生物化学 第13章 代谢调节
第十三章 脂类代谢--王镜岩《生物化学》第三版笔记(完美打印版)

第十三章脂类代谢第一节概述一、生理功能(一)储存能量,是水化糖原的6倍(二)结构成分,磷脂、胆固醇等(三)生物活性物质,如激素、第二信使、维生素等二、消化吸收(一)消化:主要在十二指肠,胰脂肪酶有三种:甘油三酯脂肪酶,水解生成2-单脂酰甘油需胆汁和共脂肪酶激活,否则被胆汁酸盐抑制;胆固醇酯酶,生成胆固醇和脂肪酸;磷脂酶A2,生成溶血磷脂和脂肪酸。
食物中的脂肪主要是甘油三酯,与胆汁结合生成胆汁酸盐微团,其中的甘油三酯70%被胰脂肪酶水解,20%被肠脂肪酶水解成甘油和脂肪酸。
微团逐渐变小,95%的胆汁酸盐被回肠重吸收。
(二)吸收:水解产物经胆汁乳化,被动扩散进入肠粘膜细胞,在光滑内质网重新酯化,形成前乳糜微粒,进入高尔基体糖化,加磷脂和胆固醇外壳,形成乳糜微粒,经淋巴系统进入血液。
甘油和小分子脂肪酸(12个碳以下)可直接进入门静脉血液。
(三)转运:甘油三酯和胆固醇酯由脂蛋白转运。
在脂蛋白中,疏水脂类构成核心,外面围绕着极性脂和载脂蛋白,以增加溶解度。
载脂蛋白主要有7种,由肝脏和小肠合成,可使疏水脂类溶解,定向转运到特异组织。
1. 乳糜微粒转运外源脂肪,被脂肪酶水解后成为乳糜残留物。
2. 极低密度脂蛋白转运内源脂肪,水解生成中间密度脂蛋白,(IDL或LDL1),失去载脂蛋白后转变为低密度脂蛋白,3. 低密度脂蛋白又称β脂蛋白,转运胆固醇到肝脏。
β脂蛋白高易患动脉粥样硬化。
4. 高密度脂蛋白由肝脏和小肠合成,可激活脂肪酶,有清除血中胆固醇的作用。
LDL/HDL称冠心病指数,正常值为2.0+_0.75. 自由脂肪酸与清蛋白结合,构成极高密度脂蛋白而转运。
第二节甘油三酯的分解代谢一、甘油三酯的水解(一)组织脂肪酶有三种,脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步水解R3、R1、R2,生成甘油和游离脂肪酸。
(二)第一步是限速步骤,肾上腺素、肾上腺皮质激素、高血糖素通过cAMP和蛋白激酶激活,胰岛素和前列腺素E1相反,有抗脂解作用。
生物化学代谢部分课件-物质代谢的调节及代谢网络3

与酶之间的相互作用;
• 有机分子与酶或蛋白质受体之间的相互作用; • 底物与酶分子之间的识别以及相互作用; • 无机金属离子与生物大分子之间的相互作用,如金
属离子与酶或蛋白质之间的络合及与生物小分子 (辅酶、ATP等)之间的络合作用。
激素可改变酶的催化活性或含量,也可改变 细胞内代谢物的浓度,从而影响代谢反应的速 度--激素水平的调节。高等动物不仅有完整的 内分泌系统,还有功能复杂的神经系统。在中 枢神经的控制下,或者通过神经递质对效应器 直接发生影响,或通过改变某些激素的分泌来 调节某些细胞的功能状态,并通过各种激素的 互相协调对整体代谢进行综合调节--整体水平 的调节。
A.
B.
疫
蛋泛 白素
泛 素
内 源
酶化 蛋 性
体的 白 抗
降内 解源 成性 肽抗
酶 体 降 解
原 在 胞
段原 途 内
被径的 28 降
S
免解
酶原的激活
• 有些酶在生物体内合成出来的是它的无活性前 体--酶原。一定的条件下,这些酶原水解去除 一部分肽链,使酶的构象发生变化,形成有活 性的酶分子—酶原激活。酶原从无活性状态转 变成有活性状态的过程是不可逆的。属于这种 类型的酶有消化系统的酶(如胰蛋白酶、胰凝 乳蛋白酶和胃蛋白酶等)以及凝血酶等。
(1)被修饰的酶可以有两种互变形式,一种为 活性形式(具有催化活性),另一种为非活性 形式(无催化活性)。正反两个方向的互变均 发生共价修饰反应,且都将引起酶活性的变化。
(2)共价修饰调节作用可以产生酶的连续激活 现象,具有信号放大效应。例如肾上腺素引起 糖原分解过程中的一系列磷酸化激活步骤,其 结果将激素的信号逐级放大了约300万倍。
生物化学 第13章 代谢调节

第十三章代谢调节一、填空题:1.生物体内的代谢调节在三种不同的水平上进行,即、和。
2.代谢途径的终产物浓度可以控制自身形成的速度,这种现象被称为。
3.酶对细胞代谢的调节是最基本的代谢调节,主要有二种方式:和。
构通糖、脂代谢的关键化合物是。
4.不同代谢途径可以通过交叉点代谢中间物进行转化,在糖、脂、蛋白质及核酸的相互转化过程中三个最关键的代谢中间物是、和。
5.1961年,法国生物学家Monod和Jacob提出了关于原核生物基因结构及表达调控的学说。
6.正调控和负调控是基因表达的两种最基本的调节形式,其中原核细胞常用调控,而真核细胞常用调控模式。
7.乳糖操纵子的天然诱导物是,实验室里常用作为乳糖操纵子的安慰诱导物诱导β-半乳糖苷酶的产生。
8.许多代谢途径的第一个酶是限速酶,终产物多是它的,对它进行,底物多为其。
9.原核细胞酶的合成速率主要在水平进行调节。
10.乳糖操纵子的诱导物是,色氨酸操纵子的辅阻遏物是。
