大三遗传学第八章 病毒的遗传分析
遗传学第二版课后部分答案专家讲座

a
c
d
eb
第17页
第七章 细菌遗传分析
10. 在接合试验中,有 Hfr a+ b+ str s × F - a - b - str r,已知 a 和 b 基因控制
营养
需要,先将杂交菌株接种在有链霉素完全培养基上生长,然后在不一样
培养基上测试100个菌落,结果以下:① 添加 a 物质,得40个菌落。② 添
遗传学第二版课后部分答案
第13页
第七章 细菌遗传分析
1. 用一个起源于一个不能合成异亮氨酸细菌菌株(ile -)噬菌体转导一 个不能合成甲硫氨酸细菌菌株(met -)。将接适用肉汤培养基稀释 后涂布在补充有异亮氨酸基本培养基上。另取相同量稀释肉汤培养 基涂布在基本培养基上。基本培养基上长出 18 个菌落, 含有异亮氨酸 基本培养基上长出 360 个菌落。计算标准化重组百分比。
绘制染色体图为何不能以
方式表示?
不能。依据绘制染色体图规则, 图距是依据试验得到重组值去掉 百分号后数字来表示。
遗传学第二版课后部分答案
第4页
第五章 连锁遗传分析
6. 粗糙脉孢菌一个菌株(基因型为 HI)与另一个菌株(基因型为 hi)杂 交, 二分之一后代为 HI 型, 另二分之一后代为 hi 型。请解释该结果。
加 b 物质,得 20 个菌落。③ 基本培养基得 10 个菌落。分别写出这 3 种
菌
Hfr a+ b+ str s × F - a - b - str r
落基因型,计算基因 a 和 b 之间重组率是多少?
40
20
10
MM + a + str MM + b + str
a+ b+ str r(10) a+ b+ str r(10) a - b+ strr(30) a + b- strr(10)
第八章:病毒的遗传分析1
公式如下:
2×rII+ 噬菌体数
重组率= 总 噬 菌 斑 数 =
2×在K上生长的噬菌体数 在B上的噬菌斑数
×100%
12
3、双重感染法绘制rII区段连锁图
操作方法:
用两种rII突变型双重感染B品系,收集溶菌液; 分别接种到B品系、K(λ)品系菌苔上; 考察两个品系菌苔上的噬菌斑数目,就可以计算两个
测得最小的重组值为0.02%,即0.02个图距,相当于2.2bp 表明重组子的最小单位是一对核苷酸。因此将该技术称为基因 的精细作图。
14
三、互补测验
(1).当不能单独在K菌株上生长的两个rII突变体混合感染时 ,它们能象野生型噬菌体那样在K株上生长,这样两种突变型 称为互补的突变型,因为每一种能补偿另一种功能。使两者都 能生长。(T4染色体在宿主细胞中暂时二倍化)
▪ 缺失作图的原理: 凡是能和某一缺失突变型进行重组而恢复野生型核苷酸顺序的,它的
位置一定不在缺失范围内;凡是不能重组的,它的位置一定在缺失范围 内。
▪ 缺失作图方法: 在0.5ml大肠杆菌B菌株培养物中加入1滴缺失型噬菌体和1滴待测的
rII突变噬菌体,几分钟后,取一滴菌液加在灭菌的纸条上,铺在长有大 肠杆菌K(λ)菌株的平板上,经培养后如在纸条覆盖的区域内形成清 晰的噬菌斑,则说明发生重组,产生了野生型的噬菌体。若不能形成噬 菌斑,则说明它们有相同重叠的缺失部位。
突变位点 于不同顺 反子中
顺式试验
反式试验
A
B +– A
B
A
B ++ A
B
总结——基因
•突变子(muton):是性状突变时,产生突变的最小单位。 一个突变子可以小到只有一对核苷酸。
