第五讲 蛋白质合成_PPT幻灯片
合集下载
蛋白质合成及转运生科课件.ppt

●胰岛素原的加工:
间插序列(C肽区)
HS SH
HS SH HS
C A链区
B链区
SH
核糖体上合成出无规 则卷曲的前胰岛素原
切除信号肽后
折叠成稳定构
信号肽
象的胰岛素原
N
N
S-S
C
S
S
S
S
胰岛素原
切除C肽后,形成 成熟的胰岛素分子
N
S S N
A链 C
S
C B链
S
胰岛素
Thanks
6、蛋白因子帮助合成的起始:
● 蛋白质合成的起始、延伸和终止的每一个阶段,都涉及到一组不同 的蛋白质因子的帮助.
● 原核生物(大肠杆菌): 三个起始因子(initiation factor):IF1、IF2、IF3 真核生物:更多种的起始因子帮助.
①IF1、IF3与30S小亚基结合:
IF3防止30S亚基与50S 亚基过早结合.
● 真核生物最靠近5’端的AUG序列通常就是起始密码.
● 原核生物mRNA 5’端的SD序列—识别16S rRNA
SD序列:在细菌的mRNA的5’端起始AUG序列上游10个碱基左右的位置, 有一段富含嘌呤碱基的序列,能与细菌的16S核糖体RNA3’端的7个嘧啶 碱基互补性识别,这段序列由Shine-Dalgarno发现,称为S-D序列.
● 由同一种tRNA合成酶合成:起始因子识别tRNAiMet
延伸因子识别tRNAMet
● 原核生物中的第一个蛋氨酸要进行甲酰化 修饰---甲酰Met:
fMet - tRNAiMet
5、翻译起始于mRNA与核糖体的结合:
● 真核生物mRNA分子的5’端有核糖体进入部位: 帽子结构帮助识别mRNA分子与核糖体的结合位点. 核糖体沿着 mRNA分子5’ → 3’扫描至起始密码AUG.
《蛋白质的合成》PPT课件

②
Met Met
③
Met-tRNAiMet-elF-2 -GTP
mRNA
ATP elF4E, elF4G, elF4A, elF4B,PAB
ADP+Pi
60S
① eIF-2B、eIF-3、
eIF-6
40S
Met
elF-5
④
各种elF释放 GDP+Pi
60S
Met
精选课件ppt
真核生物翻译起始 复合物形成过5程3
3'非 翻 译 区
11
遗
传
密
密
码
码 子
表的
第
一
个
字
母
密码子的第二个字母
精选课件ppt
12
遗传密码的特点
1. 连续性(commaless)
编码蛋白质氨基酸序列的各个三联 体密码连续阅读,密码间既无间隔也 无重叠。
精选课件ppt
13
重叠密码
非重叠连续的密码
不连续的密码
精选课件ppt
14
基因损伤引起mRNA阅读框架内的碱 基发生插入或缺失,可能导致框移突变 (frameshift mutation)。
21
摆动配对
U
精选课件ppt
22
精选课件ppt
23
二、核蛋白体是多肽链合成的装置
精选课件ppt
24
核蛋白体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基
大亚基
核蛋白 体
21种 30S 16S 34种 50S 23S
5S 70S
精选课件ppt
33种 49种
蛋白质的生物合成PPT课件

第一步
氨PPi
基
E-AMP
酸
的 氨酰腺苷酸
活 第二步
AMP
E化
AA
E
tRNA
AA
E
AA
E
tRNA
AA
3-氨酰-tRNA
tRNA
E
活化反应方程式:
氨基酸 + ATP
酶/ Mg2+
氨酰AMP-酶 + PPI
氨酰AMP-酶
氨酰tRNA + AMP + 酶
tRNA
一个氨基酸活化需要消耗2个高能磷酸键
氨酰- tRNA合成酶特点 专一性:对氨基酸有极高的专一性,每种
中心法则总结了生物体内遗传信息的流动规律,揭示遗传的分 子基础,不仅使人们对细胞的生长、发育、遗传、变异等生 命现象有了更深刻的认识,而且以这方面的理论和技术为基 础发展了基因工程,给人类的生产和生活带来了深刻的革命 。
遗 DNA
传
信
息
流
mRNA
动
示
核糖体
意
图
tRNA
第一节 RNA在蛋白质生物 合成中的重要功能
tRNA的功能
(一)被特定的氨酰- tRNA合成酶识别, 使tRNA接受正确的活化氨基酸。
(二)识别mRNA链上的密码子。
(三)在蛋白质合成过程中,tRNA起着 连结生长的多肽链与核糖体的作用。
(一)、接受正确的活化氨基酸
氨基酸 + ATP
酶/ Mg2+
氨酰AMP-酶 + PPi
氨酰AMP-酶
tRNA
合成蛋白质 ③ 被蛋白质合成的起始因子所识别,从
而促进蛋白质的合成。
AAAAAAA-OH
蛋白质生物合成PPT课件演示教学.ppt

缬 脯 苏 天冬
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
蛋白质的生物合成(共106张PPT)

现 )和真菌中发现UAG可能是编码第22种氨基酸 pyrrolysine(吡咯赖氨酸)的密码子。
遗传密码动画
2022/9/17
12
遗传密码的特点
