稻瘟病菌T-DNA插入方法优化及其突变体分析
水稻WRKY转录因子家族研究进展

水稻WRKY转录因子家族研究进展刘梦佳;李海峰【摘要】综述了水稻WRKY转录因子响应逆境胁迫、调控发育和代谢等方面的生物学功能,介绍了水稻WRKY基因表达模式和作用机制研究的新进展,并在功能解析方面做了展望。
%In this paper, we reviewed the functions of rice WRKY transcription factors on response to stress,regulation in developmental and metabolic process. Furthermore, progresses in expression pattern and molecular mechanism were introduced,and a short prospect in function analysis was made.【期刊名称】《河南农业科学》【年(卷),期】2016(045)003【总页数】8页(P1-8)【关键词】水稻;WRKY转录因子;生物胁迫;非生物胁迫;胁迫应答【作者】刘梦佳;李海峰【作者单位】西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌712100;西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌712100; 新疆农业职业技术学院,新疆昌吉831100【正文语种】中文【中图分类】S685水稻(Oryza sativa)WRKY转录因子作为植物转录因子家族的一个重要成员,通过保守的WRKY结构域特异性结合到启动子区域的W-box,直接或间接调控抗病、干旱、冷热、高盐、氧化损伤、衰老等相关基因的表达,参与水稻的生物、非生物胁迫反应、激素信号转导途经、发育和代谢过程。
近年来,水稻WRKY基因功能研究特别是在抵抗逆境方面的研究取得了很大进展。
基于此,综述了水稻WRKY转录因子响应逆境胁迫、调控发育和代谢等方面的生物学功能,介绍了水稻WRKY基因表达模式和作用机制研究的进展,并在功能解析方面做了展望。
T-DNA插入鉴定实验报告

T-DNA插入突变体的鉴定时明辉同组者:薛敏学号:201000220069摘要 Ti质粒是上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移进植物细胞,并插入植物染色体DNA中。
所以Ti质粒上的这一段能转移的DNA被叫做T-DNA。
将感兴趣的基因改造插入到T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化,得到含有突变的植株。
通过本实验,我们将学习如何用PCR的方法检测所得植株是否为T-DNA的插入突变体。
1.引言T-DNA作为一种实验常用的遗传转化方法,在插入突变过程中,插入到植物染色体上的位置是随机的。
如果T-DNA插入进某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
农杆菌Ti质粒转化植物细胞后,在获得的后代分离群体中,有T-DNA 插入的纯合突变体,杂合突变体,和野生型。
在突变体研究中,需要的材料是纯合突变体,所以必须从分离群体中将纯合突变体鉴定出来。
本次实验中,采用液CTAB(或者TSP法)提取拟南芥植株的DNA,然后PCR将所获DNA扩增,在之后采用琼脂糖凝胶电泳技术,分离处长度不一的DNA带,以确定样品是否为T-DNA插入突变纯和体。
PCR(Polymerase ChainReaction),即聚合酶链式反应是体外核算扩增技术,具有特异、敏感、产率高、快速、简便、重复性好、易自动化等突出优点;能在一个试管内将所要研究的目的基因或某一DNA片段于数小时内扩增至十万乃至百万倍,使肉眼能直接观察和判断。
(PCR基本原理如右图)DNA含有PO43-基团,在pH8.0 Buffer(本实验中为TAE)中带负电, 在电场中向正极移动。
自由电泳时,由于不同大小的DNA片段的电荷密度大致相同,各核酸分子难以分开;选用适当浓度的琼脂糖凝胶作为支持物,使之具备一定的孔径,即可发挥分子筛效应,使大小不同的核酸片段迁移率出现较大差异,达到分离的目的;同样条件对Marker电泳;起到鉴定的作用。
拟南芥T-DNA插入突变体的鉴定

拟南芥T-DNA插入突变体的鉴定09生工吴超 200900140129一、实验原理T-DNA插入法是反向遗传学研究的重要手段。
T-DNA是农杆菌的一个大质粒,长度在25kb左右。
野生型农杆菌的T-DNA上带有激素合成基因,感染植物后会导致植物细胞快速增殖形成愈伤组织,失去分化能力。
所以一般实验使用改造后的农杆菌——T-DNA中导入了卡那霉素抗性基因和抗除草剂基因。
因此在农杆菌感染植物后可用除草剂来筛选转化子。
在转化子培养到F2代出现分离后,就需要对其基因型进行鉴定。
T-DNA插入突变体鉴定方法主要有两种:三引物法和双引物法。
在本实验中使用三引物法。
三引物法的原理如图1所示,即采用三引物(LP、RP、BP)进行PCR扩增。
野生型植株目的基因的两条染色体上均未发生T-DNA插入,所以其PCR产物仅有1种,分子量约900bp(即从LP到RP);纯合突变体植株目的基因的两条染色体上均发生T-DNA插入,T-DNA本身的长度约为25kb,过长的模板会阻止目的基因特异性扩增产物的形成,所以也只能得到1种以BP与LP或RP为引物进行扩增的产物,分子量约为400-700bp;杂合突变体植株只在目的基因的一条染色体上发发生了T-DNA插入,所以PCR扩增后可同时得到两种产物。
上述3种情况的电泳结果差异明显,能有效区分不同基因型的植株。
此法优点是可同时鉴定出纯和突变体并确证T-DNA的插入情况。
图1 T-DNA插入示意图CATB,即十六烷基三甲基溴化铵,是一种离子型表面活性剂。
能溶解细胞膜和核膜蛋白,使核蛋白解聚,从而使DNA得以游离出来。
并且CATB可在高离子强度的溶液里与蛋白质和大多数多聚糖形成复合物进而形成沉淀,但不沉淀核酸。
本实验使用CATB抽提DNA。
聚合酶链式反应(Polymerase Chain Reaction,PCR)是体外核酸扩增技术。
它具有特异性高、敏感、产率高、快速、简便、重复性好、易自动化等突出优点;能在一个试管内将所要研究的目的基因或某一DNA片段于数小时内扩增至十万乃至几万倍,使肉眼能直接观察和判断。
模式植物拟南芥T-DNA插入突变体的PCR鉴定资料讲解

▪ 第一管:
▪
10xTaq buffer
min,再转移上清入新管。 5. 向上清液中加等体积的冷异丙醇,小心混匀。-20 ℃ 放置30 min,12000 rpm离心10 min,
弃上清。 6. 用70%乙醇洗涤沉淀一次,12000 rpm稍离心,弃上清。 7. 将沉淀在超净工作台上吹干,或空气中晾干。加50 μl TE (pH 8.0)放4 ℃缓慢溶解。 8. 电泳检测DNA的浓度和质量。
▪
81.8 g NaCl
▪
0.2%(V/V)β-巯基乙醇
▪注:先将除巯基乙醇之外的药品配好,高压蒸汽灭菌后加入巯基乙醇
▪酚/氯仿/异戊醇: 25:24:1
▪氯仿/异戊醇: 24:1
▪TE缓冲液:Tris-HCl (pH 8.0) 10 mmol/L,EDTA 1 mmol/L
▪3 M NaAc(pH 5.2)
左引物 LP: TCATCCACCATGGAAGAAAAG 右引物 RP:
TTGGATACGATGCGAGTAACC 中间引物LB: TGGTTCACGTAGTGGGCCATCG 用LP+RP产生大带:1107bp 用LB+RP产生小带:560-860bp
三、实验步骤
▪ 实验材料:拟南芥T-DNA插入突变体分 离群体30个株号的植株。
3. 用PCR方法鉴定T-DNA插入纯合突变体
▪ 农杆菌Ti质粒转化植物细胞后,在获得的后代分离群体中, 有T-DNA插入的纯合突变体,杂合突变体,和野生型。在 突变体研究中,需要的材料是纯合突变体,所以必须从分 离群体中将纯合突变体鉴定出来。
▪ PCR方法为鉴定纯合突变体提供了有利手段。它利用三 个引物LP、RP和BP,其中LP和RP是植物基因组上TDNA插入位点两测的引物,BP是T-DNA区段上的引物。 经过PCR,在野生型植株,LP和RP这对引物扩增出分子 量较大的产物(野生型基因,大带);在杂合突变体, LP和RP能扩增出分子量较大的产物(野生型基因,大 带),另外BP和RP还能扩增出分子量较小的产物(小 带);在纯合突变体,只有BP和RP能扩增出分子量较小 的产物(小带)。因此,利用以上三种引物做PCR,根 据扩增结果能够容易地从群体中区分出纯合突变体。
稻瘟菌基因的功能互补试验

