细胞质遗传并非都是母系遗传
母性遗传与细胞质遗传并非一回事
母性遗传与细胞质遗传并非一回事摘要细胞质遗传一般表现为具母系遗传的特征。
随着DNA分了标记技术的发展和应用,人们己发现在动物及植物中均存在有低频的线粒体DNA单亲父系遗传及双亲遗传的现象。
对质体DNA遗传的研究表明,被了植物的质体DNA大多表现为母系遗传,而裸了植物的质体DN A则卞要表现为父系遗传的方式,同时也发现存在其它的遗传规律。
关键词细胞质遗传母系遗传线粒体DNA 质体DNA本世纪初,在孟德尔定律被重新发现后的1909年,德国学者科伦斯(Co rren s)和鲍尔(B au r)分别在紫茉莉(lVl irabilis jalapa)和天竺葵((Pelargonium zonale)中发现叶色的遗传不符合孟德尔定律,而表现为细胞质遗传现象。
这一发现是对孟德尔定律的挑战和补充。
研究表明,大多数物种的细胞质性状表现为母系遗传的特征。
进而有些学者甚至某些遗传学教科书中也将细胞质遗传与母系遗传这两种现象混为一谈,将这两个概念等同起来,并认为细胞质遗传即为母系遗传。
80年代以来,随着分了生物学技术的发展,将DNA分了标记应用于细胞质遗传研究,从DNA分了水平上研究细胞质遗传物质的变异,使得人们对细胞质遗传现象有了更进一步深入的认识。
下面就细胞质遗传的卞要物质基础线粒体DNA和叶绿体DNA的遗传研究进展作一概述,使我们重新认识细胞质遗传这一现象和概念。
在遗传学教学中,经常会遇到:个体表现型与母木性状一致的现象。
其遗传方式是否是细胞质遗传呢?下面就以具体一例进行分析。
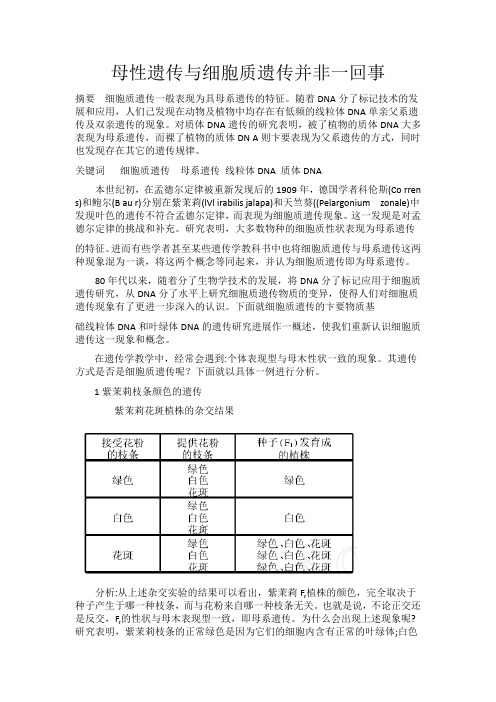
1紫茉莉枝条颜色的遗传紫茉莉花斑植株的杂交结果分析:从上述杂交实验的结果可以看出,紫茉莉F,植株的颜色,完全取决于种子产生于哪一种枝条,而与花粉来自哪一种枝条无关。
也就是说,不论正交还是反交,F,的性状与母木表现型一致,即母系遗传。
为什么会出现上述现象呢?研究表明,紫茉莉枝条的正常绿色是因为它们的细胞内含有正常的叶绿体;白色枝条是山于质体的遗传物质发生了突变,不能形成叶绿素,细胞中只含有白色体;花斑枝条的叶中则含有3种不同的细胞:只含有叶绿体的细胞,只含有白色体的细胞、同时含有叶绿体和白色体的细胞。
遗传学细胞质遗传

雄性不育得遗传决定类型
核质互作雄性不育型
由细胞质遗传因子和核基因 互相作用共同控制花粉育性 得类型
大多数正常品种与之杂交后 代仍为雄性不育
少数携带特殊恢复基因得品
种与之杂交后代正常可育
N
雄性不育得遗传决定类型
细胞质雄性不育型
决定花粉败育得基因在细胞质中 任何正常品种与其杂交,F1均为雄性不育 只能被保持而不能被恢复
胞质不育基因得载体
线粒体基因组(mtDNA)就是雄性不育基因得载体
不育系线粒体得亚显微结构和 mt DNA 与正常植株(如保持 系)有明显得差异
叶绿体基因组(cpDNA)就是雄性不育基因得载体
不育系和保持系之间在叶绿体超微结构和叶cpDNA上存在 明显不同
质核互作雄性不育性得发生机理
质核互补控制假说
终止子 亮氨酸 leu 异亮氨酸 ile 精氨酸 arg
例外情况
色氨酸 trp 苏氨酸 thr 甲硫氨酸 met
终止子
发现得生物
人和酵母得线粒体 酵母得线粒体 人得线粒体 人得线粒体
线粒体基因组
线粒体就是半自主性得细胞器
mtDNA虽能自主复制,但需要核基因组为其编码 DNA聚合酶 线粒体虽有自己得核糖体、tRNA,并能在线粒体内 翻译mtDNA转录得mRNA,但其核糖体蛋白质由核基 因编码 线粒体膜蛋白大多从核基因组中转录,在细胞质中合 成后再转运进入线粒体
叶绿体基因组
叶绿体就是半自主性得细胞器
cpDNA虽能自主复制,编码其核糖体RNA及部分 tRNA,但不能编码自身得DNA复制、转录和翻译过 程所需得全部蛋白质因子和酶 不能编码类囊体所有得结构蛋白和酶系,膜脂代谢所 需得酶系
非细胞质组分得遗传因子
细胞质遗传和细胞核遗传之间的关系