1.方向性(direction) 翻译时的阅读方向只能是5→3,即读
码从mRNA的起始密码子AUG开始,按 5→3的方向逐一阅读,直至终止密码子。
2022/9/17
为遗传密码(也称密码子)。 mRNA在核糖体小亚基就位;
-Ser-Lys-Leu-(PST序列) 顺反子(cistron):遗传学将编码一个多肽的遗传单位称为顺反子。 氨基酰-tRNA合成酶
• 开放阅读框架(open reading frame,ORF):从 开放阅读框区(open reading frame, ORF)
AMP-E或氨基酰-tRNA的酯键水解,再换上与密码子相对应
的氨基酸。
• 氨基酰-tRNA的表示方法:
• Ala-tRNAAla、Ser-tRNASer、Met-tRNAMet
氨基酸的活化形式:氨基酰-tRNA 氨基酸的活化部位:α-羧基 氨基酸与tRNA连接方式:酯键 氨基酸活化耗能:2个~P
2022/9/17
23
核糖体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基 21种 30S 16S 33种 40S 18S
大亚基 36种 50S 23S 5S
49种
28S 60S 5.8S
5S
核蛋白体
70S
80S
2022/9/17
24
30S小亚基:有mRNA结合位点 50S大亚基: E位:排出位(Exit site)
• 氨基酰-tRNA合成酶:存在于胞液中,催化氨基酸的 活化。
遗传密码动画
2022/9/17
12
遗传密码的特点
1.方向性(direction) 翻译时的阅读方向只能是5→3,即读
码从mRNA的起始密码子AUG开始,按 5→3的方向逐一阅读,直至终止密码子。
2022/9/17
为遗传密码(也称密码子)。 mRNA在核糖体小亚基就位;
-Ser-Lys-Leu-(PST序列) 顺反子(cistron):遗传学将编码一个多肽的遗传单位称为顺反子。 氨基酰-tRNA合成酶
• 开放阅读框架(open reading frame,ORF):从 开放阅读框区(open reading frame, ORF)
AMP-E或氨基酰-tRNA的酯键水解,再换上与密码子相对应
的氨基酸。
• 氨基酰-tRNA的表示方法:
• Ala-tRNAAla、Ser-tRNASer、Met-tRNAMet
氨基酸的活化形式:氨基酰-tRNA 氨基酸的活化部位:α-羧基 氨基酸与tRNA连接方式:酯键 氨基酸活化耗能:2个~P
2022/9/17
23
核糖体的组成
核蛋
原核生物
真核生物
白体 蛋白质 S值 rRNA 蛋白质 S值 rRNA
小亚基 21种 30S 16S 33种 40S 18S
大亚基 36种 50S 23S 5S
49种
28S 60S 5.8S
5S
核蛋白体
70S
80S
2022/9/17
24
30S小亚基:有mRNA结合位点 50S大亚基: E位:排出位(Exit site)
• 氨基酰-tRNA合成酶:存在于胞液中,催化氨基酸的 活化。
蛋白质的生物合成课件.ppt

如何转变?
密码子:mRNA分子中,每三个相邻的核苷酸组
成的三联体代表某一种氨基酸或其它信息,称为 密码子或三联密码.一个氨基酸密码子决定着一 个氨基酸。
遗传密码:mRNA中的核苷酸排列序列与蛋白
质中的氨基酸排列序列的关系。生物的遗传密码
是通用的。
四种核苷酸编成三联体可形成 43个即64个密码子.其中: 1.一个起始密码:AUG
小亚基:沿mRNA结合,沿5’ 3’ 方向移动.
大亚基:受位(A位): 结合氨基酰- tRNA
给位(P位):成肽
给位 (P位)
蛋 苏
大亚基
UGU
5’AUG ACA GUU
受位 (A位)
小亚基
3’
蛋白质生物合成过程
1、准备阶段: 氨基酸的活化与转运。
2、中心环节: 核蛋白体循环-活化氨基酸 在核蛋白体上的缩合组装。
氨基酸的活化与转运
1、反应式:
氨基酰-tRNA合成酶
AA + tRNA + ATP
氨基酰-tRNA+AMP+PPi 2、AA结合位置:
AA的α-羧基与tRNA活末端腺苷酸中 核糖2 ’或3’羟基以酯键相结合。
tRNA-CCA-OH(R-3’-OH)
核蛋白体循环(三阶段)
(1)、起始阶段 (2)、延伸阶段 (3)、终止阶段
基因操纵子调节系统示意图
调节基因 转录
操纵子
控制区
信息区
启动基因 操纵基因 RNA聚合酶
结构基因
DNA
(-)
(+) 转录
翻译
mRNA
阻遏蛋白
诱导剂
翻译 蛋白质
血红素对起始因子-2的调节作用
血红素
密码子:mRNA分子中,每三个相邻的核苷酸组
成的三联体代表某一种氨基酸或其它信息,称为 密码子或三联密码.一个氨基酸密码子决定着一 个氨基酸。
遗传密码:mRNA中的核苷酸排列序列与蛋白
质中的氨基酸排列序列的关系。生物的遗传密码
是通用的。
四种核苷酸编成三联体可形成 43个即64个密码子.其中: 1.一个起始密码:AUG
小亚基:沿mRNA结合,沿5’ 3’ 方向移动.
