质遗传(cytoplasmic inheritance):由细胞质内的基因即细胞质基因所决定的遗传现象和遗传规律叫做,又称染色体外遗传、核外遗传、母体遗传等。
2.母性影响(maternal effect):正反交的结果不同,子代表型受到母本基因型的影响而和母本的表型一样的现象。
又叫前定作用(predetermination)。
3.植物雄性不育:雄蕊发育不正常,不能形成有功能的正常花粉;而其雌蕊却是正常的,可以接受正常花粉而受精结实。
4.核不育型:是一种由核内染色体上基因所决定的雄性不育类型。
5.质-核不育不育型:由细胞质基因和核基因互做控制的不育类型。
6.孢子体不育:花粉的育性受孢子体(植株)基因型所控制的现象称为孢子体不育。
7.配子体不育:花粉的育性受受雄配子体(花粉)本身的基因所决定的现象称为配子体不育8.自体受精:同一个体产生的雌雄配子受精结合。
如:蚯蚓、草履虫等低等动物。
二、是非题:1.草履虫的放毒特性依赖于核基因K,因此,有K基因就是放毒型,否则就是敏感型。
(-)2.细胞质遗传的一个特点是杂种后代的性状通常表现不分离或不规则分离。
(-)3.利用化学药物杀死一个正常植株的花粉,它的雌花与正常花粉授粉,受精获得的子代也就能表现出雄性不育的特性了。
(-)4.已知一个右旋的椎实螺基因型为Dd,它自体受精产生后代应该全部是左旋。
(-)5.在植物雄性不育性遗传中,配子的育性受母体基因型的控制的现象称为配子体不育。
(-)三、选择题:1.紫茉莉的枝条有绿色、白色和花斑三种不同颜色,其颜色的遗传属于细胞质遗传,用♀花斑×♂绿色,其后代表现为(4)(1)绿色;(2)白色;(3)花斑;(4)绿色,白色,花斑。
2.现代遗传学认为, 线粒体、质体等细胞器中含有自己的(4)。
(1)蛋白质(2)叶绿体(3)糖类物质(4):遗传物质3.质核互作型雄性不育系的恢复系基因型为(4)(1)S(Rr) (2)N(rr)(3)S(rr) (4)N(RR)4.植物质核型雄性不育中的孢子体不育类型,如基因型为Rr时,产生的花粉表现(3):(1)全不育(2)一半可育(3)全可育(4)穗上分离5.植物质核型雄性不育中的配子体不育类型,如基因型为Rr时,产生的花粉表现(2):(1)全不育(2)一半可育(3)全可育(4)穗上分离6.植物质核型雄性不育中的孢子体不育类型,如基因型为Rr时,产生的花粉表现(3):(1)全不育(2)一半可育(3)全可育(4)穗上分离7.植物质核型雄性不育中的配子体不育类型,如基因型为Rr时,产生的花粉表现(2):(1)全不育(2)一半可育(3)全可育(4)穗上分离8.所谓质-核不育是由质-核中相应基因的互作而引起的,水稻杂种优势利用中成功地利用了这种雄性不育,对于如下基因型哪些能组成理想的三系配套。
遗传学 第十一章 细胞质遗传

1 dd(右)
F3: 1DD 1/2DD 1 Dd 1/2dd 1 dd
(右) 比例:
(右) (右) (右) (左)
3
:1
P: ♂ DD(右) ♀ dd(左)
F1:
Dd(左)
F2:1DD(右)
2 Dd(右)
1 dd(右)
F3: 1DD
(右) 比例:
1/2DD 1 Dd 1/2dd 1 dd
(右) (右) (右) (左)
第四节 线粒体遗传
mtDNA与核DNA有明显的不同: (1)mtDNA与原核生物的DNA一样,没有重复序列; (2)mtDNA的浮力密度比较低; (3)mtDNA的碱基成分中G、C的含量比A、T少; (4)mtDNA的两条单链的密度不同,一条称为重链(H链) ,另一条称为轻链(L链); (5)mtDNA单个拷贝非常小,与核DNA相比仅仅是后者的 十万分之一。
目前已在270多种植物中发现有细胞质雄性不育现象。
细胞质型不育系的不育性都只能被保持而不能被恢复。
2.质─核不育型(cytoplasmic male sterility CMS): 概念:
由细胞质基因和核基因互作控制的不育类型。 遗传特点: 胞质不育基因为Sterility,S; 胞质可育基因为Normal,N; 核不育基因r,不能恢复不育株育性; 核可育基因R,能够恢复不育株育性。 只有S和r同时存在时才导致雄性不育
第四节 线粒体遗传
• 线粒体内有遗传信息的复制、转录和翻 译系统,有核糖体和各种RNA。
