细胞质遗传
细胞质遗传

细胞质遗传
细胞质遗传是指通过细胞质中遗传物质的传递,影响后代性状的过程。
虽然细胞核内的核酸控制了大部分遗传信息,但细胞质中的线粒体和叶绿体也扮演着重要的角色。
细胞质遗传的机制
细胞质遗传主要通过线粒体和叶绿体中的DNA来实现。
线粒体是细胞内的能量中心,负责细胞内的能量转化,而叶绿体是光合作用的场所。
这两种细胞器都含有自己的DNA,通过雌性细胞质的传递来影响后代性状。
线粒体和叶绿体的DNA是环状的,相对于更为复杂的细胞核DNA而言,它们相对较小且编码的基因数量有限。
然而,这些基因对于细胞的正常功能至关重要。
细胞质遗传的影响
细胞质遗传可以导致一些遗传病的传递。
如果母亲携带有突变的线粒体DNA,那么她的子代继承这些突变的可能性会很高。
这种现象被称为“母系遗传”。
另外,细胞质遗传也被发现与一些植物的性状传递有关。
例如,某些植物的叶绿体DNA编码了一些影响光合作用效率的基因,这就导致了在杂交后代中出现了不同的叶形和叶色。
细胞质遗传的意义
了解细胞质遗传对于生物科学的发展至关重要。
通过研究细胞质遗传,人们可以更深入地了解遗传信息的传递机制,从而为疾病的治疗和预防提供更多可能性。
另外,细胞质遗传的研究也为农业和植物育种领域提供了一些新的思路。
通过深入研究植物叶绿体和线粒体的遗传机制,人们可以更好地改良作物,提高农作物的产量和品质。
总的来说,细胞质遗传虽然在遗传学中占据较小的一部分,但是其独特的传递方式和影响机制使其在生物科学领域中扮演着不可或缺的角色。
不断地深入研究和探索细胞质遗传,将有助于揭示生命的奥秘并推动人类社会的发展进步。
医学遗传学第八章细胞质遗传

细胞质遗传学的研究有助于发现新的药物靶点,为药物的研发提供新的思路和方法。
个体化治疗
通过对细胞质基因的研究,可以了解不同个体的基因变异情况,为个体化治疗提供依据,提高治疗效果和减少副作用。
人类起源与进化
细胞质遗传学的研究有助于深入了解人类的起源和进化历程,揭示人类在生物多样性中的地位和演化过程。
细胞质基因的表达受到多种因素的影响,包括基因的启动子、转录因子和表观遗传修饰等。
细胞质基因的表达水平在不同组织中存在差异,与细胞的能量需求和代谢活性密切相关。
细胞质基因组的复制与表达
1
2
3
细胞质基因突变是指细胞质基因组中的碱基序列发生改变,可能导致线粒体功能异常或疾病的发生。
细胞质基因突变通常由DNA复制过程中的错误或损伤引起,也可能与环境因素有关。
详细描述
线粒体疾病
VS
肌营养不良症是一类由基因突变引起的肌肉疾病,主要表现为肌肉萎缩和肌力减弱。
详细描述
肌营养不良症是一类遗传性疾病,其特点是肌肉逐渐萎缩和失去功能。常见的肌营养不良症包括杜氏肌营养不良症(DMD)和贝氏肌营养不良症(BMD)等。这些疾病通常在儿童期发病,并随着年龄的增长逐渐加重,影响患者的行走、站立、呼吸等基本功能。
05
细胞质遗传学的应用前景
疾病诊断
细胞质遗传学在疾病诊断中具有重要作用,通过对细胞质基因突变的研究,可以检测出一些与疾病相关的基因变异,为疾病的早期诊断提供依据。
预防策略
基于细胞质遗传学的知识,可以制定出针对特定疾病的预防策略,如对高危人群进行筛查和干预,以降低疾病的发生风险。
疾病诊断与预防
药物研发与治疗
深入探索细胞质遗传的机制
尽管我们对细胞质遗传有一定的了解,但是其具体机制仍有许多未知之处,需要进一步深入研究。
遗传学细胞质遗传

雄性不育得遗传决定类型
核质互作雄性不育型
由细胞质遗传因子和核基因 互相作用共同控制花粉育性 得类型
大多数正常品种与之杂交后 代仍为雄性不育
少数携带特殊恢复基因得品
种与之杂交后代正常可育
N
雄性不育得遗传决定类型
细胞质雄性不育型
决定花粉败育得基因在细胞质中 任何正常品种与其杂交,F1均为雄性不育 只能被保持而不能被恢复
胞质不育基因得载体
