第三十七章 转录后加工
合集下载
RNA转录与转录后加工

详细描述
在大多数真核生物中,RNA聚合酶在转录终止后,会在3’端加上一段多聚腺苷酸尾巴。这个过程称为加尾。加 尾的主要作用是促进RNA从核内向细胞质转运,并保护RNA免受3’核酸外切酶的降解。此外,多聚腺苷酸尾巴 也是一些RNA结合蛋白的识别位点,参与mRNA的稳定性、定位和翻译调控。
剪接
总结词
剪接是指将转录的RNA中的内含子序列 去除,并将外显子序列连接起来的加工 过程。
详细描述
C-to-U编辑由胞嘧啶脱氨酶催化,将RNA 中的胞嘧啶转变为尿嘧啶,导致RNA序列 发生变化。这种编辑可以影响RNA的翻译 和功能。
其他编辑类型
总结词
除了A-to-I和C-to-U编辑外,还存在其他类型的RNA编辑,如C-to-A编辑、C-to-G编 辑等。
详细描述
这些编辑类型在特定的生物或组织中发生,由不同的酶催化,导致RNA序列发生不同 的变化。这些编辑可以影响RNA的稳定性、翻译和功能。
肽链终止
终止密码子出现时,核糖体 释放合成的多肽链,并回收 mRNA。
蛋白质合成的起始
起始氨基酸的识别
起始密码子(AUG)被识别并结合甲酰蛋氨酸,形成甲酰蛋氨酸-tRNA。
甲酰蛋氨酸-tRNA在核糖体上的定位
甲酰蛋氨酸-tRNA与起始因子结合,定位到核糖体的P位点。
起始复合物的形成
甲酰蛋氨酸-tRNA与mRNA结合,形成起始复合物。
02
翻译水平调控
03
细胞内环境调控
翻译过程中蛋白质的表达水平可 以影响RNA的稳定性。
细胞内的pH值、离子浓度等环境 因素也可以影响RNA的稳定性。
05
RNA的翻译和蛋白质合成
mRNA的翻译
翻译起始
mRNA在核糖体上定位并结 合翻译起始因子,形成起始 复合物。
在大多数真核生物中,RNA聚合酶在转录终止后,会在3’端加上一段多聚腺苷酸尾巴。这个过程称为加尾。加 尾的主要作用是促进RNA从核内向细胞质转运,并保护RNA免受3’核酸外切酶的降解。此外,多聚腺苷酸尾巴 也是一些RNA结合蛋白的识别位点,参与mRNA的稳定性、定位和翻译调控。
剪接
总结词
剪接是指将转录的RNA中的内含子序列 去除,并将外显子序列连接起来的加工 过程。
详细描述
C-to-U编辑由胞嘧啶脱氨酶催化,将RNA 中的胞嘧啶转变为尿嘧啶,导致RNA序列 发生变化。这种编辑可以影响RNA的翻译 和功能。
其他编辑类型
总结词
除了A-to-I和C-to-U编辑外,还存在其他类型的RNA编辑,如C-to-A编辑、C-to-G编 辑等。
详细描述
这些编辑类型在特定的生物或组织中发生,由不同的酶催化,导致RNA序列发生不同 的变化。这些编辑可以影响RNA的稳定性、翻译和功能。
肽链终止
终止密码子出现时,核糖体 释放合成的多肽链,并回收 mRNA。
蛋白质合成的起始
起始氨基酸的识别
起始密码子(AUG)被识别并结合甲酰蛋氨酸,形成甲酰蛋氨酸-tRNA。
甲酰蛋氨酸-tRNA在核糖体上的定位
甲酰蛋氨酸-tRNA与起始因子结合,定位到核糖体的P位点。
起始复合物的形成
甲酰蛋氨酸-tRNA与mRNA结合,形成起始复合物。
02
翻译水平调控
03
细胞内环境调控
翻译过程中蛋白质的表达水平可 以影响RNA的稳定性。
细胞内的pH值、离子浓度等环境 因素也可以影响RNA的稳定性。
05
RNA的翻译和蛋白质合成
mRNA的翻译
翻译起始
mRNA在核糖体上定位并结 合翻译起始因子,形成起始 复合物。
简述rna转录后加工过程

简述rna转录后加工过程摘要:1.RNA转录后加工过程的概述2.RNA转录后加工的主要步骤a.剪接b.剪切c.RNA编辑d.RNA降解3.各步骤的功能和意义4.实例分析5.RNA转录后加工在生物体中的作用6.研究RNA转录后加工的意义和前景正文:在我们生物体内,基因通过转录过程将DNA信息转化为RNA,但这只是RNA生命历程中的第一步。
接下来,RNA要经历一系列复杂的加工过程,才能最终发挥其生物学功能。
这个过程被称为RNA转录后加工。
RNA转录后加工的主要步骤包括剪接、剪切、RNA编辑和RNA降解。
剪接是指将RNA前体分子中的内含子去除,并将外显子连接成成熟的RNA分子。
这一过程通过特定的酶家族,如剪接酶,来实现。
剪切是指在RNA分子的3"端添加poly(A)尾巴,这是几乎所有真核生物RNA的共同特征。