• 线粒体中有100多种蛋白质,其中只有 10种左右是线粒体自身合成的,其它蛋 白质都是由核基因组编码。
• 线粒体是半自主性的细胞器。
第五节 核质互作遗传
细胞质遗传

第十一章细胞质遗传1.基本概念(1)细胞质遗传:细胞器和细胞质颗粒中的遗传物质,统称为细胞质基因组。
把细胞质基因所决定的遗传现象和遗传规律称为细胞质遗传。
Cytoplasmic inheritance:Inheritance of genes in cytoplasm: the inheritance of genes from the female parent that are not in the nucleus but in organelles such as mitochondria that are found in the cytoplasm. This type of inheritance is not controlled by Mendel's laws.(2)母性影响:由核基因的产物积累在卵细胞中的所引起的一种遗传现象。
(3)母系遗传:杂交后代的表型和基因型与母本一致。
Maternal inheritance:A type of uniparental inheritance in which all progeny have the genotype and phenotype of the female parent.(4)(植物)雄性不育:植物雄性花粉不具有授粉能力。
Male sterility:The inability of the male to fertilise the ovum.(5)(雄性不育)保持系:与雄性不育系杂交,后代仍然为雄性不育系。
(6)(雄性不育)恢复系:与雄性不育系杂交,后代恢复为雄性可育。
2.基本问题(1)紫茉莉叶绿体的遗传现象及其机理。
实验现象:子代的性状是由母本细胞质基因决定的,即母系遗传。
机理:紫茉莉的白色叶片是由于缺乏正常的叶绿体,叶绿体的性状是由细胞质基因决定的,受精卵细胞质主要是由卵细胞提供的,所以对叶绿体遗传而言,其性状是由母本决定的。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
中性小菌落×大菌落→全是大菌落子囊孢子 细胞质 抑制性小菌落×大菌落→全是小菌落子囊孢子 小菌落
细胞质小菌落与线粒体有关
⑴ 电子传递系统中的变化
线粒体中负责ATP合成的电子呼吸链失去功能, 不得不依赖于能量产生效率较低的发酵途径。
⑵ 线粒体中蛋白质合成的变化
细胞质中某些质体改变,不能产生叶绿素 ↓
叶片呈绿白相间的条纹 ↓
表现细胞质遗传特征(不再需要ij基因)
质体具有一定程度的遗传自主性
3. 衣藻的核外基因遗传
单细胞绿色藻类 属于真核生物 含一个大的叶绿体 含较多的线粒体 两种交配型
正接合型mt+ 负接合型mt﹣
衣藻的生活周期
衣藻抗链霉素突变有两类:孟德尔方式遗传 细胞质遗传
apt
cob
bar
1: 3
第四节 其他细胞质遗传因子
一. 草履虫的放毒型特性 二. 细菌质粒 三. 真核生物的质粒 四. 果蝇体内感染因子对寄主细胞的影响
草履虫 (二倍体原生动物)
K: 决定细胞质中卡巴粒的增殖 k: 细胞质中卡巴粒不能增殖 卡巴粒: 释放草履虫素
有性接合
KK×kk→Kk
自体受精 Aa→AA or aa
抗药性来自小菌落
抗药性和决定大菌 落基因一起丢失
比较两个基因一起被保留或丢失的频 率可测知基因的连锁关系:
任何两个基因,一起被保留或丢失的
频率越高,连锁越紧密。
在酵母中一个由多种抗性(aptR barR cobR)的菌系被用来诱导 500个小菌落,其中人一对抗性标记中丢失了一种抗性的小菌落数 列于下表,请利用这些结果画一线粒体基因图。
杂合体细胞中形成芽时完成。 分离或重组可在二倍体的培养物中直接检测,
即通过观察此芽诱导成孢子时产生的减数分裂产物 检测 。
RF
=
7+1 36+48+7+1
= 6.7%
ⅱ. 丢失分析
利用小菌落诱导时标记基因的丢失来 检测位于mtDNA上的基因的连锁关系。
多个基因同时缺失 的小菌落突变体可 用于基因作图
m2 29.0 -