线粒体基因组(mtDNA)就是雄性不育基因得载体
不育系线粒体得亚显微结构和 mt DNA 与正常植株(如保持 系)有明显得差异
叶绿体基因组(cpDNA)就是雄性不育基因得载体
不育系和保持系之间在叶绿体超微结构和叶cpDNA上存在 明显不同
质核互作雄性不育性得发生机理
质核互补控制假说
终止子 亮氨酸 leu 异亮氨酸 ile 精氨酸 arg
例外情况
色氨酸 trp 苏氨酸 thr 甲硫氨酸 met
终止子
发现得生物
人和酵母得线粒体 酵母得线粒体 人得线粒体 人得线粒体
线粒体基因组
线粒体就是半自主性得细胞器
mtDNA虽能自主复制,但需要核基因组为其编码 DNA聚合酶 线粒体虽有自己得核糖体、tRNA,并能在线粒体内 翻译mtDNA转录得mRNA,但其核糖体蛋白质由核基 因编码 线粒体膜蛋白大多从核基因组中转录,在细胞质中合 成后再转运进入线粒体
叶绿体基因组
叶绿体就是半自主性得细胞器
cpDNA虽能自主复制,编码其核糖体RNA及部分 tRNA,但不能编码自身得DNA复制、转录和翻译过 程所需得全部蛋白质因子和酶 不能编码类囊体所有得结构蛋白和酶系,膜脂代谢所 需得酶系
非细胞质组分得遗传因子
遗传学第8章细胞质遗传

酸为核DNA所编码,其余十几种氨基酸为ctDNA所编码。
第四节 线粒体遗传
一、线粒体遗传的表现 二、线粒体遗传的分子基础
一、线粒体遗传的表现
举例:红色面包霉缓慢生长突变型的细胞质遗传 1。原子囊果相当于一个卵细胞,它包括细胞质和细胞核两部分;分生孢子在 受精时只提供细胞核,相当于精子。 2。红色面包霉缓慢生长突变型,在正常繁殖条件下能很稳定地遗传下去。 将突变型的原子囊果和野生型的分生孢子受精结合时,所有子代都是突变型; 反之,所有子代都是野生型的。 3。缓慢生长的特性被原子囊果携带,就能传递给所有子代;如果由分生孢子 携带,就不能传递给子代。 4。生化分析:突变型不含细胞色素氧化酶,这种酶是由线粒体控制合成的。
㈡、线粒体基因组的构成: ➢1981年Kanderson最早测出人的mt DNA全序列为16569bp。 ➢人、鼠、牛的mtDAN全序列均被测出:
2个rRNA基因;22个tRNA 基因;13个蛋白质基因等;
人、鼠和牛的mtDNA分别有 16569bp、16275bp和16338bp。
者的差异较小。
㈡、叶绿体基因组的构成: 1、低等植物的叶绿体基因组:
⑴. ct DNA仅能编码叶绿体本身结构和组成的一部分物质; 如各种RNA、核糖体(70S)蛋白质、光合作用膜蛋白 和RuBp羧化酶8个大亚基。
⑵. 特性:与抗药性、温度敏感性和某些营养缺陷有关。
2. 高等植物的叶绿体基因组:
2. cytoplasmic inheritance :由细胞质基因所决定的遗传 现象和遗传规律,也称为非孟德尔遗传,核外遗传。
染色体基因组
基因(核DNA)
遗传学细胞质遗传

㈡、草履虫放毒型的遗传:
1. 结构: 草履虫(Paramecium aurelia)是一种常见
的原生动物,种类很多。 大核(1个),是多倍体,主要负责营养; 小核(1~2个),是二倍体、主要负责遗传。
41
④.半自主性的细胞器: 线粒体内100多种蛋白质中,约有10种是线粒体本身
合成的,包括细胞色素氧化酶亚基、4种ATP酶亚基和1种 细胞色素b亚基。
∴线粒体的蛋白是由线粒体本身和核基因共同编码的, 是一种半自主性的细胞器。
42
第五节 共生体和质粒决定的染色体 外遗传
一、共生体的遗传:
㈠、共生体(symbionts): 不是细胞生存所必需的组成部分,仅以某种共生的
(二)持久的母性影响
例: 椎实螺外壳 的旋转方向受母亲基 因型控制,终生不变。 它受一对等位基因控 制,右旋(D)对左旋(d) 为显性。
椎实螺正反交,F1旋转方向都与各自母本相似,即右 旋或左旋,F2却都为右旋,F3才出现右旋和左旋的分 离。