大亚基:受位(A位): 结合氨基酰- tRNA
给位(P位):成肽
给位 (P位)
蛋 苏
大亚基
UGU
5’AUG ACA GUU
受位 (A位)
小亚基
3’
蛋白质生物合成过程
1、准备阶段: 氨基酸的活化与转运。
2、中心环节: 核蛋白体循环-活化氨基酸 在核蛋白体上的缩合组装。
氨基酸的活化与转运
1、反应式:
氨基酰-tRNA合成酶
AA + tRNA + ATP
氨基酰-tRNA+AMP+PPi 2、AA结合位置:
AA的α-羧基与tRNA活末端腺苷酸中 核糖2 ’或3’羟基以酯键相结合。
tRNA-CCA-OH(R-3’-OH)
核蛋白体循环(三阶段)
(1)、起始阶段 (2)、延伸阶段 (3)、终止阶段
基因操纵子调节系统示意图
调节基因 转录
操纵子
控制区
信息区
启动基因 操纵基因 RNA聚合酶
结构基因
DNA
(-)
(+) 转录
翻译
mRNA
阻遏蛋白
诱导剂
翻译 蛋白质
血红素对起始因子-2的调节作用
血红素
蛋白质合成PPT

原核细胞70S核糖体的A位、P位及 mRNA结合部位示意图
P 位(结
合或接受肽 基的部位)
A位(结合或
50S
接受AA- tRNA 的部位)
5
3
mRNA
30S
与mRNA结合部位
anticodon
一个 核糖体中占有2个tRNA和40bp长的mRNA
Codon 密码子
4、辅助性蛋白
在蛋白质的合成体系中,除了mRNA、 tRNA、rRNA和核糖体外,还需要一系 列辅助因子,这些因子都是蛋白质。
多种蛋白质结合而成的一种大的核糖核蛋 白颗粒,蛋白质肽键的合成就是在这种核 糖体上进行的。
两类核蛋白体: 结合型:位于粗面内质网,合成分泌蛋白(含信号肽). 游离型:游离于胞质中,参与细胞固有蛋白的合成
核蛋白体
(1)组成:蛋白质+r RNA 真核生物核蛋白体(40S+60S=80S) 原核生物核蛋白体(30S+50S=70S) ( 2)功能区 mRNA结合部位:位于小亚基,序列特异识别 受位(A位):位于大、小亚基结合处,结合AAtRNA; 给位(P位):主要位于大亚基,结合肽酰tRNA和起始Met-tRNA,肽基转移部位; 转肽酶中心、GTP:位于大亚基,形成肽键的部 位。
34 protein
5S rRNA 50 S 大 亚 基
有一段能与tRNA Met序列 互补的片段
23S rRNA
23S rRNA靠近5‘端有一段12个核苷酸的序列与 5S rRNA上的序列互补。
核蛋白体(原核生物)
34种蛋白质 21种蛋白质 50
多肽链
30
核蛋白体(原核生物)
多核糖体:一个mRNA分子与一定数目的 单个核糖体结合形成念珠状,每个核糖体 可以独立完成一条肽链合成。
蛋白质的生物合成(共74张PPT)

3)校正tRNA
tRNASu3+Tyr反密码子为CUA,能识别变异 产生的终止密码子UAG。
三、氨酰-tRNA合成酶
❖ 氨基酸在掺入肽链前,必须活化生成氨酰tRNA,获得足够的能量。
❖ 活化反应由各种氨酰-tRNA合成酶(AARS) 催化。
A.氨酰-tRNA
氨基酰-tRNA合成酶
氨基酸 + tRNA
❖ 包括:氨酰-tRNA与核糖体结合,转肽 与肽键形成和转位三个步骤。
肽链合成的延长因子
(一) 进位
Tu GTsTP
Tu GDP
Ts
GTP
5'
AUG
3'
目录
Ts 移走GDP,并与Tu 结合生成Tu-Ts复合体, 然后GTP替换Ts,生 成有活性的Tu-GTP
Poly(U) Poly(C) Poly(A) Poly(G)
poly(Phe) peptide
无细胞抽取物
poly(Pro) peptide poly(Lys) peptide poly(Gly) peptide
•核糖体 •各种tRNA •氨基酸 •AARS
•ATP, GTP
+ mRNA = 蛋白质
氨基酰- tRNA
ATP
AMP+PPi
B. 氨酰-tRNA合成酶特点
a、专一性:
•对氨基酸有极高的专一性,每种氨基酸都有专 一的酶,只作用于L-氨基酸,不作用于D-氨基酸。
•对tRNA具有极高专一性:一种AARS只能识别装
载同一种氨基酸的一组同工受体(tRNA分子)
b、校对作用:
• AARS具有底物结合位点(包括tRNA、氨 基酸和ATP结合位点)和水解位点。
5'
tRNASu3+Tyr反密码子为CUA,能识别变异 产生的终止密码子UAG。
三、氨酰-tRNA合成酶
❖ 氨基酸在掺入肽链前,必须活化生成氨酰tRNA,获得足够的能量。
❖ 活化反应由各种氨酰-tRNA合成酶(AARS) 催化。
A.氨酰-tRNA
氨基酰-tRNA合成酶
氨基酸 + tRNA
❖ 包括:氨酰-tRNA与核糖体结合,转肽 与肽键形成和转位三个步骤。
肽链合成的延长因子
(一) 进位
Tu GTsTP
Tu GDP
Ts
GTP
5'
AUG
3'
目录
Ts 移走GDP,并与Tu 结合生成Tu-Ts复合体, 然后GTP替换Ts,生 成有活性的Tu-GTP
Poly(U) Poly(C) Poly(A) Poly(G)
poly(Phe) peptide
无细胞抽取物
poly(Pro) peptide poly(Lys) peptide poly(Gly) peptide
•核糖体 •各种tRNA •氨基酸 •AARS
•ATP, GTP
+ mRNA = 蛋白质
氨基酰- tRNA
ATP
AMP+PPi
B. 氨酰-tRNA合成酶特点
a、专一性:
•对氨基酸有极高的专一性,每种氨基酸都有专 一的酶,只作用于L-氨基酸,不作用于D-氨基酸。
•对tRNA具有极高专一性:一种AARS只能识别装
载同一种氨基酸的一组同工受体(tRNA分子)
b、校对作用:
• AARS具有底物结合位点(包括tRNA、氨 基酸和ATP结合位点)和水解位点。
5'
分子生物学--蛋白质的生物合成

Protein Biosynthesis
蛋白质的生物合成