RNA编辑则是指在RNA分子上发生碱基改变,这一过程依赖于特定的编辑酶和相应的底物。
最后,RNA降解是指RNA分子在细胞内的分解过程,这对于调控RNA水平和维持细胞内稳态至关重要。
这些加工过程对于RNA最终的生物学功能具有重要意义。
以剪接为例,它能消除RNA前体中无功能的RNA片段,使成熟的RNA更具特异性和高效性。
同时,RNA编辑能够改变RNA的序列,从而影响其翻译效率和稳定性。
在生物体中,RNA转录后加工涉及多种生物过程,如基因表达调控、病毒复制和免疫反应等。
对RNA转录后加工的研究,有助于我们深入了解生命过程中的基因表达调控机制,为治疗疾病和开发新型药物提供理论依据。
随着生物科学技术的不断发展,对RNA转录后加工的研究将越来越深入。
分子生物学--转录和转录后加工

B 切割位点B和C
CA
B
报告基因
连接报告基因载体 转化E.coli 分离质粒DNA
连接接头
CA
B
2020/7/21
分别转染培养细胞
通过报告基因产物的活性,以检测基因 活化与上游调控区片段的关系
2.接头扫描突变法
• 可以找出存在于基因转录起始位点和5’端边 界之间的所有调控元件
• 方法的基础是构建一系列序列重叠的调控区突 变体,每个突变体内各有一个短序列的核苷酸 序列被打乱,
非必要亚基:Rpb4、7
2020/7/21
RNA聚合酶II最大亚基的异质性
• CTD: carboxyl terminal domain(C末端结构域)
• Amino acid sequence:
– — Tyr — Ser — Pro — Thr — Ser — Pro — Ser —
• Target for kinase • Function: trigger initiation
2020/7/21
主要内容
• 转录
• 概述 • 原核生物RNA聚合酶 • 原核生物转录过程 • 真核生物转录特点 • 真核生物基因的启动子及转录起始复
合物的组装
2020/7/21
主要内容
• 转录后加工
• rRNA前体的加工 • tRNA前体的加工 • mRNA前体的加工 • RNA的自我剪接 • 真核mRNA前体的剪接 • 真核mRNA前体的选择性剪接 • RNA的编辑
转录起始位点
基本启动子 (initiation site, Inr) (basal promotor)
上游元件 (upstream elements)
2020/7/21
转录后加工学习.pptx

催化的 RNA 有一个鸟苷结合位点和一个底物结合位点
G414
鸟苷结合位点
5’ CUCUCU 底物结合位点
GGGAGG
3’ 含 A16 和 U20 UUUACCU
第一次转移:G-OH 进入 G-结合位点,5’ 外显子进入底物结合位点
3’ G414
3’ G414
G-OH
5’ CUCUCU GGGAGG
聚体蛋白。它能水解各种核苷三磷酸,实际上 是一种NTP酶。由于催化了NTP的水解,ρ因 子能促使新生的RNA链从三元转录复合物中解 离出来,从而终止转录。
作用:ρ因子即附着在新生的RNA链上,靠 ATP水解产生的能量,沿着5‘→3’方向朝RNA 聚合酶移动,到达RNA的3‘-OH端后取代了暂 停在终止位点上的RNA聚合酶,并从模板和酶 上释放RNA,完成转录过程。终止过程需要消 耗能量,所以,ρ因子具有终止转录和核苷三 磷酸酶两种功能。
3’G-OH
5’ pCCCCC-OH3’ GGGAGG 5’
帽 子 的 类 型
第6页/共78页
(4)加帽生化过程
Ⅰ 剪切前加帽 如呼肠病毒, 牛豆病毒
Ⅱ 剪切后加帽 如疱疹病毒和口炎病毒
第7页/共78页
G N1 N2 N3 PPP P P P P…
磷酸水解酶
G N1 N2 N3 PP P P P P… + Pi
G N1 N2 N3
G
PP P
PP
P… + PPP
• (1) 有回文结构存在; • (2)茎的区域富内含G-C; • (3) 强终止子3′端上有6个U;
第24页/共78页
•
N
•
N
N
•
•
N
N
•
生物化学转录后加工

内含子的特点: 位置在反密码子环的下游 外显子和内含子交界处没有明显的保守序列
真核细胞tRNA加工过程
真核生物RNA的转录后加工
❖真核生物rRNA前体的加工 ❖真核生物tRNA前体的转录后加工 ❖真核生物mRNA前体的加工