P ♀DD × dd♂
右旋 左旋
♀dd × DD♂
左旋 右旋
36
线粒体数目及mt DNA大小:
生物种类 酵母
几种生物的 mt DNA
每细胞中 线粒体数
mt DNA 大小 (kb)
22
84
鼠(L 细胞)
500
16.2
人(Hela 细胞) 800
16.6
mt DNA 与 核 DNA 比值
0.18
0.002 0.01
37
㈡、线粒体基因组的构成:
1981年Kanderson最早测出人的mt DNA全序列为16569 bp。 人、鼠、牛的mtDAN全序列中测出:
遗传学细胞质遗传

第四节 线粒体遗传
一、线粒体遗传旳体现
(一)红色面包霉生长缓慢突变型旳遗传
突变型原子囊果 × 野生型分生孢子 (缓慢生长型) (生长速度正常)
↓ 突变型
野生原子囊果 × 突变型分生孢子 ↓
野生型
成果分析:
缓慢生长突变型在幼嫩培养阶段不含细胞 色素氧化酶,且线粒体构造不正常 细胞色素氧化酶旳产生与线粒体直接有关
二、叶绿体基因组
叶绿体DNA(chloroplast DNA,ctDNA) ctDNA与细菌旳DNA相同,闭合双链旳环状 构造,裸露旳DNA ; 每一种叶绿体内ctNDA分子数目不拟定,高等 植物中约30~60拷贝。 ctDNA能自主复制、转录和翻译 叶绿体性状同步受核DNA和ctDNA控制,在遗传 上有半自主性
的枝条
的表现
白色
白色
绿色
白色
花斑
白色
绿色
绿色
绿色
花斑
白色
花斑
绿色
花斑
花斑
杂种植株所体现旳性状完全由母本枝条所决定,
与提供花粉旳父本枝条无关。可知控制紫茉莉花
斑性状旳遗传物质是经过母本传递旳。
原因:
正常质体
绿色
细胞正常 随机分配 细 白色质体
白色
质体和白 色质体
胞 正常质体和白
色质体
花斑
花斑枝条:
基因组
附加体基因组(细菌质粒)
一、细胞质遗传概念
二、 细胞质遗传旳特点
1.正交和反交旳体现不同 2.杂交后裔旳表型分离不符合Mendel百分比; 3.经过连续回交,能把母体旳核基因全部置换掉, 母体旳细胞质基因及其控制旳性状不消失; 4.由附加体或共生体决定旳性状,其体现类似于病 毒旳转导和感染; 5.只能经过卵细胞传递给后裔。
细胞质遗传

三系二区配套育种
六、母性影响
母性影响又称母性效应,母系影响, 是指子代的表型不受自身基因型控制, 受母亲基因型影响,其表型同目前相 同的现象。 其分为二种类型: 一种是短暂的,只影响子代个体的幼 龄期; 另一种是持久的,影响子代个体终生。
母性影响所表现的遗传现象和细胞质遗 传十分相似,但它在本质上时另一类遗 传现象。 它属于细胞质遗传的范畴。而且,母性 影响经过几代后,影响消失,则性状遗 传也随之改变。
但是一般由核基因所决定的性状,正交 和反交的遗传表现是完全一致的。
2.杂交后代子代不出现孟德尔式的分离比。
3. 通过连续的回交能把母本的核基因全部臵 换掉,但母本的细胞质基因及其控制的性状 仍不消失。 4.具有细胞质异质性与细胞质分裂和重组。
同一细胞内含有不同基因型细胞器(如叶绿 体或线粒体)的现象称为细胞质异质性。
这类细胞在分裂过程中,细胞器无规则地随 机分离,子代细胞获得不同基因型细胞器或 获得的比例不同,导致细胞或个体间的表型 差异,这种分配过程称为细胞质分裂和重组。
二、细胞器基因组的遗传
1.叶绿体的遗传
紫茉莉花斑叶色的遗传
母本枝条的类型 父本枝条的类型 白 白 绿 花斑 白 绿 绿 花斑 白 花斑 绿 花斑 子代的类型 白
有色 褐眼
幼虫: 有色 成虫: 褐眼
幼虫:有色 成虫:红眼
2.持久的母性影响
在椎实螺发育中,其受精卵是螺 旋卵裂形式,它的整个发育决定于第 一次卵裂,而第一次卵裂决定于卵母 细胞的来源。所以椎实螺发育归根决 定于母体的基因型和这种基因型所影 响的卵母细胞。(见下图)
椎实螺外壳右旋(D)对左旋(d)显性。