完整版课件ppt
1
主要内容
概述
遗传密码
tRNA
核糖体
蛋白质的生物合成过程
一般特征
原核生物
真核生物
蛋白质的运输和翻译后修饰
完整版课件ppt
2
二、蛋白质的生物合成过程
完整版课件ppt
3
(一)蛋白质合成的一般特征
三种RNA:mRNA、tRNA、rRNA 原料:20种aa 需要酶及众多蛋白因子、ATP、GTP等 蛋白质合成方向:N端→C端 mRNA的翻译方向:5ˊ→3ˊ
需GTP和两个延伸因子 EFTu和EFTs
完整版课件ppt
24
延长因子EFTu和EFTs催化进位
完整版课件ppt
25
完整版课件ppt
26
2.成键:肽酰基从P位点转位到A位点,同时形成肽键。 由肽酰转移酶催化
完整版课件ppt
27
完整版课件ppt
28
3.移位
核糖体沿mRNA的5ˊ→3ˊ方向移动一个密码子的距离。 肽酰tRNA由A位移到P位 空载的tRNA进入 E位。 完整版需课件GppTt P和延伸因子G(EFG29 )
❖AUG,上游无SD序列,通常mRNA上最靠近5’ 端的AUG为起始部位。 ❖40S核糖体与mRNA 5’端帽子结合,向3’方向移 动,搜索AUG密码子,这过程要消耗ATP。
完整版课件ppt
37
完整版课件ppt
38
4.起始因子
命名为eIF-n eIF-3:与40S亚基结合,使大小亚基解离; eIF-2:与Met-tRNAiMet及GTP结合; eIF-4:与mRNA的帽子结合。 eIF-5:释放起始因子,使核糖体亚基结合
蛋白质的生物合成
完整版课件ppt
1
主要内容
概述
遗传密码
tRNA
核糖体
蛋白质的生物合成过程
一般特征
原核生物
真核生物
蛋白质的运输和翻译后修饰
完整版课件ppt
2
二、蛋白质的生物合成过程
完整版课件ppt
3
(一)蛋白质合成的一般特征
三种RNA:mRNA、tRNA、rRNA 原料:20种aa 需要酶及众多蛋白因子、ATP、GTP等 蛋白质合成方向:N端→C端 mRNA的翻译方向:5ˊ→3ˊ
需GTP和两个延伸因子 EFTu和EFTs
完整版课件ppt
24
延长因子EFTu和EFTs催化进位
完整版课件ppt
25
完整版课件ppt
26
2.成键:肽酰基从P位点转位到A位点,同时形成肽键。 由肽酰转移酶催化
完整版课件ppt
27
完整版课件ppt
28
3.移位
核糖体沿mRNA的5ˊ→3ˊ方向移动一个密码子的距离。 肽酰tRNA由A位移到P位 空载的tRNA进入 E位。 完整版需课件GppTt P和延伸因子G(EFG29 )
❖AUG,上游无SD序列,通常mRNA上最靠近5’ 端的AUG为起始部位。 ❖40S核糖体与mRNA 5’端帽子结合,向3’方向移 动,搜索AUG密码子,这过程要消耗ATP。
完整版课件ppt
37
完整版课件ppt
38
4.起始因子
命名为eIF-n eIF-3:与40S亚基结合,使大小亚基解离; eIF-2:与Met-tRNAiMet及GTP结合; eIF-4:与mRNA的帽子结合。 eIF-5:释放起始因子,使核糖体亚基结合
蛋白质合成PPTPPT课件

蛋白质合成的细胞定位
总结词
蛋白质合成主要发生在细胞内的核糖体上,核糖体是 细胞内蛋白质合成的场所。
详细描述
核糖体是细胞内一种由RNA和蛋白质组成的颗粒状结 构,主要存在于细胞质中。核糖体在蛋白质合成过程中 起着至关重要的作用,它能够读取mRNA上的遗传信 息,将一个个氨基酸按照特定的顺序连接起来形成多肽 链。同时,核糖体还具有催化肽键形成的酶活性,促进 蛋白质合成的进行。除了核糖体外,细胞内还有其他一 些细胞器也参与了蛋白质的合成过程,如内质网、高尔 基体等。这些细胞器在蛋白质的修饰、加工和运输等方 面起着重要作用。
蛋白质合成PPT课件
目录
• 蛋白质合成简介 • 蛋白质合成的过程 • 蛋白质合成的调控 • 蛋白质合成与疾病的关系 • 研究展望
01
蛋白质合成简介
蛋白质合成的基本概念
总结词
蛋白质合成是指细胞内利用已有的小分子物质作为原料,通过一系列酶促反应将氨基酸 按照特定的顺序连接起来形成多肽链,进而形成具有特定结构和功能的蛋白质的过程。
翻译后加工与修饰
总结词
翻译后加工与修饰是蛋白质合成的重要环节,涉及多 种酶促反应和化学修饰。
详细描述
翻译后加工与修饰是蛋白质合成的最后阶段,涉及到 多种酶促反应和化学修饰。这些加工和修饰包括剪切 、磷酸化、糖基化、乙酰化等,有助于完善蛋白质的 结构和功能。这些加工和修饰过程通常在特定的细胞 器或细胞部位进行,需要特定的酶和化学环境的支持 。通过翻译后加工与修饰,蛋白质的结构和性质得以 最终确定,从而发挥其在细胞生命活动中的重要功能 。
04
蛋白质合成与疾病的关系
蛋白质合成异常与疾病的发生
癌症
蛋白质合成异常可能导 致细胞增殖失控,引发
基因指导蛋白质的合成(5)PPT

A.人的成熟红细胞 B.成熟的白细胞
C.卵细胞
D.受精卵
16
4.某基因中含有1200个碱基对,则由它控制合 成的含有两条肽链的蛋白质分子中最多含有肽 健的个数是 ( )
A.198个
B.398个
C.400个
D.798个
17
5.一条多肽链中有氨基酸1000个,则合成该肽链 的mRNA和基因分别至少有碱基多少个?
为 (B )
A.20个 C.60个
B.40个 D.200个
14
2、已知一段mRNA中含有30个碱基, 其中A和G有12个,转录该段mRNA的
DNA分子中应有C和T的个数是 ( D )
A.12个
B.24个
C.18个
D.30个
15
3、研究基因控制蛋白质的合成过程,最好
选择下列哪一项为实验材料( D )
密码子
密码子
密码子
U U A GAU AUC
mRNA
11
翻译模板
➢信息传递
DNA上的遗传信息 (脱氧核苷酸排序)
➢中心法则
转录 细胞核
mRNA(核糖核苷 酸排序)
翻译
蛋白质
(特定的氨基
细胞质 酸顺序)
遗传信息的传递只有这种模式吗?