二、选择题(只有一个最佳答案):1.下列与能量代谢有关的过程除哪个外都发生在线粒体中?()A、糖酵解B、三羧酸循环C、脂肪酸的β-氧化D、氧化磷酸化2.IPTG可以诱导乳糖操纵子(lacOperon)的表达,这是因为:()A、IPTG与乳糖操作子(lacoperator)结合,诱导转录B、IPTG与LACI基因产物结合,并抑制其活性C、抑制β-半乳糖苷酶的活性D、促进Lac阻遏物的活性E、IPTG与LACI基因产物结合,并激活其活性3.在什么情况下,乳糖操纵子的转录活性最高?()A、高乳糖,低葡萄糖B、高乳糖,高葡萄糖C、低乳糖,低葡萄糖D、低乳糖,高葡萄糖4.真核细胞参与基因表达调节的调控区比原核细胞复杂是因为()A、真核细胞的细胞核具有双层膜B、原核细胞的基因总是以操纵子的形式存在C、原核细胞调节基因表达主要是在翻译水平D、真核细胞需要控制细胞特异性的基因表达E、真核细胞基因组含有太多的重复序列5.调节物质代谢体内最基础的层次是()A、细胞水平B、激素水平C、神经调节D、整体水平E、器官水平6.磷酸果糖激酶是什么代谢途径中的别构调节酶()A、三羧酸循环B、糖异生C、葡萄糖分解D、糖原合成E、糖原分解7.三羧酸循环中的别构调节酶是()A、柠檬酸合成酶B、α-酮戊二酸脱氢酶C、琥珀酸脱氢酶D、延胡索酸酶E、苹果酸脱氢酶8.催化糖酵解与磷酸戊糖途径的酶主要分布在细胞中什么部位()A、核B、胞质C、线粒体D、微粒体E、质膜9.催化三羧酸循环与脂肪酸β-氧化的酶分布在细胞内的什么部位()A、胞质B、胞膜C、胞核D、内质网E、线粒体10.氨基酸分解代谢调节的别构酶是()A、转氨酶B、脱羧酶C、转甲基酶D、己糖激酶E、谷氨酸脱氨酶11.糖异生限速酶的别构调节激活剂是()A、A TPB、ADPC、AMPD、dA TPE、cAMP 12.各种分解途径中,放能最多的途径是:()A、糖酵解B、三羧酸循环C、 -氧化D、氧化脱氨基13.操纵子调节系统属于哪一种水平的调节?()A、复制水平的调节B、转录水平的调节C、转录后加工的调节D、翻译水平的调节14.下列关于操纵基因的论述哪个是正确的?()A、能专一性地与阻遏蛋白结合B、是RNA聚合酶识别和结合的部位C、是诱导物和辅阻遏物的结合部位D、能于结构基因一起转录但未被翻译15.以下有关阻遏蛋白的论述哪个是正确的?()A、阻遏蛋白是调节基因表达的产物B、阻遏蛋白妨碍RNA聚合酶与启动子结合C、阻遏蛋白RNA聚合酶结合而抑制转录D、阻遏蛋白与启动子结合而阻碍转录的启动16.糖酵解中,下列哪一个催化的反应不是限速反应?()A、丙酮酸激酶B、磷酸果糖激酶C、己糖激酶D、磷酸丙糖异构酶17.磷酸化酶通过接受或脱去磷酸基而调节活性,因此它属于:()A、别(变)构调节酶B、共价调节酶C、诱导酶D、同工酶18.下列与能量代谢有关的途径不在线粒体内进行的是:()A、三羧酸循环B、脂肪酸β氧化C、氧化磷酸化D、糖酵解作用19.关于共价修饰调节酶,下列哪种说法是错误的?()A、这类酶一般存在活性和无活性两种形式,B、酶的这两种形式通过酶促的共价修饰相互转变C、伴有级联放大作用D、是高等生物独有的代谢调节方式20.阻遏蛋白结合的位点是:()A、调节基因B、启动因子C、操纵基因D、结构基因21.下面哪一项代谢是在细胞质内进行的:()A、脂肪酸的β-氧化B、氧化磷酸化C、脂肪酸的合成D、TCA22.在乳糖操纵子模型中,操纵基因专门控制是否转录与翻译。
糖代谢的生物化学调节

糖代谢的生物化学调节糖代谢是生物体内一个重要的代谢过程,通过一系列的生物化学反应,将摄入的碳水化合物转化为能量和存储形式。
这一过程涉及多个关键酶的调节,以保持机体内部代谢平衡。
本文将探讨糖代谢的生物化学调节机制。
1. 糖代谢的基本过程糖代谢的基本过程主要包括糖的吸收、储存、释放和利用。
当我们进食含糖食物时,消化系统中的酶将复杂的糖类分解为单糖,如葡萄糖。
这些单糖通过细胞膜转运蛋白进入细胞内,并在细胞质中进行代谢。
2. 葡萄糖调节机制葡萄糖是糖代谢的主要物质,其浓度在血液中需要维持在一定的范围内。
当血糖浓度过高时,胰岛素释放,促进葡萄糖的摄入和利用。
胰岛素通过激活葡萄糖转运蛋白和糖原合成酶,促使葡萄糖转化为糖原储存起来。
当血糖浓度过低时,胰岛素的分泌减少,肝细胞将糖原分解为葡萄糖释放到血液中,以维持血糖水平。
3. 糖原和糖酵解的调节糖原是一种储存在肝脏和肌肉中的多糖,能够释放葡萄糖以满足机体能量需求。
糖原的合成受到胰岛素的促进,而其分解则受到胰高血糖素和肾上腺素的调节。
当机体需要能量时,肾上腺素的分泌增加,激活糖原磷酸化酶,使得糖原分解为葡萄糖。
4. 糖酵解调节糖酵解是将葡萄糖分解为乳酸或丙酮酸的过程,产生少量的ATP。
当氧气供应不足时,糖酵解是细胞的主要能源来源。
糖酵解的过程中,多个关键酶受到调节,如磷酸果糖激酶、葡萄糖激酶和磷酸三磷酸异构酶等。
这些酶的活性可以通过磷酸化、糖酮-糖磷酸酯循环以及底物浓度等因素进行调节。
5. 糖异生的调节糖异生是指在机体无法通过摄入糖类满足能量需求时,通过非糖类物质合成葡萄糖。
糖异生主要发生在肝细胞中,其中多糖、脂肪和氨基酸是糖异生的补给物。
多个酶参与糖异生的调节,其中磷酸烯醇式还原酶和磷酸果糖-6-磷酸酶是关键酶,其活性受到内分泌激素和底物浓度的调控。
总结:糖代谢的生物化学调节涉及多个酶的活性调控,其中胰岛素和肾上腺素是重要的调节激素。
胰岛素在血糖浓度高时促进糖的储存和利用,而肾上腺素则在能量需求增加时促进糖原分解和糖酵解。
生物化学:第十三章 代谢调节

• 代谢调节普遍存在于生物界
单细胞生物
通过细胞内代谢物浓度的变 化,影响酶活性及含量,从而调 节代谢
——原始调节/细胞水平调节
高 等 生 细胞水平代谢调节 物
激素水平代谢调节
整体水平代谢调节
细胞水平代谢调节
细胞的膜结构及酶分布 在代谢的调节作用
酶活性的调节
多酶体系的 区域化分布
同工酶在调 节中的作用
GTP