Figure 6-21 Molecular Biology of the Cell (© Garland Science 2008)
原核生物rRNA前体的加工过程
初转录产物内部碱基互补配对, 折叠成一些茎环结构
continue
茎环结构有助于一些蛋白质结合形成核糖核 蛋白复合体(RNP, ribonucleoprotein)
(甲基化)
(M16)
(RNaseIII和RNaseE酶的剪切)
(M23) (M5)
原核生物RNA的转录后加工
分或全部poly(A),此时mRNA的寿命亦接近终点。
加尾反应所需的特定序列元件
特定序列元件:1)5‘-AAUAAA-3’’ 聚腺苷酸信号序列; 2)在其后11~20nt处紧随着一个“ 5‘-YA-3’ ”结构; 3)在其下游方向的GUGUGUG 。这些序列共同决定了聚 腺苷酸化位点(polyadenylation site)。
真核生物mRNA poly (A)长度并非固定不变 细胞核中的poly (A)长度平均为210±20 nt,细胞质poly (A )长度
190±20 nt。 输送到细胞质中的mRNA其poly (A)可由RNase切短,但又能经细胞质
poly(A)酶重新加长,保持有限的长度。 细胞质中mRNA的poly (A)长度总的趋势是逐渐变短,直到丢失大部
真核生物rRNA的特点
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、 28S rRNA和5S rRNA。其中前三者的基因组成一个转录单位, 由RNA pol Ⅰ合成,产生47S的前体,并很快转变成45S前 体;真核生物的5S rRNA由RNA pol Ⅲ合成121nt的转录产 物,几乎不需要加工。
真核细胞tRNA加工过程
真核生物RNA的转录后加工
❖真核生物rRNA前体的加工 ❖真核生物tRNA前体的转录后加工 ❖真核生物mRNA前体的加工
Figure 6-21 Molecular Biology of the Cell (© Garland Science 2008)
原核生物rRNA前体的加工过程
初转录产物内部碱基互补配对, 折叠成一些茎环结构
continue
茎环结构有助于一些蛋白质结合形成核糖核 蛋白复合体(RNP, ribonucleoprotein)
(甲基化)
(M16)
(RNaseIII和RNaseE酶的剪切)
(M23) (M5)
原核生物RNA的转录后加工
分或全部poly(A),此时mRNA的寿命亦接近终点。
加尾反应所需的特定序列元件
特定序列元件:1)5‘-AAUAAA-3’’ 聚腺苷酸信号序列; 2)在其后11~20nt处紧随着一个“ 5‘-YA-3’ ”结构; 3)在其下游方向的GUGUGUG 。这些序列共同决定了聚 腺苷酸化位点(polyadenylation site)。
真核生物mRNA poly (A)长度并非固定不变 细胞核中的poly (A)长度平均为210±20 nt,细胞质poly (A )长度
190±20 nt。 输送到细胞质中的mRNA其poly (A)可由RNase切短,但又能经细胞质
poly(A)酶重新加长,保持有限的长度。 细胞质中mRNA的poly (A)长度总的趋势是逐渐变短,直到丢失大部
真核生物rRNA的特点
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、 28S rRNA和5S rRNA。其中前三者的基因组成一个转录单位, 由RNA pol Ⅰ合成,产生47S的前体,并很快转变成45S前 体;真核生物的5S rRNA由RNA pol Ⅲ合成121nt的转录产 物,几乎不需要加工。
分子生物学-转录后加工

小鼠免疫球蛋白μ重链基因可变剪接
14-14
可变剪接的多种模式
• 转录物可以按不同的模式进行可变剪接,产生具有多样性的转录本, 许多基因有2种以上的剪接方式,有的甚至达上千种。
• 几种常见的选择性剪接模式: • 1)不同启动子;2)忽略外显子;3)5’选择性剪接;4)3’选择性 剪接;5)保留内含子;6)多腺苷酸化
• 如果装配在snRNP的剪接因 子识别外显子,则称为外显子 界定(exon definition)