细胞质遗传和细胞核遗传之间的关系1.引言1.1 概述细胞质遗传和细胞核遗传是细胞传递遗传信息的两个重要方面,它们在细胞功能和特征的传递中起着不可或缺的作用。
细胞质遗传主要指的是通过细胞质中的线粒体和叶绿体等细胞质组织传递的遗传信息,而细胞核遗传则是指通过细胞核中的染色体传递的遗传信息。
相较于细胞核遗传,细胞质遗传具有一定的特点和重要性。
首先,细胞质遗传发生在细胞质中的染色体外DNA上,其特点是遗传信息的传递相对稳定。
细胞质中的线粒体和叶绿体内含有独立的DNA分子,通过细胞分裂和有性生殖过程中的细胞质遗传,这些细胞质DNA可以在后代细胞中相对稳定地传递下去。
因此,细胞质遗传在维持细胞的正常功能和代谢过程中具有重要的作用。
其次,细胞质遗传与细胞功能的表现密切相关。
许多重要的细胞功能和特征,如能量代谢、细胞呼吸以及光合作用等,都与细胞质中的线粒体和叶绿体密切相关。
细胞质遗传的变异可能会引起这些细胞功能的异常或改变,进而导致细胞的生理和形态特征发生变化。
与细胞质遗传相比,细胞核遗传也具有其独特的特点和重要性。
细胞核遗传的主要特点是通过细胞核中的染色体传递遗传信息,这些染色体内含有大部分的基因和遗传信息,编码着细胞的大部分蛋白质和生物功能。
细胞核遗传对于细胞的正常发育和功能发挥着至关重要的作用。
在细胞遗传过程中,细胞质遗传和细胞核遗传之间存在着密切的关系和相互作用。
细胞质中的线粒体和叶绿体所带有的DNA,与细胞核中的染色体共同决定了细胞的特征和功能。
细胞核中的基因调控和表达也会对细胞质中的线粒体和叶绿体功能产生影响。
因此,细胞质遗传和细胞核遗传的相互作用是细胞遗传调控的重要方面,它们共同决定了细胞的功能和特征表现。
综上所述,细胞质遗传和细胞核遗传之间存在着密切的关系和相互作用。
它们在细胞的正常发育、功能表现以及遗传特征的传递中起着不可或缺的作用。
深入理解和研究细胞质遗传和细胞核遗传之间的关系,对于揭示细胞生物学的奥秘以及人类疾病的治疗和预防具有重要的意义。
遗传图解的判断及习题

1.首先判定是否为母系遗传(细胞质遗传)若系谱中,女性患者的子女全部患病,女性正常的子女全部正常,即子女的性状总是与母方一致,则为母系遗传;否则不是母系遗传,即为细胞核遗传。
2. 其次判定是否为伴Y染色体遗传若系谱中,遗传具有“父传子,子传孙”的特点,即患者都是男性且有“父→子→孙”的传递规律,则为伴Y染色体遗传;否则不是伴Y染色体遗传。
3.再次判定是显性遗传还是隐性遗传(1)若双亲正常,其子代中有患者,则此单基因遗传病一定为隐性遗传(即“无中生有为隐性”)。
(2)若患病的双亲生有正常后代,则此单基因遗传病一定为显性遗传(即“有中生无为显性”)。
如果没有上述明显特征,则采用假设法,即先假设为显性或隐性遗传,如果与遗传图谱相符,则假设成立,否则假设不成立。
4.判定是伴X染色体遗传还是常染色体遗传(1)在确定是隐性遗传的前提下:①若女患者的父亲和儿子都是患者,则最可能为伴X染色体隐性遗传病,如图1所示。
②若女患者的父亲和儿子中有正常的,则一定是常染色体隐性遗传病,如图2、3所示。
(2)在确定是显性遗传的前提下:①若男患者的母亲和女儿都是患者,则最可能为伴X染色体显性遗传病,如图4所示。
②若男患者的母亲和女儿中有正常的,则一定为常染色体显性遗传病,如图5、6所示。
遗传图解综合练一.选择题(1-6为单选题;7-8为 双选题)1.右图为人类某种单基因遗传病的系谱图,II4为患者。
相关叙述不合理的是A .该病属于隐性遗传病,但致病基因不一定在常染色体上B .若I2携带致病基因,则I1、I2再生一患病男孩的概率为1/8C . II3是携带致病基因的概率为1/2D .若I2不携带致病基因,则I1的一个初级卵母细胞中含2个致病基因 2.(07广东理基)下列最可能反映红绿色盲的遗传系谱图是3.(09广东理基)图10是一种伴性遗传病的家系图,下列叙述错误的是A .该病是显性遗传病,Ⅱ—4是杂合体B .Ⅲ—7与正常男性结婚,子女都不患病C .Ⅲ—8与正常女性结婚,儿子都不患病D .该病在男性人群中的发病率高于女性人群4.以下为遗传系谱图,2号个体无甲病致病基因。
遗传分析中“正反交结果”的几点探究