-
m3 18.0 10.9
-
m4 18.4 26.2
8.8
画出叶绿体基因的遗传图。
m1
18.0
29.0 m2
m3
26.2
m4
8.8 注:数字表示共分离频率
二. 线粒体的遗传表现
1. 红色面包霉的生长迟缓突变 2. 啤酒酵母的小菌落突变 3. 啤酒酵母线粒体基因作图
1. 红色面包酶的生长迟缓突变
第十一章 细胞质遗传
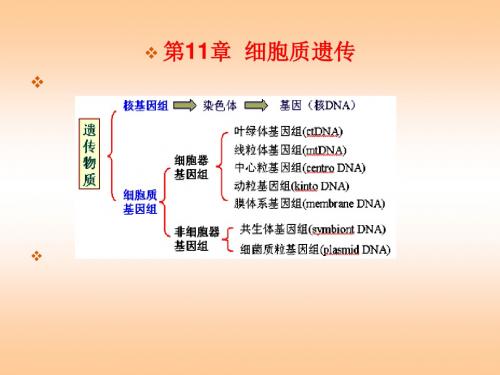
生物的遗传体系
核基因组——染色体——基因(核DNA)
遗传物质
(染色体基因组)
细胞器基因组——线粒体(mtDNA)
细胞质基因组
叶绿体(cpDNA)
非细胞器基因组——质粒DNA
共生体DNA
第一节 细胞质遗传的概念和特点
一. 概念 细胞质基因所决定的遗传现象和遗传规律,称
细胞质遗传。 染色体外遗传 核外遗传
正常线粒体中有独特的蛋白质合成装置,但细 胞质小菌落没有。
⑶ mtDNA组成的变化
中性小菌落缺乏线粒体DNA,抑制性小菌落 mtDNA的结构、组成均有较大改变
抗药性, 在二倍体芽经诱导形成孢子时, 减数分裂产物表现单亲遗传。
3. 啤酒酵母线粒体基因作图
ⅰ 重组分析 在酵母中,核外分离和重组是在在二倍体胞质
生化分析表明: 抗性来自叶绿体,只能由mt+亲本传递(单亲遗传)。
后来发现,mt﹣型的叶绿体在结合后总是被丢失 (5%共存,最终会分离)
4. 衣藻叶绿体基因作图
strr mt+ × strs mt﹣ ↓
0.1%后代同时含有strr和 strs (双亲合子体) 紫外线处理mt+ 亲本, 频率可达40—100%,
正交:poky(♀) ×野生型(♂)→ poky突变型
反交:野生型(♀) 生×活po周ky(期♂) →全为野生型
Poky突变体缺少细胞色素a和b, 但细胞色素c含量超过正常值
2. 啤酒酵母的小菌落突变
生活周期
1940年,Boris Ephrussi等发现小菌落突变, 从遗传方式上分成三类:
分离型小菌落 中性小菌落 抑制性小菌落
左旋
右旋
锥实螺旋壳方向的遗传属于核遗传范畴 由于受精前母体卵细胞状态决定子代性状 表现的母性影响也称为前定作用
其孟德尔分离比例(3:1)在F3 代才表现 也称为延迟遗传
二. 面粉蛾的眼色
暂时的母性影响,缺乏A基因,不能制造色素, 随个体发育色素逐渐消耗,成虫时已成为红色。
第三节 叶绿体和线粒体遗传
母性遗传(少数例外)
二. 特点
1. 正交、反交遗传表现不同。 F1的表型与母本相同, 后代表型分离不呈孟德尔比例。
2. 连续回交,细胞质基因及其控制的性状仍不消失。
3. 非细胞器基因及其决定的性状遗传类似病毒的转导。
第二节 母性影响
一 . 锥实螺外壳的旋转方向
右旋
左旋
右旋
左旋
旋向取决于受精卵最初 几次卵裂纺锤体的方向
↓ 分析叶绿体基因的分离与重组建立遗传图
一个基因经历细胞质分离,另一紧密连锁的基 因也会随着经历细胞质分离的现象,叫做共分离。
共分离频率与基因间的距离成反比
在衣藻的共分离作图中,有四个叶绿体基因m1、m2、m3、 m4。在每对基因都呈杂合状态的胞质杂合体中,基因共分离 的频率如下:
m1
m2
m3
m1 -
基因对 apt bar apt cob bar cob
第一抗性基因丢失
87 aptS barR 27 aptScobR 48 barS cobR
第二抗性基因丢失
120 aptR barS 最高 207/500 18 aptR cobS 最低 45/500 69 barR cobS 中等 117/500
一. 叶绿体的遗传表现
1. 紫茉莉的花斑叶色遗传 2. 玉米叶片的埃型条纹遗传 3. 衣藻的核外遗传 4. 衣藻叶绿体基因作图
1. 紫茉莉花斑的遗传
亲本枝条♀卵细胞×Fra bibliotek花粉 后代表型
白色
任一
白色
绿色
任一
绿色
花斑
任一 花斑,绿和白
?
2. 玉米叶片的埃型条纹遗传
表现细胞核遗传
表现细胞质遗传
核隐性突变基因ij(7染色体上) ↓
短时间 长时间
KK+卡巴粒(放毒型)×kk无卡巴粒(敏感型)
↓
两种情况
短时间接合
长时间接合
Kk+卡 放毒
↓ KK+卡 稳定放毒
Kk无卡 敏感
↓ KK无卡
敏感
kk+卡 不稳定放毒