细胞生物学翟中和第四版蛋白质分选李绍军
细胞生物学-王版第8章蛋白质分选与膜泡运输—2016
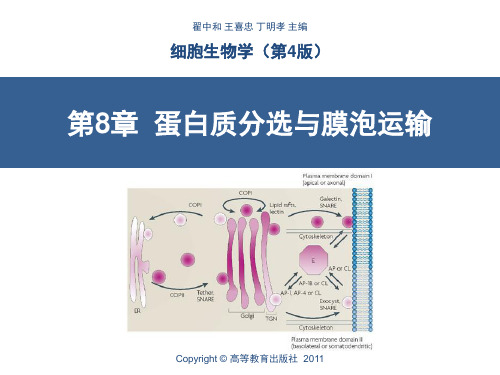
第一节 细胞内蛋白质的分选
01
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)© 高等教育出版社 2011
一、信号假说与蛋白质分选信号
1999 年诺贝尔生理 学或医学奖
发现蛋白质由内部信号决定 其在细胞内的转移和定位
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)© 高等教育出版社 2011
(3)分泌性蛋白的合成与其共翻译转运
02
分泌性蛋白的合成与其跨越内质网膜的共翻译转运图解 图示信号肽、SRP、DP 及移位子之间的相互作用
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)© 高等教育出版社 2011
(4)内质网膜整合蛋白3
Ⅰ型: 如LDL 受体、流感HA、胰岛素受体、 生长素受体;Ⅱ型:无唾液酸糖蛋白受 体、转铁蛋 白受体、高尔基半乳糖苷转移酶、高尔基唾液酸转 移酶;Ⅲ型:细胞色 素P450 ;Ⅳ型:G 蛋白偶联受 体、葡萄糖转运蛋白、电压门Ca2+ 通道、ABC 小分 子泵、CFTR(Cl-)通道、Sec61 蛋白
• 开始转移序列(start
transfer sequence)
• 内在停止转移锚定序 列(internal stop-
transfer anchor sequence, STA)
• 内在信号锚定序列
(internal signal anchor sequence, SA)
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)© 高等教育出版社 2011
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)© 高等教育出版社 2011
1. 信号假说
信号肽(signal peptide) 信号识别颗粒(signal recognition particle,SRP) 信号识别颗粒的受体(又称停泊蛋白,docking protein,DP)
(2021年整理)细胞生物学翟中和第四版教案

细胞生物学翟中和第四版教案编辑整理:尊敬的读者朋友们:这里是精品文档编辑中心,本文档内容是由我和我的同事精心编辑整理后发布的,发布之前我们对文中内容进行仔细校对,但是难免会有疏漏的地方,但是任然希望(细胞生物学翟中和第四版教案)的内容能够给您的工作和学习带来便利。
同时也真诚的希望收到您的建议和反馈,这将是我们进步的源泉,前进的动力。
本文可编辑可修改,如果觉得对您有帮助请收藏以便随时查阅,最后祝您生活愉快业绩进步,以下为细胞生物学翟中和第四版教案的全部内容。
第一章绪论一.细胞生物学研究的内容和现状1.细胞生物学是现代生命科学的重要基础学科细胞生物学是研究细胞基本生命活动规律的科学,它是在不同层次(显微、亚显微与分子水平)上以研究细胞结构与功能、细胞增殖、分化、衰老与凋亡、细胞信号传递、真核细胞基因表达与调控、细胞起源与进化等为主要内容。
核心问题是将遗传与发育在细胞水平上结合起来。
细胞生物学的主要研究内容一般可分为细胞结构功能与细胞重要生命活动两大基本部分:大致归纳为下面几个领域:1)细胞核、染色体以及基因表达的研究2)生物膜与细胞器的研究3)细胞骨架体系的研究4)细胞增殖及其调控5)细胞分化及其调控6)细胞的衰老与凋亡7)细胞的起源与进化8)细胞工程当前细胞生物学研究的总趋势与重点领域1)细胞生物学与分子生物学(包括分子遗传学与生物化学)相互渗透与交融是总的发展趋势2)当前研究的重点领域:I:染色体DNA与蛋白质相互作用关系——主要是非组蛋白对基因组的作用II:细胞增殖、分化、凋亡的相互关系及其调控III:细胞信号转导的研究IV:细胞结构体系的组装二.细胞学与细胞生物学发展简史1.细胞的发现2.细胞学说的建立其意义1838~1839年,德国植物学家施莱登和动物学家施旺提出了“细胞学说”。
3.细胞学的经典时期4.实验细胞学时期5.细胞生物学学科的形成与发展第二章细胞基本知识概要细胞的基本概念1.细胞是生命活动的基本单位.1)一切有机体都由细胞构成,细胞是构成有机体的基本单位2)细胞具有独立的、有序的自控代谢体系,细胞是代谢与功能的基本单位3)细胞是有机体生长与发育的基础4)细胞是遗传的基本单位,细胞具有遗传的全能性5)没有细胞就没有完整的生命2.细胞概念的一些新思考细胞是多层次非线性的复杂结构体系:细胞具有高度复杂性和组织性2)细胞是物质(结构)、能量与信息过程精巧结合的综合体细胞是高度有序的,具有自组装能力与自组织体系。