第42讲细胞质遗传

第42讲细胞质遗传一、考点内容全解(一)本讲考点是什么1.细胞质遗传(1)细胞质遗传的物质基础:叶绿体、线粒体、质粒等细胞质结构中的DNA。
(2)细胞质遗传的主要特点是:母系遗传;后代不出现一定的分离比。
(3)细胞核遗传和细胞质遗传是相互依存、相互制约,不可分割的,共同控制生物的性状。
2.核质互作杂交育种—三系配套一区:培育不育系S(rr)和保持系N(rr)S(rr)×N(rr)S(rr)N(rr)二区:培育杂交种S(Rr)和恢复系N(RR)S(rr)×N(RR)S(Rr)N(RR)一、细胞质遗传的概念:细胞质遗传:由胞质遗传物质引起的遗传现象(又称非染色体遗传、非孟德尔遗传、染色体外遗传、核外遗传、母性遗传)。
细胞质基因组:所有细胞器和细胞质颗粒中遗传物质的统称。
二、细胞质遗传的特点:(1)母系遗传:具有相对性状的亲本杂交,F1总是表现出母本性状的遗传现象。
(2)分离比不确定:两个亲本杂交,若出现性状分离,不会像细胞核遗传那样出现一定的分离比。
3. 形成特点的原因:(1)受精卵中的细胞质几乎全部来自卵细胞。
(2)生殖细胞在进行减数分裂时,细胞质中的遗传物质不能像核内的遗传物质那样进行有规律的分离,而是随机地、不均等地分配到子细胞中去。
细胞质遗传的物质基础线粒体和叶绿体中存在DNA,都能够进行自我复制,并通过转录和翻译控制某些蛋白质的合成,控制生物的某些性状。
细胞质遗传在实践中的应用:细胞质遗传主要应用在作物育种上。
(一)杂种优势1. 概念:在农业生产上,杂交种表现出生长整齐、植株健壮、产量高、抗虫抗病能力强等特点。
2. 弊端:杂种优势往往只表现在两个品种杂交后的第一代上。
杂种第一代所结的种子再种下去,长出来的杂种第二代(F2)就会出现生长不齐、产量下降等现象。
因此要保持作物的杂种优势,就必须年年配制第一代杂交种。
提问:若利用矮秆(A)植株和抗病(B)植株,培育既矮秆又抗病的植株,其杂交的遗传图解如何?杂交的后代性状分离的情况如何?(二)雄性不育植株1. 概念:雄蕊发育不正常,不能产生可育的花粉,但是雌蕊正常可育,可以接受其他植株的花粉而产生种子,这种现象在遗传学上称为雄性不育。
第10章细胞质遗传 - 副本

细胞质 基因组
非细胞器 基因组
细胞质遗传:又称核外遗传,指核内染色体以外的 遗传物质所决定的遗传现象和规律。
1.1 细胞质遗传的特点
卵细胞:含细胞核,细胞质及 各种细胞器 为子代提供 核基因及全部或大部分细胞 质基因; 精子:含细胞核,没有或极少 细胞质和细胞器 仅为子 代提供核基因。 胞质基因决定的性状,遗传信 息只能通过卵细胞传给子代。
S rr S RR S Rr
可育
S Rr S rr
可育
可育
不育
S Rr
×
不育
N Rr
可育
N RR S Rr
可育
可育
N rr S rr
可育
可育
不育
生产上的应用——三系配套
不育系 (A) —S(r/r) 保持系 (B) —N(r/r) 恢复系 (R)—N(R/R)或S(R/R)
5.2 质核不育性遗传的复杂性
细胞质遗传的特点
正交和反交遗传表现不同。F1只表现母本性状; 非孟德尔遗传,杂交后代不表现有比例的分离, 基因定位困难; 通过连续回交能将母本 核基因置换掉,但母本 细胞质基因及其所控制 性状仍不消失。
1.2 细胞质遗传与细胞核遗传的比较
细胞质DNA 突变频率大 较强的定向突变性 正反交不一样 基因通过雌配子传递 基因定位困难 分离无规律 细胞间分布不均匀 某些基因有感染性 核DNA 突变频率较小 难于定向突变性 正反交一样 基因通过雌雄子传递 杂交方式进行基因定位 有规律分离 细胞间分布均匀 无感染性
3.2.2 叶绿体基因组的构成 低等植物的叶绿体基因组