已糖激酶
6-P-G
磷酸果糖激酶
6-P-F
磷酸果糖激酶1
6-磷酸果糖
1,6-二磷酸果糖
各种腺苷酸对磷酸果糖激酶的变构调节
变构调节的机制和特点
➢多数变构酶由多亚基构成,所以存在四级结构。 它们的变构调节一般体现在亚基的解聚和聚合上。 ➢多数变构酶由两种亚基组成:催化亚基和调节亚 基。 ➢变构酶有两种构象。 ➢变构剂与调节亚基以非共价键结合,两者的结合 程度取决于变构剂的浓度。 ➢变构调节快速短暂,一般在数分钟内完成。
甘油
某些非必需氨基酸
磷酸甘油醛
糖酵解途径
丙酮酸
其他α-酮酸
ቤተ መጻሕፍቲ ባይዱ
• 但不能说,脂类可转变为氨基酸
• 氨基酸可以转变为脂类
氨基酸
乙酰CoA
脂肪
• 氨基酸可作为合成磷脂的原料
丝氨酸
丝氨酸磷脂
胆胺
脑磷脂
胆碱
卵磷脂
4. 核酸与糖、蛋白质
代谢的相互联系
• 氨基酸是体内合成核酸的重要原料
甘氨酸
天冬氨酸
谷氨酰胺
一碳单位
酶结构 的调节
酶数量 的调节
一、代谢途径的区域化分布 1、代谢途径有关酶类常组成酶体系,分布
于细胞的某一区域或亚细胞结构中。
生物化学习题及答案_代谢调节

代谢调节(一)名词解释1.诱导酶(Inducible enzyme)2.标兵酶(Pacemaker enzyme)3.操纵子(Operon)4.衰减子(Attenuator)5.阻遏物(Repressor)6.辅阻遏物(Corepressor)7.降解物基因活化蛋白(Catabolic gene activator protein)8.腺苷酸环化酶(Adenylate cyclase)9.共价修饰(Covalent modification)10.级联系统(Cascade system)11.反馈抑制(Feedback inhibition)12.交叉调节(Cross regulation)13.前馈激活(Feedforward activation)14.钙调蛋白(Calmodulin)(二)英文缩写符号1. CAP(Catabolic gene activator protein):2. PKA(Protein kinase):3. CaM(Calmkdulin):4. ORF(Open reading frame):(三)填空题1. 哺乳动物的代谢调节可以在、、和四个水平上进行。
2. 酶水平的调节包括、和。
其中最灵敏的调节方式是。
3. 酶合成的调节分别在、和三个方面进行。
4. 合成诱导酶的调节基因产物是,它通过与结合起调节作用。
5. 在分解代谢阻遏中调节基因的产物是,它能与结合而被活化,帮助与启动子结合,促进转录进行。
6. 色氨酸是一种,能激活,抑制转录过程。
7. 乳糖操纵子的结构基因包括、和。
8. 在代谢网络中最关键的三个中间代谢物是、和。
9. 酶活性的调节包括、、、、和。
10.共价调节酶是由对酶分子进行,使其构象在和之间相互转变。
11.真核细胞中酶的共价修饰形式主要是,原核细胞中酶共价修饰形式主要是。
(四)选择题1. 利用操纵子控制酶的合成属于哪一种水平的调节:A.翻译后加工 B.翻译水平 C.转录后加工 D.转录水平2. 色氨酸操纵子调节基因产物是:A.活性阻遏蛋白 B.失活阻遏蛋白C.cAMP受体蛋白 D.无基因产物3. 下述关于启动子的论述错误的是:A.能专一地与阻遏蛋白结合 B.是RNA聚合酶识别部位C.没有基因产物 D.是RNA聚合酶结合部位4. 在酶合成调节中阻遏蛋白作用于:A.结构基因 B.调节基因 C.操纵基因 D.RNA聚合酶5. 酶合成的调节不包括下面哪一项:A.转录过程 B.RNA加工过程C.mRNA翻译过程 D.酶的激活作用6. 关于共价调节酶下面哪个说法是错误的:A.都以活性和无活性两种形式存在 B.常受到激素调节C.能进行可逆的共价修饰 D.是高等生物特有的调节方式7. 被称作第二信使的分子是:A.cDNA B.ACP C.cAMP D.AMP8.反馈调节作用中下列哪一个说法是错误的:A.有反馈调节的酶都是变构酶 B.酶与效应物的结合是可逆的C.反馈作用都是使反速度变慢 D.酶分子的构象与效应物浓度有关(五)是非判断题()1.分解代谢和合成代谢是同一反应的逆转,所以它们的代谢反应是可逆的。
专科(生物化学)第13章 酸碱平衡

KHb+H2CO3
Hb02
KHCO3+HHb
Hb+ 02
血浆
组织液
内流
(2)在肺部:
血液运行至肺部时。HHb与02结合成HHb02,后者 与KHCO3作用生成KHbO2和H2CO3,H2CO3再分解成CO2 呼出: HHb+ 02
HHb02+KHCO3
HHb02
KHbO2+ H2CO3
但肺不能调节NaHCO3的浓度,NaHCO3浓度 的恢复还需要肾进行调节。
三、肾对酸碱平衡的调节
肾通过重吸收与再生NaHCO3以及排出过多 的碱来调节血中NaHCO3浓度,以维持 [NaHCO3]/[ H2CO3]的正常比值。
这种调节作用是:由肾小管细胞的
泌氢、泌氨、泌钾及重吸收钠作用来完 成的。
血浆PH值可按亨德森一哈塞巴方程式计算: [ NaHCO3] PH=pKa+lg [ H2CO3] 式中的 pKa是H2CO3解离常数的负对数,在 37℃时为 6.l,将此值与[NaHCO3]/[H2CO3]的比值代入上式. pH=6.l+lg 20/l=6.l+1.3=7.4
血浆通过肾小球滤过而产生的原尿其 pH未变,仍 为7.4左右,[Na2HPO4]/[ NaH2PO4]的比值