稻瘟菌基因的功能互补试验随着几种模式生物,尤其是人的全基因组序列的完成,生命科学的研究进入了一个功能基因组学的时代。
人们已经不仅仅关心一个基因组中包含有多少基因,更重要的是包含有多少有功能的基因,各个基因的功能分别是什么。
稻瘟病以其在农业生产中所处的地位以及稻瘟菌的生物学特性,已作为研究植物与病原真菌相互作用的模式系统(Valent, 1990)。
同时稻瘟菌具有易于人工培养,已有高效率的人工转化体系,在实验室可进行有性杂交,菌丝体为单倍体等特点,是分子遗传研究的良好材料。
稻瘟菌的基因组一共包括七条染色体,全基因组的大小为38Mb,包括大约10000个基因,平均3-4kb就有一个基因。
由于稻瘟菌的菌丝体为单倍体,每一个基因的突变都能引起其相应生理生化功能的变化,因此当在此基因组中引入相应基因的野生型基因时,就能从功能上互补此基因的突变,恢复野生型的功能,即稻瘟菌基因的功能互补试验。
基因的功能互补试验一般包括以下几个步骤:首先,将基因的全长克隆至细菌载体上;然后,在此载体中连入一个真核生物的选择标记,一般为潮霉素抗性基因和新霉素抗性基因等,或基因的全长重新克隆至连有真核生物的选择标记细菌载体上;第三,选用合适的限制性内切酶将质粒切为线形;第四,利用REMI转化体系转化突变体;第五,挑取抗性转化子进行功能互补鉴定;最后,PCR鉴定或基因组Southern杂交鉴定。
以上所用的实验步骤是基于REMI转化的,目前稻瘟菌的转化还有农杆菌介导转化等方法,利用这种方法也能进行基因的功能互补试验,其鉴定的方法与上面一致,只是在载体构建时是将基因全长和真核生物选择标记连入农杆菌的T-DNA中。
基于REMI转化和农杆菌介导转化的方法都是比较好的用于基因功能互补试验的方法,但是这两种方法都有一个共同的弱点,就是在这两种方法中,所引入的DNA都是以随机插入的方式整合入稻瘟菌的基因组中,这样,就有可能导致另一个功能基因的失活,从而产生新的性状或者不能完全互补原来的基因突变。
实验五 拟南芥TDNA插入突变纯合体的鉴定

0.5 ml dNTP〔10 mmol/L〕
1 ml 046 5’引物〔10 mmol/L〕
1 ml 046 5’引物〔10 mmol/L〕
1 ml LBa 引物〔10 mmol/L〕
1 ml LBa 引物〔10 mmol/L〕
135号突变体的鉴定〔7-11组〕
30ml反响体系1:
30ml反响体系3:
实验五 拟南芥TDNA插入突变 纯合体的鉴定
实验目的
1.熟练掌握植物基因组DNA快速提取的方法; 2.掌握利用PCR方法鉴定拟南芥T-DNA插入突
变体的方法。
T-DNA插入鉴定的原理
突变体鉴定的步骤
1. T-DNA插入基因的基因组序列; 2. T-DNA插入方向确实定; 3. T-DNA插入位置确实定; 4. 引物设计; 5. PCR扩增; 6. 电泳检测。
条件下,离心5分钟; 〔6〕弃上清后,用70%乙醇润洗沉淀,并在室温下枯燥沉淀; 〔7〕100 ml TE溶解沉淀,将制备好的样品在4℃保存备用。
〔此方法提取的基因组DNA只适用于PCR的鉴定,不适合酶切和大片段基因的扩增〕
PCR鉴定
30ml反响体系: 2ml 植物基因组DNA样品 3ml 10×扩增缓冲液 ml Taqase 〔5U/ml〕 0.5 ml dNTP〔10 mmol/L〕 1ml 引物1〔10 mmol/L〕 1ml 引物2〔10 mmol/L〕
1ml 135 5’引物〔10 mmol/L〕
1ml 135 3’引物〔10 mmol/L〕
1ml 135 3’引物〔10 mmol/L〕
30ml反响体系2:
30ml反响体系4:
2ml 植物基因组DNA样品〔WT〕 2ml 植物基因组DNA样品〔111〕
T-DNA

T-DNA插入突变具有受体材料范围广、操作简便、转化效率高、插入拷贝数少,转化子稳定等优点,是目前标记和获得植物病原真菌功能基因组的最有效的途径之一。
T-DNA插入突变也叫T-DNA标签(tagging)、(Agrobacterium tumefaciens 一mediated transformation,ATMT),是根癌农杆菌介导的利用T-DNA作为插入突变原或分子标记,来分离或克隆因为插入而失活的基因并研究其基因功能的方法。
所谓的根癌农杆菌是指侵染植物根部的能诱发植物产生肿瘤的革兰氏阴性细菌,它们在自然状态下趋化性感染大多数双子叶植物的受伤部位,继而侵入根部细胞,最后在其中裂解,释放出Ti质粒,其上的T-DNA片段便会与植物细胞的核基因组整合,合成正常植株所没有的冠瘿碱(opines),破坏控制细胞分裂的激素调节系统,从而使它转为癌细胞并诱导其产生冠瘿瘤。
1974年,比利时根特大学遗传学实验室的Zaenen观察到了农杆菌中的巨型质粒,并首次命名为Ti质粒。
后来发现这个约200kb的环状Ti质粒,主要包括毒性区(Vir区)、转移DNA 区(T-DNA区)、接合转移区和复制起始区这四个区域。
它能从致病植株转移到无病的植株中,使正常植株致病,T-DNA转移要穿越植物和细菌的细胞壁,再经过其原生质膜,最后穿透核膜。
主要包括4个过程:1)农杆菌首先识别植物敏感细胞再进行吸附和对信号的感受;2)Ti质粒上vir基因的激活;3) T-DNA切割以及T-DNA复合物的形成;4)新形成的T-DNA复合物从农杆菌转移到植物细胞中;5)整合T-DNA到植物的核基因并开始表达。
整个过程中,T-DNA自身并不编码任何与转移及整合有关的酶。
ATMT技术正是利用这一特性来携带外来基因进入植物细胞。
由于T-DNA两端各有一段25 bp的相对保守的同向重复序列,即T-DNA 边界序列。
将要转入植物细胞核的外源目的基因就是通过质粒构建的方法连接于这两段DNA区域之间。
【国家自然科学基金】_t-dna插入突变_基金支持热词逐年推荐_【万方软件创新助手】_20140803
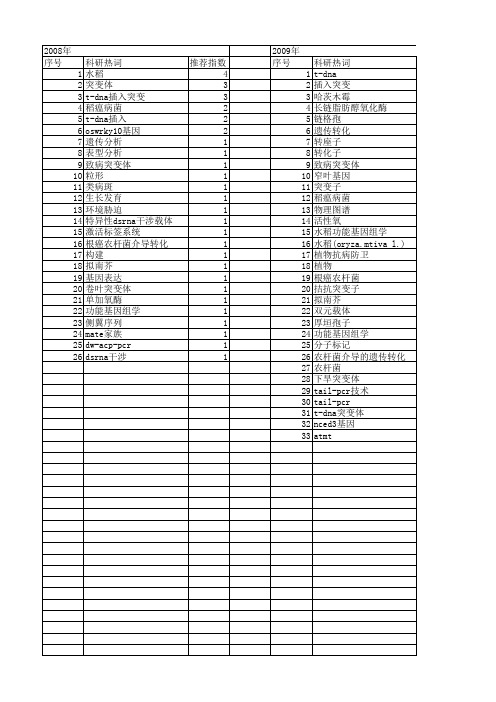
推荐指数 3 2 2 2 2 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13
2014年 科研热词 致病性 致病力 脱落酸 生物学性状 淡紫拟青霉 棉花黄萎病 插入突变 拟南芥 大丽轮枝菌 图位克隆 农杆菌介导的遗传转化 t-dna插入突变 asm1 推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1
推荐指数 4 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9
科研热词 雄性不育 花药发育 木霉 拟南芥 序列分析 图位克隆 tail-pcr t-dna插入突变体 ms1521
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32
科研热词 拟南芥 水稻 t-dna插入突变 雄性不育 鉴定 角果角度 花药发育 芥子油苷 绒毡层 突变表型 突变体库 生殖发育 玉米大斑病菌 漆酶 根癌农杆菌介导的转化 根发育 新型隐球酵母 反向遗传学 共分离 tos17突变系 tail-pcr t-dna突变系 t-dna插入缺失突变体 t-dna插入突变体 st273基因 spa基因 myb51 dna损伤修复 atmyb123 atmt atkor1 atccap
2011年 科研热词 推荐指数 水稻 4 拟南芥 4 遗传分析 3 短根毛突变体 2 基因定位 2 黄叶 1 遗传转化 1 转录因子 1 转基因 1 编码 1 红曲霉 1 红曲色素 1 精细定位 1 类病变突变体 1 筛选 1 突变俸库 1 突变体 1 病变组织 1 生物学机制 1 生物合成途径 1 水稻(oryza sativa l.) 1 氧化酶 1 桔霉素 1 根癌农杆菌 1 根形态 1 核基因控制 1 整合 1 插入突变 1 图位克隆 1 单萜合酶attps03 1 分子标记 1 农杆菌 1 侧翼序列 1 β -罗勒烯 1 tail-pcr 1 t-dna插入突变 1 t-dna 1 rei1 1 nadk3 1 gfp 1 cry1 1 bzip 1 abre元件 1 abl1 1 aba响应基因 1 aba信号传导 1
最新高考专题高考专题汇编【王梦新】【20180913】【高考专题生物 第7讲 孟德尔遗传定律】