仅能编码叶绿体本身结构和 组成的一部分物质。如各种 RNA、核糖体(70S)蛋白质、 光合作用膜蛋白和RuBp羧化 酶 8个大亚基; 基因功能与抗药性、温度敏 感性和某些营养缺陷有关。
细胞质遗传

紫茉莉
天竺葵
♀ ♂♀ ♂
亲代 绿色×花斑 花斑×绿色
↓ 子一代 绿色
↓ 绿色、花斑
白色
♀♂ ♀♂
绿色×花斑 花斑×绿色
↓
↓
大部分绿色 大部分花斑
少部分花斑 少部分绿色
紫茉莉花斑完全通过母本遗传 天竺葵由双亲遗传,但母本是叶绿体的主要供体
四、衣藻的核外基因
单细胞绿藻 真核生物 含有一个大的叶绿体
生活史包括有性过程和无性过程 雌雄异株,两种交配类型称为
二、面粉蛾的眼色 ---短暂的母性影响
A 幼虫皮肤有色 复眼深褐色
a 幼虫皮肤不着色 复眼红色 AA × aa ↓ Aa
正、反交幼虫都表现为有色
♂Aa × aa♀ 有色 无色 褐眼 红眼
1/2 Aa 1/2 aa 幼虫 有色 无色
褐眼 红眼
♀Aa × aa♂ 有色 无色 褐眼 红眼
1/2 Aa 1/2 aa 有色 有色?? 褐眼 红眼
细胞质: N1S1、N2S2、……、NnSn 核基因: r1R1、r2R2、……、rnRn
核内的育性基因总是与细胞质中的育性基因发 生对应的互作: r1(或R1)对N1(或S1)、r2(或R2)对N2(或S2)、……、 rn(或Rn)对Nn(或Sn)
3、单基因不育性和多基因不育性
单基因不育性:一对或两对核内主基因与对应 的不育胞质基因决定的不育性 多基因不育性:由两对以上的核基因与对应的 胞质基因共同决定的不育性
1、 遗传方式是非孟德尔式的; 杂交后代一般不表现一定比例的分离;
2、 正交和反交的遗传表现不同; F1通常只表现母本的性状;
3、 通过连续回交能将母本的核基因几乎全部置 换掉,但母本的细胞质基因及其所控制的性状 仍不消失;
细胞质遗传

11.1 细胞质遗传的概念和特点
11.1.2 细胞质遗传的特点
p244
❖ 正反交结果不同,且F1代的表现总是和母本 一致,故又称之为母性遗传。
❖ 不出现Mendel式分离比
❖ 通过连续回交能把母本的核基因全部置换掉,
但母本的细胞质基因及其控制性状仍不消失。
❖ 非细胞器基因组中遗传物质的传递类似病毒
链霉素抗性表型同正接合型(mt+)亲代的表型一致, mt-型细胞的叶绿体在接合中被丢失
玉米埃型条纹叶的遗传
玉米第7染色体上 (核基因)
正常绿色叶(Ij) 白色条纹叶(ijij) 母本为正常绿叶表现
孟得尔式遗传
p248-249
母本为条形叶不表现孟得尔式遗传
玉米条纹叶遗传的解释
条纹叶基因纯合(ijij)时,会引起叶绿体基因变异
花的植物,雄蕊发育不正常,不能产生有正常功能 的花药、花粉
雌蕊发育正常,能接受正常花粉而受精结实
雄性不育现象的存在与意义:
广泛存在
用于杂种优势利用中,杂交种制种
以不育系作为母本,免除人工去雄的麻烦,
节约人力,降低种子成本,并能保证种子纯度。
雄性不育在植物界是很普遍的,迄今已在18个 科的110多种植物中发现了雄性不育性的存在。
与野生型菌落接合
– 二倍接合子的线粒体基因组均来于野生型菌落 – 后代均表现为野生型菌落
11.2.3 叶绿体基因组遗传
p168
叶绿体的遗传特点
叶绿体具有遗传自主性:
既能自我复制,又能独立表达(决定性状表现)
叶绿体遗传的自主性是相对的:
可能与核基因共同决定性状表现 在某些情况下受核基因影响(甚至产生细胞质基因突变)
花斑:
– 局部不能形成叶绿素 具有叶绿体白色体两类
细胞质和遗传

Dd 1/4DD(右) 1/2Dd(右) 1/4dd(右)
1/4X1/2 1/4X1/2 1/2X1/2
3/8DD 右
2/8Dd 右
1/8dd 2/8dd 右 左
第三节
叶绿体遗传
一、紫茉莉花斑性状的遗传
白色植株 (幼年死亡)
绿色植株
花斑植株
(白色)
白色体 细胞核
叶绿体 (绿色) (白色) (绿色) (绿色)
实验结果分析
1.