也与血浆相同,为4/l。
当原尿流经肾小管时,肾小管细胞分泌出 的 H+与 Na2HPO4中的 Na+进行交换, Na2HPO4转变成 NaH2PO4随尿排出。 被重吸收的Na+则与肾小管细胞内的HCO3- 一起转运入血液形成NaHCO3。
糖、脂肪和蛋白质彻底氧化的终产物是二氧化碳和 水,两者化合生成碳酸(CO2+ H2O→ H2CO3 )。
物质代谢的联系与调节《生物化学》复习提要

物质代谢的联系与调节第一节物质代谢的特点(一)整体性体内各种物质包括糖、脂、蛋白质、水、无机盐、维生素等的代谢不是彼此孤立各自为政,而是同时进行的,而且彼此互相联系,或相互转变,或相互依存,构成统一的整体。
(二)代谢调节机体存在精细的调节机制,不断调节各种物质代谢的强度、方向和速度以适应内外环境的变化。
代谢调节普遍存在于生物界,是生物的重要特征。
(三)各组织、器官物质代谢各具特色由于各组织、器官的结构不同,所含有酶系的种类和含量各不相同,因而代谢途径及功能各异,各具特色。
例如肝在糖、脂、蛋白质代谢上具有特殊重要的作用,是人体物质代谢的枢纽。
(四)各种代谢物均具有各自共同的代谢池无论是体外摄人的营养物或体内各组织细胞的代谢物,只要是同一化学结构的物质在进行中间代谢时,不分彼此,参加到共同的代谢池中参与代谢。
(五)ATP是机体能量利用的共同形式糖、脂及蛋白质在体内分解氧化释出的能量,均储存在ATP的高能磷酸键中。
(六)NADPH是合成代谢所需的还原当量参与还原合成代谢的还原酶则多以NADPH为辅酶,提供还原当量。
如糖经戊糖磷酸途径生成的NADPH既可为乙酰辅酶A合成脂酸,又可为乙酰辅酶A 合成固醇提供还原当量。
第二节物质代谢的相互联系一、在能量代谢上的相互联系乙酰辅酶A是三大营养物共同的中间代谢物,三羧酸循环是糖、脂、蛋白质最后分解的共同代谢途径,释出的能量均以ATP形式储存。
从能量供应的角度看,这三大营养素可以互相代替,并互相制约。
二、糖、脂和蛋白质代谢之间的联系体内糖、脂、蛋白质和核酸等的代谢不是彼此独立,而是相互关联。
它们通过共同的中间代谢物,即两种代谢途径汇合时的中间产物,三羧酸循环和生物氧化等联成整体。
(一)糖代谢与脂代谢的相互联系当摄人的糖量超过体内能量消耗时,除合成少量糖原储存在肝及肌肉外,生成的柠檬酸及ATP可变构激活乙酰辅酶A竣化酶,使由糖代谢源源而来的大量乙酰辅酶A得以羧化成丙二酰辅酶A,进而合成脂酸及脂肪在脂肪组织中储存,即糖可以转变为脂肪。
《生物化学》-物质代谢的调节与控制

1.酶量调节机理
酶量调节的两种基本调节机制是诱导和阻遏
诱导:一些分解代谢的酶类只在有关的底物或底物 类似物存在时才被诱导合成。依赖于诱导物才能合 成的酶称为诱导酶。
阻遏:对于合成代谢的酶类,在产物或产物类似物 足够量存在时,其合成被阻遏。(反馈阻遏)
共价修饰调节是酶蛋白中的活性基团(-OH、SH、-COOH、-NH2)在其他酶的作用下发生共价 修饰,从而改变酶的活性。
共价修饰调节具有级联放大作用,效率高。
(三)酶量变化对代谢的调节(基因表达的调节控制)
细胞内酶浓度的改变也可以改变代谢速度。
主要是通过调节酶蛋白的合成过程实现的。 (1)活化基因则合成相应的酶,酶量增加; (2)钝化基因则停止酶的合成,酶量降低。
柠檬酸
+
–
乙酰辅酶A羧化酶 6-磷酸果糖激酶
促进脂酸的合成 抑制糖的氧化
2.共价修饰调节
(1)有些酶,在其它酶的催化下,其分子结构中的某 些基团,如:Ser、Thr或Tyr 的-OH 基,能与特殊的 化学基团,如ATP分子上脱下的磷酸基或腺苷酰基 (AMP),共价结合或解离,从而使酶分子活性形式发生 改变。这种修饰作用称为共价修饰调节。这种被修饰 的酶称为共价调节酶。
葡萄糖
分解代 谢产物
变构调节的生理意义
① 代谢终产物反馈抑制 (feedback inhibition) 反应途径中的酶,使代谢物不致生成过多。
乙酰CoA
丙二酰CoA
乙酰CoA羧化酶
长链脂酰CoA
②变构调节使能量得以有效利用,不致浪费。
G-6-P
+
–
糖原合酶
糖原磷酸化酶
生物化学基础第13章-血液的生物化学

血液化学成分与临床
人体在生理状态下血液化学成分含量相对恒定, 仅在一定范围波动。
如正常血NPN含量为 14.28~24.99 mmol/L, 其中血尿素氮(blood urea nitrogen,BUN)约占 NPN的1/2。
分析血液的化学成分,可以了解体内物质代谢 状况,有助于诊断和治疗疾病。
➢ 催化作用 许多具有催化作用的酶是血浆蛋 白。这些酶分为:血浆功能酶(凝血酶、纤溶 酶)、外分泌酶(AMS)与细胞酶(LDH、 ALT)。
➢ 营养作用 血浆蛋白在体内分解产生氨基酸, 参与氨基酸代谢池,用于组织蛋白合成,修复 损伤的组织,也可氧化分解供能,或转变为其 他含氮物质。
第二节 红细胞的代谢
➢ 在 组 织 , 红 细 胞 内 的 2 , 3-BPG 显 著 增 加 HbO2释放O2供组织需要。
➢ 缺O2时,红细胞内产生的2,3-BPG增加,有 利于释放更多的O2供组织需要。由平原登高 山、或贫血、肺气肿等,红细胞内2,3-BPG 增加,有利于组织获得较多的O2。
二、二氧化碳的运输
血液运输CO2的形式: (1)物理溶解