遗传学第八章 核外遗传分析课件

2、 性比(sex-ratio,SR)因子
◇SR因子是胞质中的一种原生动物,在雌蝇 和雄蝇中都能发现,但对发育中的雄性幼 虫是致死的,所以后代中雌蝇比例远大于 雄蝇比例。
◇将SR雌蝇的卵细胞质注入正常雌蝇可诱导 SR现象。
◇有证据表明,产生雄性致死毒素的可能是原 生动物内的病毒。
基因型与表型的关系
细胞质基因 正常(N) 不育(S)
核基因型
RfRf(可育) Rfrf(可育) rfrf(不育) N(RfRf)可育 N(Rfrf)可育 N(rfrf)可育 S(RfRf)可育 S(Rfrf)可育 S(rfrf)不育
遗传学 第八章 核外遗传分析
(二)可能的遗传机制 1、线粒体与雄性不育的关系 2、叶绿体与雄性不育的关系
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
• KSS综合征(Keams-Sayre Syndrome)是多 系统线粒体病,主要症状为眼肌麻痹和色素性视 网膜炎。患者骨骼肌细胞mtDNA有2.0kb7.0kb的缺失。 遗传学 第八章 核外遗传分析
五、叶绿体遗传及其分子基础
(一)衣藻的叶绿体遗传
◇ 衣藻细胞中只有一个叶绿体,约含50个 拷贝的环状ds-DNA分子。 ◇ 不同交配型(mt+,mt-)的单倍体衣藻 杂交,形成短暂2n时期,进行减数分裂。 ◇ 虽然杂交双方融合时为合子提供等量细 胞质,但叶绿体只由mt+方传递,表现单亲 遗传。
遗传学 第八章 核外遗传分析
遗传学 第八章 核外遗传分析
(二)叶绿体遗传的分子基础 1、叶绿体基因组
大小:环状双链DNA分子。大小120-190kb。 其基因序列中不含5-甲基胞嘧啶。
◆ cpDNA编码约100种蛋白质和RNAs,包 括45个编码RNA的基因,27个编码与基因表达 有关的蛋白的基因,18个编码类囊体膜的蛋白基 因和10个与电子传递功能有关的基因。
病毒的遗传与变异ppt课件

1.2、突变率
DNA病毒复制具有校正(proofreading) 功能 ,RNA的复制缺少校正机制,病毒 RNA复制的错误率远远高于病毒DNA。
1.3、诱变
物理诱变 化学诱变 定点诱变:将病毒基因组任何既定部位的核
苷酸替换,或者使之缺失,或者插入另一段 核苷酸。
2、基因重组
分子内重组 重配
3.1、补偿作用
是指在感染的细胞中,病毒蛋白质之间由 于相互作用的结果,拯救了一种或两种病 毒或增加了病毒的产量。
Complementation
和 (或)
Progeny virus assembled using wt N and wt M proteins Genomes in progeny are either ts M or ts N
1.1、突变类型
基因组与表型变异
点突变:少数几对碱基的缺少、插 入或置换。
遗传结构改变:大段染色体的缺失、 重复、移位和倒位等。
自发突变与诱发突变
1.1.2、基因组变异 位点变异
缺损型干扰(defective interfering,DI)突变株
这是一种缺失突变的产物。自身不能复制, 只有在亲本野生株作为辅助病毒存在时才 能复制,但又干扰亲本病毒的复制,导致后 者数量减少。
Reassortment by independent assortment during dual infection.