翟中和第四版细胞生物学 章习题及答案(doc)

翟中和第四版《细胞生物学》习题集及答案第一章绪论一、名词解释细胞生物学:是研究和揭示细胞基本生命活动规律的科学,它从显微、亚显微与分子水平上研究细胞结构与功能、细胞增殖、分化、代谢、运动、衰老、死亡,以及细胞信号传导,细胞基因表达与调控,细胞起源与进化等重大生命过程。
二、填空题1、细胞分裂有直接分裂、减数分裂和有丝分裂三种类型。
2、细胞学说、能量转化与守恒和达尔文进化论并列为19世纪自然科学的“三大发现”。
3、细胞学说、进化论和遗传学为现代生物学的三大基石。
4、细胞生物学是从细胞的显微、亚显微和分子三个水平,对细胞的各种生命活动展开研究的科学。
5、第一次观察到活细胞有机体的人是荷兰学者列文虎克。
三、问答题:1、当前细胞生物学研究中的3大基本问题是什么?答:①基因组是如何在时间与空间上有序表达的?②基因表达产物是如何逐级组装成能行使生命活动的基本结构体系及各种细胞器的?这种自组装过程的调控程序与调控机制是什么?③基因及其表达的产物,特别是各种信号分子与活性因子是如何调节诸如细胞的增殖、分化、衰老与凋亡等细胞最重要的生命活动过程?2、细胞生物学的主要研究内容有哪些?答:①生物膜与细胞器②细胞信号转导③细胞骨架体系④细胞核、染色体及基因表达⑤细胞增殖及其调控⑥细胞分化及干细胞生物学⑦细胞死亡⑧细胞衰老⑨细胞工程⑩细胞的起源与进化3、细胞学说的基本内容是什么?答:①细胞是有机体,一切动植物都是由细胞发育而来,并由细胞和细胞产物所构成。
②每个细胞作为一个相对独立的单位,既有它自己的生命,又对与其他细胞共同组成的整体的生命有所助益。
③新的细胞可以通过已存在的细胞繁殖产生。
第二章细胞的统一性与多样性一、名词解释1、细胞:生命活动的基本单位。
2、病毒(virus):非细胞形态生命体,最小、最简单的有机体,必须在活细胞体内复制繁殖,彻底寄生性。
3、原核细胞:没有核膜包裹的和结构的细胞,细菌是原核细胞的代表。
4、质粒:细菌的核外DNA。
细胞生物学翟中和编第8章蛋白质分选与膜泡运输
负责从内质网高尔基体的物质运输; COPII包被蛋白由5种蛋白亚基组成:Sar1、 Sec12、 Sec23/Sec24、 Sec13/Sec31、Sec16 包被蛋白的装配是受控的; COPII包被膜泡具有对转运物质的选择性并使之浓缩。 COPII蛋白能识别并结合跨膜内质网蛋白胞质面一端的信号序列(Asp-X-Glu) 内质网腔面的受体能与ER腔中的可溶性蛋白(如分泌蛋白)结合。
02
五、转运膜泡与靶膜的锚定与融合
3
2
4
1
SNAREs
结构:具有一个螺旋结构域,相互缠绕形成跨SNAREs复合体,将小泡与靶膜拉在一起。
功能:介导运输小泡与靶膜的融合。
类型:v-SNAREs和t-SNAREs。
膜泡运输的定向机制
SNAREs in vesicle transport
(二)Rabs
蛋白质要解折叠与重新折叠
导肽转运线粒体蛋白质的特点
(二)叶绿体基质蛋白与类囊体蛋白的靶向输入
外膜蛋白 内膜蛋白 叶绿体基质蛋白 类囊体膜蛋白 类囊体腔蛋白
叶绿体蛋白质的运送及组装
(三)过氧化物酶体蛋白的分选
信号序列不被切除
过氧化物酶体膜受体
可溶性细胞质受体
ATP水解
C端SKL(Ser-Lys-Leu)靶向序列
开始转移序列(start transfer sequence)
内在停止转移锚定序列(internal stop-transfer anchor sequence, STA)
内在信号锚定序列(internal signal anchor sequence, SA)
01
03
02
多肽跨膜次数的确定
(完整版)翟中和版-细胞生物学各章习题及解答完整修订版

《细胞生物学》习题及解答第一章绪论本章要点:本章重点阐述细胞生物学的形成、发展及目前的现状和前景展望。
要求重点掌握细胞生物学研究的主要内容和当前的研究热点或重点研究领域,重点掌握细胞生物学形成与发展过程中的主要重大事件及代表人物,了解细胞生物学发展过程的不同阶段及其特点。
一、名词解释1、细胞生物学cell biology:是研究细胞基本生命活动规律的科学,是在显微、亚显微和分子水平上,以研究细胞结构与功能,细胞增殖、分化、衰老与凋亡,细胞信号传递,真核细胞基因表达与调控,细胞起源与进化等为主要内容的一门学科。
2、显微结构microscopic structure:在普通光学显微镜中能够观察到的细胞结构,直径大于0.2微米,如细胞的大小及外部形态、染色体、线粒体、中心体、细胞核、核仁等,目前用于研究细胞显微结构的工具有普通光学显微镜、暗视野显微镜、相差显微镜、荧光显微镜等。