➢线粒体内δ-氨基-γ-酮戊酸(δ-aminolevulinic acid, ALA) 合酶(ALA synthase)催化琥珀酰CoA与甘 氨酸缩合生成ALA。 ➢ALA合酶的辅酶为磷酸吡哆醛。维生素B6缺乏 时,血红素合成发生障碍,造成维生素B6反应性 贫血。
琥珀酰CoA
甘氨酸
ALA由线粒体转运到胞液。 ALA脱水酶催化 两分子ALA缩合成胆色素原(PBG)。
PBG
在尿卟啉原Ⅰ同合酶(UPGⅠcoynthase) 及UPGⅢ同合酶协同作用下,4分子PBG脱 氨缩合生成尿卟啉原Ⅲ。
尿卟啉原Ⅲ
生物化学课件-代谢调节

所有這些調節機制都是在基因產物蛋白質(或RNA) 的作用下進行的。也就是說與基因表達調控有關。
二、酶水準的調節
酶水準調節是生物體內最基本、最普遍的調節方式。 包括酶含量調節、酶活性調節、酶的定位調節
(一)酶含(數)量調節 1、酶降解調節 蛋白水解酶催化使酶降解,控制酶的數量。 2、酶的合成調節--基因表達調控 在轉錄和翻譯水準上,都能進行調節,但主要是 在轉錄水準上。
細胞是如何經濟有效地轉化各類物質的?
代謝的總原則和方略是什麼?
誰在代謝調節中起主導作用?
一、代謝網路
細胞代謝有一套總原則和方略,來解決經濟有 效轉化問題:
(1)將各類物質分別納入各自的共同代謝途徑, 以少數種類的反應,轉化種類繁多的分子,例如, 氧化還原,基團轉移,水解合成,基團脫加,異 構反應等。
‥‥‥
基因
Lac z
Lac y
Lac a
基因關閉 不轉錄
(有活性)
阻遏蛋白
‥‥‥
阻遏蛋白阻擋操縱基因,結構基因不表達
基因打開
Lac z Lac y Lac a 轉錄
mRNA
阻遏蛋白
誘導物 (乳糖等)
(無活性)
-半乳 -半乳 糖苷酶 糖苷透
性酶
-半乳糖 苷乙醯基
轉移酶
誘導物與阻遏蛋白結合,阻遏蛋白不能阻擋操縱基因,結構基因表達。
第十二章 代謝調節
第一節 代謝途徑的相互聯繫 第二節 代謝調節
第一節 代謝途徑的相互聯繫
一、代謝網路 二、物質代謝的相互關係
一、代謝網路
細胞在新陳代謝過程中,細胞內數百種小分子在 起作用,它們構成了成千上萬的生物大分子,如 果這些分子單獨代謝互不相干,那麼代謝將變的 無比複雜,可想像細胞是無法容納的。那麼:
《代谢调节生物化学》课件

岛素和生长因子等相关。
3
MAPKs (mitogen-activated
protein kinases)
调节细胞增殖、分化和细胞死亡等重要
mTOR (mammalian target of rapamycin)
4
过程,与多个代谢疾病相关。
参与细胞生长和代谢调节,对于蛋白质 合成和能量平衡起重要作用。
2. Hardie DG. (2014). AMPK - sensing energy while talking to other signaling pathways. Cell Metab. 20(6): 939-952.
3. Lin SC, Hardie DG. (2018). AMPK: Sensing Glucose as well as Cellular Energy Status. Cell Metab. 27(2): 299-313.
糖皮质激素
调节糖、脂肪和蛋白质的代谢, 影响细胞能量平衡和炎症反应。
胰高血糖素
反调节胰岛素,升高血糖水平, 在饥饿状态下保持血糖稳定。
代谢调节的细胞信号传导机制
1
蛋白激酶A (PKA)
通过磷酸化酶和蛋白质结合,调节多种
蛋白激酶B (PKB)
2
酶和转录因子的活性,影响能量代谢。
参与细胞生长、存活和代谢调控,与胰
代谢物及其在代谢调节中的作用
ATP
作为能量储存和释放的分子,ATP在细胞能量代 谢以及信号传导中起着关键作用。
cAMP
腺苷环化酶产生的第二信使,调节多种细胞功 能和代谢途径。
A MPK
AMP激活的蛋白激酶,参与调节能量代谢平衡, 对疾病如糖尿病和肥胖症具有调节作用。
生物化学 第13章 糖原代谢和糖异生作用

贮存糖原的动用(主要发生在肝脏中)可以为脑和红细胞 提供对葡萄糖部分需要。当进食之后葡萄糖很丰富时,葡萄糖便 会很快地以糖原的形式贮存起来。然而,肝脏贮存糖原的能力仅 能满足脑每日对葡萄糖的一半的需要。在禁食的条件下,机体所
需的大部分葡萄糖则通过葡萄糖的异生作用从非糖前体(例如氨基
糖原合酶催化糖原合成 UDP-葡萄糖的生成具备了将糖基转移用
于糖原合成的条件。在糖原合酶(glycogen
synthase)的催化下,UDP-葡萄糖的糖基被转移到 分子较小的糖原(作为引物)的非还原性末端C-4– OH上,形成α-1,4-糖苷键:
UDP-葡萄糖 + 糖原(n个葡萄糖残基) →
UDP + 糖原(n + 1个葡萄糖基)
酸化酶b和有活性的磷酸化酶a。 1959年,E.Krebs和E.Fischer证实这种转换
涉及到磷酸化的共价修饰机制。
糖原磷酸化酶的共价修饰部位是每个亚基
第14位Ser残基。在Ser14被磷酸化酶激酶(一种转
换酶)催化而磷酸化时,糖原磷酸化酶由低活性的 b形式转变成有活性的a形式。
糖原磷酸化酶的活性变化是由于Ser14残基的
二
糖原合成酶催化糖原合成 UDP-葡萄糖的生成具备了将糖基转移用于糖
原合成的条件。在糖原合酶(glycogen synthase)的
催化下,UDP-葡萄糖的糖基被转移到分子较小的糖原 (作为引物)的非还原性末端C-4–OH上,形成α-1,4糖苷键:
UDP-葡萄糖 + 糖原(n个葡萄糖残基) →
UDP + 糖原(n + 1个葡萄糖基)
一.