遗传分析中“正反交结果”的几点探究在细胞质遗传的教学中,我发现在一些辅导材料上经常出现“正反交结果相同的是细胞核遗传,正反交结果不同的是细胞质遗传”,实际上,对这种简单化的表述如不能真正深入理解,就会出现错误或误解。
作者经过一番探究归纳,总结了若干理解要点和分析问题的注意事项,在此发表,以与大家共享。
1.正反交结果相同正反交结果相同,F1表现显性性状的是细胞核遗传的特点,特别强调的是它包含了性染色体上的同源部分的遗传。
2.正反交结果不相同2.1 与母本不一样,有明显的性别差异,是伴性遗传,它是指性染色体非同源部分的遗传。
伴性遗传是一种特殊的核遗传。
因为Y染色体上一般没有X染色体的等位基因,雄性个体不论是显性性状,还是隐性性状,只要有一个基因就表现出性状来。
而雌性个体,必须要有两个隐性基因才表现出隐性性状。
所以,基因位于X染色体上的伴性遗传,正反交时,后代的表现型也会不相同。
2.2 与母本一样,可能是细胞质遗传,也可能是母性影响,还可能是果皮或种皮的遗传。
2.2.1 细胞质遗传与果皮、种皮遗传的比较无论正交还是反交,子代性状总与母本相同,这是质遗传的一个显著特点。
这个特点很容易与被子植物的果皮、种皮遗传相混淆。
我们知道,果皮和种皮的基因型总是与母本相同的。
但要特别注意的是母本上结出的果实,其果皮和种皮并不是子代的,而是母本的一部分,它们分别是由母本的子房壁和珠被发育而成的。
种子的胚以及由胚发育而成的植株才是子代。
2.2.2 细胞质遗传与母性影响有些性状的遗传,看起来似乎是细胞质遗传,实际上仍然是细胞核遗传控制的。
比如锥实螺螺壳的螺旋方向有左旋和右旋两种类型,让右旋雌螺和左旋雄螺杂交,F1全部是右旋的,让左旋雌螺和右旋雄螺杂交,F1都是左旋的,从表面上看,锥实螺的螺旋方向的遗传属于细胞质遗传,但如果是细胞质遗传,那么F1自交,得到的F2自交得到的F3,乃至Fn,都应该与F1性状相同,但事实上,F3就开始出现了性状分离。
遗传学课程论文——细胞质遗传并非完全是母系遗传

生命科学学院2010级本科遗传学课程论文细胞质遗传并非完全是母系遗传姓名:孙颖雯班级:2010级生物科学(师范)2班学号:222010317011053指导教师:帅小蓉【摘要】细胞质遗传一般表现为具母系遗传的特征,但随着科学技术的日益发达,DNA分子标记技术的发展和应用,科学家们已发现在动物及植物中均存在有低频的线粒体DNA单亲父系遗传以及双亲遗传的现象。
对质体DNA遗传的研究表明:被子植物的质体DNA大多表现为母系遗传,而裸子植物的质体DNA则主要表现为父系遗传的方式,同时也发现存在其它的遗传规律[1]。
由此证实了细胞质基因的遗传并非全部都是母系遗传。
【关键词】细胞质遗传母系遗传线粒体DNA遗传质粒DNA遗传【引言】母系遗传是细胞质遗传的普遍形式,高等生物的细胞质遗传包括母系遗传、父系遗传和双亲遗传。
母系遗传是指:正交和反交的遗传表现不同,通常子代只表现母本性状,这种表型和母本表型一致而与父本表型无关的细胞质遗传又称母系遗传。
异配生殖的生物,其合子中细胞质主要来自雌配子,同时父本所形成的配子中,细胞质基因或丢失、或以某种方式被破坏、或失去活性,即只有母本的细胞质基因得到表达,父本的细胞质基因没有表达,因此,后代通常只表现母本的性状,这种细胞质遗传才是母系遗传[2]。
但是,如果过分强调雌配子中的细胞质多,而雄配子中细胞质少是母系遗传的主要原因,则以后就难以理解父系遗传和双亲遗传。
通常所说的细胞质遗传特点其实是指母系遗传的特点,如子代通常只表现母本性状,母系遗传和细胞质遗传是包容关系的概念。
因此,母系遗传一定是细胞质遗传,而细胞质遗传却不一定是母系遗传[3]。
例如花斑紫茉莉白色枝叶、绿色枝叶、花斑枝叶的遗传和链孢霉的慢性生长性状遗传,属于母系遗传;而酵母菌小菌落遗传属于细胞质遗传但不属于母系遗传。
【正文】细胞质遗传中,正、反交结果是不同的,因此,我们常常根据正、反交结果和后代的性状分离比来判断一种遗传是否是细胞质遗传。
浅析一些细胞质基因不表现母系遗传的现象

浅析一些细胞质基因不表现母系遗传的现象摘要:细胞质遗传的主要特点: 一是母系遗传; 二是后代的性状不会出现一定的分离比。
但是,由于一些特殊的遗传原因,有些细胞质基因并不表现母系遗传,而表现为父系遗传,或者两种遗传都表现。
关键词:细胞质基因母系遗传父系遗传一细胞质遗传并非都是母系遗传本世纪初, 在孟德尔定律被重新发现后的1909 年, 德国学者科伦斯( Corr ens) 和鲍尔(Baur) 分别在紫茉莉(Mir abilis j alapa ) 和天竺葵( Pelargonium zonale) 中发现叶色的遗传不符合孟德尔定律, 而表现为细胞质遗传现象。
这一发现是对孟德尔定律的挑战和补充。
研究表明, 大多数物种的细胞质性状表现为母系遗传的特征。
进而有些学者甚至某些遗传学教科书中也将细胞质遗传与母系遗传这两种现象混为一谈, 将这两个概念等同起来, 并认为细胞质遗传即为母系遗传。
细胞质遗传一般表现为母系遗传的特征。
然而据有关资料表明, 细胞质遗传并非都是母系遗传。
20 世纪80 年代以来, 随着DNA 分子生物学技术的发展, 将DNA分子标记应用于细胞质遗传研究, 从DNA 分子水平上研究细胞质遗传物质的变异, 使得人们对细胞质遗传现象有了更进一步的认识。
研究表明,在所有高等真核生物中, 线粒体DNA一般表现为母系遗传的特征, 包括人类、其他哺乳类动物、两栖类动物、鱼类及高等植物等。
但也发现, 老鼠、衣藻、被子植物的月见草、大麦和黑麦的属间杂种、甘蓝型油菜、北美红杉等生物体中线粒体DNA 是父系遗传的。
而对植物叶绿体DNA 的研究发现, 在被子植物中, 大多数植物表现为母系遗传特征, 而其中20%的物种存在着双亲遗传的现象, 紫花苜蓿、胡萝卜等植物表现为典型的父系遗传特征。
与被子植物相比, 大多数裸子植物的质体DNA 则表现为父系遗传特征。
二细胞质基因不表现母系遗传的原因细胞质遗传表现为多种形式的复杂性, 没有一种简单的机制去解释这种现象。
(整理)细胞质基因遗传与母系遗传现象