专题7 孟德尔遗传定律2018高考真题(填空题30)(17分)水稻是我国最重要的粮食作物。
稻瘟病是由稻瘟病菌(Mp)侵染水稻引起的病害,严重危害我国粮食生产安全。
与使用农药相比,抗稻瘟病基因的利用是控制稻瘟病更加有效、安全和经济的措施。
(1)水稻对Mp表现出的抗病与感病为一对相对__________。
为判断某抗病水稻是否为纯合子,可通过观察自交子代____________来确定。
(2)现有甲(R1R1r2r2r3r3)、乙(r1r1R2R2r3r3)、丙(r1r1r2r2R3R3)三个水稻抗病品种,抗病(R)对感病(r)为显性,三对抗病基因位于不同染色体上。
根据基因的DNA序列设计特异性引物,用PCR方法可将样本中的R1、r1、R2、r2、R3、r3区分开。
这种方法可用于抗病品种选育中基因型的鉴定。
①甲品种与感病品种杂交后,对F2不同植株的R1、r1进行PCR扩增。
已知R1比r1片段短。
从扩增结果(下图)推测可抗病的植株有____________。
Array②为了在较短时间内将甲、乙、丙三个品种中的抗病基因整合,选育新的纯合抗病植株,下列育种步骤的正确排序是________。
a.甲×乙,得到F1b.用PCR方法选出R1R1R2R2R3R3植株c.R1r1R2r2r3r3植株×丙,得到不同基因型的子代d.用PCR方法选出R1r1R2r2R3r3植株,然后自交得到不同基因型的子代(3)研究发现,水稻的抗病表现不仅需要自身抗病基因(R1、R2、R3等)编码的蛋白,也需要Mp基因(A1、A2、A3等)编码的蛋白。
只有R蛋白与相应的A蛋白结合,抗病反应才能被激活。
若基因型为R1R1r2r2R3R3和r1r1R2R2R3R3的水稻,被基因型为a1a1A2A2a3a3的Mp侵染,推测这两种水稻的抗病性表现依次为___________。
(4)研究人员每年用Mp(A1A1a2a2a3a3)人工接种水稻品种甲(R1R1r2r2r3r3),几年后甲品种丧失了抗病性,检测水稻的基因未发现变异。
2022北京高三一模生物汇编:基因突变及其他变异

2022北京高三一模生物汇编基因突变及其他变异一、单选题1.(2022·北京延庆·一模)家族性高胆固醇血症是一种单基因遗传病(由A、a控制),该病在人群中的发病率没有明显的性别差异。
如图为某家族部分系谱图,其中已故男女的基因型及表现型无法获知,Ⅱ6不携带该病的致病基因。
下列分析错误..的是()A.该病属于常染色体上的显性遗传病B.Ⅱ5个体的基因型为AA或AaC.Ⅲ8和Ⅲ9再生的孩子一定患病D.Ⅱ4和Ⅲ10个体一定为杂合子2.(2022·北京延庆·一模)皱粒豌豆形成的分子机理如下图所示,相关分析正确的是()A.皱粒豌豆的形成是因为DNA中插入了一段外来DNA序列引发了染色体变异B.r基因转录的mRNA可能提前出现终止密码子,形成无活性的SBEI蛋白C.RR的植株与rr的植株杂交,F1表现为圆粒是由于r基因无法表达D.该图可以说明基因通过控制蛋白质的结构直接控制生物的性状3.(2022·北京延庆·一模)将相关内容依次填入下图各框中,其中包含关系错误..的选项是()A.A B.B C.C D.D4.(2022·北京海淀·一模)栽培品种香蕉染色体组成为AAA(字母代表染色体组),易患黄叶病。
野生蕉染色体组成为BB,含有纯合的抗黄叶病基因。
经过杂交、筛选,获得染色体组成为AAB的抗黄叶病香蕉新品种。
下列有关叙述,不正确...的是()A.栽培品种高度不育B.新品种含有三个染色体组C.新品种细胞中无同源染色体D.栽培品种减数分裂时联会紊乱5.(2022·北京丰台·一模)人类Y染色体末端的SRY基因能激活睾酮合成相关基因的表达睾酮在5α-还原酶的作用下转化为二氢睾酮,促使未分化的性腺发育成睾丸。
下列叙述不正确的是()A.SRY基因所在染色体片段缺失,后代可能出现XY的女性B.SRY基因所在染色体片段移接,后代可能出现XX的男性C.5α-还原酶基因缺陷的XY个体,雄性生殖器官发育不良D.XYY男性染色体异常是母亲减数分裂产生异常卵细胞所致6.(2022·北京丰台·一模)研究发现肿瘤细胞中谷氨酰胺合成酶(GS)高表达,GS通过推动有丝分裂中期到后期的转化促进细胞增殖。
稻瘟病(Magnaporthe grisea)研究进展图片

• 在多种杂草种病原M.grisea中说明与PWL2有同 源性的DNA序列的分布。
• 水稻的多数病菌和马唐属的病菌含有与PWL2 相似的同源序列。
• 稷属病菌同源性较少。
• 含有PWL1基因的病菌与PWL2有较少同源性片 断,这种同源性导致克隆PWL1基因的全部功 能。
•
• 第二个惰性基因,PWL3,来自同一稷属 病菌和另一个惰性基因PWL4来自弯叶画 眉草,都已克隆。克隆的PWL3基因编码 一个无活性的蛋白质,PWL4基因编码一 个活性的蛋白质,但不表达
四.寄主专化性的遗传控制
• 1. PWL寄主专化基因族 • 2.AVR2-YAMO无毒基因族
• 遗传分析鉴定2个基因,来自狗尾草病菌的 PWL1 , 来 自 水 稻 病 菌 的 PWL2 , 各 自 对 M.grisea菌系侵染弯叶画眉草能力起主要作用。
• 在水稻致病系统是典型的基因对基因系统,病 菌中的AVR基因与寄主中特定的抗病基因功能 性表现对应关系。
• 这种不稳定性也在减数分裂时发生。
• 在M.grisea变异潜在机制包括像B染色体, 准性循环,或在双链RNAS多态性。
• 在病菌中遗传不稳定看来与基因组不稳 定区频繁的正常发生的缺失和其他突变 有关。
• 在其它一些情况中,至少在实验室研究 中,转位也导致变异。
1.与染色体位置相关的不稳定 性
六.结论
• 插入诱变技术正在鉴定,致病性或孢子 形成的所要求的基因。
• 不同的cDNA 克隆技术在鉴定附着胞形成 期间所表达的一些基因或病菌在寄主植 物侵染性生长期表达的一些基因。
• 使用基因干扰技术突变分析已有可能鉴 定克隆的一些基因的功能。
• 主要的突破将来自发现单个AVR基因作用,例 如PWL2和AVR2-YAMO对病菌起什么作用。
农杆菌(Agrobacterium tumefaciens)介导的稻瘟病菌转化及致病突变体的筛选

浙江大学硕士学位论文农杆菌(Agrobacterium tumefaciens)介导的稻瘟病菌转化及致病突变体的筛选姓名:刘朋娟申请学位级别:硕士专业:植物病理学指导教师:王政逸20050501本研究受国家自然科学基金项目30270861资助单位代码:——究生学号:——硕士学位论文农杆菌(Agrobacteriumtumefaciens)介导的稻瘟病菌转化及致病突变体的筛选论文评阅人:.郄鏖压潘废垡蒸友基答辩委员会主席:.郑康乐答辩委员会成员:奎蕉夔筮回盐国昌三遮逸论文答辩日期:摘要稻瘟病菌(Magnaporthegrisea)可以引起水稻上的重要病害一稻瘟病。
了解彪grisea的致病分子机理不仅对防治稻瘟病具有重要的意义,而且它作为一个理想的研究体系对了解其他植物病原真菌与寄主的相互作用也有重要的意义。
分离和鉴定稻瘟病菌的致病相关基因,是达到这一目标的关键所在。
BNA插入突变是标记、分离功能基因的有效途径之一,ATMT是丝状真菌遗传转化的一个的新的技术,具有转化效率离、成本低,操作方便和重复性好多重优点。
我们在构建了二个含有潮霉素B磷酸转移酶基因双元载体的基础上,成功地实现了农杆菌介导的稻瘟病菌转化,转化效率达每106个分生孢子>300个转化子,并得到了4000多个转化子。
通过继代培养和PCR检测,证明插入到稻瘟病菌基因组中的潮霉素抗性基因可稳定遗传。
Southern杂交分析表明,大约有2/3转化子的T-DNA插入是单拷贝的。
用大麦叶片离体接种的方法快速测定部分稻瘟病菌转化子的致病性,发现一个致病缺陷突变体:A1-412。
该突变体不能侵入水稻叶片及擦伤的大麦或水稻叶片,说明A1.412突变体在寄主组织中扩展进程被阻断。
进一步的表型分析,发现A1,412突变体的产孢量显著下降,仅为野生菌株的7%,在疏水表面不能形成附着胞,部分分生孢子萌发的速度也略有延迟。
Southcm杂交显示A1-412基因组中T-DNA插入是单拷贝的。
t-dna插入失活技术原理