转录 DNA
逆转录
➢基因对性状的控制
翻译
RNA
蛋白质
▪控制蛋白质分子的结构而直接影响性状
mrnarnatrnarnarrnarna整理课件规则的双螺旋结构通常呈单链结构脱氧核苷酸核糖核苷酸腺嘌呤a鸟嘌呤g腺嘌呤a鸟嘌呤g胞嘧啶c胸腺嘧啶t胞嘧啶c尿嘧啶u脱氧核糖核糖磷酸磷酸dna与rna分子的比较单击画面继续整理课件dna解旋以一条链为模板合成rna组成rna的核糖核苷酸一个个连接起来场所
《蛋白质的合成》课件

特异性。
翻译水平的调控
翻译起始
翻译后修饰
翻译起始是蛋白质合成的关键步骤之 一,涉及核糖体与mRNA的结合以及 起始密码子的识别。
翻译后的蛋白质还需要经过一系列的 修饰才能成为有功能的分子,如磷酸 化、乙酰化、糖基化等。
翻译延长与终止
在翻译过程中,核糖体的移位速度和 肽酰-tRNA的形成受到多种因素的调 节,如mRNA的结构、tRNA的浓度 和种类、核糖体的构象等。
04
CATALOGUE
蛋白质合成的应用
蛋白质工程
蛋白质工程是通过修改或设计蛋白质 的氨基酸序列,以达到改善蛋白质的 某些功能或创建新功能的技术。
蛋白质工程在医药、农业和工业领域 有广泛应用,例如设计新的药物、改 良酶的催化效率和稳定性、优化蛋白 质的表达和纯化等。
药物设计与开发
01
蛋白质合成在药物设计与开发中 发挥着关键作用,因为许多药物 是直接与蛋白质相互作用来发挥 其治疗作用的。
蛋白质合成的分子机制
mRNA在蛋白质合成中的作用
信息传递者
mRNA是DNA遗传信息的传递者 ,将遗传信息从DNA传递到核糖 体,用于指导蛋白质的合成。
遗传信息的解码
mRNA上的密码子与tRNA上的反 密码子互补配对,实现遗传信息 的解码,确保氨基酸按照正确的 顺序连接起来。
tRNA在蛋白质合成中的作用
氨基酸转运
tRNA作为氨基酸的转运载体,将氨基酸按照mRNA上的指令准确运送到核糖 体的A位点,参与蛋白质的合成。
反密码子识别
tRNA的反密码子能够与mRNA上的密码子进行互补配对,确保氨基酸按照正确 的顺序加入到多肽链中。
rRNA在蛋白质合成中的作用
核糖体的组成
rRNA是核糖体的主要组成部分,核 糖体是蛋白质合成的场所。
翻译水平的调控
翻译起始
翻译后修饰
翻译起始是蛋白质合成的关键步骤之 一,涉及核糖体与mRNA的结合以及 起始密码子的识别。
翻译后的蛋白质还需要经过一系列的 修饰才能成为有功能的分子,如磷酸 化、乙酰化、糖基化等。
翻译延长与终止
在翻译过程中,核糖体的移位速度和 肽酰-tRNA的形成受到多种因素的调 节,如mRNA的结构、tRNA的浓度 和种类、核糖体的构象等。
04
CATALOGUE
蛋白质合成的应用
蛋白质工程
蛋白质工程是通过修改或设计蛋白质 的氨基酸序列,以达到改善蛋白质的 某些功能或创建新功能的技术。
蛋白质工程在医药、农业和工业领域 有广泛应用,例如设计新的药物、改 良酶的催化效率和稳定性、优化蛋白 质的表达和纯化等。
药物设计与开发
01
蛋白质合成在药物设计与开发中 发挥着关键作用,因为许多药物 是直接与蛋白质相互作用来发挥 其治疗作用的。
蛋白质合成的分子机制
mRNA在蛋白质合成中的作用
信息传递者
mRNA是DNA遗传信息的传递者 ,将遗传信息从DNA传递到核糖 体,用于指导蛋白质的合成。
遗传信息的解码
mRNA上的密码子与tRNA上的反 密码子互补配对,实现遗传信息 的解码,确保氨基酸按照正确的 顺序连接起来。
tRNA在蛋白质合成中的作用
氨基酸转运
tRNA作为氨基酸的转运载体,将氨基酸按照mRNA上的指令准确运送到核糖 体的A位点,参与蛋白质的合成。
反密码子识别
tRNA的反密码子能够与mRNA上的密码子进行互补配对,确保氨基酸按照正确 的顺序加入到多肽链中。
rRNA在蛋白质合成中的作用
核糖体的组成
rRNA是核糖体的主要组成部分,核 糖体是蛋白质合成的场所。
第蛋白质的生物合成(共23张PPT)
在一条mRNA链上可结合多个核糖体,各自独立进 行多肽的合成,能提高mRNA的翻译效率。
6 多肽链的加工
1)N-端甲酰基及多余氨基酸的切除 按蛋白质合成机理,蛋白质N-端氨基酸应是甲酰蛋 氨酸(原核)或蛋氨酸(真核),但事实上并非如此。这是由 于脱甲酰酶除去了N-端的甲酰基,氨肽酶切除了N-端 一或几个多余氨基酸。此过程常在肽链延伸约有40个
氨基酸左右就开始了。
2)蛋白质内部某些氨基酸的修饰
被修饰的部位通常是丝氨酸或苏氨酸铡链上 的羟基;天冬氨酸、谷氨酸铡链上的羧基;天冬 酰胺铡链上的酰胺基;精氨酸、赖氨酸上的氨基; 半胱氨酸上的巯基等。修饰作用是在专一性的酶 催化下进行的。