细胞质基因遗传与母系遗传现象摘要:细胞质遗传的主要特点是母系遗传和后代的性状不会出现一定的分离比。
因此,很多师生在遇到有关细胞质遗传的问题时,都会不加思索地认为:细胞质遗传一定是母系遗传。
然而据有关资料表明,细胞质遗传并非都是母系遗传。
但是随着20世纪80年代DNA分子生物技术、以及相继的电镜和抗DNA免疫电镜技术等的技术的发展发现细胞质遗传中也存在着低频的父系遗传与双亲遗传。
并且目前的大多数学者公认为现在没有任何一种简单的理论可以概括解释这种现象,细胞质遗传的内容也随着科学技术的发展在不断地扩充丰富着。
关键词:细胞质基因母系遗传父系遗传线粒体DNA 叶绿体DNA细胞质遗传一般表现为具母系遗传的特征。
随着DNA分子标记技术的发展和应用,人们已发现在动物及植物中均存在有低频的线粒体DNA单亲父系遗传及双亲遗传的现象。
对质体DNA遗传的研究表明,被子植物的质体DNA大多表现为母系遗传,而裸子植物的质体DNA则主要表现为父系遗传的方式,同时也发现存在其它的遗传规律。
一、母系遗传现象母体遗传可区分为迟延遗传和细胞质遗传二类。
所谓迟延遗传,例如在蚕卵的颜色、田螺壳的右旋、左旋等所见的现象。
虽然这些性状本身是由细胞核基因决定的,表现出明显的孟德尔式遗传,但是作为卵细胞质的性质却是在母体内形成的,因此子代的表型常常和母亲的基因型相同。
这样在F2中基因型虽以3∶1比例分离,但表型并不分离,与母体的性状完全一样,延迟一代后,在F3中表型才以3∶1的比例分离。
细胞质遗传一般也称为偏母遗传。
真核生物的绝大多数遗传物质存在于细胞核染色体中,但有少量遗传物质DNA存在于细胞质的细胞器中,如线粒体和叶绿体中,为小型环状DNA结构,一般含几千个碱基。
这种细胞质中的基因控制某些遗传性状,如水稻雄性不育症状。
在受精过程中,卵细胞中的细胞质基因完全遗传给子代,而精子由于本身缺乏细胞质,所以细胞质遗传表现为母系遗传。
二、细胞质遗现象(一) 常见的细胞质遗传(1). 高等植物叶绿体的遗传有几种高等植物有绿白斑植株,如紫茉莉、藏报春、加荆介等。
细胞质遗传

(二)胞质不育型: 胞质不育型: 指由细胞质内的基因所决定的雄性不育类 型. 在不育系的细胞质内有雄性不育基因S 在不育系的细胞质内有雄性不育基因 S , 但在细胞核内没有相应的雄性不育基因和 可育基因. 可育基因. 这种雄性不育性仅受细胞质的基因所控制, 这种雄性不育性仅受细胞质的基因所控制, 具有S基因的个体是雄性不育的,具有N 具有S基因的个体是雄性不育的,具有N基 因的个体是雄性可育.因此,有性生殖不 因的个体是雄性可育.因此,有性生殖不 能恢复胞质不育型的育性. 能恢复胞质不育型的育性.
这表明卡巴粒只通过细胞质来传递, 这表明卡巴粒只通过细胞质来传递 , 但 是它们的保持却要依赖于核中显性基因K 是它们的保持却要依赖于核中显性基因K 的存在. 的存在. 然而, 基因型为Kk 的放毒型并不稳定, Kk的放毒型并不稳定 然而 , 基因型为 Kk 的放毒型并不稳定 , 一旦经自体受精 基因型分离为KK,Kk 自体受精, KK,Kk和 一旦经自体受精,基因型分离为KK,Kk和 kk, kk个体细胞质里的卡巴粒不能保持 kk , kk 个体细胞质里的卡巴粒不能保持 和增值, 几次无性分裂后, 和增值 , 几次无性分裂后 , 就将因卡巴 粒的消失而成为敏感型 敏感型. 粒的消失而成为敏感型.
2)线粒体基因组的半自主性
线粒体基因组的相对独立性 mtDNA合成的调节与核 DNA合成的调节彼 合成的调节与核DNA ① mtDNA 合成的调节与核 DNA 合成的调节彼 此独立,可能存在多种复制形式,其中D 此独立,可能存在多种复制形式,其中D 环复制是线粒体特有的复制形式; 环复制是线粒体特有的复制形式; 线粒体基因组有自己独立的表达系统, ② 线粒体基因组有自己独立的表达系统 , 自己编码两种rRNA 22~ 24种 tRNA, rRNA, 自己编码两种 rRNA , 22 ~ 24 种 tRNA , 在 线粒体内合成mtDNA编码的蛋白质; mtDNA编码的蛋白质 线粒体内合成mtDNA编码的蛋白质;
细胞质遗传不等同母系遗传