• 如果装配在snRNP的剪接因 子识别内含子,则称为内含子 界定(intron definition)
• 通过外显子-内含子边界(剪 接位点)突变,可以分析外显 子-内含子的界定类型
14-12
RNA Pol II的CTD参与外显子界定
内含子类型
特征
剪接体内含子(spliceosomal introns)
细胞核,mRNA,由剪接体催化切除
tRNA内含子(tRNA introns)
细胞核或古菌tRNA基因,由蛋白催化切除
自切割I类内含子(self-splicing group I introns) 细胞器,由RNA催化切除
自切割II类内含子(self-splicing group II introns)细胞器,由RNA催化切除
• 如果在6个位点发生2个不同事件,将会产生26 = 64种结果
14-15
可变剪接示例:果蝇性别决定
• 果蝇的性别决定涉及sxl(sex lethal,性别致死)、 tra (transformer, 转换)和dsx(doublesex, 双性别)三个基因前体mRNA的可变剪接
• 这3个基因的剪接存在级联反应:sxl基因雌性特异性剪接可产生活性 蛋白,进一步增强sxl基因的雌性特异性剪接,同时引发tra基因的雌性 特异性剪接,再进一步引发dsx的雌性特异性剪接
转录后的加工过程及RNA复制

➢ 外显子 (exon) : 基因组中出现在成熟mRNA分子中的序列。
➢ 内含子 (intron) : 位于外显子之间,出现在mRNA前体分子中, 剪接时被去除,不出现在成熟mRNA分子中。
E1
②
U6 U4
UG U5
UACUACA - AG
U1
U2
E2
U1、U4、U5 E1
③
U6 UG
UACUACA - AG
RNA复制酶 (RNA replicase)
RNA指导的RNA聚合酶 (RNA directed RNA polymerase, RDRP)
复制和转录的区别
模板 原料 酶 产物
配对
复制
转录Biblioteka 两股链均复制 模板链转录(不对称转录)
dNTP DNA聚合酶 子代双链DNA (半保留复制) A-T,G-C
mRNA前体经转酯反应切除内含子
剪接体参与mRNA前体的剪接(spliceosome)
剪接体: UsnRNP与mRNA前体结合形成的复合物
UsnRNP
snRNA
Uridine-rich small nuclear
ribonucleoprotein
核蛋白
分类:U1,U2,U4,U5,U6
可变剪接(alternate splicing of mRNA)
1. 各种RNA前体的加工主要有哪些类型? 2. 真核生物mRNA前体剪接部位的结构有 何特点? 3. 真核生物mRNA前体剪接部位的结构有何 特点?mRNA前体的内含子如何被切除的? 4. 为什么转录生成错误RNA远没有复制产 生错误DNA对细胞的影响大?
➢ 剪接和剪切 ➢ 3’-末端添加-CCA ➢ 碱基修饰
真核生物转录后的加工
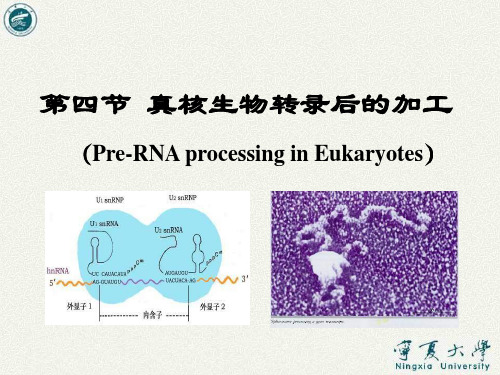
其5´端有一保守序列:3´CAUUCAU-5´。 这一序列可 与内含子5´端的边界序列互补 结合。
U2与分枝点配对
U6 snRNA既能与U4配对 也能与U2配对
U6与mRNA 5´端配对
3、剪接机制(简化)
第一次转酯--左外显子、 内含子剪切套索
第二次转酯--外显子连接、 套索状内含子释放
2、 帽子结构的生物学功能
① 使mRNA免受核酸酶的降解,增加mRNA的稳定性;
② 有助于mRNA越过核膜,进入细胞质;
③被蛋白质起始因子识别,使 mRNA 能与核糖体小亚基
结合并开始合成蛋白质。
(二)3´-端加上多聚腺苷酸尾巴(polyA)
几乎所有真核生物成熟的 mRNA末端都有一串约250个 腺嘌呤核苷酸尾巴。它们并 非模板 DNA 编码,而是在转 录完成时由 poly (A) 多聚酶 合成。加尾位置不在转录物 的最末端,而是在接近末端 的内部位点。在切除 mRNA 3' 末端的一段序列后,再加 上多聚腺苷酸。 加尾是在核内完成 ,先于 mRNA 中 段 的 剪 接 , 和 转 录终止同时进行。
② 协助mRNA从细胞核向细胞质转运。
③作为核糖体的识别信号,使 mRNA分子有效翻译。 ④ 对基因的表达调控有重要作用。
(三)mRNA的内部甲基化