F1总是表现出母本性状。
•
象这样,具有相对性状的亲本杂交,F1总是表现 母本性状的遗传现象,叫做母系遗传。
2. F1的性状不会出现一定的分离比。
结论:这种遗传现象不符合孟德尔遗传的定律。
推论:具有以上两种特征的遗传现象就是细胞质遗传。
原因: 叶绿体是由前体变异形成的.
正常质体 正常质体 白色质体 细胞 白色质体 正常和白色
瘦小、空
不开裂 少、不散出 没有或少 不结实
肥大、饱满
开裂 多、散出 多 结实
雌性不育:无实践意义
不 育
雄性不育
质遗传型 质核互作遗传型(应用价值最大) 核遗传型
一、雄性不育的遗传特点:
1、质不育型:
不育性只能被保持而不能被恢复。
通过远缘杂交培育。 2、核不育型: 由核内染色体上基因决定。 败育十分彻底,可育株与不育株有明显界限。
杂交试验:
结合时间短
结合时间长
放毒现象与内共生体 共生体:
不是细胞生存所需要的必要组成部分,仅以某 种共生的形式存在于细胞之中的细胞质颗粒。
能够自我复制,或在核基因组的作用下进行 复制; 对寄主表现产生影响,类似细胞质遗传。
1、假设下面每种草履虫细胞都自体受精,给定两 种细胞基因型,指出它们是放毒型还是敏感型: KK有卡巴粒; KK没有卡巴粒;
第十章细胞质遗传

基因组印迹(基因组印记) 与表观遗传学
来自父母双方的同源染色体或等位基因存在着功能上的 差异,子代中来自不同性别亲本的同一染色体或基因,当 发生改变时可以引起不同的表型,这种现象称为基因组印 迹。 基因组印迹的概念也可以理解为,基因组在传递信息的过 程中对基因或DNA片段打下标识、烙印的过程。 如果母源染色体的基因被印迹,则父源染色体上的等位 基因得到表达;如果父源染色体的基因被印迹,则母源染 色体上的等位基因得到表达。 基因组印迹不服从孟德尔定律,是一种典型的非孟德尔 遗传。
基因组印迹(基因组印记) 与表观遗传学
孟德尔遗传规律认为不管遗传物质来自双亲的哪 一方,都具有相同的表型效应。但是,有些遗 传现象是例外的—— 公驴与母马杂交生成骡子,公马与母驴的 后代称为駃騠(jueti),两者有相当显著的差别。 具有两条母源的11号染色体的小鼠在胚胎 期比正常小鼠小,而具有两条父源的11号染色 体的小鼠在胚胎期比正常小鼠大,但这两种胚 胎都死于发育阶段。
♂Igf-2m / Igf-2m × ♀Igf-2 / Igf-2 ↓ Igf-2▲ / Igf-2m 母源的Igf-2被印迹,父源的Igf-2m表 达,后代为矮小型小鼠。 试验结果表明:
1. 2. 3.
正反交结果不一样; 都是母源的基因被印迹,是一种母系印迹; 被印迹的基因表达受抑制,失去生物学功能。
质核互作不育性由不育的细胞质基因和 与其相对应的核内不育基因共同决定。 胞质不育基因 S 对应的可育基因 N 核内不育基因 r 对应的可育基因 R R又称为育性恢复基因
核 质 互 作 不 育 性 遗 传 示 意 图
核质互作雄性不育性的遗传
三系配套
细胞质遗传

条纹♂ ijij
F1
绿色
Ijij
×
F2 绿色 绿色 条纹 IjIj : Ijij : ijij
3
:1
母本正常,表现孟德尔 遗传,显示绿色与非绿 色是一对基因的差别。
反交:
P 条纹♀ ×
ijij
绿色♂ IjIj
F1 绿色 白色 条纹 Ijij Ijij Ijij
(无一定比例)
(连续回交)
条纹♀ × 绿色♂
卵 细 胞
卵
卵
细
细
胞
胞
• 母体遗传:
真核生物有性过程: 卵细胞:有细胞核、大量的细胞质和细胞器;
能为子代提供核基因和胞质基因。 精细胞:只有细胞核,细胞质或细胞器极少或没有;
只能提供其核基因,不能或极少提供胞质基因。 所以:一切受细胞质基因所决定的性状,其遗传信息只能通过