3、亚显微结构submicroscopic structure:在电子显微镜中能够观察到的细胞分子水平以上的结构,直径小于0.2微米,如内质网膜、核膜、微管、微丝、核糖体等,目前用于亚显微结构研究的工具主要有电子显微镜、偏光显微镜和X线衍射仪等。
4、细胞学cytology:研究细胞形态、结构、功能和生活史的科学,细胞学的确立是从施莱登(1838)和施旺(1839)的细胞学说的提出开始的,而大部分细胞学的基础知识是在十九世纪七十年代以后得到的。
在这一时期,显微镜的观察技术有了显著的进步,详细地观察到核和其他细胞结构、有丝分裂、染色体的行为、受精时的核融合等,细胞内的渗透压和细胞膜的透性等生理学方面的知识也有了发展。
对于生殖过程中的细胞以及核的行为的研究,对于发展遗传和进化的理论起了很大作用。
5、分子细胞生物学molecular cell biology:是细胞的分子生物学,是指在分子水平上探索细胞的基本生命活动规律,主要应用物理的、化学的方法、技术,分析研究细胞各种结构中核酸和蛋白质等大分子的构造、组成的复杂结构、这些结构之间分子的相互作用及遗传性状的表现的控制等。
细胞生物学翟中和第四版教案

高效、精确、灵活,可实现多种基因编辑需求。
3
应用领域
基因治疗、基因功能研究、农作物遗传改良等。
单细胞测序技术在肿瘤研究中的应用
单细胞测序技术原理
对单个细胞进行基因组、转录组或蛋 白质组测序,揭示单细胞水平上的基
因表达和变异情况。
发现肿瘤干细胞
通过分析单细胞数据,可发现具有自 我更新和分化潜能的肿瘤干细胞,为
再生医学原理和应用前景
治疗退行性疾病
如帕金森病、阿尔茨海默病等神经系统疾病
修复损伤组织
如心肌梗塞后的心肌再生、糖尿病患者的胰 岛细胞移植等
个性化医疗
利用患者自身细胞进行再生治疗,避免免疫 排斥反应和伦理问题
现代细胞生物学研究技术与
06
方法
显微镜技术原理及应用领域
显微镜技术原理
利用光学或电子显微 镜对细胞进行高倍率 放大观察,揭示细胞
VS
细胞生物学发展现状
目前,细胞生物学已经成为生命科学领域 最活跃的研究方向之一。随着高通量测序 、蛋白质组学、代谢组学等技术的发展, 细胞生物学研究已经进入系统生物学时代 。同时,细胞生物学与医学、药学、生物 工程等领域的交叉融合也促进了相关领域 的快速发展。
本课程教学目标与要求
教学目标
本课程的目标是使学生掌握细胞生物学的基本概念和原理,了解细胞的结构、功能、代谢、遗传与发 育等方面的知识,培养学生的实验技能和科研能力,为后续的专业课程学习和科研工作打下基础。
细胞周期调控因子和信号通路
细胞周期蛋白依赖性激酶(CDK)和细胞周期蛋白(cy…
通过不同时相的CDK-cyclin复合物的形成和激活,推动细胞周期的进行。
检查点蛋白
监控细胞周期的关键节点,确保DNA复制和染色体分离的准确性。
细胞生物学(翟中和第四版)第8章-第12章

• 转运到线粒体和过氧化物酶体的蛋白质靠的是线粒体蛋白N 端的导肽(leader peptide)或过氧化物酶体蛋白C端的靶 向序列(targeting sequence)。
• 注意:蛋白质最终的定位还需要其它空间定位信号;蛋白质 必须在分子伴侣的帮助下解折叠或维持非折叠状态,以利于 通过膜;蛋白质输入通常需要能量。
(一)蛋白质从细胞质基质输入到 线粒体
1. 线粒体蛋白从细胞质基质输入到线粒体基质:两性的N端靶 向信号序列(形成α螺旋构象)对于指导蛋白质输入线粒 体基质是至关重要的;需要分子伴侣胞质蛋白Hsc70和线 粒体基质蛋白Hsc70协助;需要从内外膜接触点的Tom (外膜移位子)和Tim(内膜移位子)处输入。
核基因编码的蛋白质的分选大体可分2条途径: (1)共翻译转运(cotranslational translocation)途径:
即蛋白质合成在游离核糖体上起始之后,由信号肽和与之结 合的SRP引导转移至糙面内质网,然后新生肽边合成边转 入糙面内质网腔或定位在ER膜上,经转运膜泡运到高尔基 体加工包装再分选至溶酶体、细胞质膜或分泌到细胞外。这 种蛋白在信号肽引导下边翻译边跨膜转运的过程称为共翻译 转运。注意:内质网和高尔基体本身的蛋白质分选也按此途 径来完成。 (2)翻译后转运(post-translational translocation)途径: 即蛋白质在细胞质基质游离核糖体上合成以后,再转移到膜 围绕的细胞器,如细胞核、线粒体、叶绿体和过氧化物酶体, 或者成为细胞质基质的可溶性驻留蛋白和骨架蛋白。酵母中 有些分泌蛋白由结合ATP的分子Bip蛋白(Bip-ATP)与膜整 合蛋白Sec63复合物相互作用,水解ATP提供动力驱动翻译 后转运途径,即分泌蛋白在细胞质基质游离核糖体上合成后, 再转运至内质网中。
新细胞生物学课件翟中和第四版13篇