t-dna插入失活技术原理t-dna插入失活技术是基因工程技术中的一种重要方法,它是利用外源DNA片段t-DNA 插入到目标基因中导致目标基因的失活从而研究基因功能的技术。
本文将分别从t-DNA插入原理、插入失活原理和t-DNA插入失活技术操作方法等方面进行详细介绍。
一、t-DNA插入原理t-DNA插入原理是指将来自农杆菌的具有植物感染活性的农杆菌(Agrobacterium tumefaciens)或农杆菌样细菌(Agrobacterium rhizogenes)分泌出来的Ti质粒或Ri质粒中的t-DNA随机插入到植物基因中,从而导致基因的失活或突变。
t-DNA(转移DNA)是一种由病原细菌通过T型四边形转移机制将其质粒中的特定DNA片段转移至植物细胞中的DNA。
t-DNA的大小和形状有很大的差异,但一般都包含左右边界(LB和RB)和一定的非编码序列。
通过LB和RB,t-DNA片段可以在植物基因组内随机插入到目标基因的某个位点中,从而会导致基因的失活或突变。
二、插入失活原理通常来说,t-DNA插入至基因区之下或之上才可能导致基因的失活或突变。
一旦t-DNA 片段成功插入至植物基因组的不适当位置上,将会导致基因的失活或突变。
在植物细胞中,t-DNA的LB和RB片段会被发现和被绑定到某些植物基因组中的端粒Repeat(telomeric repeat)上。
然后,t-DNA将和相邻的组织特异启动子、转录子或其他基因序列合并在一起,形成插入位点。
一旦t-DNA插入到目标基因中,其对目标基因的影响主要包括以下两种方式:第一是t-DNA插入至基因区之下,从而导致基因沉默或失活。
第二种是t-DNA插入至基因区之上,从而产生空穴(null alleles)或欠表达的突变形式。
三、t-DNA插入失活技术操作方法t-DNA插入失活技术是一种通过肆意插入t-DNA片段的方式探究植物基因功能的方法。
t-DNA的插入通常是随机发生的,因此需要通过重复筛选最终获得所需的突变体。
水稻突变体介绍及鉴定(很详细)

RMD水稻突变体信息及基因型鉴定1.背景介绍:突变体对于遗传学研究有着重要作用,随着拟南芥和水稻等物种全基因组测序的开展,人类积累了前所未有的基因序列信息,为了弄清这些基因序列的生物学信息,寻找该基因区段序列发生变异的突变体是阐释基因功能最直接最有效的方法。
植物在自然的环境条件下也会产生突变性状,早期普通正向遗传学研究往往通过寻找与某种生物学特性相关的突变体来发掘或定位某个特定基因。
为配合植物功能基因组研究高通量的策略,构建水稻等物种的大型突变体库已成为必然,借助水稻全基因组测序信息、通过反向遗传学的手段大规模地筛选突变体库,理论上可以获得基因组中任一基因的突变体,最终实现阐释基因功能的目的。
2.原理:2.1农杆菌介导的T-DNA 插入农杆菌是寄主范围非常广泛的土壤杆菌,它能通过伤口侵染植物导致冠瘿瘤和毛状根的发生。
1974从根癌农杆菌中分离出一种与肿瘤诱导相关的质粒,称为致瘤质粒(Tumor-inducing plasmid),简称Ti 质粒。
Ti 质粒上存在一段DNA,能够转移并整合到植物基因组中,称为Transferred DNA,简称T-DNA。
研究发现,T-DNA 两端存在非常保守的同向重复的25bp 序列,分别称为左边界(LB)和右边界(RB)。
T-DNA 的转移只与边界序列相关,尤其是RB,而与T-DNA区段的其它基因或序列无关。
我们将T-DNA 区段上的致瘤基因和其它无关序列去掉,利用其转移的特性,实现农杆菌介导的T-DNA 转入水稻愈伤,从而构建水稻突变体库。
大量研究表明,农杆菌T-DNA 整合到植物基因组中的位置是随机的,并且整合到植物基因组中的T-DNA 能稳定遗传。
由于插入到植物基因组中的T-DNA 区段序列已知,这样随机插入到植物基因组中的T-DNA类似于给植物基因“贴”了一个序列标签。
我们利用这个标签,通过各类PCR技术最终可以获取其插入的位点。
2.2 水稻Tos17 反转录转座子创造水稻突变体的另一种方法是利用植物的反转录转座子,它们是以DNA→RNA→DNA 的方式进行转座,在水稻上已发现大约40 种长未端重复的反转录转座子,它们是Tos1-Tos32,RIRE1-RIRE8,其中5 类被证明是有转座活性的,分别是Tos10、Tos17、Tos19、Tos25 和Tos27。
T-DNA插入鉴定实验报告

T-DNA插入突变体的鉴定时明辉同组者:薛敏学号:201000220069摘要 Ti质粒是上有一段特殊的DNA区段,当农杆菌侵染植物细胞时,该DNA区段能自发转移进植物细胞,并插入植物染色体DNA中。
所以Ti质粒上的这一段能转移的DNA被叫做T-DNA。
将感兴趣的基因改造插入到T-DNA区段中,通过农杆菌侵染植物细胞,实现外源基因对植物的遗传转化,得到含有突变的植株。
通过本实验,我们将学习如何用PCR的方法检测所得植株是否为T-DNA的插入突变体。
1.引言T-DNA作为一种实验常用的遗传转化方法,在插入突变过程中,插入到植物染色体上的位置是随机的。
如果T-DNA插入进某个功能基因的内部,特别是插入到外显子区,将造成基因功能的丧失。
所以利用农杆菌Ti质粒转化植物细胞,是获得植物突变体的一种重要方法。
农杆菌Ti质粒转化植物细胞后,在获得的后代分离群体中,有T-DNA插入的纯合突变体,杂合突变体,和野生型。
在突变体研究中,需要的材料是纯合突变体,所以必须从分离群体中将纯合突变体鉴定出来。
本次实验中,采用液CTAB(或者TSP法)提取拟南芥植株的DNA,然后PCR将所获DNA 扩增,在之后采用琼脂糖凝胶电泳技术,分离处长度不一的DNA带,以确定样品是否为T-DNA 插入突变纯和体。
PCR(Polymerase Chain Reaction),即聚合酶链式反应是体外核算扩增技术,具有特异、敏感、产率高、快速、简便、重复性好、易自动化等突出优点;能在一个试管内将所要研究的目的基因或某一DNA片段于数小时内扩增至十万乃至百万倍,使肉眼能直接观察和判断。
(PCR基本原理如右图)3-基团,在pH8.0 BufferDNA含有PO4(本实验中为TAE)中带负电, 在电场中向正极移动。
自由电泳时,由于不同大小的DNA片段的电荷密度大致相同,各核酸分子难以分开;选用适当浓度的琼脂糖凝胶作为支持物,使之具备一定的孔径,即可发挥分子筛效应,使大小不同的核酸片段迁移率出现较大差异,达到分离的目的;同样条件对Marker电泳;起到鉴定的作用。
稻瘟菌T-DNA插入所致Pi9致病突变体的获得

l rr fMa a oteg e edsri J 5 5 B b ce nn h ievreyw t e i a c e eP9 h o iv ln h i ayo gnp r nf l tan F9 0 4 ysre igteep st eco ei te b h i c h t i n
M a n po t rs a T- g a rhe g ie DNA n e to a i s ri n lmut nt r l n o Pi a s viu e tt 9
L N n,P h — i I Ho g y ,C I Ya AN C u y ,L n - u HEN L - u i a,L o d n W ANG Z n - u h U Gu — o g, o gh a
维普资讯
福建农林大学学报 ( 自然科 学版) Ju a o ul g c tm adF r t n e i N t a Si c dtn o r l f j nA f u u n oe r U i r t a rl c neE io ) n F a il s y v sy( u e i
第3 5卷 第 3期
20 0 6年 5月
稻 瘟菌 T D A插人所致 P9致病突变体 的获 得 -N i
林 艳, 潘初沂 , 李宏宇 , 陈丽华 , 国东 , 鲁 王宗华
( 福建农林大学教育部生物农 药与化学生物 学重点实验 室, 福建 福州 3 00 ) 502
摘要: 利用 自 建稻 瘟菌菌株 F904 .N J55B TD A插 入突变体库, 选获得 2个对 水稻 品种 P 筛 i 9致病的 突变 体 P - 8 i2 9—
由稻瘟菌 Manpt re H r r a ( g ao e ia( e e )Br 无性态为 P ru r re ac ) h gs bt r y cl i giaSc. 侵染水稻引起 的稻瘟病 i aa s 是水稻最重要的病害之一. 该病害的经济重要性及其遗传可操作性 , 使之成为研究植物病原真菌与寄主互
稻瘟菌生长缓慢突变体的筛选及遗传分析