糖原磷酸化酶和糖原合酶的别构调节
糖原磷酸化酶可被AMP别构激活,而ATP和葡萄糖-
代谢调节与代谢工程2
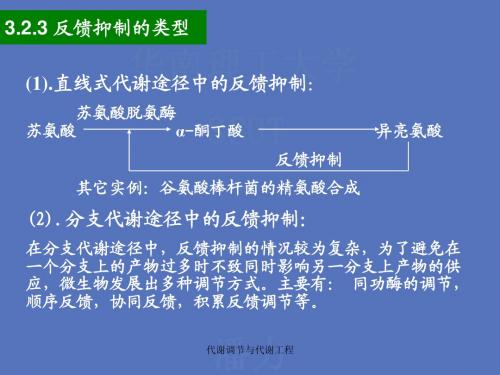
代谢调节与代谢工程
天冬氨酸族 (谷氨酸棒 杆菌)
E
二氢吡啶二羧酸 R 六氢吡啶二羧酸 二氨基庚二酸
天冬氨酸酸半醛
同型丝氨酸 E,R
E,R
E 同型丝氨酸磷酸 苏氨酸 E,R 2-酮丁酸
O-琥珀酰同型丝氨酸 胱硫醚 R 同型半胱氨酸 R 甲硫氨酸 代谢调节与代谢工程
赖氨酸
异亮氨酸
合作反馈抑制
定义:两种末端产物同时存在时,共同的反馈抑制作用大于二 者单独作用之和。 举例:在嘌呤核苷酸合成中,磷酸核糖焦磷酸酶受AMP和 GMP (和IMP)的合作反馈抑制,二者共同存在时,可以完 全抑制该酶的活性。而二者单独过量时,分别抑制其活性的 70%和10%。
代谢调节与代谢工程
(3)积累反馈抑制
代谢调节与代谢工程
代谢调节与代谢工程
(2)协同反馈抑制
定义:分支代谢途径中几个末端产物同时过量时才能抑制 共同途径中的第一个酶的一种反馈调节方式。 举例:谷氨酸棒杆菌(Corynebacterium glutamicum) 多粘芽孢杆菌(Bacillus polymyxa) 天冬氨酸族氨基酸合成中天冬氨酸激酶受赖氨酸和 苏氨酸的协同反馈抑制和阻遏。
代谢调节与代谢工程
回 补 顺 序
谷氨酸脱氢酶
谷氨酸
-酮戊二酸氧化酶
+NH3
代谢调节与代谢工程
1.谷氨酸生产菌中存在2个糖酵解途径EMP/HMP 生物素参与糖代谢作用:增加糖代谢的速度 而丙酮酸氧化脱羧的速度未改变
丙酮酸积累
乳酸积累 2.α-酮戊二酸氧化酶缺失
TCA环阻断, α-酮戊二酸积累
代谢调节与代谢工程
谷氨酸发酵 一、谷氨酸发酵菌种: Corynebacterium pekinense
生物化学-考试知识点_7物质代谢调节整理(1)

物质代谢调节1.细胞水平调节:细胞水平的调节主要是细胞内酶水平的调节。
方式:细胞内酶呈隔离分布、代谢调节作用点(限速酶、关键酶)、酶的别构调节、酶的化学修饰、同工酶对物质代谢的调节、酶含量的调节2.关键酶:催化代谢途径定向步骤的酶,往往是代谢途径反应的第一个酶。
在可逆反应中偏向一个方向,决定着多酶体系的催化方向。
限速酶:体内代谢是一系列酶促反应的总和。
整个代谢途径速度取决于多酶体系中催化活力最低、米氏常数最大、催化反应速度最慢的酶。
此酶起着限速作用,代谢调节的作用点。
生理意义:①限速酶的催化活力最低,Km最大,催化反应速度最慢,故它的速度决定了整个代谢途径的总速度。
②关键酶多为催化各代谢途径反应的第一个酶,在催化可逆反应中往往极度偏向一个方向,故它的定向决定着多酶体系催化代谢反应的方向。
③代谢调节主要是通过对限速酶与关键酶活性的调节而实现的,而关键酶大多同时又是限速酶,所以它们是代谢调节的作用点。
例:己糖激酶3.酶的别构调节:小分子化合物与酶分子活性中心以外的某一部位特异结合,引起酶蛋白分子构象的轻微改变,从而引起酶活性的改变,这种调节称为酶的别构调节。
方式:生理意义:①代谢终产物反馈抑制反应途径中的酶,使代谢物不致生成过多。
②别构调节使机体维持在相对恒定的生理状态。
例:HMG-CoA还原酶4.酶的化学修饰:酶蛋白肽链上某些残基在酶的催化下发生可逆的共价修饰,从而引起酶活性改变,这种调节称为酶的化学修饰。
生理意义:①催化的反应具有放大效应,比别构调节调节效率高。
②消耗的ATP少于酶蛋白合成所需。
③比酶蛋白合成的调节迅速。
④是体内酶活性经济、高效的调节方式。
例:磷酸化酶。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第十三章代谢调节一、填空题:1.生物体内的代谢调节在三种不同的水平上进行,即、和。
2.代谢途径的终产物浓度可以控制自身形成的速度,这种现象被称为。
3.酶对细胞代谢的调节是最基本的代谢调节,主要有二种方式:和。
构通糖、脂代谢的关键化合物是。
4.不同代谢途径可以通过交叉点代谢中间物进行转化,在糖、脂、蛋白质及核酸的相互转化过程中三个最关键的代谢中间物是、和。
5.1961年,法国生物学家Monod和Jacob提出了关于原核生物基因结构及表达调控的学说。
6.正调控和负调控是基因表达的两种最基本的调节形式,其中原核细胞常用调控,而真核细胞常用调控模式。
7.乳糖操纵子的天然诱导物是,实验室里常用作为乳糖操纵子的安慰诱导物诱导β-半乳糖苷酶的产生。
8.许多代谢途径的第一个酶是限速酶,终产物多是它的,对它进行,底物多为其。
9.原核细胞酶的合成速率主要在水平进行调节。
10.乳糖操纵子的诱导物是,色氨酸操纵子的辅阻遏物是。