卵细胞传给子代,而不能通过精细胞遗传给子代。
↓异体受精 左旋(Dd)
↓自体受精 1DD∶2Dd∶1dd(均右旋) ↓↓↓ 右旋 右旋 左旋 (DD)(分离)(dd)
3∶1
3∶1
② 原因:
16
椎实螺外壳旋转方向是由受精卵螺旋式分裂时纺锤体分裂方向 决定的,并由受精前的母体基因型决定。
左旋──受精卵纺锤体向中线左侧分裂 ;
右旋──受精卵纺锤体向中线右侧分裂 。
③ 分析:
缓慢生长突变型在幼嫩培养阶段时无细胞色素氧化酶, 酶是正常代谢所必需的。
由于细胞色素氧化酶的产生与线粒体有关,缓慢生长突变 型中的线粒体结构不正常。
结论:有关基因存在于线粒体之中,由母本遗传。
2.酵母小菌落的遗传:
小菌落酵母细胞缺少细胞色 素a和b、细胞色素氧化酶
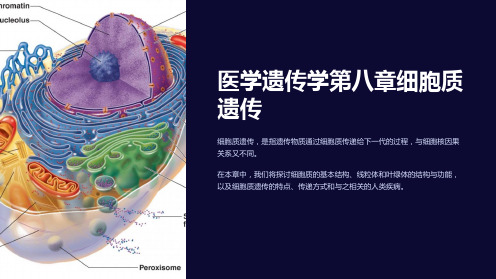
医学遗传学第八章细胞质遗传
叶绿体的功能
叶绿体的主要功能是进行光合作用,将太阳能 转化为植物所需的能量。
细胞质遗传的特点
• 细胞质遗传受到母系遗传的支配,由母亲通过卵细胞传递给下一代。 • 线粒体和叶绿体携带着自己的遗传物质,与细胞核基因共同决定个体
性状。 • 细胞质遗传的特征包括不完全性和限制性。
医学遗传学第八章细胞质 遗传
细胞质遗传,是指遗传物质通过细胞质传递给下一代的过程,与细胞核因果 关系又不同。
在本章中,我们将探讨细胞质的基本结构、线粒体和叶绿体的结构与功能, 以及细胞质遗传的特点、传递方式和与之相关的人类疾病。
细胞质的基本结构
细胞质是细胞核外的胞质区域,包括细胞质基质和细胞内膜系统。它起到了维持细胞形态、提供营养 和能量等重要功能。
线粒体的结构
线粒体是由两层膜组成的,其内膜上有许多折 叠形成的结构,称为线粒体内膜嵴。
线粒体的功能
线粒体的主要功能是进行细胞呼吸,产生细胞 所需的能量。
叶绿体的结构和功能
叶绿体是植物细胞中的特殊细胞器,负责进行光合作用,将阳光转化为化学能。 • 叶绿体内部有叶绿素颗粒,其中含有光合色素和光合膜。 • 叶绿体能够利用光能将二氧化碳和水转化为有机物质和氧气。 • 叶绿体具有独立的DNA和RNA分子,可以自主进行蛋白质合成。
细胞质遗传的传递方式
• 细胞质遗传通过卵细胞传递给后代,通常不受精子细胞的影响。 • 通过杂交和纯合线等方法,可以确定细胞质的遗传特性。 • 细胞质遗传的发生与遗传物质在细胞质中的复制和传递过程有关。
与细胞质遗传相关的人类疾病
1. 临床表现为线粒体功能障碍,如线粒体病。 2. 罕见的细胞质遗传性疾病,如MELAS综合征和MERRF综合征。 3. 一些癌症和神经退行性疾病与细胞质遗传也有关联。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
例题分析:
1、紫茉莉枝条颜色遗传属于 ,若用花斑枝条接受白 色枝条植株的花粉,发育成的植株的表现型为 。
2、透明金鱼(T)和普通金鱼(t)杂交实验,正交和反 交的结果一样,F1均为五花鱼。金鱼的体色遗传属于 。 3、果实白色的南瓜(A)与果实黄色(a)的南瓜杂交, 正交和反交所结的南瓜颜色都与母本一致,但是F1所结 的南瓜全为白色的。南瓜颜色的遗传属于 。 4、椎实螺外壳的螺旋方向,右旋对左旋是显性。杂交实 验,当右旋(母本) ×左旋(父本)时,F1为右旋;当 左旋(母本) ×右旋(父本)时,F1为左旋,则椎实螺 外壳螺旋方向的遗传为 。
巩固练习:
1 豌豆的硬豆荚对软豆荚为显性,现有两种经过多 年种植的豌豆品种硬豆荚M和软豆荚N。用M作母本, N作父本杂交,M结硬豆荚;用M作父本,N作母本杂 交,N结软豆荚。则控制豆荚硬度基因的位置( ) A、在细胞核中 B、在细胞质中 C、在质粒中 D、不能确定 D