adapted from Treanor JJ Infect. Med. 15:714
最近重配
此次甲型H1N1流感病毒包含北美和欧 亚猪流感病毒、人流感病毒以及禽流感 病毒的基因片段。正因为其基因片段不 同于传统的H1N1病毒,因此属于新病 毒。
第八章病毒的遗传分析

第八章病毒的遗传分析第八章病毒的遗传分析8.1 病毒的形态结构与基因组 8.2 噬菌体的增殖和突变型 8.3 噬菌体突变型的重组测验 8.4 噬菌体突变的互补测验 8.5 噬菌体T4rⅡ的缺失突变与作图8.6 λ噬菌体的基因组与位点专一性重组 8.7 环状排列与末端重复补充:细菌和病毒遗传研究的意义细菌和病毒在遗传研究中的优越性细菌和病毒的拟有性过程一、细菌和病毒在遗传研究中的优越性1.繁殖世代所需时间短。
2.易于管理和进行化学分析。
3.遗传物质较简单。
4.便于研究基因的突变。
5.便于研究基因的作用。
6.可作为研究高等生物的简单模型。
二、细菌和病毒的拟有性过程1.真核生物基因分离、自由组合及连锁交换均通过有性过程(减数分裂——受精)实现。
2.细菌细胞内除了染色体外还有一些寄生性复制因子(如噬菌体和质粒,也被称为核外或染色体外因子),它们可以在细胞间传递,并且形成细菌染色体间以及细菌染色体与核外遗传因子间的重组体。
这种重组体结构类似于真核生物减数分裂过程中形成的重组体结构。
8.1 病毒的形态结构与基因组二、细菌和病毒的拟有性过程3.拟有性过程——引起细菌、病毒间遗传物质转移与重组的过程。
拟有性过程的存在是细菌、病毒在遗传学研究,特别是作为真核生物的模型研究遗传重组和基因结构的重要前提。
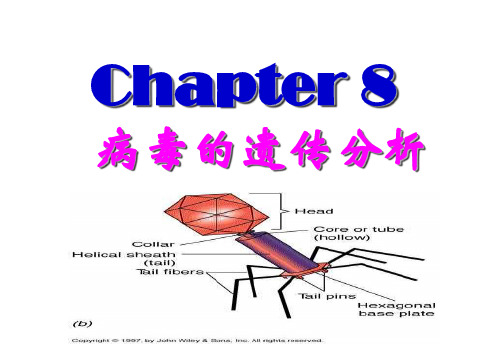
8.1.1 病毒的形态结构1.形体极微小,只有在电子显微镜下才能观察到;2.化学组成简单,主要是核酸和蛋白质;无细胞结构,为蛋白质外壳包裹核而成的颗粒;3.只含一种核酸(DNA或RNA),是单倍体,只有一条染色体;4.病毒的多样性;5.缺乏独立代谢能力;16.繁殖方式独特,只能依赖宿主活细胞的代谢机器;7.具有双重存在方式,或营专性寄生在活细胞内,或在细胞外以大分子颗粒状态进行传播;8.对干扰素敏感,而对抗生素不敏感。
病毒的基本结构病毒粒子(毒粒):基本结构:蛋白质衣壳位于毒粒中心的核酸构成的核壳包膜——脂质或脂蛋白蛋白质或糖蛋白突出的结构——刺突螺旋对称,二十面体对称,复合对称8.1.2 病毒的基因组病毒的类型根据宿主细胞的不同可将病毒分为: 1.植物病毒 2.动物病毒 3.细菌病毒(又称噬菌体) 4.亚病毒类病毒拟病毒朊病毒每种病毒只含一类核酸(DNA或RNA)含DNA的病毒称DNA病毒含RNA的病毒称RNA病毒病毒基因组的类型极为多样化有单链(ss)与双链(ds) 正链(+)与负链(-) 正链:基因组碱基排列顺序与mRNA相同负链:基因组碱基排列顺序与mRNA互补有线状(L)与环状(O)噬菌体以线状的dsDNA居多。
遗传学病毒的遗传分析

(二)基因内互补 例外情况。如:沙门氏杆菌甘油磷酸脱氢 酶基因;大肠杆菌和脉孢菌色氨酸合成酶 基因;脉孢菌谷氨酸脱氢酶基因。
1、基因内互补和基因间互补的区别 P200 2、基因内互补的作用机制 P202图8-14
遗传学病毒的遗传分析
五、噬菌体T4 rⅡ缺失突变与作图
(一)缺失作图的原理 ◇ 缺失突变(deletion mutation):一个
K(λ)
野生型 rⅡ
小噬菌斑 小噬菌斑 大噬菌斑 无噬菌斑
遗传学病毒的遗传分析
3、宿主范围突变型