1 . 1 . 1 菌 株 野 生 菌 株 S 1 5 2 8 、 野生菌株 P 1 3 1 、 P 1 3 1的 A T M T ( a g r o b a e t e r i u m—t u me f a c i e n s m e d i a t e d t r a n s f o r m a t i o n ) 转
的A T MT转化子 的菌落 大小 , 将在 菌落生 长速度上 与野生 型 菌株 P I 3 1 有 明显差异的转化子挑出 , 进行复筛 。 复筛 : 用5 a r i n打孔器分别打取 上述 培养 的 P 1 3 1和菌 落
直径小 的 A T MT转化子边缘 菌落 , 接种到西 红柿燕麦 培养基 平板上 , 每个培养皿接种 1 个 菌株 , 2 5℃培 养 7 d , 并 在培养 3 、 5 、 7 d时测量其菌落直径 。
获得致病性变异突变体 型与插入标记 共分离 的插入 突变体 , 为 以后的基因克隆和功能分析及 阐述其致病的分子机理提供了
试验材料。 1 材 料与方法
1 . 1 材料
接到另一西 红柿 燕麦 培养基平 板 中央 , 每皿 1 块, 并标 记 编 1 . 2 . 2 . 3 生长缓慢型突变体 的筛选 初筛 : 观察培养 7 d后
1 0 mg / m L链霉素 4 0 0 w L 。链霉素使用 时待培养基温度 降至
断深入研究稻瘟 病菌生物学特性与其致病性之间的关系以及
侵染寄主的过程等 , 对于开发新型杀菌剂具有重要意义 。 目前已从 稻瘟 病 菌 中鉴 定 出包括 A V R—C O 3 9 、 A V R~ Y A MO 、 A V R一 1 S u V等在 内的 2 0多个 无毒 基 因。病 原菌 突
稻瘟病菌MoSCJ1基因的生物学功能分析