2 叶绿体是植物光合作用的细胞器,其内部 具有独立遗传叶绿体基因。下列细胞全都不含叶 绿体基因的是 A、仙人掌茎细胞; B、洋葱根尖细胞 C、C3植物维管束鞘细胞; D、 光合细菌细胞 D 3 植物有性生殖产生的子代常常发生性状分离,无性 生殖产生的子代一般保持了母本的性状。用紫茉莉花 斑枝条所开的花自花授粉,所得子代的表现型 为: 绿枝条、白枝条、花斑枝条 。用花斑紫 茉莉植株的根尖细胞通过组织培养大量生产幼苗,所 得植株的表现型为 绿枝条、白枝条、花斑枝条 。
Hale Waihona Puke 白色花斑花斑 绿色 白色 花斑
绿色、白色、花斑 绿色、白色、花斑
二、对细胞质遗传现象的解释(一)一般情况
绿枝条 母本细胞 (二)紫 茉莉枝条 颜色遗传 原因 各种父 本精子 卵 细 胞
白枝条 母本细胞
花斑枝条 母本细胞
三、细胞质遗传的特点 (一)F1的性状由母本决定(母系遗传); (二)后代不出现一定的性状分离比(细胞质中遗 传物质随机的、不均等分配到子细胞)。
4、某家庭,母亲患肌无力遗传病,其所生儿子均 患病。要确定该遗传病由细胞核基因决定,还是由 细胞质基因决定,可采用的杂交方法是正交和反交 。 5、紫茉莉花色是受细胞核基因控制的,枝条颜色 (花斑、绿色、白色)是受细胞质基因控制的。基因 型为RR、Rr、rr的紫茉莉花色分别为红色、粉色、 白色。现有一开粉花花斑枝条紫茉莉植株,在互相传 粉的条件下,若其花斑枝条上所得子代经培育全部完 成个体发育并开始有性生殖,这些子代的表现型为 9 种。若用该植株选育红花子代,最好采用 绿色 枝条所 结的种子。该枝条所结种子长成的植株的表现型及 比例为 。 绿枝条红花:绿枝条粉花:绿枝条白花=1:2:1
例题:
1、杂交子一代雄性不育系,基因型为S(rr),则它的双 亲可能的基因型为( ) A、♀N(rr)×♂S(rr) B、♀S(Rr)×♂N(rr)C、 ♀S(rr)×♂S(rr) D、♀S(rr)×♂S(RR)
2、某农场由于不慎,在一块培养不育系的地里把雄性不 育系、保持系和恢复系种到了一块地里,其中(1和3保持 系,2和4不育系,5恢复系),请回答: (1) 在1和3植株上获得的种子的基因型是 。 (2) 在2和4植株上获得的种子的基因型是 。 (3) 在5植株上获得的种子的基因型是 。 (4) 如果该农场是为了培育不育系品系,在幼苗期发现了这一 现象时,可采取的补救措施是 。
第 三 章
复习:
遗 传 与 基 因 工 程
1、自然界中那些生物可以固氮? 2、采用何种措施水稻、小麦、大肠杆菌自行固氮?
3、将固氮基因转移到水稻、大肠杆菌中,那种生物更 容易获得自行固氮能力?为什么?
第一节 细胞质遗传
一、实例 1909年德国生物学家科伦斯----紫茉莉质体遗传试验
接受花粉的枝条:♀ 提供花粉的枝条:♂ 绿色 绿色 白色 花斑 绿色 白色 种子发育成的植株:F1 绿色 绿色 绿色 白色 白色 白色 绿色、白色、花斑
5、生物体的遗传是由( ) A、细胞核遗传作用的结果 B、细胞质遗传作用的结果 C、细胞核遗传和细胞质遗传共同作用的结果 D、外界环境条件变化而引起的结果 6、下列关于细胞质基因的存在位置的说法中正确的是 A、存在于细胞质内某些细胞器的染色体上 B、存在于细胞质中某些细胞器内的DNA上 C、存在于细胞质和细胞核内 D、存在于染色体上 7、紫罗兰的胚表皮颜色有黄色和深蓝色之别。将胚表皮 为黄色的紫罗兰花粉传授于胚表皮为深蓝色的雌蕊上,所 得后代的胚表皮呈深蓝色;将胚表皮为深蓝色的紫罗兰花 粉传授于胚表皮为黄色的紫罗兰雌蕊上,所得后代的胚表 皮则呈黄色。紫罗兰胚表皮颜色遗传属于 。