二、选择题(只有一个最佳答案):1.下列与能量代谢有关的过程除哪个外都发生在线粒体中?()A、糖酵解B、三羧酸循环C、脂肪酸的β-氧化D、氧化磷酸化2.IPTG可以诱导乳糖操纵子(lacOperon)的表达,这是因为:()A、IPTG与乳糖操作子(lacoperator)结合,诱导转录B、IPTG与LACI基因产物结合,并抑制其活性C、抑制β-半乳糖苷酶的活性D、促进Lac阻遏物的活性E、IPTG与LACI基因产物结合,并激活其活性3.在什么情况下,乳糖操纵子的转录活性最高?()A、高乳糖,低葡萄糖B、高乳糖,高葡萄糖C、低乳糖,低葡萄糖D、低乳糖,高葡萄糖4.真核细胞参与基因表达调节的调控区比原核细胞复杂是因为()A、真核细胞的细胞核具有双层膜B、原核细胞的基因总是以操纵子的形式存在C、原核细胞调节基因表达主要是在翻译水平D、真核细胞需要控制细胞特异性的基因表达E、真核细胞基因组含有太多的重复序列5.调节物质代谢体内最基础的层次是()A、细胞水平B、激素水平C、神经调节D、整体水平E、器官水平6.磷酸果糖激酶是什么代谢途径中的别构调节酶()A、三羧酸循环B、糖异生C、葡萄糖分解D、糖原合成E、糖原分解7.三羧酸循环中的别构调节酶是()A、柠檬酸合成酶B、α-酮戊二酸脱氢酶C、琥珀酸脱氢酶D、延胡索酸酶E、苹果酸脱氢酶8.催化糖酵解与磷酸戊糖途径的酶主要分布在细胞中什么部位()A、核B、胞质C、线粒体D、微粒体E、质膜9.催化三羧酸循环与脂肪酸β-氧化的酶分布在细胞内的什么部位()A、胞质B、胞膜C、胞核D、内质网E、线粒体10.氨基酸分解代谢调节的别构酶是()A、转氨酶B、脱羧酶C、转甲基酶D、己糖激酶E、谷氨酸脱氨酶11.糖异生限速酶的别构调节激活剂是()A、ATPB、ADPC、AMPD、dATPE、cAMP 12.各种分解途径中,放能最多的途径是:()A、糖酵解B、三羧酸循环C、 -氧化D、氧化脱氨基13.操纵子调节系统属于哪一种水平的调节?()A、复制水平的调节B、转录水平的调节C、转录后加工的调节D、翻译水平的调节14.下列关于操纵基因的论述哪个是正确的?()A、能专一性地与阻遏蛋白结合B、是RNA聚合酶识别和结合的部位C、是诱导物和辅阻遏物的结合部位D、能于结构基因一起转录但未被翻译15.以下有关阻遏蛋白的论述哪个是正确的?()A、阻遏蛋白是调节基因表达的产物B、阻遏蛋白妨碍RNA聚合酶与启动子结合C、阻遏蛋白RNA聚合酶结合而抑制转录D、阻遏蛋白与启动子结合而阻碍转录的启动16.糖酵解中,下列哪一个催化的反应不是限速反应?()A、丙酮酸激酶B、磷酸果糖激酶C、己糖激酶D、磷酸丙糖异构酶17.磷酸化酶通过接受或脱去磷酸基而调节活性,因此它属于:()A、别(变)构调节酶B、共价调节酶C、诱导酶D、同工酶18.下列与能量代谢有关的途径不在线粒体内进行的是:()A、三羧酸循环B、脂肪酸β氧化C、氧化磷酸化D、糖酵解作用19.关于共价修饰调节酶,下列哪种说法是错误的?()A、这类酶一般存在活性和无活性两种形式,B、酶的这两种形式通过酶促的共价修饰相互转变C、伴有级联放大作用D、是高等生物独有的代谢调节方式20.阻遏蛋白结合的位点是:()A、调节基因B、启动因子C、操纵基因D、结构基因21.下面哪一项代谢是在细胞质内进行的:()A、脂肪酸的β-氧化B、氧化磷酸化C、脂肪酸的合成D、TCA22.在乳糖操纵子模型中,操纵基因专门控制是否转录与翻译。
()A、结构基因B、调节基因C、起动因子D、阻遏蛋白23.按照操纵子学说,对基因转录起调控作用的是:()A、诱导酶B、阻遏蛋白C、RNA聚合酶D、DNA聚合酶三、是非题(在题后括号内打√或×):1.蛋白激酶和蛋白磷酸酶对蛋白质进行磷酸化和去磷酸化的共价修饰是真核细胞代谢的重要方式。
()2.共价修饰调节酶被磷酸化后活性增大,去磷酸化后活性降低。
()3.操纵基因又称操纵子,如同启动基因又称启动子一样。
()4.别构酶又称变构酶,催化反应物从一种构型转化为另一种构型。
()5.细胞内区域化在代谢调节上的作用,除把不同的酶系统和代谢物分隔在特定区间外,还通过膜上的运载系统调节代谢物、辅助因子和金属离子的浓度。
()6.组成酶是细胞中含量较为稳定的酶。
()四、问答题:1.为什么说三羧酸循环是糖、脂和蛋白质三大物质代谢的共同通路?2.简单举例说明生物体代谢调控的三个水平。
3.用实例说明能荷调节的重要性。
4.举例说明原核生物基因表达的调节。
5.何谓酶的共价修饰(或化学修饰)?举例说明通过共价修饰来调节酶的活性。
6.何谓反馈调节?可分为哪些类型?7.举例说明什么叫级联放大作用。
8.哪些化合物可以被认为是联系糖、脂、蛋白质和核酸代谢的重要环节?为什么?五、名词解释:酶的级联系统酶的共价修饰反馈抑制操纵子第二信使诱导酶组成酶阻遏物前馈激活CaM参考答案:第十三章代谢调节一、填空题:1.生物体内的代谢调节在三种不同的水平上进行,即分子水平、细胞水平和多细胞整体水平。