新细胞生物学课件翟中和第四版1第一篇:细胞的起源和进化细胞是构成生物的基本单位,但细胞是如何起源和发展的呢?关于细胞的起源有两种观点:一种是自然发生说,也称为原细胞说;另一种是造物主说,也称神创说。
自然发生说认为,最初的细胞是在原始地球中,由非生物物质逐渐复杂组合而成的。
支持这一观点的证据有以下几点:首先,现代生命最早的祖先是单细胞生物,它们的构成和功能与原始地球上的自然环境有着密切的关系;其次,多种有机分子可以在实验条件下通过自然反应生成,而这些有机分子就是构成生物体的基本物质;最后,多功能的核酸分子和细胞膜的组装能够在实验室条件中模拟出来。
造物主说则认为,最初的细胞是由一个造物主创造的。
但是,这一观点并不能提供具体的科学证据,因此被认为是一种宗教信仰。
无论哪种观点,都不能否认细胞的进化。
细胞的进化可以通过细胞分裂和基因突变等方式进行。
在分裂过程中,细胞将自身物质平均分配到两个新细胞中,并维持了原始的遗传信息;在基因突变中,细胞的遗传信息会发生变异、重组等,从而产生了新的功能和适应环境的能力。
总的来说,细胞的起源和进化是生物学领域中一个非常重要的问题,它们的研究将有助于我们更好地了解生命的本质和进化的规律。
第二篇:细胞的组成和功能细胞作为生物的基本单位,它的组成和功能非常复杂。
大多数细胞都具有以下三个主要组成部分:细胞质、细胞核和细胞膜。
细胞质是细胞内的非核物质,其主要成分是水、有机分子、无机盐和细胞器等。
细胞器是细胞质中一些具有特定功能的亚细胞结构,如线粒体、内质网、高尔基体、溶酶体等,它们分工合作共同维持细胞的正常代谢和生长。
细胞核是细胞的控制中心,主要包括染色体和核仁两部分。
染色体是细胞内质粒的一种,其中含有生物体遗传信息的DNA,核仁则参与到蛋白质合成的过程中。
细胞膜是细胞内外分隔的关键部分,其主要成分是脂质双层和膜蛋白。
脂质双层是由两层疏水性和亲水性的脂质分子组成,能有效地阻止水溶性物质的自由进出。
细胞生物学(翟中和第四版)

包括显微镜观察、细胞计数、细胞体积测量和生物量测定等方法。
细胞的分裂方式及过程
有丝分裂
真核细胞进行的一种分裂方式, 包括前期分离和细胞的分
裂。
无丝分裂
原核细胞和某些真核细胞进行的 分裂方式,不形成纺锤丝,直接 通过细胞膜的内陷将细胞一分为
细胞株和细胞系
通过选择法或克隆形成法从原代培养物或细胞系中获得具有特殊性质或标志物的培养物称为细胞株;当培养 超过50代时,大多数的细胞不再分裂,出现危机,但是有部分细胞可以度过危机继续传代,这些细胞称为 细胞系。
细胞融合技术及应用
细胞融合方法
包括病毒诱导融合、化 学融合法(如PEG融合 法)和电融合法等。
THANKS
感谢观看
探索生物进化机制
通过研究不同物种细胞的结构 和功能差异,可以探索生物进
化的机制和规律。
02
细胞的基本结构与功能
细胞膜的结构与功能
细胞膜的主要成分
01
脂质、蛋白质和糖类。
细胞膜的结构
02
磷脂双分子层构成基本支架,蛋白质分子以不同方式镶嵌其中
。
细胞膜的功能
03
作为细胞的边界,维持细胞内外环境的相对稳定;控制物质进
杂交瘤技术
将具有分泌特异性抗体 能力的致敏B细胞和具 有无限繁殖能力的骨髓 瘤细胞融合为B细胞杂 交瘤。
单克隆抗体
由单一B细胞克隆产生 的高度均一、仅针对某 一特定抗原表位的抗体 。
干细胞技术及应用
干细胞类型
包括胚胎干细胞、成体干细胞和诱导多能干细胞等。
干细胞应用
可用于研究细胞发育和分化机制、建立疾病模型、进行药物筛选和基因治疗等 。如利用干细胞治疗帕金森病、糖尿病、心肌梗死等疾病。
《细胞生物学》第四版翟中和、王喜忠、丁明孝名词解释
癌基因 oncogene通常表示原癌基因(proto oncogene )的突变体,这些基因编码的蛋白使细胞的生长失去控制,并转变成癌细胞,故称癌基因。
氨酰-tRNA 合成酶 aminoacyl tRNA synthetase将氨基酸和对应的tRNA 的3′端进行共价连接形成氨酰-tRNA 的酶。
不同的氨基酸被不同的氨酰-tRNA 合成酶所识别。
暗反应light independent reaction光合作用中的另外一种反应,又称碳同化反应(carbonassimilation reaction)。
该反应利用光反应生成的ATP 和NADPH 中的能量,固定CO 2生成糖类。
白介素-1β转换酶 interleukin-1β converting enzyme,ICECaspase-1,Caspase 家族成员之一,线虫Ced3在哺乳动物细胞中的同源蛋白,催化白介素-1β前体的剪切成熟过程。
半桥粒 hemidesmosome 位于上皮细胞基底面的一种特化的黏着结构,将细胞黏附到基膜上。
胞间连丝 plasmodesma 相邻植物细胞之间的联系通道,直接穿过两相邻细胞的细胞壁。