第50卷第3期2021年5月福建农林大学学报(自然科学版)Journal of Fujian Agriculture and Forestry University ( Natural Science Edition )稻瘟病菌MoSCJ l 基因的生物学功能分析陈浩,王敏,杨子峰,廖煌,吴欢,凌菡,汤蔚(福建农林大学闽台作物有害生物生态防控国家重点实验室,福建福州350002)摘要:以稻瘟病菌野生型菌株Guyll 为材料,利用基因敲除和回补得到稻瘟病菌的突变体\Moscj 1和互补菌株A Moscjl / MoSCJ l,并以野生型为对照,分析了 MoSCJ l 的生物学功能.结果发现:突变体b Moscj l 在基本培养基上的生长速率显著下 降,同时,MoSCJ l 基因的缺失导致突变体对细胞壁胁迫剂更加敏感;但MoSCJ l 基因的缺失并不影响稻瘟病菌的无性繁殖、 附着胞发育及致病性.这说明MoSCJ l 参与调控稻瘟病菌的营养生长和对细胞壁胁迫剂的应答过程,不参与调控该菌的无性繁殖及致病过程.关键词:稻瘟病菌;MoSCJ l ;营养生长;细胞壁完整性中图分类号:S432.44 文献标识码:A文章编号:l67l-5470(202l)03-0309-08DOI :l0.l3323/ki.j.fafu(nat.sci.) .202l.03.003开放科学(资源服务)标识码(OS1D )Biological function of MoSCJl in Magnaporthe oryzaeCHEN Hao, WANG Min, YANG Zifeng, L1AO Huang, WU Huan, L1NG Han, TANG Wei (State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops , Fujian Agriculture and Forestry University ,Fuzhou, Fujian 350002, China)Abstract : We obtained the MoSCJ l gene deletion mutant from the wild type Guyll of Magnaporthe oryzae and the complemented strain via gene knock-out and complementation technologies , and then analyzed the biological functions of MoSCJ l in rice blast fun gus. We found that the deletion of MoSCJ l significantly impaired the vegetative growth of M. oryzae on basic medium and led to high er sensitivity to cell wall stressors. However , the loss of MoSCJ l had no effects on the sporulation , appressorium development and pathogenicity. 1n conclusion , MoSCJ l is likely to be involved in the regulation of vegetative growth and cell wall integrity of M. oryzae , instead of the regulation of asexual propagation and pathogenicity.Key words : Magnaporthe oryzae ; MoSCJ l ; vegetative growth ; cell wall integrity稻瘟病是全球水稻生产上的重要病害之一,对水稻的产量和品质具有巨大的威胁,发生严重时能造成 整片稻田绝收.稻瘟病菌(Magnaporthe oryzae )通过分生抱子传播,分生抱子在水稻叶片表面萌发产生芽管 并形成附着胞,后产生侵染栓穿透叶片的角质层和表皮细胞壁,在寄主细胞内分化产生次生菌丝并侵染邻 近细胞和组织,继而发病.内质网是由囊状、管状和泡状结构组成的一个连续的网膜系统.真核细胞中,在折叠酶、伴侣结合蛋白 等协助下,内质网对分泌性蛋白质合成、转运和加工并运输到相关的细胞部位发挥功能.为了保证蛋白质 能正确折叠和修饰,以达到天然构象从而转运到目标细胞器或分泌到胞外,细胞内部有一套严格的蛋白质 质量控制系统,一旦蛋白质由于转录翻译失败或不同条件压力等原因发生错误折叠并聚集在内质网中,内 质网就会产生压力,影响细胞的正常生理功能['-2].真核细胞存在两条途径调控内质网压力:非折叠蛋白响应(unfolded protein response , UPR)和内质网 相关蛋白降解(endoplasmic reticulum-associated protein degradation , ERAD)⑶.其中,ERAD 主要通过泛素 化降解内质网中错误折叠或未折叠的蛋白[4],其包括底物识别、定位、泛素化、逆向运输和蛋白酶体降解 等步骤.底物识别水平决定底物的降解程度.蛋白质错误折叠是由于其本应在蛋白质内部的疏水区暴露在 外从而引起蛋白质之间的聚集而发生,在底物识别的过程中,热休克蛋白Hsp70家族成员(Bip/Kar2)会收稿日期:2020-07-28 修回日期:2020-l0-22基金项目:福建农林大学科技创新专项基金项目(CXZX20l9007G 、CXZX20200l4A ).作者简介:陈浩(l998-),男.研究方向:植物保护.Email : chenho998@ .通信作者汤蔚( l988-),男,讲师,博士.研究方向:真菌功能基 因 组 学 .Email : tangw@ .-310-福建农林大学学报(自然科学版)第50卷与疏水区结合促进多肽链折叠,从而避免蛋白质分子的聚集[5].与Hsp70长时间作用后的底物会被E3泛素连接酶Hrdl复合物识别⑷.Hsp40/DnaJ是一个可以调节Hsp70分子伴侣活性的蛋白质家族,Hsp40s刺激Hsp70蛋白固有的弱腺苷三磷酸(adenosine triphosphate,ATP)酶活性,使其由ATP状态转变为腺苷二磷酸(adenosine diphosphate,ADP)状态,从而对底物有强亲和性,促进Hsp70与多肽底物的相互作用. Hsp70家族成员通常有多个Hsp40伴侣,其特定的配对控制Hsp70伴侣参与调控特定过程.所有Hsp40均含有高度保守的75个氨基酸J结构域,该结构域与Hsp70的ATP酶结构域相互作用,促进ATP水解[7].研究表 明,酿酒酵母(Saccharomyces cerevisiae)基因组的Hsp40/DnaJ家族中有22种蛋白质,其中包括Scjl蛋白囚.Scjl作为一种蛋白分子伴侣参与蛋白的折叠过程.已有研究发现,缺失Scjl会影响Hsp70的活性,进而影响ERAD途径中底物的识别及降解过程,最终启动细胞的凋亡程序冈.目前,有关Scjl在丝状真菌中的研究较少.本试验利用基因敲除技术获得MoSCJ l基因敲除突变体(^Moscj l),研究其营养生长、无性繁殖、致病力及对细胞壁胁迫剂的应答等情况,以探讨MoSCJ l基因在稻瘟病菌中的生物学功能,为其他病原真菌中SCJl基因的研究提供一定的参考.1材料与方法1.1供试菌株及其他材料本研究中所用稻瘟病菌野生型菌株Guyll由福建农林大学闽台作物有害生物生态防控国家重点实验室保存,△M oscj l突变体和互补菌株A Moscj l/MoSCJ l均由本试验获得.试验所用的水稻品种CO39、大麦种子均保存于福建农林大学闽台作物有害生物生态防控国家重点实验室的4冷库中.完全培养基(complete medium,CM):20x硝酸盐50mL•L_l,l000x微量兀素l mL•L_l,l000x维生素溶液l mL-LT,葡萄糖l0g-LT,蛋白胨2g-LT,水解酪蛋白l g-LT,酵母提取物l g-LT,琼脂粉l5g•L_l.基本培养基(minimal medium,MM):NaNO36g,KCl0.52g,MgSO4•7H2O0.l52g,KH2PO4 l.52g,维生素Bl0.0l g,微量元素溶液l mL,葡萄糖l0g,琼脂粉l5g.稻秆培养基(straw decoction and corn,SDC):稻秆l00g-L-l(煮沸20min,两层纱布过滤),玉米粉40g-,琼脂粉l5g-⑼.TB3培养基:水解酪蛋白3g-LT,酵母提取物3g-LT,蔗糖20%,琼脂粉l2g-L»色氨酸缺陷型合成葡萄糖基础培养基(synthetic dextrose minimal medium without tryptophan,SD-Trp):无氨基酵母氮源6.7g,D-山梨糖醇l82.2g,缺色氨酸氨基酸粉末0.74g,葡萄糖20g,琼脂粉l5g,pH5.8[l0].LB培养基:胰蛋白胨l0g-L'1,酵母提取物5g•L_1,NaCl10g-L_1,琼脂15g-L_1;酵母膏胨葡萄糖琼脂培养基(yeast extract peptone dextrose medium,YPD):葡萄糖20g-L'1,蛋白胨20g-L'1,酵母提取物10g-L'1,琼脂粉15g•L'1.试剂与仪器:潮霉素B,北京索莱宝科技有限公司;博来霉素,美国Innivogen公司;Southern杂交试剂盒,美国罗氏公司;rTaq酶、RNase,宝日医生物技术(北京)有限公司;刚果红(congo red,CR),西格玛奥德里奇(上海)贸易有限公司;microcloth,EMD Millipore公司;T100PCR仪,美国Bio-Rad公司;智能培养箱,宁波江南仪器厂;Nikon E200生物显微镜、激光扫描共聚焦显微镜,日本尼康公司;血球计数板,北京索莱宝科技有限公司;Centrifuge5420离心机,德国艾本德股份公司;PowerShot G7X Mark H相机,佳能(中国)有限公司.1.2MoScjl的系统发育分析在酵母基因组数据库中得到酿酒酵母Scjl的蛋白质序列,根据其与本试验稻瘟病菌Guyll的序列同源性比对结果,得到的同源蛋白为MoScjl.在NCBI中利用Blast比对得到一组MoScjl的同源蛋白序列,选取8个不同物种的序列在MEGA5.0软件中采用邻接法(neighbor-joining,NJ)建立系统发育树[11].1.3MoSCJ l突变体的获得1.3.1MoSCJ l基因的敲除以稻瘟病菌野生型菌株Guyll基因组为模板,以MoSCJ1-F1/MoSCJ1-F2和MoSCJ1-F3/MoSCJ1-F4(表1)两对引物对MoSCJ l基因上游和下游各1000bp左右的DNA片段进行PCR 大量扩增.与此同时,将pCX62质粒作为模板,以引物HYH-F/HYH-R(表1)对潮霉素磷酸转移酶基因全长进行PCR扩增,并对扩增结果进行纯化回收.将扩增好的MoSCJ l基因上游和下游各1000bp左右的第3期陈浩等:稻瘟病菌MoSCJ l基因的生物学功能分析-311-DNA片段及潮霉素磷酸转移酶基因全长中的目的片段进行融合PCR,获得长度约为3400bp的重组片段.以引物MoSCJ1-F1/MoSCJ1-F4(表1)对重组片段进行大量扩增,纯化回收所得片段,将回收产物用于原生质体的转化[12].用潮霉素(600愿-mf)筛选转化子.