◆ 宿主范围(host range,h):h-表示可同
时感染两种不同的E.coli菌株B,B-2,噬
菌斑透明。h+表示只感染菌株B,噬菌斑半 透明。
遗传学病毒的遗传分析
三、噬菌体突变型的重组测验 (一)Benzer的重组测验和基因的
(1)温度敏感突变 • 热敏感突变 • 冷敏感突变 (2)抑制因子敏感突变(sus突变)
sus突变 su+突变:宿主菌含有抑制基因su+, sus突变噬菌体感染此类菌时能产生后代。
遗传学病毒的遗传分析
2、噬菌斑形态突变
(1)T4噬菌体简介 T4噬菌体为T-偶列噬菌体之一,基因组为
165Kb的双链DNA分子,包括160个基因, 宿主为E.coli。 形态特征 环状遗传学图
遗传学病毒的遗传分析
遗传学病毒的遗传分析
遗传学病毒的遗传分析
在反式测验中,如两个突变之间能互补,则表 明两个突变是位于两个基因(顺反子)内的突变。 如两个突变之间不能互补,则表明两个突变是位于 同一个基因内的突变。
遗传学病毒的遗传分析
• 顺反子:一个不同突变之间没有互补的 功能区称为顺反子(cistron)。
8、病毒的遗传分析z

含多个基因位点,是遗传上的一个作用(功能)单
位,但不是一个突变单位或重组单位。
同一顺反子内发生的突变间不能互补
• 顺反试验:指将两个待测突变分别处于顺 式和反式,根据其表型确定两个突变是否 是同一基因的试验。
溶源性细菌
• 原噬菌体 (prophage):整合进细菌基因组中
的噬菌体称为原噬菌体。在细菌染色体的特异位
点整合进整个噬菌体基因组。
• 溶源菌(lysogenic bacterium):含有原噬菌
体的细菌具有产生和释放噬菌体的潜力,这种细
菌称为溶源菌。 • 非溶源性细菌(non-lysogenic bacterium): 失去原噬菌体的细菌。
• 重组值计算:rxry的数量与r+r+ 相同,计算 时r+r+ 噬菌体数×2。
最小的结构单位
• 此种测定方法称为重组测验,理论上此方法测定 重组频率非常灵敏可以获得小到0.002%,即 0.002个图距单位。 • 但实际结果不会低于0.02%;可推断基因内存在 最小重组单位,Benzer将最小重组单位定义为重 组子。
• 微细的遗传图
噬菌体的基因重组与作图
• 凯泽(Kaiser A. D., 1955)最先进行λ噬菌 体的重组作图试验。紫外线照射处理 获5个噬 菌体突变系,产生不同噬菌斑:
s系:小噬菌斑; mi系:微小噬菌斑; c系:完全清亮的噬菌斑; co1系:中央环之外部分表现清亮的噬菌斑; co2系:更浓密的中央环噬菌斑。
8、病毒的遗传分析
1、病毒的形态结构与基因组
根据宿主不同,可以分为: 动物病毒 植物病毒 细菌病毒-----噬菌体 基本形态:
Genetics 8 病毒遗传分析
图 8-10 顺反位置效应示意图
• 遗传上的突变单位和重组单位是突变子 (muton)和重组子(recon),突变子是基因 内改变后可以产生突变表型的最小单位。 它只相当与一个核苷酸对,不能将其定义 为一个基因。重组子是基因内不能有重组 分开的遗传单位,也不能将其定义为一个 基因。 • 所以:基因可分,可分为很多突变子和重 组子。
• 若双重感染的细菌不产 生子代噬菌体,那么这 两种突变型一定有一个 相同功能受到损伤。
• 互补测验结果发现:
• rⅡ 突变型可分成rⅡ A和rⅡ B两个互补群。
所有rⅡ A突变型的突变位点都在rⅡ 区的
一头,是一个独立的功能单位,所有rⅡB突
变型的突变位点都在rⅡ 区的另一头,也是
一个独立的功能单位.