胞内体 endosome 动物细胞内由膜包围的细胞器,其作用是转运由胞吞作用新摄取的物质到溶酶体被降解。
胞内体被认为是胞吞物质的主要分选站。
胞吐作用 exocytosis 携带有内容物的膜泡与质膜融合,将内容物释放到胞外的过程。
胞吞作用 endocytosis通过质膜内陷形成膜泡,将细胞外或细胞质膜表面的物质包裹到膜泡内并转运到细胞内(胞饮和吞噬作用)。
胞外基质 extracellular matrix分布于细胞外空间、由细胞分泌的蛋白质和多糖所构成的网络结构,如胶原和蛋白聚糖等,在决定细胞形状和活性的过程中起着一种整合作用。
胞质动力蛋白 cytoplasmic dynein 由多条肽链组成的巨型马达蛋白,利用ATP 水解释放的能量将膜泡或膜性细胞器等沿微管朝负极转运。
2015细胞生物学(翟中和第四版第8章 蛋白质分选(李绍军)

蛋白分选(蛋白质寻靶): 蛋白分子在蛋白内部的分选 信号指导下从细胞质运往各种目标细胞器或细胞表面。
• 细胞内合成的蛋白质、脂类等物质之所以能够定向的转运到特定的细胞器 取决于两个方面: – 其一是蛋白质中包含特殊的信号序列(signal sequence)。 – 其二是细胞器上具特定的信号识别装置(分选受体,sorting receptor)。
进入内质网腔的可溶性蛋白没有停止转移序列
含内在停止转移锚定序列(STA)和内在信号锚定序列(SA)的 蛋白在内质网膜上的拓扑特征
蛋定 膜 白序 整 膜列 合 定) 蛋 位和 白 中开 的 的始 内 作转 部 用移 停 序止 列转 (移 内序 部列 信( 号内 锚部 定停 序止 列转 )移 在锚
葡萄糖转运蛋白1等多跨膜α螺旋蛋白的合成和转运:多个开始转移序列(内部信号锚定序列,不切除的信号 肽)和停止转移序列(内部停止转移锚定序列)形成多次跨膜。
高尔基体在细胞分裂中的解体和分裂后重新组装Golgi assembly occurs in 2 stages upon mitotic exit: (a) Video frames illustrating post-mitotic Golgi assembly in mCherry(一种红色荧光蛋白)-Rab6(定位于高尔基体或后高尔基体区室上的GTP结合蛋白,调节膜 泡运输、融合) expressing RPE1(一种视网膜上皮细胞株) cells. Time zero marks approximate onset of telophase(末期开始). Boxed area is enlarged below. (b) Postmitotic Golgi particle size based on live imaging experiments in NT-control cells. Average fold increase of Golgi particles relative to time zero is shown. (c) Enlarged box from (a) showing Golgi mini-stack (red) clustering (6–9', blue and yellow arrows indicate two separate clusters) prior to re-location toward the centrosome (10'). Miller et al. Golgi-derived microtubules cluster and organize Golgi ministacks (G-stage of Golgi assembly). At the same time, centrosomal microtubules collect Golgi stacks in the cell center (C-stage of Golgi assembly). In CLASP高尔基 体膜上组织微管组装的一种蛋白)depleted cells that have no 细胞分裂末期高尔基体重新组装由两套微管体系完成——中心体微管体系(CGolgi-derived microtubules, G-
翟中和第四版细胞生物学1~9章习题及答案

翟中和第四版细胞生物学1~9章习题及答案翟中和第四版《细胞生物学》习题集及答案第一章绪论一、名词解释细胞生物学:是研究和揭示细胞基本生命活动规律的科学,它从显微、亚显微与分子水平上研究细胞结构与功能、细胞增殖、分化、代谢、运动、衰老、死亡,以及细胞信号传导,细胞基因表达与调控,细胞起源与进化等重大生命过程。
二、填空题1、细胞分裂有直接分裂、减数分裂和有丝分裂三种类型。
2、细胞学说、能量转化与守恒和达尔文进化论并列为19世纪自然科学的“三大发现”。