表1试验所用引物Table1Primers used in this study序列(5'f3‘)引物MoSCJl-Fl MoSCJ1-F2 MoSCJ1-F3 MoSCJ1-F4 MOSCJ1-BY-F ACTATAGGGCGAATTGGGTACCGCATCGTCTCCATGGCTGA GCGCCGAATTCGATATCAAGCTTGGCACCTAAGGCTCACTGC TGCCGACCGGGCCGGCCGGATCCGCGAAGGTGCGGTTAGAC GGTGGCGGCCGCTCTAGACGATGTTGTCAACCTCGATG CCTCAGTGAGGTAGCAAGHY-R GTATTGACCGATTCCTTGCGGTCCGAAMoSCJ1-KO-F MoSCJl-KO-R HYH-F CTGAGTTCCAGTGGGAGAGG CCTCCTTCCAGAACAGATCG TGGAGCTAGTGGAGGTCAACAATGAATGHYH-R CGGTCGGCATCTACTCTATTCCTTTGC GFP-R CGGTGGTGCAGATGAACTTCNP-SCJ1-F NP-SCJ1-R ACTCACTATAGGGCGAATTGGGTACTCAAATTGGTTGAACTTCTCCTGTCCGTACT CACCACCCCGGTGAACAGCTCCTCGCCCTTGCTCACCAACTCATCATGACCGGAGG1.3.2MoSCJ1突变体的鉴定用灭菌的牙签挑选转化子至CM培养基上,培养4d后,用CTAB法粗提转化子DNA[I3然后以粗提的转化子DNA为模板,用引物MoSCJ1-KO-F/MoSCJ1-KO-R(表1)进行PCR验证,阳性对照以野生型菌株Guy11基因组为模板,阴性对照以无菌水为模板,在阳性对照有条带、阴性对照无条带的前提下,对无条带出现的转化子进行下一步验证;以无条带的粗提转化子DNA为模板,用引物MoSCJ1-BY-F/HY-R(表1)再次进行PCR验证,阳性对照和阴性对照同上,在阳性对照、阴性对照均无条带的前提下,有条带的转化子即为突变体.1.4回补菌株的获得1.4.1回补载体的构建以野生型菌株Guy11基因组为模板,以引物NP-SCJ1-F/NP-SCJ1-R(表1)进行PCR扩增,得到长度为4247bp的片段.将其与pYF11骨架片段共同转入XK-125酵母感受态细胞中,并于SD-Trp培养基上培养24h.以酵母转化子为模板,用引物MoSCJ1-KO-F/GFP-R(表1)进行PCR验证,取有条带的酵母转化子在YPD培养基中摇培18h,提取质粒.将酵母质粒转入大肠杆菌中,再进行PCR验证,并将结果测序,获得回补载体.1.4.2回补菌株的鉴定将回补质粒转入突变体的原生质体中,培养4d后挑出转化子,在激光扫描共聚焦显微镜下用博来霉素(200jig-mL_1)进行筛选.1.5MoSCJl突变体的生物学表型分析1.5.1突变体在不同培养基上的营养生长从保菌袋中取出野生型菌株Guy11、突变体和回补菌株的存菌滤纸片分别接种于CM培养基上,于28T黑暗培养箱中培养5d.然后在各菌落边缘切取3mmX3mm 的菌块并接种于CM、MM、SDC培养基上,每个菌株设3个重复.在28弋黑暗培养箱中培养7d后,测量、记录其直径并拍照.1.5.2突变体对细胞壁胁迫剂的响应从野生型菌株Guy11、突变体和回补菌株的菌落边缘切取3mmX 3mm的菌块,分别接种于含有胁迫剂CR(400ig•mL"1)的CM培养基上,每个菌株设3个重复.在28°C 黑暗培养箱中培养7d后,测量、记录其直径并拍照,同时计算抑制率,抑制率=(空白组菌株直径-处理组菌株直径)/空白组菌株直径X100%.1.6MoSCJ l突变体的产抱量测定及分生抱子梗观察1.6.1产孢量的测定从野生型菌株Guy11、突变体和回补菌株的菌落边缘切取3mmX3mm的菌块,分别接种于SDC培养基上,每个菌株设3个重复,在28C黑暗培养箱中培养5d.刮去表面气生菌丝,置于黑光灯下培养5d后,用无菌水冲洗培养基,2层microcloth过滤于10mL离心管中得到抱子液,并最终定容・3l2・福建农林大学学报(自然科学版)第50卷至2mL.在血球计数板上统计抱子数目,并记录数据.l.6.2分生孢子梗的观察菌株培养方法同l.6.l.切2cmx2cm的菌块于载玻片上,菌丝面贴载玻片,黑光灯下培养2d后,在Nikon E200生物显微镜下观察分生抱子梗产抱情况并拍照.试验重复3次.1.7MoSCJ l突变体的附着胞萌发试验抱子液的制备方法同l.6.l,调整抱子液浓度为8xl05个・mL-l・将疏水性盖玻片置于载玻片上,向盖玻片上滴40|xL抱子液后,将载玻片放入保湿盒中,于28°C黑暗培养箱中培养,分别于4、8、l2、24h在Nikon E200生物显微镜下观察分生抱子的萌发情况并拍照,每个菌株统计l00个抱子,计算抱子萌发率和附着胞形成率.试验重复3次.1.8MoSCJ l突变体的侵染试验l.8.l水稻侵染种植感病水稻品种CO39,于26C温室中培养至三叶一心期.抱子液的制备方法同l.6.l,用血球计数板调整抱子液浓度为8xl05个・mL-1,向抱子液中加入0.2%(体积分数)明胶,混匀,用喷壶均匀地喷洒在水稻叶片上.然后将水稻置于28C黑暗培养箱中培养24h,再放在接种室培养6d,观察水稻的感染情况,统计并记录病斑等级[l4],对感病水稻叶片进行拍照.试验重复3次.l.8.2离体大麦侵染种植大麦,于温室中培养9d.抱子液的制备方法同l.6.l,用血球计数板调整抱子液浓度为8xl05个・mL-1,向抱子液中加入0.2%(体积分数)明胶.剪取长度一致的大麦叶片,滴20応抱子液于大麦叶片背部,将大麦叶片置于28C黑暗培养箱中保湿培养24h.撕取大麦背部表皮,在Nikon E200生物显微镜下观察菌丝侵染情况并拍照.试验重复3次.1.9数据处理用Excel20l0软件对试验数据进行分析和处理,用DPS数据处理系统进行单因素统计分析.2结果与分析2.1MoScjl系统发育分析系统发育树分为两大分支,MoScjl与酿酒酵母S288C具有高度同源性,且从整体来看与其他7个物种裂殖酵母(Schizosaccharomyces pombe)、拟南芥(Arabidopsis thaliana)、粘菌(Dictyostelium discoideum AX4)、果蝇(Drosophila melanogaster)、利什曼原虫(Leishmania donovani)、秀丽线虫(Caenorhabditis ele-gans)、人类(Homo sapiens)的同源蛋白序列均有相似性.由此推测这些物种的Scjl同源蛋白在功能上可能具有保守性.85---------------------------------------------------酿酒酵母Saccharomyces cerevisiae S288C98--------------------------------------------Magnaporthe oryzae---------------------------------裂殖酵母Schizosaccharomyces pombe-------------------------------拟南芥Arabidopsis thaliana99-------------------------------------------------ttW Dictyostelium discoideum AX4---------------------------------------------果蝇Drosophila melanogaster59------------------------------------------------禾Leishmania donovani88-----------------------------------------Caenorhabditis elegans91-----------------------------人类Homo sapiens图l稻瘟病菌Scjl与其他8个物种同源蛋白序列的系统发育树Fig.l Phylogenetic tree of the homologous amino acid sequences of Scjl in M.oryzae and other8species2.2MoSCJ l突变体的鉴定利用同源重组原理构建了基因敲除载体(图2A),通过原生质体转化的方法,得到具潮霉素抗性的转化子.粗提转化子基因组DNA,利用基因内部引物、臂外引物进行PCR验证,基因内部引物验证无条带、臂第3期陈浩等:稻瘟病菌MoSCJl 基因的生物学功能分析・313・外引物验证有条带的转化子即为突变体.结果显示,转化子'Mosc/1-4、'Mosc/1-22、'Mosc/1-23和'Mosc/1 - 37为符合条件的MoSCJ l 基因敲除突变体(图2B ).随机选取'Moscj l -22突变体用于后续试验.2.3 MoSCJ 1突变体的营养生长图3显示,突变体菌株在CM 和SDC 培养基上生长的直径与野生型菌株和回补菌株相比无明显差异, 而在MM 培养基上生长的直径相比于野生型菌株和回补菌株极显著缩小(P <0.01).可见,MoSCJ l 参与调 控稻瘟病菌在基本培养基上的营养生长过程.A 1 kbB MoSCJl -Guyll -----------------------——---------------------- 寻 gdDsowd 0 旨owd 寸\ hph I\Moscj\2F 2RA.MoSCJl 基因的敲除策略;B.PCR 验证结果(Guyll 为以野生型菌株为模板的阳性对照,Mock 为以无菌水为模板的阴性对照).图2 MoSCJ1基因的靶向敲除Fig.2 The targeted deletion of MoSCJl geneB ■Guyll □ ^Moscjl**表示同一培养基下不同菌株之间的差异极显著(P <0.01).图3 野生型菌株Guy11、突变体NMoscj1、回补菌株^Moscj1/MoSCJ1在CM 、MM 、SDC 培养基上的营养生长情况(A )和菌落直径(B )Fig.3 Vegetative growth (A) and colony diameters ( B) of Guyl 1, 'Moscjl and 'Moscjl /MoSCJl on CM , MM, SDC plates2.4 MoSCJ 1突变体对细胞壁胁迫剂的响应相比于野生型菌株和回补菌株,突变体对CR 更加敏感(图4),抑制率呈现极显著差异(P <0. 01).这 说明MoSCJ l 参与调控稻瘟病菌对细胞壁胁迫剂的应答过程.CK 为不加入胁迫剂的对照组;CR 为加入刚果红的处理组.**表示差异极显著(P <0.01).图4 野生型菌株Guy11、突变体\Moscj1、回补菌株^Moscj1/MoSCJ1在含有CR 的CM 培养基上的生长情况(A )和抑制率(B )Fig.4 The growth (A ) and inhibition rate of Guyll , 'Moscjl and 'Mosc/l/MoSCJl on CM plates containingCR・3l4・福建农林大学学报(自然科学版)第 50 卷2.5 MoSCJ l 突变体的无性繁殖结果显示,野生型菌株、突变体和回补菌株均能产生大量抱子(图5A ),且三者之间的产抱量无显著 差异(图5B ).由此推测MoSCJ l 不参与调控稻瘟病菌的无性繁殖过程.AB 二 A n u 204 O 1612Guyl 1 bMoscj\ NMoscj'l MoSCJX 菌株类型旨owaUUSOW 己。
PCR法鉴定T-DNA插入突变体1概论