2.代谢途径的终产物浓度可以控制自身形成的速度,这种现象被称为反馈。
3.酶对细胞代谢的调节是最基本的代谢调节,主要有二种方式:酶活性的调节和酶量的调节。
构通糖、脂代谢的关键化合物是乙酰CoA 。
4.不同代谢途径可以通过交叉点代谢中间物进行转化,在糖、脂、蛋白质及核酸的相互转化过程中三个最关键的代谢中间物是乙酰CoA 、丙酮酸和6-磷酸-葡萄糖。
5.1961年,法国生物学家Monod和Jacob提出了关于原核生物基因结构及表达调控的操纵子学说。
6.正调控和负调控是基因表达的两种最基本的调节形式,其中原核细胞常用负调控,而真核细胞常用正调控模式。
7.乳糖操纵子的天然诱导物是别乳糖,实验室里常用IPTG 作为乳糖操纵子的安慰诱导物诱导β-半乳糖苷酶的产生。
8.许多代谢途径的第一个酶是限速酶,终产物多是它的别构抑制剂,对它进行反馈抑制,底物多为其别构激活剂。
9.原核细胞酶的合成速率主要在转录水平进行调节。
10.乳糖操纵子的诱导物是乳糖,色氨酸操纵子的辅阻遏物是色氨酸。
二、选择题(只有一个最佳答案):1.下列与能量代谢有关的过程除哪个外都发生在线粒体中?( A )A、糖酵解B、三羧酸循环C、脂肪酸的β-氧化D、氧化磷酸化2.IPTG可以诱导乳糖操纵子(lacOperon)的表达,这是因为:( B )A、IPTG与乳糖操作子(lacoperator)结合,诱导转录B、IPTG与LACI基因产物结合,并抑制其活性C、抑制β-半乳糖苷酶的活性D、促进Lac阻遏物的活性E、IPTG与LACI基因产物结合,并激活其活性3.在什么情况下,乳糖操纵子的转录活性最高?( A )A、高乳糖,低葡萄糖B、高乳糖,高葡萄糖C、低乳糖,低葡萄糖D、低乳糖,高葡萄糖4.真核细胞参与基因表达调节的调控区比原核细胞复杂是因为( D )A、真核细胞的细胞核具有双层膜B、原核细胞的基因总是以操纵子的形式存在C、原核细胞调节基因表达主要是在翻译水平D、真核细胞需要控制细胞特异性的基因表达E、真核细胞基因组含有太多的重复序列5.调节物质代谢体内最基础的层次是( A )A、细胞水平B、激素水平C、神经调节D、整体水平E、器官水平6.磷酸果糖激酶是什么代谢途径中的别构调节酶( C )A、三羧酸循环B、糖异生C、葡萄糖分解D、糖原合成E、糖原分解7.三羧酸循环中的别构调节酶是( A )A、柠檬酸合成酶B、α-酮戊二酸脱氢酶C、琥珀酸脱氢酶D、延胡索酸酶E、苹果酸脱氢酶8.催化糖酵解与磷酸戊糖途径的酶主要分布在细胞中什么部位( B )A、核B、胞质C、线粒体D、微粒体E、质膜9.催化三羧酸循环与脂肪酸β-氧化的酶分布在细胞内的什么部位( E )A、胞质B、胞膜C、胞核D、内质网E、线粒体10.氨基酸分解代谢调节的别构酶是( E )A、转氨酶B、脱羧酶C、转甲基酶D、己糖激酶E、谷氨酸脱氨酶11.糖异生限速酶的别构调节激活剂是( A )A、ATPB、ADPC、AMPD、dATPE、cAMP 12.各种分解途径中,放能最多的途径是:( B )A、糖酵解B、三羧酸循环C、 —氧化D、氧化脱氨基13.操纵子调节系统属于哪一种水平的调节?( B )A、复制水平的调节B、转录水平的调节C、转录后加工的调节D、翻译水平的调节14.下列关于操纵基因的论述哪个是正确的?( A )A、能专一性地与阻遏蛋白结合B、是RNA聚合酶识别和结合的部位C、是诱导物和辅阻遏物的结合部位D、能于结构基因一起转录但未被翻译15.以下有关阻遏蛋白的论述哪个是正确的?( A )A、阻遏蛋白是调节基因表达的产物B、阻遏蛋白妨碍RNA聚合酶与启动子结合C、阻遏蛋白RNA聚合酶结合而抑制转录D、阻遏蛋白与启动子结合而阻碍转录的启动16.糖酵解中,下列哪一个催化的反应不是限速反应?( D )A、丙酮酸激酶B、磷酸果糖激酶C、己糖激酶D、磷酸丙糖异构酶17.磷酸化酶通过接受或脱去磷酸基而调节活性,因此它属于:( B )A、别(变)构调节酶B、共价调节酶C、诱导酶D、同工酶18.下列与能量代谢有关的途径不在线粒体内进行的是:(D)A、三羧酸循环B、脂肪酸β氧化C、氧化磷酸化D、糖酵解作用19.关于共价修饰调节酶,下列哪种说法是错误的?(D)A、这类酶一般存在活性和无活性两种形式,B、酶的这两种形式通过酶促的共价修饰相互转变C、伴有级联放大作用D、是高等生物独有的代谢调节方式20.阻遏蛋白结合的位点是:(C)A、调节基因B、启动因子C、操纵基因D、结构基因21.下面哪一项代谢是在细胞质内进行的:(C)A、脂肪酸的β-氧化B、氧化磷酸化C、脂肪酸的合成D、TCA22.在乳糖操纵子模型中,操纵基因专门控制是否转录与翻译。