的特 征 ,包 括 人 类、其 它 哺 乳类 动 物、
两 栖 动 物 、 鱼 类 及 高 等 植 物 等 。 随 着 D NA 分 子 标 记 技 术 在 细 胞 质 遗 传 研 究 中 的应 用 ,于 伦 斯 滕 ( G y l l e n s t e n)等
典 型的例证 , 是 1 9 8 9年舒 曼 ( S c h u ma n n) 和汉考克 ( Ha n c o c k)报 道 了第 1例 用R F L P分 析发现 紫花苜 蓿 ( Me d i c a g o s a t i v a)的质体 D NA 表现 为父 系遗传 的 特 征 ,而 胡 萝 卜属则 是在 被 子植 物 中 发 现质 体 D NA 由雄 配 子传 递 的第 2个 属 ( B o b l e n z等 ,1 9 9 0 o后来 又 在苜 蓿 ( Ma s o u d等 ,1 9 9 0) 矮 牵牛 ( P e t u n i a h y b r i d a) ( De r e p a s ,1 9 9 1)牛尾 草 与黑 麦 草 的属 间 杂 种 ( Ki a n g等 ,1 9 9 4) 和 猕
( 1 9 9 1 )报道在 老 鼠中线粒体 D NA 经雄
配 子 传 递 ,表 现 为低 频 的 父 系 遗 传 。 博 因顿 ( B o y n t o n)等 ( 1 9 8 7)发 现 在 衣 藻 ( C h l a my d o mo n a s r e i n h a r d t i i ) 中 线
e mml e) ( 1 9 8 4) 发现 月见 草属 的一 个杂
种 【 Oe n o t h e r a b e r t e r i a n a ×Oe . o d o r  ̄a) ,
23、细胞质遗传

皮为深蓝色的紫罗兰花粉传授于胚表皮为黄色的紫罗兰雌蕊上,所得后代的胚表
皮则呈黄色。紫罗兰胚表皮颜色遗传属于( )
A 显性遗传
B 伴性遗传
C 细胞质遗传 D 随机遗传
【答案】C
【解析】可根据细胞质遗传区别于细胞核遗传的特点:F1总是表现出母本性状, 所以紫罗兰胚表皮颜色遗传属于细胞质遗传。
延伸、拓展
具有可遗传的雄性不育性状的同种植 株群体叫做雄性不育系。
• 植物发生雄性不育的原因
(1)雄性是否可育,是由核基因和质基因 共同决定的。
核基因 :可育基因R对不育基因r是显性 质基因 :可育基因为N,不育基因为S
(2)核基因和质基因的关系: 质可育基因N可使花粉正常发育; 核可育基因R能够抑制细胞质不育基因
正确顺序是( B )
①N(rr)②S(rr)③N(RR)④S(Rr)
A ①②③④B ②①③④
C ③①②④D ④①②③
2 保持系的特点是( A ) A 既能使母本结实,又能使后代保持不育特性
B 质基因是不育基因,核基因为可育基因
C 质基因为可育基因,核基因为可育基因
D A、B、C都对
3
(D )
A 雄性不育系、保持系和杂交系配套使用
【答案】海拉细胞的氯霉素抗性遗传属于细胞质遗传
【解析】解答此题时首先应明确:小鼠细胞有无氯霉素抗性是一对相对性状。 将分别具有这一相对性状的两细胞进行核移植,形成的杂交细胞性状并不随核 的转移而改变。说明控制这一相对性状的基因位于细胞质中,氯霉素抗性遗传 属细胞质遗传。 【点评】解答此题的思维障碍是对杂交细胞的性状表现不明确。 排除障碍的方法是:分析杂交细胞的生长情况。可以在含氯霉素的培养基中分 裂生长,说明杂交细胞具有氯霉素抗性,与海拉细胞的性状相同,受细胞质基 因控制;若不能正常生长,说明杂交细胞无氯霉素抗性,与核源细胞性状相同 ,受核基因控制。
细胞质遗传

染色体外遗传染色体以外的遗传因子所表现的遗传现象。
在真核生物中常称为细胞质遗传,也称为核外遗传、非染色体遗传、非孟德尔式遗传或母体遗传。
植物正反杂交子代的某些性状都相同于母本,这是最早发现的染色体外遗传现象。
这里性状相同于母本的原因是由于控制这些性状的遗传因子是在细胞质中,而高等植物的合子的细胞质又几乎全部来自雌性配子。
母体影响在现象上相似于染色体外遗传或母体遗传,即正反杂交子代的某些性状相同于雌性亲本,不过这是由于核基因的产物积累在卵细胞的细胞质中造成的,因此不属于染色体外遗传的范畴。
例如在地中海粉螟中,基因A使幼虫含色素,基因a使幼虫不含色素。
正反杂交aa♀×Aa♂和Aa♀×aa♂的子代的基因型都是Aa和aa 两种。
基因型Aa的幼虫含有色素。
但是基因型aa的幼虫则因正反交而不同,由前一杂交得来的aa幼虫不含色素而由后一杂交得来的则含有色素。
原因是后者的卵来自Aa雌性亲本,由于卵母细胞中A基因的存在,因而经减数分裂产生的卵不论基因型是A或是a,它们的细胞质中都含有A基因的产物,即色素的前体以及由前体所合成的色素。
简史1900年德国植物学家兼遗传学家C.E.科伦斯重新发现孟德尔定律后,又于1909年报道了不符合于孟德尔定律的遗传现象。
他发现在紫茉莉(Mirabilis jalapa)中黄绿色叶♀×绿色叶♂的杂交子代都是黄绿色叶,绿色叶♀×黄绿色叶♂的杂交子代都是绿色叶。
德国学者E.鲍尔在同一年中报道了天竺葵(Pelar.onium zonale)中的类似现象,并且认为这是由于叶绿体的独立自主性所造成的。
1924 年德国学者 R.von 韦特施泰因报道了葫芦藓(Furnaria)中的母体遗传现象,并提出细胞质基因组这一名词,用来概括细胞质中的全部遗传物质。
P.米凯利斯从 20年代开始对柳叶菜属(Epilobium)植物连续进行了二、三十年的研究,分析了若干种性状的母体遗传现象。
质遗传