真核生物mRNA分子中有许多甲基化的碱基; 具体的甲基化位点还不太清楚; 主要是:N6-甲基腺嘌呤(m6A); 推测可能为mRNA的剪切提供信号。
转录后加工的主要方式:
加帽(Capping) 加尾(Tailing) 剪接(Splicing ) 碱基修饰(Bases modification) 编辑(Editing )
Pre-RNA
转录后加工及RNA编辑

线粒体和叶绿体rRNA基因的排列方式和转录后加工过
程一般与原核生物的rRNA基因类似。
Processing scheme of 45S human (HeLa) rRNA precursor
真核生物rRNA前体的 甲基化、假尿苷酸化 和切割都是由核仁小 RNA(snoRNA)指导 的。含有C/D框 snoRNA指导2’-O-P 甲基化;H/ACA框 snoRNA指导假尿苷酸 化
OH OH NH
CH2
H2 N
R
二、原核生物中RNA的转录后加工
1、原核生物rRNA前体的加工
16S
23S
tRNA
5S
tRNA
RNaseIII识别部位是特定的RNA 双螺旋区, RNaseIII在16S rRNA
和23S rRNA茎部有两个切割位点
只相差2bp,切割产生P16和P23。
同样的5S rRNA的前体P5由
5’-AAUAAA-3’聚腺苷酸化信号序列、其后11-20nt紧随5’YA-3’结构(Y-嘧啶)、下游有GU rich 序列,这些序列共 同决定了聚腺苷酸化位点 真核生物mRNA的3’端常有20~200个腺苷酸残基 hnRNA的多聚腺苷酸平均长度为150~200nt。
AAUAAA是切割产生3’端和多聚腺苷酸化所必需的。
hnRNA与特定蛋白结合形成hnRNP。 这些蛋白被分为A—U类,其中A、B、C最丰富 hnRNP蛋白:有助于保持hnRNA的单链状态,辅
助各种RNA加工反应
• pre-mRNAs are not exported until processing is complete, hnRNPs are found only in the nucleus.
分子生物学--转录与转录后加工课件

• 序列名称 位置 序列 反式作用因子
• TATA-box -25 ~ –30 TATAAAA
TBP
作用是确定精确的转录起始位点,而不起调节转录 效率的功能
2020/6/3
上游元件 (upstream elements)
常表现出细胞类型特异性,功能主要是提高转 录的效率和特异性
• 序列名称 位置 序列 反式作用因子
转录起始位点
基本启动子 (initiation site, Inr) (basal promotor)
上游元件 (upstream elements)
2020/6/3
启动子近侧序列元件
下游元件 (downstream elements) 上游元件
下游元件
基本启动子 (basal promotor)
A
活化因子结合
β rpo 150 1 结合底物,催化磷酸二脂键形成 B
β rpo 160 1 ’C
结合DNA模板
σ rpo 32-90 1 识别启动子,促进转录起始 D
ω
9
2020/6/3
未知
2020/6/3
2020/6/3
σ因子的结构
σ70因子的一级结构
2020/6/3
2020/6/3
σ因子与启动子的特异性相互作用
2020/6/3
σ因子的更替可控制转录起始
2020/6/3
2020/6/3
转录的起始
2020/6/3
转录的延伸
• RNA链的合成方向5′→3′
2020/6/3
σ循环:RNA聚合酶与启动子结合,使DNA局部熔 解,当转录延伸时, σ因子与核心酶分离,与其它 新的核心酶结合,起始新的转录事件.
RNA的转录后加工分子生物学
• 把这2个位点集结到 一起;
• 催化或协助催化剪接 和连接反应。
U2AF (U2 auxiliary factor), recognizes the polypyrimidine (Py) tract/3' splice important in splicing
• What rules ensure that recognition of splice sites is restricted so that only the 5’and 3’ sites of the same intron are spliced?