• T-DNA大多为单拷贝插入,使其利于进行遗传分析。
鉴定原理
BP: T-DNA载体上的插 入片段的左边界序列 >>>通用引物(LB)
LP、RP: 用已知的被插入 的基因的侧翼序列设计的 特异性引物
鉴定原理
Mix 17 ul
16℃ ∞
32 cycles
• 电泳检测
Mix 1: LP+RP Mix2: LB+RP
PCR法鉴定T-DNA插入突变体
T-DNA插入
• T-DNA,转移DNA(transferred DNA)是根瘤农 杆菌Ti质粒中的一段DNA序列,可以从农杆菌中转移 并稳定整合到植物基因组
• 将目的基因转入到经过改造的T-DNA区,借农杆菌 的感染实现外源基因向植物细胞的转移与整合,获得 转基因植株
WT 没有插入
HM 纯合体
HZ 杂合体
鉴定原理
本次实验材料:
插入位点
Exon Intron UTR promoter
1100 bp 500 bp
实验安排
提取植物基因组
PCR反应鉴定
实验安排
提取植物基因组
• 每2个人一个小组,每小组提 1 个样品 • 每行一个大组,另提1个对照WT • 注意写好样品编号源自实验安排提取植物基因组
(1)剪取约0.5-1 cm2大小的幼嫩植物叶片,放入离心管中; (2)在液氮中,用研磨棒将叶片磨碎; (3)研磨结束后,向管中加入400 uL的提取缓冲液,混匀; (4)室温,12000 rpm离心3 min; (5)吸取300 uL上清液至一新的离心管中,加入300 uL异丙醇,混匀; (6)室温,12000 rpm离心5 min; (7)倒掉上清,加入300 uL 70%乙醇; (8)室温,12000 rpm离心3 min; (9)倒掉上清,待管中液体晾干后,加入50 uL ddH2O溶解。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
杆菌的抗生素头孢噻肟钠和羧苄青霉素的配比, 不同转化阶段培养基的选择等。转化 ! G !%4 个孢子平均可获得约 5%% 个左右的转化子, HIJ 和 2EKL0HIJ 检测表明约 15M 转化子中含 20D,E 插入。对 !5$% 个突变体进行形态变异 观察, 发现菌落颜色突变的有 !5 个; 随机取 51 个突变体进行比较, 发现产孢量减少的 # 个, 孢子萌发率降低的 1 个, 附着胞形成率降低的 " 个; 还获得对水稻品种 I!%!LEI ( H90!) 和 ’50!0!$’ ( H90") 致病的突变体, 为进一步克隆相 应的无毒基因奠定了基础。 关键词 稻瘟病菌,20D,E,插入突变,功能基因组学 N’5! 文献标识码 E 文章编号 !%%%0&%4! ($%%&) %#0%#!"0%5
[’] ( .+7 O! Z 9#$ ;") 的 XFOR! , 由美国宾夕法尼 >X>
年新发展的农杆菌介导转化 ( 51*(6+0/%*#)7 /)7%-+8 的 20D,E 插入方 0#%$&073;98R3; RQ8<TB)Q78R9)<, E2U2) 一是转化 法解决了这一问题。 E2U2 方法的优点: 材料易得, 可以用原生质体、 菌丝、 孢子和蘑菇的菌 二是得到 素等, 而 J6UK 只能用原生质体进行转化;
[4] 中 , 这给进一步的突变体分析带来很多麻烦。近 [5]
&
&’&
材料与方法
供试菌株 稻瘟病菌菌株 4VF!5、 "5%5#F、 1!$’# 和测 14%54
均为本实验室保存的从福建田间分离纯化的单孢菌 系。 农杆菌菌株为 EWL0!。 20D,E 插入所用质粒为 XIEUFKE!&%% 中的 XI8U(&5Y0>X> 被 置 换 为 X2QXI0
[/@] [-]
[-]
将 /13@ 个突变菌株每 3@ 个一组混合接种于含 单个抗瘟基因的水稻品种 ./@/5J.( R+M/) 、 ./@/J1/ (R+M3) 和 -1M/M/3( R+M’) 及感病对照 .W8’ 上, 31D 保 湿 3A>, 然后置于控温玻璃温室中让其发病, C Z /@E
[/8] 后调查病情, 参照 9",4%, 等 @ Z 1 级的标准记载 病斑反
将稻瘟病菌 A 个田间 -"!"! 抗生素的选用和优化: 菌株 B[9/1、 ’1@1A9、 C/3-A 和测 CB@1B 的分生孢子置 于不同浓度的潮霉素 9 水溶液中, 在 3CD 下培养, 并分别在 C、 AC、 ’B> 观察其萌发情况。结果对照 C> 内, 分生孢子萌发率就已达 ’@2 ; 但在 3@@ :645 潮 ! 霉素 9 水 溶 液 中, 只 有 /2 萌 发; 而 在 A@@、 B@@ 和 孢子都没有萌发。对潮霉素 9 水溶液 C@@ :645 中, ! 进一步观察的结 中的分生孢子培养 AC> 和 ’B> 后, 果表明经 3@@ :645 处理的分生孢子萌发率略有增 ! 加, 分别是 32 和 12 ,A@@、 B@@、 C@@ :645 中仍然未 ! 见孢子萌发, 菌株间没有差异。可见 3@@ :645 潮霉 ! 素 9 水溶液对稻瘟病菌分生孢子萌发有较好的抑 制效果。 而将上述 A 个菌株的分生孢子均匀地铺于含不 同浓度潮霉素 9 的培养基平板上, 结果是 3CD 培养 其余则未见; 3E 后在对照中就可以观察到菌落, 8E 后对 照 菌 落 已 基 本 长 满 平 板, 所有菌株均可在 而且 3@@ :645 潮霉素 9 平板上见到少量菌落生长, ! 菌株 ’1@1A9 在 A@@ 潮霉素 平板上仍可见少 :645 9 ! 量菌落生长; AE 后所有菌株在 A@@ :645 潮霉素 9 ! 平板上均有菌落生长, 菌株 ’1@1A9 在 B@@ :645 潮 ! 霉素 9 平板上也可见少量菌落生长。结果表明, 在 平板培养基上, 需要较高浓度的潮霉素 9 才能达到 较好的抑菌效果, 而且不同稻瘟病菌菌株间所需的 抑菌浓度也有差异。因此, 在进行转化选择时必须 而且要尽早将 根据所用菌株确定潮霉素 9 的用量, 转化子转移到新的培养基上, 以免出现太高比例的 假转化子。 稻瘟病菌与农杆菌共培养后, 选用合适的抗生 素抑制农杆菌的生长极为重要。结果表明, 在稻瘟 病菌 与 农 杆 菌 共 培 养 后, 从只用头孢噻肟钠 ( 3@@
基金项目: 国家自然科学基金 ( ,)+ &%%%%!!!) , 福建省 “百千万” 人才工程资助。 23*: 1405"!0&’1"#’1; 60789*: :.);)<:*.= /8>))+ ?)7。 23*: 1405"!0&’’"5&4; 60789*: @8<:A>= BC8. + 3;. + ?< " 通讯作者:
!" 卷 # 期 $%%& 年 ’ 月
生 物 工 程 学 报 !"#$%&% ’()*$+, (- .#(/%0"$(,(12
()*+ !" ,)+ # -.*/ $%%&
! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! !
稻瘟病菌 !"#$% 插入方法优化及其突变体分析
$, & $" 李宏宇!, 潘初沂$ 陈 涵! 赵长江$ 鲁国东$" 王宗华!,
$ (! 福建农林大学生命科学学院; 福建农林大学植物保护学院, 福州 &5%%%$) & (中国人民解放军军需大学, 长春 !&%%4$)
摘
要
优化了农杆菌介导转化稻瘟病菌获得 20D,E 插入突变的条件, 包括选择转化子的潮霉素 F 用量, 抑制农
收稿日期: 修回日期: $%%&0%!0$1, $%%&0%#0$1。
亚州立大学的 Y3):?>8< [8<: 博士赠送。 &’( 培养基 酵母粉 $:, 琼脂 !%: 加水 酵母培养基: 淀粉 !%:, 至 !%%%7L。 琼脂 !1 :, 加水至 米糠产孢培养基: 米糠 $% :, !%%% 7L, XO4\5。
[’, 1] 。 E2U2 法 构 建 真 菌 突 变 体 库 已 用 于 多 种 真 菌
并获得一批稻 为此, 我们优化了 E2U2 方法的条件, 包括形态发育和致病性 瘟病菌 20D,E 插入突变体, 突变体。
为进一步从基因组水平认识水稻与稻瘟病菌相互作 用的机制奠定了基础。稻瘟病菌致病性复杂易变, 给水稻抗病育种及抗病品种的利用带来了极大困 难。因此, 利用功能基因组学的方法从全基因组水 平分析该菌的致病性变异机制非常必要。 基因插入失活获得能够覆盖全基因组的突变体 库是功能基因组学研究的基础性工作。在真菌中最 常用的插入失活方法是限制酶诱导整合 ( J3TRQ9?R9)< 但是 J6UK 有许 J6UK) , 3<A/73073;98R3; 9<R3:Q8R9)<, 多缺 点, 最大的缺点是所产生的转化子中约有 高 的 可 达 到 1%M , 其标签没有插入基因组 $%M ,
。
=期
李宏宇等: 稻瘟病菌 45673 插入方法优化及其突变体分析
=(+
的选择培养基上分离纯化的稻瘟病菌株中, !"#$) ! 约有 %&’ 在培养时仍可见农杆菌生长; 而在含头孢 噻肟钠 ((&& 和羧苄青霉素 ((&& 的选择 !"#$) !"#$) ! ! 培养基上分离纯化的稻瘟病菌株中, 只有 %’ 在继 代培养时仍可见农杆菌生长, 但在选择培养基上培 养时间延长, 带农杆菌的比例还会增加。因此, 在选 和 择培养基中, 应同时加入头孢噻肟钠 ((&& !"#$) ! 羧苄青霉素 ((&& 以有效抑制农杆菌的生长。 !"#$) ! !"#"! 培养基的优化: ( 种共培养培养基比较结果 表明, ( 培养基中葡萄糖含量为 , ##-."$) 的共 )*+ 培养效果好, 共培养 /01 在硝酸纤维素膜上可见稻 瘟病菌菌丝生长, 但又不过于茂盛, 结果能得到较多 的转化子; 而在 )*( ( 培养基中葡萄糖含量提高到 , 共培养 /01 看不到稻瘟病菌菌丝, 最后 +& ##-."$) 在选择培养基上也得不到转化子, 这可能是因为葡 萄糖含量高不利于稻瘟病菌生长。 共培养后的稻 ( 种选择培养基比较结果表明, 瘟病菌在 2*+ ( 基本培养基) 、 ( 酵母培养基) 和 2*( (米糠产孢培养基) 上均能良好生长, 说明 34*4 2*/ 方法在选择转化子时, 对培养基要求不严。米糠产 孢培养基不易污染, 而且可以直接产孢, 是比较适宜 的培养基。 !"! 转化子 $%&’( 插入的 )*+ 验证及其插入的 稳定性 随机抽取获得的转化子, 以 45673 左右边界序 列设计的引物进行 8)9,结果从转化子 +5+:5%、 +5+:5 ,、 +58;<5+=、 +58;<5+(、 +58;<5<、 +58;<5%、 +58;<5= 都扩增 到一条约 +>,?@ 大小的片段, 野生型菌株对照和清 水中均未见该条带, 但 8;<5+/ 也没有条带 (图 +) , 可 能没有 45673 插入, 或是 45673 边界序列丢失。多 次转化结果平均每 + A +& 个孢子可以得到约 ,&& 个转化子, 而 8)9 结果则表明其中约有 +,’ 的转化