3)切除非必需肽段 有些酶、激素等要经过此加工过程。如胃蛋白酶、 胰蛋白酶等,初合成的是没有活性的酶原,在一定条 件下除去一段肽才能转变为有活性的酶。如胰岛素, 初产物为前胰岛素原,经切除N-端信号肽变为胰岛素 原,再切除C肽成为有活性的胰岛素。
在结构上类似于遗传信息传递中的某些底物的物质可用于治疗肿瘤,如6-巯基嘌呤、5-氟尿嘧啶等碱基类似物可抑制DNA的复制。
此过程常在肽链延伸约有40个氨基酸左右就开始了。
tRNA合成酶的催化下先生成氨酰-AMP 逆转录病毒以RNA基因组合成cDNA时以tRNA为引物;
有些酶、激素等要经过此加工过程。
,再将活化的氨
2 核糖体
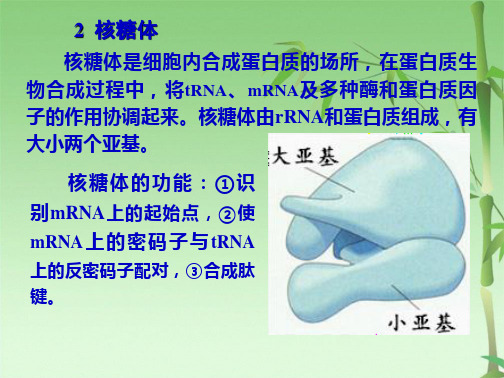
核糖体是细胞内合成蛋白质的场所,在蛋白质生 物合成过程中,将tRNA、mRNA及多种酶和蛋白质因 子的作用协调起来。核糖体由rRNA和蛋白质组成,有 大小两个亚基。
核糖体的功能:①识 别mRNA上的起始点,②使 mRNA 上 的 密 码 子 与 tRNA
上的反密码子配对,③合成肽 键。
核糖体的种类、结构和组分
6 多肽链的加工
1)N-端甲酰基及多余氨基酸的切除 按蛋白质合成机理,蛋白质N-端氨基酸应是甲酰蛋 氨酸(原核)或蛋氨酸(真核),但事实上并非如此。这是由 于脱甲酰酶除去了N-端的甲酰基,氨肽酶切除了N-端 一或几个多余氨基酸。此过程常在肽链延伸约有40个
氨基酸左右就开始了。
2)蛋白质内部某些氨基酸的修饰
被修饰的部位通常是丝氨酸或苏氨酸铡链上 的羟基;天冬氨酸、谷氨酸铡链上的羧基;天冬 酰胺铡链上的酰胺基;精氨酸、赖氨酸上的氨基; 半胱氨酸上的巯基等。修饰作用是在专一性的酶 催化下进行的。
3)切除非必需肽段 有些酶、激素等要经过此加工过程。如胃蛋白酶、 胰蛋白酶等,初合成的是没有活性的酶原,在一定条 件下除去一段肽才能转变为有活性的酶。如胰岛素, 初产物为前胰岛素原,经切除N-端信号肽变为胰岛素 原,再切除C肽成为有活性的胰岛素。
在结构上类似于遗传信息传递中的某些底物的物质可用于治疗肿瘤,如6-巯基嘌呤、5-氟尿嘧啶等碱基类似物可抑制DNA的复制。
此过程常在肽链延伸约有40个氨基酸左右就开始了。
tRNA合成酶的催化下先生成氨酰-AMP 逆转录病毒以RNA基因组合成cDNA时以tRNA为引物;
有些酶、激素等要经过此加工过程。
,再将活化的氨
2 核糖体
核糖体是细胞内合成蛋白质的场所,在蛋白质生 物合成过程中,将tRNA、mRNA及多种酶和蛋白质因 子的作用协调起来。核糖体由rRNA和蛋白质组成,有 大小两个亚基。
核糖体的功能:①识 别mRNA上的起始点,②使 mRNA 上 的 密 码 子 与 tRNA
上的反密码子配对,③合成肽 键。
核糖体的种类、结构和组分
《蛋白质合成》PPT课件

5
5′-末端 碱基
U C
A
G
中间 碱 基
U
苯丙氨酸 苯丙氨酸
亮氨酸 亮氨酸 亮氨酸 亮氨酸 亮氨酸 亮氨酸
异亮氨酸 异亮氨酸 异亮氨酸 甲硫氨酸、起始 甲酰甲硫氨酸
缬氨酸 缬氨酸 缬氨酸 缬氨酸
C
A
丝氨酸 丝氨酸 丝氨酸 丝氨酸
脯氨酸 脯氨酸 脯氨酸 脯氨酸
苏氨酸 苏氨酸 苏氨酸 苏氨酸
丙氨酸 丙氨酸 丙氨酸 丙氨酸
T2
GTP和 Mg2+
肽链的延长分四步进行:
1、进位
2、转肽
3、脱落
4、移位
精选课件ppt
24
1、进位
fMet-
下一个氨酰- tRNA结合
到70s核糖体的A位点上。
A A
~~~~~U~AC~~~~~~~
AA
新进入的氨酰- tRNA上的
AUG
反密码子必须与A位点上 mRNA的密码子互补。
AA Tu-GTP
专一性 主要决 定于第 一、二 碱基。
精选课件ppt
9
5)通用性
近于完全通用。
哺乳动物线粒体中,UGA不是终止密码子,而是色氨 酸的密码;AUA是甲硫氨酸的密码,而不是异亮氨酸 的密码,CUA是亮氨酸的密码,线粒体中却是苏氨酸 的密码。
精选课件ppt
10
(二)核糖体
核糖体是细胞质中球状颗粒,游离存在或结合在 内质网上。 1、核糖体是蛋白质合成的部位 2、核糖体的组成
这一过程需要延长因子Tu、
OOO
Ts参与,消耗一分子GTP
Pi OOO
Tu -GTP
fMet-
Tu -GDP AA
Ts GDP
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
根据摆动假说,在密码子与反密码子的配对中,前两 对严格遵守碱基配对原则,第三对碱基有一定的自由度, 可以摆动,因而使某些tRNA可以识别1个以上的密码子。
▪ 2)Francis Crick等人第一次证实只有用三联子密 码的形式才能把包含在由AUGC四个字母组成的遗 传信息(核酸)准确无误地翻译成由20种不同氨基 酸组成的蛋白质序列,实现遗传信息的表达。 实验1:
用吖啶类试剂(诱导核苷酸插入或丢失)处理T4噬菌体 rII位点上的两个基因,使之发生移码突变(frame-shift), 就生成完全不同的、没有功能的蛋白质。
基因表达包括转录(transcription)和翻译 (translation)两个阶段。转录是指拷贝出一条与DNA链 序列完全相同(除了T→U之外)的RNA单链的过程,是基 因表达的核心步骤。翻译是指以新生的mRNA为模板,把核 苷酸三联子遗传密码翻译成氨基酸序列、合成蛋白质多肽链 的过程,是基因表达的最终目的。
▪ 实验6:
以随机多聚物指导多肽合成。Nirenberg等及Ochoa
等又用各种随机的多聚物作模板合成多肽。例如,以只 含A、C的多聚核苷酸作模板,任意排列时可出现8种三 联子,即CCC、CCA、CAC、ACC、CAA、ACA、 AAC、AAA,获得由Asn、His、Pro、Gln、Thr、Lys 等6种氨基酸组成的多肽。
▪ (3)氨基酸的“活化”与核糖体结合技术。
如果把氨基酸与ATP和肝脏细胞质共培养,氨基酸就会 被固定在某些热稳定且可溶性RNA分子(transfer RNA, tRNA)上。现将氨基酸活化后的产物称为氨基酰-tRNA (aminoacyl-tRNA),并把催化该过程的酶称为氨基酰合 成酶(aminoacyl-tRNA Synthetase)。
实验2:
研究烟草坏死卫星病毒发现,其外壳蛋白亚基由400个氨 基酸组成,相应的RNA片段长1200个核苷酸,与密码三联 子体系正好相吻合。
▪ 实验3: 以均聚物为模板指导多肽的合成。 在含有tRNA、核糖
体、AA-tRNA合成酶及其它蛋白质因子的细胞抽提物中加 入mRNA或人工合成的均聚物作为模板以及ATP、GTP、氨 基酸等成分时又能合成新的肽链,新生肽链的氨基酸顺序由 外加的模板来决定。
(2)密码的兼并性
4种核苷酸组成61个编码氨基酸的密码子和3个终止 密码子(即UAA、UAG、UGA),终止密码不能与 tRNA的反密码子配对,但能被终止因子或释放因子识 别,终止肽链的合成。
由一种以上密码子编码同一个氨基酸的现象称为简 并(degeneracy),对应于同一氨基酸的密码子称为同 义密码子(synonymous codon)。
▪ 一. 基因与基因表达的一般概念 ▪ 二. 遗传密码——三联子 ▪ 三.tRNA ▪ 四.AA- tRNA合成酶 ▪ 五. 核糖体 ▪ 六. 信使核糖核酸 ▪ 七、蛋白质的生物合成 ▪ 八、氨基酸及功能蛋白质合成后的修饰 ▪ 九、蛋白质的运输和降解
一.基因与基因表达的一般概念
基因作为唯一能够自主复制、永久存在的单位,其生 理学功能以蛋白质形式得到表达。DNA序列是遗传信息的 贮存者,它通过自主复制得到永存,并通过转录生成 mRNA,翻译生成蛋白质的过程控制所有生命现象。
的核糖体经常与特异性氨基酰-tRNA相结合。如果把核糖 体与poly(U)和Phe-tRNA Phe共温育,核糖体就能同时 与poly(U)和Phe-tRNA Phe相结合。
ቤተ መጻሕፍቲ ባይዱ 5–1
2、遗传密码的性质
(1)每个AA密码子的数量。 Met, Trp 一个密码子。 Phe, Tyr, His, Gln, Glu, Asn, Asp, Lys, Cys, 2个密码子。 Ile, 3个密码子。 Val,Pro, Thr, Ala, Gly 4个密码子。 Leu, Arg, Ser 6个密码子。
▪
(3)普遍性与特殊性。 遗传密码无论在体内还是在体外,也无论对病毒、细菌、 动物还是植物而言都是适用的,所以,遗传密码具有普遍 性。但在支原体、线粒体中也存在例外。
(4)无间隔,无标点。
(5)遗传密码的摇摆性
蛋白质生物合成过程中,tRNA的反密码子通过碱基的 反向配对与mRNA的密码子相互作用。1966年,Crick根 据立体化学原理提出摆动假说(wobble hypothesis), 解释了反密码子中某些稀有成分如I以及许多有2个以上同 源密码子的配对问题。
多聚三核苷酸为模板时也可能只合成2种多肽: 5’…GUA GUA GUA GUA GUA…3’或 5’…UAG UAG UAG UAG UAG…3’或 5’…AGU AGU AGU AGU AGU…3’由第二种读码方式产 生的密码子UAG是终止密码,不编码任何氨基酸,因此, 只产生GUA(Val)或AGU(Ser)。
▪ 实验5:
以共聚三核苷酸作为模板可得到有3种氨基酸组成的多 肽。如以多聚(UUC)为模板,可能有3种起读方式:
5’…UUC UUC UUC UUC UUC…3’或 5’…UCU UCU UCU UCU UCU…3’或
5'…CUU CUU CUU CUU CUU…3‘ 分别产生UUC(Phe)、UCU(Ser)或CUU(Leu).