外显子遗漏
The first guard mechanism
• 5 种核内小RNA( snRNA) :U1,U2,U4,U5,U6 (序列中富含U) • 与蛋白质形成 RNA-protein 复合物称为小的核内核糖核蛋白 (snRNP)。 • AG前一位核苷酸可以影响剪切效率:CAG=UAG>AAG>GAG
The snRNPs 在剪接中 有3个功能:
• 识别5' 剪接位点和分 支点;
Alternative Splicing Is Regulated by Activators and Repressors
• ESE: exonic splicing enhancer • ISE: intronic splicing enhancer • ESS: exonic splicing silencer • ISS: intronic splicing silencer
正常情况
外显子遗漏
外显子延伸
内含子保留
可变剪接
肌钙蛋白T
两种抗原的比例因剪接相关蛋白SF2/ASF的表达水平而不同。SF2/ASF is an SR protein, when abundant, this protein directs the machinery to favor use of the closest 5' splice site.
• 催化或协助催化剪接 和连接反应。
U2AF (U2 auxiliary factor), recognizes the polypyrimidine (Py) tract/3' splice important in splicing
• What rules ensure that recognition of splice sites is restricted so that only the 5’and 3’ sites of the same intron are spliced?
外显子遗漏
The first guard mechanism
• 5 种核内小RNA( snRNA) :U1,U2,U4,U5,U6 (序列中富含U) • 与蛋白质形成 RNA-protein 复合物称为小的核内核糖核蛋白 (snRNP)。 • AG前一位核苷酸可以影响剪切效率:CAG=UAG>AAG>GAG
The snRNPs 在剪接中 有3个功能:
• 识别5' 剪接位点和分 支点;
Alternative Splicing Is Regulated by Activators and Repressors
• ESE: exonic splicing enhancer • ISE: intronic splicing enhancer • ESS: exonic splicing silencer • ISS: intronic splicing silencer
正常情况
外显子遗漏
外显子延伸
内含子保留
可变剪接
肌钙蛋白T
两种抗原的比例因剪接相关蛋白SF2/ASF的表达水平而不同。SF2/ASF is an SR protein, when abundant, this protein directs the machinery to favor use of the closest 5' splice site.
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
锥体虫COXIII mRNA前体在编 辑过程中插入大量的尿苷酸
gRNA介导的编辑过程
小肠上皮细胞内发生的Apo B蛋白的Pre-mRNA的编辑
编辑的意义
利什曼原虫细胞色素b mRNA通过编辑创造终止密码子
rRNA前体的后加工
*
*
细菌-剪切、修剪和修饰 真核生物 1)剪切、修剪和修饰 (需要snoRNA) 2)剪接 (某些真核生物,如四膜虫)
细菌rRNA前体的后加工
真核细胞核rRNA前体的后加工
由snoRNA 指导的rRNA前体的修饰
snoRNA指导的2′-O甲基化核糖和假尿苷形成
四膜虫rRNA前体的自我剪接
tRNA前体的后加工
*
*
细菌 1) 剪切和修剪 2) 修饰 3) 添加CCA (如何需要) 真核生物 1) 剪切和修剪 2) 修饰 3) 添加CCA 4) 剪接(某些真核生物)
加尾反应由两步组成:
1. 剪切——在3′-UTR一个特定序列上游10-30 核 苷酸序列的位置切开 2. 添加腺苷酸(100-200个)产生多聚A尾巴
加尾信号
真核细胞核mRNA前体的加尾反应
为什么必须有尾巴?
保护mRNA免受3′-外切核酸酶的消化,提高 mRNA的稳定性。 PABP能够与帽子相互作用增强mRNA的可翻 译性,提高其翻译的效率; 影响最后一个内含子的剪接; 某些本来缺乏终止密码子的mRNA通过加尾 反应创造终止密码子。在UG后加尾可产生 UGA,在UA后加尾产生UAA; 通过可变加尾调节基因的表达。
第三十七章 转录后加工
提纲
一.
细菌的转录后加工
mRNA前体的后加工 rRNA前体的后加工 tRNA前体的后加工
1. 2. 3.
二.
真核生物的转录后加工
mRNA前体的后加工 rRNA前体的后加工 tRNA前体的后加工
1. 2. 3.
三.