• 缺失作图:Benzer根据这一原理很方便地把数 千个独立的rⅡ突变定位在rⅡ遗传图上更小的区 段内,此方法称缺失作图。 • 凡是能和某一缺失突变进行重组的,他的位置一 定不在缺失范围内,凡是不能重组的,它的位置 一定在缺失范围内。
缺失 1 缺失 2 a b c 细线表示缺失区,二者分别与各种突变体杂交,缺失2只 有与a区中突变体杂交才能产生野生型重组体,缺失1只 有与 c区突变体杂交才能产生野生型重组体,但2个缺失 与b区的突变体杂交均不能产生野生型重组体。
噬菌体突变型 r- 快速溶菌,大噬菌斑 野生型 r+ 缓慢溶菌,小噬菌斑 寄主范围突变型 h- 感染大肠杆菌品系1和2 野生型 h+ 只能感染品系1 h+r- 与 h-r+同时感染大肠杆菌品系1,二者发生重组。子代噬 菌体感染品系1/2 的混合物,计算重组子出现的频率。p193
第8章 病毒的遗传分析[1]
![第8章 病毒的遗传分析[1]](https://img.taocdn.com/s3/m/ba81a46addccda38376bafa8.png)
4. 在数分钟内,所有的细菌核
酸和蛋白质合成都被抑制
5. 噬菌体大分子合成,细菌的核酸被降解
(1) 噬菌体DNA复制。 (2) 噬菌体外壳蛋白合成。
6. 噬菌体组装
(1) DNA被包到头部 (2) 组装尾部 (3) 装上尾丝
su+
产生子代噬菌体(许可条件)
su能侵入,不能产生正常子代噬菌体 (限制条件)
Tyr: UAU Anticodon: AUA Muton: AUC Stop codon:UAG
UAC AUG AUC UAG
这种sus突变型在效应的抑制基因宿主中可产生后代,原因是将 终止密码子(UAA/UAG/UGA)代换成一个特殊的氨基酸,防止在 终止密码子上提前终止
•温度敏感 突变
冷敏感突变型(cold sensitive mutants):突变型在低温下致死
•抑制敏感突变(suppressor-sensitive mutation,简称
sus):是原来正常的密码子变成了终止密码子,产生
无活性蛋白质
无义突变与无义抑制基因
终止密码子因为不编码任何氨基酸,所以称为无义密 码子,它们是蛋白质合成的终止信号
两者与tsC距离较远,tsC属非选择标记。
测验步骤:
amA + tsC × +amB +
su许可条件
野生型重组子 ++tsC、++tsC+
限制条件
总数
tsC和tsC+所占比例, 确定基因排列顺序
tsC+
amA + tsC
正交
+ + tsC
遗传学_第二版_课后答案(1~8章)

40 20 10
MM + a + str
MM + b + str
MM + str
a+ b+ str r(10) a+ b+ str r(10) a - b+ str r(30) a + b- str r(10)
a+ b+ str r
第七章 细菌的遗传分析
6. 测验 5 个点突变(a-e)与下面拓扑图表示的 5 个缺失杂交产生野生型重组 的情况。(+ = 重组,0 = 没有重组)。结果列在表中。确定点突变的顺序。
缺 1 2 3 失 4 5来自ab c d e
0
+ 0 0 0
0
+ 0 + +
+
+ + 0 0
+
0 + 0 0
+
+ 0 0 + a c d e b
第六章 真核生物的遗传分析
• 第6题: • 含1号染色体的克隆B、D不同时有任何酶活性; • 含2号染色体的克隆A、D都有II、IV酶活,而不含 该染色体的克隆都不具有这两个酶的活性,表明 这两个基因定位于2号染色体; • 3号染色体不含上述基因; • 因此只能判断II、IV两个基因定位于2号染色体.