1961年,Nirenberg等以poly(U)作模板时发现合成 了多聚苯丙氨酸,从而推出UUU代表苯丙氨酸(Phe)。以 poly(C)及poly(A)做模板分别得到多聚脯氨酸和多聚 赖氨酸。
▪ 实验4: 以特定序列的共聚物为模板指导多肽的合成。以多聚二
核苷酸作模板可合成由2个氨基酸组成的多肽, 5'…UGU GUG UGU GUG UGU GUG…3',不管读码从U开 始还是从G开始,都只能有UGU(Cys)及GUG(Val)两 种密码子。
以人工合成的三核苷酸如UUU、UCU、UGU等为模板, 在含核糖体、AA-tRNA的反应液中保温后通过硝酸纤维素滤 膜,只有游离的AA-tRNA因相对分子质量小而通过滤膜,而 核糖体或与核糖体结合的AA-tRNA则留在滤膜上,这样可把 已结合与未结合的AA-tRNA分开。
当体系中带有多聚核苷酸模板时,从大肠杆菌中提取
▪ 2)Francis Crick等人第一次证实只有用三联子密 码的形式才能把包含在由AUGC四个字母组成的遗 传信息(核酸)准确无误地翻译成由20种不同氨基 酸组成的蛋白质序列,实现遗传信息的表达。 实验1:
用吖啶类试剂(诱导核苷酸插入或丢失)处理T4噬菌体 rII位点上的两个基因,使之发生移码突变(frame-shift), 就生成完全不同的、没有功能的蛋白质。
基因表达包括转录(transcription)和翻译 (translation)两个阶段。转录是指拷贝出一条与DNA链 序列完全相同(除了T→U之外)的RNA单链的过程,是基 因表达的核心步骤。翻译是指以新生的mRNA为模板,把核 苷酸三联子遗传密码翻译成氨基酸序列、合成蛋白质多肽链 的过程,是基因表达的最终目的。
▪ 实验6:
以随机多聚物指导多肽合成。Nirenberg等及Ochoa
等又用各种随机的多聚物作模板合成多肽。例如,以只 含A、C的多聚核苷酸作模板,任意排列时可出现8种三 联子,即CCC、CCA、CAC、ACC、CAA、ACA、 AAC、AAA,获得由Asn、His、Pro、Gln、Thr、Lys 等6种氨基酸组成的多肽。
▪ (3)氨基酸的“活化”与核糖体结合技术。
如果把氨基酸与ATP和肝脏细胞质共培养,氨基酸就会 被固定在某些热稳定且可溶性RNA分子(transfer RNA, tRNA)上。现将氨基酸活化后的产物称为氨基酰-tRNA (aminoacyl-tRNA),并把催化该过程的酶称为氨基酰合 成酶(aminoacyl-tRNA Synthetase)。
实验2:
研究烟草坏死卫星病毒发现,其外壳蛋白亚基由400个氨 基酸组成,相应的RNA片段长1200个核苷酸,与密码三联 子体系正好相吻合。
▪ 实验3: 以均聚物为模板指导多肽的合成。 在含有tRNA、核糖
体、AA-tRNA合成酶及其它蛋白质因子的细胞抽提物中加 入mRNA或人工合成的均聚物作为模板以及ATP、GTP、氨 基酸等成分时又能合成新的肽链,新生肽链的氨基酸顺序由 外加的模板来决定。
(2)密码的兼并性
4种核苷酸组成61个编码氨基酸的密码子和3个终止 密码子(即UAA、UAG、UGA),终止密码不能与 tRNA的反密码子配对,但能被终止因子或释放因子识 别,终止肽链的合成。
由一种以上密码子编码同一个氨基酸的现象称为简 并(degeneracy),对应于同一氨基酸的密码子称为同 义密码子(synonymous codon)。
▪ 一. 基因与基因表达的一般概念 ▪ 二. 遗传密码——三联子 ▪ 三.tRNA ▪ 四.AA- tRNA合成酶 ▪ 五. 核糖体 ▪ 六. 信使核糖核酸 ▪ 七、蛋白质的生物合成 ▪ 八、氨基酸及功能蛋白质合成后的修饰 ▪ 九、蛋白质的运输和降解
一.基因与基因表达的一般概念
基因作为唯一能够自主复制、永久存在的单位,其生 理学功能以蛋白质形式得到表达。DNA序列是遗传信息的 贮存者,它通过自主复制得到永存,并通过转录生成 mRNA,翻译生成蛋白质的过程控制所有生命现象。
的核糖体经常与特异性氨基酰-tRNA相结合。如果把核糖 体与poly(U)和Phe-tRNA Phe共温育,核糖体就能同时 与poly(U)和Phe-tRNA Phe相结合。
ቤተ መጻሕፍቲ ባይዱ 5–1
2、遗传密码的性质
(1)每个AA密码子的数量。 Met, Trp 一个密码子。 Phe, Tyr, His, Gln, Glu, Asn, Asp, Lys, Cys, 2个密码子。 Ile, 3个密码子。 Val,Pro, Thr, Ala, Gly 4个密码子。 Leu, Arg, Ser 6个密码子。
▪
(3)普遍性与特殊性。 遗传密码无论在体内还是在体外,也无论对病毒、细菌、 动物还是植物而言都是适用的,所以,遗传密码具有普遍 性。但在支原体、线粒体中也存在例外。
(4)无间隔,无标点。
(5)遗传密码的摇摆性
蛋白质生物合成过程中,tRNA的反密码子通过碱基的 反向配对与mRNA的密码子相互作用。1966年,Crick根 据立体化学原理提出摆动假说(wobble hypothesis), 解释了反密码子中某些稀有成分如I以及许多有2个以上同 源密码子的配对问题。
多聚三核苷酸为模板时也可能只合成2种多肽: 5’…GUA GUA GUA GUA GUA…3’或 5’…UAG UAG UAG UAG UAG…3’或 5’…AGU AGU AGU AGU AGU…3’由第二种读码方式产 生的密码子UAG是终止密码,不编码任何氨基酸,因此, 只产生GUA(Val)或AGU(Ser)。
▪ 实验5:
以共聚三核苷酸作为模板可得到有3种氨基酸组成的多 肽。如以多聚(UUC)为模板,可能有3种起读方式:
5’…UUC UUC UUC UUC UUC…3’或 5’…UCU UCU UCU UCU UCU…3’或
5'…CUU CUU CUU CUU CUU…3‘ 分别产生UUC(Phe)、UCU(Ser)或CUU(Leu).
1961年,Nirenberg等以poly(U)作模板时发现合成 了多聚苯丙氨酸,从而推出UUU代表苯丙氨酸(Phe)。以 poly(C)及poly(A)做模板分别得到多聚脯氨酸和多聚 赖氨酸。
▪ 实验4: 以特定序列的共聚物为模板指导多肽的合成。以多聚二
核苷酸作模板可合成由2个氨基酸组成的多肽, 5'…UGU GUG UGU GUG UGU GUG…3',不管读码从U开 始还是从G开始,都只能有UGU(Cys)及GUG(Val)两 种密码子。
以人工合成的三核苷酸如UUU、UCU、UGU等为模板, 在含核糖体、AA-tRNA的反应液中保温后通过硝酸纤维素滤 膜,只有游离的AA-tRNA因相对分子质量小而通过滤膜,而 核糖体或与核糖体结合的AA-tRNA则留在滤膜上,这样可把 已结合与未结合的AA-tRNA分开。
当体系中带有多聚核苷酸模板时,从大肠杆菌中提取