古菌的转录后加工
转录后加工
基因转录的直接产物被称为初级转录物。初 级转录物一般是无功能的,它们在细胞内必 → 须经历一些结构和化学的变化即所谓的转录 后加工以后才会有功能。转录后加工可能是 RNA的功能所必需的,也可能提供基因表达 调控的一种手段。 RNA所能经历的后加工方式可达10种以上, 但后加工反应的本质要么是增减一些核苷酸 序列,要么是修饰某些特定的核苷酸。
剪接体的组装与剪接反应(续)
*
*
*
*
U6一方面代替U1-snRNP与5'-剪接点的一致序列结合, 另一方面与U4脱离,转而与U2结合; U6的“脚踩两只船”将分支点上突出的腺苷酸上的 2'-OH拉近到5'-剪接点,为第一次转酯反应创造了条 件;在进行第一次转酯反应以后,紧接着U5-snRNP 经历了依赖于ATP的重排,将相邻的外显子并置在一 起,为第二次转酯反应创造了条件; 第一次转酯反应中游离出来的外显子上的3'-OH 亲核 进攻5'-剪接点上的磷酸二酯键,导致内含子以套索结 构释放出来并很快被水解,而外显子则被连接起来; 一旦剪接反应完成,剪接体的所有成分即解体,U6snRNP 与U4-snRNP重新结合参与下一轮剪接反应。
加帽和甲基化
* * * * 帽子结构 加帽反应:共转录 为什么只有mRNA才会加帽? 功能
为什么要有帽子?
提高mRNA的稳定性 参与识别起始密码子的过程,提高mRNA 的可翻译性 有助于mRNA通过核孔从细胞核运输到细 胞质 提高剪接反应的效率。
为什么只有mRNA加帽?
* 之所以只有mRNA和某些snRNA才有帽子 结构,是因为它们都由聚合酶II催化,当 TFIIH磷酸化CTD重复序列中的Ser5以后, 它即可以将转录因子DSIF 招募到转录复合 物。DSIF随后将另一种转录因子NELF招 募进来,以阻滞转录。上述暂停允许加帽 酶进入,来修饰转录物的5'-端。第三种转 录因子P-TEFb是一种激酶,在帽子结构形 成不久也被招募到复合物,然后磷酸化 CTD的Ser2和NELF,NELF随之失活,聚 合酶II继续延伸。
细菌mRNA前体的后加工
在细菌,mRNA很少有后加工。但某些 噬菌体mRNA会发生最简单的剪切反应, 将一个多顺反子切割成单顺反子,也有 某些噬菌体的mRNA需要经过相对复杂 的剪接反应才能成熟(如T4噬菌体编码 的胸苷酸合酶)。
真核细胞mRNA前体的后加工
* 加工形式 1) 5′-端 = 加帽 2) 3′-端 = 加尾 3) 内部 = 剪接 4) 内部=甲基化 5) 编码区=编辑 * 后加工机制
为什么只有mRNA才会加尾?
* 与加帽反应一样,只有mRNA才会加尾, 也是因为聚合酶 II最大亚基上的 CTD 重复 序列被 TFIIH 磷酸化,但是磷酸化位点为 Ser2 。 Ser2 的 磷 酸 化 将 加 尾 因 子 招 募 到 mRNA 前体上进行加尾反应。
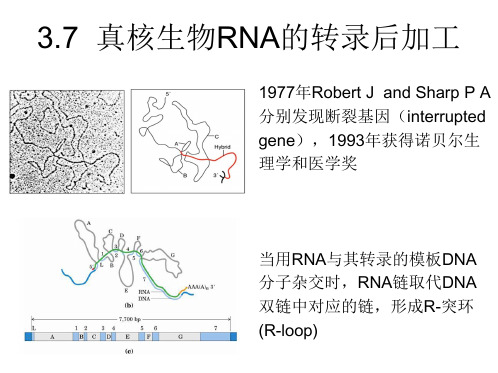
mRNA前体的剪接
剪接这种后加工方式是在发现基因断裂的现象后确定 的。1977年,由Phillip Sharp和Richard Roberts领导两个 实验小组几乎同时在腺病毒的晚期表达基因中发现蛋 白质基因断裂现象。 进一步研究表明,基因断裂是真核细胞及其病毒的基 因组中的普遍现象,在高等生物的基因组中,只有很 少的蛋白质基因是连续的(如组蛋白和干扰素),但 在低等的真核生物,断裂基因却不多见。 不同断裂基因含有的内含子数目不一定相同,同样内 含子大小也会有差别。一般说来,一个典型的真核生 物蛋白质的基因由10% 的外显子序列和90%的内含子 序列组成。
酵母Pre-tRNA的剪接
古菌的转录后加工