(30+10)/(30+10+10)= 80%
第七章 细菌的遗传分析
12. 大肠杆菌 Hfr gal + lac +(A)与 F — gal — lac —(B)杂交,A 向 B 转移 gal + 比较早而且频率高,但是转移 lac + 迟而且频率低。菌株 B 的 gal + 重组子仍 旧是 F — 。从菌株 A 可以分离出一个突变体称为菌株 C ,菌株 C 向 B 转移 lac +早而且频率高,但不转移 gal + 。在 C × B 的杂交中,B 菌株的 lac + 重 组子一般是 F+ 。问菌株 C 的性质是什么?试设计一个实验分离这个菌株。
遗传学_第二版_课后答案

幻灯片 1习题参考答案第四章第五章幻灯片2第四章孟德尔式遗传分析2. 在小鼠中,等位基因 A 引起黄色皮毛,纯合时不致死。
等位基因 R 可以单独引起黑色皮毛。
当 A 和 R 在一起时,引起灰色皮毛;当 a 和 r 在一起时,引起白色皮毛。
一个灰色的雄鼠和一个黄色雌鼠交配,F1 表型如下:3/8 黄色小鼠, 3/8 灰色小鼠, 1/8 黑色小鼠, 1/8 白色小鼠。
请写出亲本的基因型。
A_R_A_rrAaRrAarraaR_aarrA_rrA_R_幻灯片3第四章孟德尔式遗传分析3. 果蝇中野生型眼色的色素的产生必需显性等位基因 A。
第二个独立的显性基因 P 使得色素呈紫色,但它处于隐性地位时眼色仍为红色。
不产生色素的个体的眼睛呈白色。
两个纯系杂交,结果如下:AaXPXp AaXpY解释它的遗传模式,并写出亲本、F1 和F2 的基因型。
A/a 位于常染色体上,P/p 位于X染色体上;基因型aa 的个体眼睛呈白色,基因型A_XP_ 的个体眼睛呈紫色,基因型A_XpXp、A_XpY 的个体眼睛呈红色。
幻灯片4第四章孟德尔式遗传分析4. 一条真实遗传的棕色狗和一条真实遗传的白色狗交配,所有F1 的表型都是白色的。
F1 自交得到的 F2 中有 118 条白色狗、32 条黑色狗和 10 条棕色狗。
给出这一结果的遗传学解释。
分析: 子二代分离为 12:3:1,可看作9:3:3:1 的衍生,白色与有色(黑 + 棕)之比 3:1 ,而在有色内部,黑与棕之比也是 3:1,表明遗传很有可能涉及有两对基因之差。
假设: 1. 基因 A 控制白色,即基因型A_B_、A_bb 为白色。
2. 有显性基因 A 时,B(黑色)和 b(棕色)不表现显隐性关系;3. 无显性基因 A 即 aa 时, B(黑色)和 b(棕色)表现显隐性关系。
P 棕色×白色F1 白色118 32 10F2 12白色 : 3黑色 : 1棕色aabb AABBAaBbA_B_ A_bb aaB_ aabb在此,显性基因A 对另一显性基因B 是上位性的。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
3) 反式构型:两个突变分别位于两条同源染色
体上的基因组合。 4) 顺式构型:两个突变位于同一染色体上的基
因组合。
- + - +
- + + -
顺式
反式
5) 互补测验与重组测验的区别: 重组测验中检测的是重组类型的产生,以遗传 图距的方式确定基因的空间关系;
互补测验中检测的是具有亲代基因型的子代噬
理论上,重组率=2×10-6
实际上,重组率=0.02%。
T4噬菌体染色体1.8×105bp,1500图距单位。
0.02 1500 x 2.4bp 5 x 1.8 10
即0.02个图距单位约2bp 。 说明:基因内相邻核苷酸对的突变可发生重组,
重组子的单位可小到2bp 。
基因内部可分,基因是一个功能单位,但
8.5.1 缺失作图原理 ⑴ 缺失突变的特点 ① 多碱基对的缺失; ② 具不可逆性; ③ 有部分相同缺失的突变型间不能通过重组 恢复野生型表型。
⑵ 缺失作图的原理 凡是能重组且产生野生型的,点突变一定 不在缺失区内; 凡是不能重组且不产生野生型的,点突变 一定在缺失区内。
8.5.2 缺失作图的方法 ⑴ 将待测点突变(X)型先与几个最大的缺失
菌体,即在限制条件下突变型能否生长。
T4噬菌体的rⅡ区3000多个突变可分为rⅡA和
rⅡB两个互补群。
1
y
2
+ 3 +
+ 4
6) 顺反子、突变子、重组子 1 顺反子(cistron):不同的突变型之间没 有互补的功能区,即一个功能水平上的基因。 2 突变子(recon) :顺反子内部能发生突变的 最小单位. 3 重组子(muton ):顺反子内部出现重组的 最小区间.