并非所有细胞质基因都是母系遗传内容摘要大家都知道真核生物的遗传基因包括质基因和核基因,存在于细胞核内的基因叫做核基因,存在与细胞质内的基因叫做质基因,细胞质基因又都存在于线粒体和质体中,所以要讨论质基因只要讨论线粒体基因和质体基因就行了。
细胞质基因大多数都表现为母系遗传,但是有些质基因并不表现为母系遗传,却表现为父系遗传和双亲遗传。
而这种遗传机制还没有完全研究清楚。
关键词质基因父系遗传双亲遗传母系遗传正文细胞质遗传一般表现为具母系遗传的特征。
随着DNA分子标记技术的发展和应用,人们已发现在动物及植物中均存在有低频的线粒体DNA单亲父系遗传及双亲遗传的现象,而质体中也存在这种现象。
这是对细胞质母系遗传的挑战。
1. 线粒体基因不表现母系遗传的例证随着DNA分子标记技术在细胞质遗传研究中的应用,于伦斯滕(Gyllensten)等(1991)报道在老鼠中线粒体DNA经雄配子传递,表现为低频的父系遗传。
博因顿(Boynton)等(1987)发现在衣藻(Ch lam y d om onas rein2ha rd tii)中线粒体DNA表现为父系遗传。
在被子植物中,利用线粒体DNA的限制性片段分析方法,Brennicke和施韦姆勒(Schwemm le)(1984)发现月见草属的一个杂种(O enotheraberteriana×O e.od ora ta),苏莱曼(So lim an)等(1987)报道在大麦(H ord eum v u lga re)和黑麦(S eca le cerea le)的属间杂种,K iang等(1994)观察到在牛尾草(Festuca p ratensis)黑麦草(L olium p erenne)的属间杂种中均表现为线粒体DNA的父系遗传。
埃里克森(Erickson)和肯布尔(Kemble)(1990)在双子叶的甘蓝型油菜(B rassica nap us)中同样发现了高等植物中存在的线粒体DNA父系遗传现象,在F1子代中有10%的植株的线粒体DNA来自于父本。
浅析细胞质遗传和母性效应

浅析细胞质遗传和母性效应作者:后治中来源:《读写算》2011年第16期【摘要】一般来说,细胞质遗传和母性效应都与母本有关,都是真核生物的两种遗传方式,不同的是,细胞质遗传具有两个明显的主要特点:一是母系遗传,二是后代无一定分离比,不遵循孟德尔遗传定律;而母性效应虽与母本有关,但本质上属于细胞核遗传,遵循孟德尔遗传定律【关键词】细胞质遗传;母性效应1 细胞质遗传紫茉莉的枝条一般都是绿色的。
但是这种植物有多种变异类型,如出现花斑植株。
在花斑植株上有时还会生有3种不同的枝条——绿色的、白色的和花斑状的。
紫茉莉枝叶的这种性状是怎样向后代传递的呢?用紫茉莉不同枝条上的花相互授粉,得到的结果如下表:从紫茉莉枝叶颜色遗传的杂交结果,我们知道,F1植株的颜色,完全取决于种子产生于哪一种枝条,而与花粉来自哪一种枝条无关,也就是说,F1的性状,完全是由母本决定的。
像这样,具有相对性状的亲本杂交,F1总是表现出母本性状的遗传现象,叫做母系遗传。
研究表明:卵细胞中含有大量的细胞质,而精子中只含极少量的细胞质。
这就是说,受精卵中的细胞质,几乎全部来自卵细胞。
这样,受细胞质内遗传物质控制的性状实际上是由卵细胞传给了子代,因此,子代总是表现出母本性状。
这是细胞质遗传不同于细胞核遗传的一个主要特点。
这种遗传现象可简单地表示为:即细胞质遗传中,正、反交结果是不同的,因此,我们常常根据正、反交结果和后代的性状分离比来判断一种遗传是否是细胞质遗传。
细胞质遗传的另一个主要特点是:两个亲本杂交,后代的性状不会像细胞核遗传那样出现一定的分离比。
之所以会出现这一现象,是因为生殖细胞在进行减数分裂时,细胞质中的遗传物质不能像核内的遗传物质那样进行有规律的分离,而是随机的、不均等的分配到子细胞中去。
例如,如果让紫茉莉花斑枝条上的雌蕊接收绿色枝条上的花粉,由于花斑枝条细胞质中的遗传物质不均等的分配,F1呈现出来的3种表现型不会出现一定的分离比。
也就是说,孟德尔的遗传定律的实验是根据细胞核内的染色体上的基因来进行的,并未选择细胞质内的遗传物质;母系遗传的遗传物质存在于细胞质,细胞质内的染色体复制、转录等方式和核内染色体有着很大的区别,甚至密码子都不一样。
细胞质遗传不满是母系遗传