古菌的转录后加工在某些方面分别与细菌和真核生物一样,在 某些方面仅类似于细菌或真核生物,还有少数是古菌特有的。 tRNA后加工包括:5′-端和3′-端的剪切和修剪 ;核苷酸的修饰 ; 添加CCA ;含有内含子的还包括剪接,但古菌的tRNA剪接与 真核生物一样,由一系列专门的蛋白质组成的酶按照一定的次 序催化完成的 rRNA后加工:其基因以多顺反子的形式存在,通过剪切和修剪 释放出三种rRNA。核苷酸的修饰与真核生物一样,需要各种特 殊的小RNA(相当于真核生物的snoRNA)与蛋白质形成的 RNP去锁定修饰目标。但对于含有内含子的rRNA的剪接与其 tRNA剪接机制相似,完全不同于真核生物 mRNA后加工:后加工类似于细菌,没有帽子结构,一般也没 有多聚A尾巴。对于少数具有多聚A尾巴的mRNA而言,其加尾 的机制和功能完全不同于真核生物,倒与含有多聚A尾巴的细 菌相似。含有内含子的mRNA的剪接机制既不同于真核生物, 也不同于细菌,而是类似于它的tRNA的剪接
U5
上游外显子 和下游外显子
ห้องสมุดไป่ตู้U6
U4 (和U2)
在剪接体中与U2结合。
snRNP的重要性
小核糖核酸蛋白颗粒(发音为 snurps)—— 参与剪接 一种 snRNP 由一个snRNA (100-200核苷酸长) 和10种不同的蛋白质组成 snRNPs和mRNA前体形成剪接体 剪接体大小与核糖体差不多,其组装需要 ATP
剪接信号
剪接通过2次转酯反应
在转酯反应中,1个磷酸二酯键被转移到另1个羟基上 没有水解,无能量的损失
参与剪接反应的5种snRNA
snRNA U1 U2 U4 互补性 内含子的5' -端 分支点 U6 snRNA 功能 识别和结合5'-剪接点 识别和结合分支点。在剪接体组装中,也与 U6 snRNA 配对。 结合并失活U6。在剪接体组装中,U4-U6之间 的碱基配对被U2-U6之间的剪接配对所取代。 U6也取代U1 与5' -剪接点的相互作用。 与相邻的2个外显子结合,以防止它们离开剪 接体。
大肠杆菌 tRNATyr的后加工
核糖核酸酶 P –一种真正的核酶
一种核酸内切酶–参与tRNA前体5′-端的后加 工 大肠杆菌核糖核酸酶 P: 14-k的多肽链 + 一 种 377核苷酸长的RNA ( M1 RNA) 在高浓度的Mg2+下,分离的M1 RNA能单独 和剪切大肠杆菌的tRNA前体。核糖核酸酶P 中的多肽链提高 M1 RNA的剪切速率,允 许它在生理Mg2+浓度下起作用。
R-环技术:真核基因内含子数目与结构分析
变性
与成熟的mRNA杂交; 在电镜下观察
DNA 模板链 成熟的mRNA
R环实验的结果及其对结果的解释
鸡卵清蛋白基因结构及其Pre-mRNA的后加工
mRNA前体的剪接机制
*
*
*
* *
mRNA前体的剪接是高度精确的。其精确 性一方面取决于位于外显子和内含子交界 处的剪接信号(可以将其视为内因),另 外一方面取决于5种被称为snRNP的核糖核 酸蛋白质复合物(可以将其视为外因)。 剪接反应的“内因”——剪接信号 剪接反应的“外因”——snRNP和剪接因子 剪接反应——两次转酯反应 剪接体的组装
RNA polII CTD的磷酸化与加尾和加帽反应的关系
3′-加尾
加尾内因:加尾信号 ATG 加尾外因:
终止密码子
mRNA
1. CPSF = “剪切/多聚腺苷酸化特异性因子(3个 亚基)识别mRNA前体 3′-UTR上的 AAUAAA 一致序列,并与此结合 2. CFI和CFII = “剪切因子” 3. PAP = “poly(A) 聚合酶”
为什么只有mRNA被snRNPs剪接?
* * *
RNA聚合酶的CTD是剪接必需的 剪接体内的剪接因子与CTD作用 这保证来自RNA聚合酶II的转录物处于 和剪接机构正确的相互作用的位置。
可变剪接和反式剪接
可变剪接
1. 2. 剪接是精确的,但是... 一种mRNA前体可以通过去除不同的内含子/ 外显子组合而剪接成两种以上的mRNA,从 而导致一个基因可以编码多种蛋白质 可变剪接受到调节 果蝇的性别决定与可变剪接有关 发生在两个mRNA分子之间的剪接 比较罕见 自然界只发现在锥体虫和秀丽线虫