3、细胞学说、进化论和遗传学为现代生物学的三大基石。
4、细胞生物学是从细胞的显微、亚显微和分子三个水平,对细胞的各种生命活动展开研究的科学。
5、第一次观察到活细胞有机体的人是荷兰学者列文虎克。
三、问答题:1、当前细胞生物学研究中的3大基本问题是什么?答:①基因组是如何在时间与空间上有序表达的?②基因表达产物是如何逐级组装成能行使生命活动的基本结构体系及各种细胞器的?这种自组装过程的调控程序与调控机制是什么?③基因及其表达的产物,特别是各种信号分子与活性因子是如何调节诸如细胞的增殖、分化、衰老与凋亡等细胞最重要的生命活动过程?2、细胞生物学的主要研究内容有哪些?答:①生物膜与细胞器②细胞信号转导③细胞骨架体系④细胞核、染色体及基因表达⑤细胞增殖及其调控⑥细胞分化及干细胞生物学⑦细胞死亡⑧细胞衰老⑨细胞工程⑩细胞的起源与进化3、细胞学说的基本内容是什么?答:①细胞是有机体,一切动植物都是由细胞发育而来,并由细胞和细胞产物所构成。
②每个细胞作为一个相对独立的单位,既有它自己的生命,又对与其他细胞共同组成的整体的生命有所助益。
③新的细胞可以通过已存在的细胞繁殖产生。
第二章细胞的统一性与多样性一、名词解释1、细胞:生命活动的基本单位。
2、病毒(virus):非细胞形态生命体,最小、最简单的有机体,必须在活细胞体内复制繁殖,彻底寄生性。
3、原核细胞:没有核膜包裹的和结构的细胞,细菌是原核细胞的代表。
细胞生物学(翟中和,高教四版)

细胞生物学(翟中和,高教四版)第一章绪论 (3)第二章细胞的统一性和多样性 (5)第三章细胞生物学研究方法 (9)第四章细胞质膜 (11)第五章物质的跨膜运输 (13)第六章线粒体和叶绿体 (15)第七章细胞质基质与内膜系统 (17)第八章蛋白质分选与膜泡运输 (20)第九章细胞信号转导 (21)第十章细胞骨架 (25)第十一章细胞核与染色质 (27)第十二章核糖体 (34)第十三章细胞周期与细胞分裂 (35)第十四章细胞增殖调控与癌细胞 (36)第十五章细胞分化与胚胎发育 (37)第十六章细胞死亡与细胞衰老 (39)第十七章细胞的社会联系 (40)第一章绪论第一节细胞生物学研究的内容与现状一、现代生命科学的一门重要的基础前沿学科当前细胞生物学研究的课题归纳起来包括3个根本性问题:(1)基因组是如何在时间与空间上有序表达的?(2)基因表达产物是如何逐级组装成能行使生命活动的基本结构体系及各种细胞器?(3)基因及其表达的产物,特别是各种信号分子与活性因子,是如何调节诸如细胞的增殖、分化、衰老、与凋亡等细胞最重要的生命活动过程的?二、细胞生物学得主要研究内容10个方面:(一)生物膜与细胞器(二)细胞信号转导基本研究内容3个方面:①细胞间信号传递:信号分子-受体作用②受体与信号跨膜转导:G蛋白与一系列受体③细胞内信号传递途径与网络调控-生物学效应(三)细胞骨架体系(四)细胞核、染色体及基因表达(五)细胞增殖及其调控增殖调控研究从两方面进行:①找控制增殖的因子②研究控制增殖的主要检验点相关的周期蛋白与依赖于周期蛋白的激酶的调控机理(六)细胞分化及干细胞生物学(七)细胞死亡(八)细胞衰老(九)细胞工程(十)细胞的起源于进化目前全球最热门的研究方向是:①细胞周期调控②细胞凋亡③细胞衰老④信号转导⑤DNA的损伤修复第二节细胞学与细胞生物学发展简史生物科学发展的3个阶段:①>19世纪形态描述为主-生物科学②20世纪前半个世纪(1950年前)-实验生物学③20世纪50年代后-现代生物学一、细胞的发现二、细胞学说的建立及其意义当时“细胞学说”的基本内容:①细胞是有机体,一切动植物都是有细胞发育而来,并有细胞产物所构成②每个细胞作为一个相对独立的单位,既有它自己的生命,又对其他细胞共同组成的整体的生命有所助益③新的细胞可以通过已存在的细胞繁殖产生三、细胞学的经典时期(一)原生质理论的提出(二)细胞分裂的研究(三)细胞器的发现四、实验细胞学与细胞学的分支及其发展(一)细胞遗传学(二)细胞生理学(三)细胞化学五、细胞生物学学科的形成与发展这个新阶段的基本特点可归纳如下:(1)研究对象:细胞(及社会),尤其活细胞(2)研究内容:细胞重大生命活动(3)研究重点:细胞信号调控网络,作为揭示生命活动分子机制方面(4)研究目标:多层次上特别是纳米层次揭示生命活动本质(5)研究特征:多领域、多学科交叉结构&流程示意图1.细胞重大生命活动及其相互关系示意图思考题1.根据细胞生物学研究的内容与你所掌握的生命科学知识,恰当地评价细胞生物学在生命科学中所处的地位及它与其它学科的关系。
第8章--蛋白质分选与膜泡运输 (1)
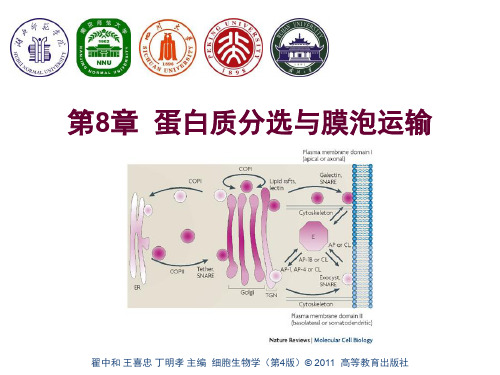
翟中和 王喜忠 丁明孝 主编 细胞生物学(第4版)配套PPT 欢迎使用!