生命科学学院生物科学遗传学课程论文生科6班2220刘秦杰细胞质遗传并非都是母系遗传摘要:自从德国植物学家兼遗传学家Correns(1908)第一次宣布了他的紫茉莉叶色杂交实验结果以后,大量的实验证明细胞质基因是通过母体遗传给子代的。
细胞质遗传确实是母系遗传的那个观点为公共所同意,乃至很多教科书也是如此提到。
可是,事实真是如此吗?答案是不是定的:细胞质遗传指由细胞质内的基因操纵的遗传,是与细胞核遗传相对的。
而母系遗传指子代性状由母本决定的遗传,与它相对的是双亲遗传和父系遗传。
Russell等依照5种代表植物配子融合时精细胞质传递的状况,将质体遗传区分为单亲母系传递型和双亲传递型。
哈格曼(Hagemann, 1992) 将被子植物的花粉传递分为3 类:第1 类为月见草属型,表现为明显的母系传递特性;第2 类为天竺葵属型,表现为双亲传递特性;第3 类为苜蓿属型,表现为明显的父系传递特性。
即细胞质遗传除母系遗传外,还有双亲遗传和父系遗传。
关键词:细胞质遗传母系遗传双亲遗传父系遗传例证机制一、母系遗传。
第一,咱们应该先明确母系遗传的概念。
母系遗传(matrilinear inheritance)是指只受母本遗传物质操纵,子代只表现母本性状的现象。
在那个地址就有一个问题了,凡是F1代表现出母本形状的都是母系遗传吗?以下例证能够回答那个问题。
例证一、豌豆豆荚颜色的表现。
豌豆豆荚的颜色有绿色和黄色之分,将绿色豆荚豌豆的花粉授给黄色豆荚的豌豆,该植株所结出的豌豆豆荚的颜色均表现为黄色,与母本性状一致;反之,将黄色豆荚豌豆的花粉授给绿色豆荚的豌豆,该植株所结出的豌豆豆荚的颜色均表现为绿色,亦与母本性状一致。
可是豌豆豆荚颜色的遗传却不是母系遗传:豌豆的豆荚事实上是果皮,它是由子房壁发育而来的,而子房壁是母本的一部份,固然其基因型及表现型应与母本的一致。
例证二、雄蜂体色的表现。
褐色雄蜂与黑色蜂王交配,子代中所有雄蜂都是黑色的,与母本性状一致。
细胞质遗传并非都是母系遗传

细胞质遗传并非都是母系遗传
田志宏
【期刊名称】《生物学通报》
【年(卷),期】1999(034)001
【摘要】细胞质遗传一般表现为具母系遗传的特征。
随着DNA分子标记技术的发展和应用,人们已发现在动物及植物中均存在有低频的线粒体DNA单亲父系遗传及双亲遗传的现象,对质体DNA遗传的研究表明,被子植物的质体DNA大多表现为母系遗传。
而裸子植物的质体DNA则主要表现为父系遗传的方式,同时也出现存在其它的遗传规律。
【总页数】2页(P14-15)
【作者】田志宏
【作者单位】华中农业大学作物遗传改良国家重点实验室
【正文语种】中文
【中图分类】Q343.3
【相关文献】
1.与母本性状一致都是细胞质遗传吗 [J], 毕景花;刘鸿
2.耳鸣,并非都是肾虚惹的祸 [J], 张乾
3.并非所有"阴道流血"都是先兆流产 [J], 陈晓琴
4.并非所有的午睡都是健康的 [J], 吴施楠
5.与母本性状一致的现象都是细胞质遗传吗 [J], 王小飞
因版权原因,仅展示原文概要,查看原文内容请购买。
细胞质遗传

细胞质遗传并非都为母系遗传一.细胞质遗传并非都是母系遗传细胞质遗传现象的发现最早可追溯到1909 年,德国学者科伦斯(Carl Correns)和鲍尔(Baur)分别在紫茉莉和天竺葵中发现叶色的遗传不符合孟德尔定律,而表现为细胞质遗传现象。
后来的研究表明,大多数物种的细胞质性状表现为母系遗传的特征,因而有些学者甚至某些遗传学教科书中也将细胞质遗传与母系遗传这两种现象混为一谈,将这两个概念等同起来,并认为细胞质遗传即为母系遗传。
20 世纪80 年代以来,随着分子生物学技术的发展,将DNA 分子标记应用于细胞质遗传研究,从DNA 分子水平上研究细胞质遗传物质的变异,使得人们对细胞质遗传现象有了更进一步的认识。
据研究表明,在所有高等真核生物中,线粒体DNA 一般表现为母系遗传的特征,包括人类、其他哺乳类动物、两栖动物、鱼类及高等植物等。
但也发现,老鼠、衣藻、被子植物月见草属的一个杂种、大麦和黑麦的属间杂种、甘蓝型油菜、北美红杉等生物体中线粒体DNA 是父系遗传的。
在被子植物中,对近60 个物种的质体DNA 的遗传研究,发现大多数表现为母系遗传特征,而其中20%的物种中存在着双亲遗传的现象,紫花苜蓿、胡萝卜等植物表现为典型的父系遗传特征。
与被子植物相比,大多数裸子植物的质体DNA 则表现为父系遗传特征。
可见,细胞质遗传表现为多种形式的复杂性,没有一种简单的机制去解释这种现象。
母系遗传是细胞质遗传的主要特征,而不能代表细胞质遗传的全部内容。
随着分子生物学技术的发展和应用,为人们对细胞质遗传规律的研究和认识提供了强有力的手段,科学家们已揭示出了生物细胞质DNA 遗传的新规律和新现象,在细胞质遗传方面表现为单亲的母系遗传、父系遗传及双亲遗传多种形式,大大丰富和逐步丰富了细胞质遗传研究的内容。
由于细胞质基因与细胞核基因在细胞中分布位置不同,在细胞分裂时分配规律不同,所以在传种接代时细胞质基因表现出不同于细胞核基因的遗传特点:(1)母系遗传:具有相对性状的亲本杂交时,F1总是表现出母本性状的遗传现象。
遗传病遗传方式的判断