ra 24 h rb 12.3 h rc 1.6 h
4个基因可能的排列: ra ra ra ra rb rb rc rc h h h h rc rc rb rb
再做杂交:
rcrb+ × rc+rb
结果表明: rc——rb的重组值>rb——h 所以,h应位于rb 及rc之间,
即:rc——h——rb。
(3)重组值的计算
重组噬菌斑数 重组值 噬菌斑总数 (h r ) ( h r ) 100% 噬菌斑总数
要测定不同速溶突变体ra-、rb-、rc-之
间的距离,可以分别进行h+ra-×h-ra+、
h+rb-×h-rb+、h+rc-×h-rc+重组实验。
h+rx-×h-rx+ 噬菌斑数目及重组值
3467
3729
520
474 853 965 162 172
∨
∨ ∨ ∨
∨
∨
合计
10342 12.9 20.8 27.1
m 12.9
r
20.8
tu
8.4 噬菌体突变型的互补测验
8.4.1 互补测验与顺反子 1)互补测验:根据基因的功能确定两个基因是 否等位的测验方法. 2)互补作用:两个突变型细胞的两条同源染色 体同处在一个杂合子时,野生型基因补偿突变基 因的缺陷而使表型恢复正常。否则,两种突变型 一定具有相同的功能损伤。
8 病毒的遗传分析
主要内容: 1 噬菌体的突变型及基因重组的特点; * 2 噬菌体的重组测验与互补测验;
* 3 噬菌体T4rⅡ的缺失作图;
4 噬菌体基因组与位点专一性重组; 5 噬菌体的线状染色体与环状连锁图。
8.1 病毒的形态结构与基因组
8.1.1 病毒的形态结构 病毒:一种不具备细胞结构的生物体,只有寄 生在宿主细胞中才能生存。其基本结构包含核 酸和外壳蛋白。有些病毒的外壳外还有一层脂 质或脂蛋白组成的包膜,有的包膜表面还有蛋 白质或糖蛋白突出的结构刺突。
个基因。
在限制条件下,不能长出噬菌斑: 说明:两个突变型不能发生功能互补,是 同一基因。
2 互补测验及结果
X174突变的互补测验结果 顺反子 突 变 型 A am8,am18,am30,am33,am35,am50,am86,tsl28, B am14,am16,och5,ts9,tsl16,och1,och8,och11, C och6 D am10,amH81, E am3,am6,am27, F am87,am88,am89, amH57,op6, op9,tsh6,ts41D G am9,am32,ts,ts79 H amN1,am23,am80,am90,ts4
突变体分别杂交,以确定该突变位点所属的大
范围; ⑵ 将点突变与有关的的几个小缺失突变体分 别杂交,以缩小突变位点的范围。
A5
利用T4rⅡ缺失突变型的缺失部位所构建的精细结构图
8.6 λ噬菌体的基因组与位点专一性重组 8.6.1 λ噬菌体的基因组 头部基因: 7个 尾部基因:11个 噬菌斑形 成 必需基因
杂交组合 h+ r h+ra-×h-ra+ h+rb-×h-rb+ h+rc-×h-rc+
-
噬菌斑基因型的% h-r + 42.0 56.0 59.0 h+ r + 12.0 5.9 0.7 h-r
-
重组值
34.0 32.0 39.0
12.0 6.0 0.9
24% 12.3% 1.6%
则:ra、rb、 rc与 h之间的顺序:
是双链RNA,藻类病毒都是双链DNA。
8.2 噬菌体的增殖与突变型
8.2.1 噬菌体的繁殖 (1)烈性噬菌体的增殖 如:E.coli的T噬菌体系列(T1-T7)。 T偶列 噬菌体 头部:双链DNA分子 颈部:中空的针状结构及外鞘
尾部:由基板、尾针和尾丝组成