– 从高尔基体TGN向胞内体或向 溶酶体、 黑(色)素体、血 小板囊泡和液泡的运输。
(表8-3 )
– 受体介导的胞吞途径中负责将 物质从细胞表面运往胞内体转 而到溶酶体的运输。
第8章 蛋白质分选与膜泡运输
图8-13 网格蛋白、多角形网格包被 结构及发动蛋白介导的网格蛋白/ 接头蛋白包被膜泡的形成示意图 A.三腿网格蛋白分子:3 条重链,3 条轻链 B. 在体外缺乏膜泡的情况下自组装 的网格蛋白网格包被。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第二节 膜泡运输
内膜系统之间及与质膜之间 的物质传递主要通过膜泡运 输进行。
运输小泡在膜的特定区域以 出芽、内吞的方式产生。表 面具有一个笼子状的由蛋白 质构成的包被(coat)。包 被在运输小泡与靶细胞器的 膜融合之前解体。
膜泡运输及内膜系统结构、 位置的组织依赖细胞骨架, 特别是细胞微管体系。
跨膜运输:
• 进入内质网依赖信号肽; • 进入线粒体、叶绿体、过氧化物酶体依赖导肽或转运肽; • 通过膜上的易位子、内外膜转运复合体蛋白跨膜; • 以单个未折叠的肽链形式跨膜; • 需要通过分子伴侣协助。
膜泡运输:
• 通过出芽,转运,驻泊及最后与靶膜融合运输蛋白、脂质; • 在膜泡的外围组装包被蛋白; (网格蛋白、COPII包被蛋白、COPI包被蛋
( 微 管 组 装 抑 制 剂 )
When the G-stage is missing, Golgi organization is disturbed. Compare normally organized Golgi (left) to a partially scattered circular Golgi assembled by radial centrosomal microtubules in CLASP-depleted cells (right).
高尔基体在细胞分裂中的解体和分裂后重新组装Golgi assembly occurs in 2 stages upon mitotic exit: (a) Video frames illustrating
post-mitotic Golgi assembly in mCherry(一种红色荧光蛋白)-Rab6(定位于高尔基体或后高尔基体区室上的GTP结合蛋白,调节膜
第一节 蛋白质的分选(定向转运)
1 真核细胞中核编码蛋白的分选概述(共翻译转运和后翻译转运)
蛋 白 质 分 选 运 输 的 路 径 图
蛋白通过四种机制转运:
• 跨膜运输:蛋白运入内质网、 线粒体、叶绿体、过氧化物酶 体;
• 膜泡运输:内质网、高尔基体、 质膜、溶酶体、胞内体之间的 蛋白运输;
• 门控通道运输:通过核孔运输; • 细胞质基质中的蛋白转运。
白等); • 只有正确折叠、组装的蛋白才能被运输; • 在运输中被运输蛋白、脂质的方向不会改变。
门控通道转运:
• 细胞核蛋白通过核孔复合体运输; • 蛋白存在核定位信号 (NLS); • 以折叠或组装的形式进行运输。
2 信号假说和内膜系统蛋白质的转运
--G.Blobel和D.Sabatini,1975年提出共翻译转运的信号假说。
• 不仅蛋白质进入内质网需要信号引导,蛋白在内膜系统间运输也需 要信号。如内质网驻留蛋白的KDEL(HDEL)信号和KKXX信号(膜蛋 白),质膜整合蛋白由内质网进入高尔基体的双酸分选信号(AspX-Gln),溶酶体蛋白有高尔基体进入溶酶体的甘露糖-6-磷酸信号 等。
• 水疱性口炎病毒包膜蛋 白的分选信号是存在于 细胞质基质一侧的双酸 分选信号,这种信号也 存在于其他膜蛋白上。
细胞生物学
第八章 蛋白质分选与膜泡运输
西北农林科技大学 生命科学学院细胞生物学教研室
李绍军
本章内容
第一节 细胞内蛋白质的分选 1. 信号假说与蛋白质分选信号 2. 蛋白质分选的基本途径与类型 3. 蛋白质向线粒体、叶绿体和过氧化物酶体的分选 第二节 细胞内膜泡运输 1. COPII与COPI包被膜泡的装载与运输 2. 网格蛋白/接头蛋白包被膜泡的装载与运输 3. 转运膜泡与靶膜的锚定和融合 4. 细胞结构体系的组装
高尔基体重新组装依赖微管:(c) Showing MT nucleation (成核) at Golgi mini-stacks (red), binding of mini-stacks to MT (yellow arrow), and transport along MT (white arrow) resulting in clustering along Golgi-nucleated MT. mCherry-Rab6 (red), GFP-EB3(一种 微管结合蛋白, green). (d) Mini-stack clustering from (c), mCherry-Rab6 alone. Blue arrow, mini-stack where MT nucleates. Yellowarrow, transported mini-stack.