遗传病遗传方式的判断太原外国语学校 刘保东 lbdong277@一,遗传病的类型及其特点;1,细胞质遗传:特点是:母系遗传。
(母患病子女都患病,与父无关) 2,Y 染色体遗传:特点是:父患病儿子都患病,女儿都不患病。
(传子不传女,子孙无穷尽) 3,常染色体显性遗传:特点是:①男女患病的概率相等; ②连续遗传;③家族中发病率高。
4,常染色体隐性遗传:特点是:①男女患病的概率相等; ②隔代遗传;③家族中发病率低。
5,X 染色体显性遗传:特点是:①女性患者多于男性患者; ②连续交叉遗传; ③家族中发病率高;④男性患病其母亲和女儿都患病; ⑤女性正常其父亲和儿子都正常。
6,X 染色体隐性遗传:特点是:①男性患者多于女性; ②隔代交叉遗传; ③家族中发病率低;④男性正常其母亲和女儿都正常; ⑤女性患病其父亲和儿子都患病。
二,遗传方式判断方法;首先;判断细胞质遗传和Y 染色体遗传。
观察遗传系谱图,如果符合细胞质遗传和Y 染色体遗传的特点,则优先判断为细胞质遗传或Y 染色体遗传。
例如:图一可能为Y 染色体遗传、常染色体显性遗传、常染色体隐性遗传、X 染色体隐性遗传,最可能是Y 染色体遗传;图二可能为为常染色体显性遗传、常染色体隐性遗传、细胞质遗传,最可能是细胞质遗传。
如果不符合细胞质遗传和Y染色体遗传的特点,则可以确定不是细胞质遗传或Y 染色体遗传。
(即:如果父亲患病儿子有不病、女儿有患病的,或儿子患病而父亲不病,则一定不是Y染色体遗传。
如果母亲患病子女有不病的,则一定不是细胞质遗传。
)其次;确定不是细胞质遗传或Y染色体遗传后,判断其他四种方式的遗传。
1,大致判断;(基本能知道遗传的类型,但不一定正确)观察遗传系谱图:(1)如果某种遗传病在家族中发病率高、且连续遗传,可以大致判断为显性遗传。
(2)如果某种遗传病在家族中发病率低,可以大致判断为隐性遗传。
(3)如果某种遗传病男女患病的概率相等,可以大致判断为常染色体遗传。
高中生物中几个问题的辨析

高中生物中几个问题的辨析摘要:随着分子生物学技术的发展和应用,人们对细胞质遗传规律、胰高血糖素、淀粉酶、细菌的研究和认识提供了强有力的手段,大大丰富和逐步完善了研究的内容。
关键词:生物问题辨析一、无籽西瓜发育形成时生长素从何而来我们知道,果实是由子房发育而来的,在子房发育成为果实的过程中,需要一定量的生长素,一般情况下,生长素是由胚珠发育形成的幼嫩种子提供的,没有种子的无籽西瓜,生长素从何而来呢?一般来说,生长素在植物体内的合成部位是叶原基、嫩叶和发育中的种子。
在这些部位,存在着与生长素的合成有关的酶体系,在多种酶的催化作用下,植物体内的色氨酸经过氨基转换、脱羧作用和两个氧化步骤,最终变成生长素(吲哚乙酸)。
在二倍体西瓜的花粉中,除含有少量的生长素外,同样也含有使色氨酸转变成生长素的酶体系。
在无籽西瓜栽培中,三倍体植株开花时必须授以二倍体西瓜正常的花粉,当二倍体花粉萌发时,形成的花粉管伸入到三倍体植株的子房内并将自身合成生长素的酶体系转移到其中,在子房内仍能合成大量的生长素,从而发育形成无籽西瓜。
二、细胞质遗传并非都是母系遗传细胞质遗传的主要特点:一是母系遗传;二是后代的性状不会出现一定的分离比。
因此,很多学生在遇到有关细胞质遗传的问题时,都会不加思索地认为:细胞质遗传一定是母系遗传。
然而据有关资料表明,细胞质遗传并非都是母系遗传。
细胞质遗传一般表现为母系遗传的特征。
20世纪80年代以来,随着dna分子生物学技术的发展,将dna分子标记应用于细胞质遗传研究,从dna分子水平上研究细胞质遗传物质的变异,使得人们对细胞质遗传现象有了更进一步的认识。
研究表明,在所有高等真核生物中,线粒体dna一般表现为母系遗传的特征,包括人类、其他哺乳类动物、两栖类动物、鱼类及高等植物等。
但也发现,老鼠、衣藻、被子植物的月见草、大麦和黑麦的属间杂种、甘蓝型油菜、北美红杉等生物体中线粒体dna是父系遗传的。
而对植物叶绿体dna 的研究发现,在被子植物中,大多数植物表现为母系遗传特征,而其中20%的物种存在着双亲遗传的现象,紫花苜蓿、胡萝卜等植物表现为典型的父系遗传特征。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
细胞质遗传并非都是母系遗传
细胞质遗传一般表现为母系遗传的特征。
20世纪80年代以来,随着DNA分子生物学技术的发展,将DNA分子标记应用于细胞质遗传研究,从DNA分子水平上研究细胞质遗传物质的变异,使得人们对细胞质遗传现象有了更进一步的认识。
据研究表明,在所有高等真核生物中,线粒体DNA一般表现为母系遗传的特征,包括人类、其他哺乳类动物、两栖动物、鱼类及高等植物等。
但也发现,老鼠、衣藻、被子植物中的月见草、大麦和黑麦的属间杂种、甘蓝型油菜、北美红杉等生物体中线粒体DNA是父系遗传的。
而对植物叶绿体DNA的研究发现,在被子植物中,大多数植物表现为母系遗传特征,而其中20%的物种中存在着双亲遗传的现象,紫花苜蓿、胡萝卜等植物表现为典型的父系遗传特征。
与被子植物相比,大多数裸子植物的质体DNA则表现为父系遗传特征。
细胞质遗传表现为多种形式的复杂性,没有一种简单的机制去解释这种现象。
母系遗传是细胞质遗传的主要特征,而不能代表细胞质遗传的全部内容。
随着分子生物学技术的发展和应用,为人们对细胞质遗传规律的研究和认识提供了强有力的手段,科学家们己揭示出了生物细胞质DNA遗传的新规律和新现象,在细胞质遗传方面表现为单亲的母系遗传,父系遗传及双亲遗传多种形式,大大丰富和逐步完善了细胞质遗传研究的内容。