(2)温和噬菌体的增殖 ①λ噬菌体及其生活周期 λ噬菌体的宿主是大肠杆菌K12。噬菌体侵 入后,细菌不裂解→附着在E.coli染色体的gal 和bio位点间的attλ座位上→整合到细菌染色 体上,并能阻止其它λ噬菌体的超数感染。 超数感染:一个细菌受一个以上同种噬菌体感 染的现象。
8.1.2 病毒的基因组 DNA病毒和RNA病毒;
噬菌体的核酸大多为DNA;植物病毒的核酸大多
为RNA,少数为DNA;动物病毒部分是RNA ,部
分是RNA;真菌病毒的核酸大多是RNA。
RNA病毒多为单链,线状,有正、负链之分; DNA病毒多为双链,少数为单链正DNA,腺病毒 为-DNA;噬菌体多为线状双链DNA。真菌病毒都
进受体细胞后, λ噬菌体立即从供体染色体上脱
落下来自主繁殖,最终使受体细胞裂解,释放出游 d+ c+ λ 离的噬菌体。
b+ a+ 原点
8.6.3 原噬菌体的整合与切除 ⑴ 原噬菌体的整合与正常切除
⑵ 原噬菌体的异常切除
整合态
⑶ 温和性噬菌体的互补试验与作图
某些d gal品系与图谱左臂顺反子中典型sus突变之间重组
② P1噬菌体
P1噬菌体并不整 合到细菌的染色
体上,而是独立
地存在于细菌细 胞质内。
8.2.2 噬菌体的突变型
(1)条件致死突变型 ① 温度敏感突变型:实质是氨基酸的替换,蛋 白质失去活性。 ② 抑制因子敏感突变(sus):实质是原来正常 的密码子变成了终止密码子,因而翻译提前终止, 不能形成完整肽链而产生有活性的蛋白质。 宿主 su-:不产生子代(限制条件) su+:产生子代(许可条件)
co1 + + mi + mi co1 +
双交换型 s + 合计
s 3.83 co1 6.21 8.32+2×0.86=10.04
mi
8.3.4 T4噬菌体的突变型的三点测交作图
类型 数目 重组率
亲本类型 单交换 Ⅰ
单交换 Ⅱ 双交换型
m r tu + + + m + + + r tu
m r + + + tu m + tu + r +
① 噬菌斑形态突变型
野生型r+:小而边缘模糊的噬菌斑。
原因:有两个以上的噬菌体侵染一个细菌时,
出现溶菌阻碍现象,混有裂解和未裂解的细胞。
突变型r:大而边缘清晰的噬菌斑。 原因:无溶菌阻碍现象。
鉴别:噬菌斑大小。
② 寄主范围突变型 表8-4 T4野生型和突变型的区别
类 型
不同大肠杆菌菌斑平板上表型
E.coli B E.coli K() 小噬菌斑 S 小噬菌斑
复制所需基因:O、P
裂解、释放所需基因:S、R基因 附着区:att 专一性重组:int、xis
噬菌斑形 成非必需 基因
溶源化所需:CI、CII、CIII
重组必需:exo、rex
8.6.2 λ原噬菌体与合子诱导
合子诱导: Hfr(λ)× F-的杂交中,供体菌带有 λ噬菌体,受体菌对λ噬菌体敏感,染色体从供体 向受体转移时,当带有λ噬菌体的染色体部位转移
SUS噬菌体
根据在带有专一性抑制基因的宿主中的非致死性: Ⅰ 琥珀突变(amb): UAG;Ⅱ 赭石突变(och):UAA Ⅲ 乳白突变(op): UGA 表8-2 不同宿主菌中sus突变噬菌体的表型
噬菌体基因型 野生型 sus amber su+ -
宿主菌基因型 su+amb su+och su+op
+ + + + + -sus ochres Nhomakorabeas opal
-
-
+
-
+
5种琥珀抑制基因的性质 琥珀型抑 制基因 su1+ su2+ su3+ su4+ su5+ 插入的 氨基酸 丝氨酸 谷氨酰胺 酪氨酸 酪氨酸 赖氨酸 合成的蛋白质 占野生型% 28 14 55 16 5 赭石型抑 制基因