• 没有任何信号(序列)的蛋白是细胞质基质驻留蛋白。
信号肽及信号斑
• 蛋白建立分选信号的两种方式: • 信号位于一段氨基酸序列中被称为信号序列,它暴露在蛋白外部通常位
于肽链端部,也有存在于其他部位的。
• 信号斑由分隔开的氨基酸序列在蛋白折叠时相互接近组成,或者在蛋白 折叠后形成暴露的固定距离的多个信号斑。
泡运输、融合) expressing RPE1(一种视网膜上皮细胞株) cells. Time zero marks approximate onset of telophase(末期开始).
Boxed area is enlarged below. (b) Postmitotic Golgi particle size based on live imaging experiments in NT-control cells. Average fold
increase of Golgi particles relative to time zero is shown. (c) Enlarged box from (a) showing Golgi mini-stack (red) clustering (6–9',
blue and yellow arrows indicate two separate clusters) prior to re-location toward the centrosome (10'). Miller et al.
• Palade发现内质网附着的核糖体 合成分泌蛋白。
• Milstein等发现:在非细胞体系中 合 成 的 IgG 轻 链 比 体 内 真 正 的 IgG轻链N端长出数个氨基酸。
• 在非细胞体系中添加ER膜组分 则可合成正常长度的IgG轻链。
• 分泌蛋白的信号肽:由多肽N端16-26个氨基酸组成。具有三个区域,带正 电荷的N端区,中间的疏水核心区和极性的与成熟肽链连接的C端剪切区, 新生分泌蛋白的信号肽使肽链定向运输到内质网,随后被切除。
• 双酸分选信号指导膜蛋 白从内质网向高尔基体 运输。
上皮细胞顶端和基膜蛋白的分选: 水疱性口炎病毒(VSV)与普通感冒 病毒共同侵染上皮细胞时在内质网 上合成的两种病毒的包膜蛋白被高 尔基体转运小泡分别运输至上皮细 胞的基膜端和游离端。有些膜蛋白 分泌到基膜一侧在通过转胞吞作用 聚集到游离面质膜上。
进入内质网腔的可溶性蛋白没有停止转移序列
含内在停止转移锚定序列(STA)和内在信号锚定序列(SA)的 蛋白在内质网膜上的拓扑特征
蛋定膜 白序整 膜列合 定)蛋 位和白 中开的 的始内 作转部 用移停
序止 列转 (移 内序 部列 信( 号内 锚部 定停 序止 列转 )移 在锚
葡萄糖转运蛋白1等多跨膜α螺旋蛋白的合成和转运:多个开始转移序列(内部信号锚定序列,不切除的信号 肽)和停止转移序列(内部停止转移锚定序列)形成多次跨膜。
蛋白分选(蛋白质寻靶): 蛋白分子在蛋白内部的分选 信号指导下从细胞质运往各种目标细胞器或细胞表面。
• 细胞内合成的蛋白质、脂类等物质之所以能够定向的转运到特定的细胞器 取决于两个方面: – 其一是蛋白质中包含特殊的信号序列(signal sequence)。 – 其二是细胞器上具特定的信号识别装置(分选受体,sorting receptor)。
• 信号识别颗粒, SRP: 由6种不同的多肽和一条7S(300bp左右)RNA组成 的核糖核蛋白复合体,可结合GTP)。
• 信号识别颗粒受体:内质网停泊蛋白 (DP,一种GTP结合蛋白)。 • 易位子:由3-4个Sec61蛋白构成的通道,每个Sec61由3条肽链组成。 • 停止转移序列:与内质网膜的亲合力
in the cell center (C-stage of
Golgi assembly). In CLASP高尔基
体膜上组织微管组装的一种蛋白)-
细胞分裂末期高尔基体重新组装由两套微管体系完成——中心体微管体系(Cstage)和高尔基体微管体系(G-stage,高尔基体有微管组织中心的功能)
depleted cells that have no Golgi-derived microtubules, G-
很高,阻止肽链继续进入网腔,成为 跨膜蛋白。
• SRP 有三个主要活性区: • 信号肽识别区:P54甲硫氨酸与信号肽疏水核
心结合; • 核糖体结合区:P9、P14,阻断进一步翻译; • 结合内质网停泊蛋白的区域。
粗面内质网合成分泌蛋白共翻译转运示意图
除N端信号肽引导蛋白转运 至内质网外,内质网定位 蛋白还可能有开始转移序 列,内在停止转移锚定序 列(STA)、内在信号锚定 序列(SA,起信号肽、开 始转移、锚定的序列)等 内部信号序列指导内质网 膜蛋白的转运。
Polarity of post-Golgi trafficking cannot be maintained in cells that have a disorganized, circular Golgi. Compare normal polarized trafficking toward the cell front (left) to non-directional trafficking pattern in a CLASP-depleted cell (right).