Section N 真核生物的转录因子
分子生物学

单亚基的因子(35 kD) 能把TFIID与TFIIF/Pol II相连在一起,即是聚合
酶II结合到预起始复合物所必需的 能与一些基因特异转录因子相互作用,促进转录
四、真核生物的基因转录及其调控
4. 真核生物的通用转录因子 (1)II类因子(class II factors) TFIIF的结构及功能
TFIIA
在酵母中有2个亚基,在果蝇和人中有3个亚基 TFIIA可以看成是一种TAFII(与TBP结合,
能稳定TFIID与启动子之间的结合) 在体外体系中,TFIIA并非必不可少
四、真核生物的基因转录及其调控
4. 真核生物的通用转录因子 (1)II类因子(class II factors) TFIIA与TFIIB的结构及功能
四、真核生物的基因转录及其调控
4. 真核生物的通用转录因子 (1)II类因子(class II factors) TFIIE和TFIIH的结构及功能
TFIIH
最后一个结合到预起始复合物的通用转录因子, 结构、功能均复杂
功能之一是使Pol II最大一个亚基的羧基末端域 (CTD)磷酸化,即使Pol IIA变为Pol IIO,从 而导致转录起始到转录延伸过渡
有些基因甚至没有TATA区
看家基因(housekeeping genes) 控制发育的基因
四、真核生物的基因转录及其调控
2. 真核RNA聚合酶识别的启动子 (1)RNA聚合酶II识别的启动子 起始子(initiator)
转录起始位点前后的保守序列 共同序列为:PyPyANT/APyPy
分子生物学
四、真核生物的基因转录及其调控
2. 真核RNA聚合酶识别的启动子 (1)RNA聚合酶II识别的启动子(II类启动子,
真核通用转录因子

• TAF的主要功能: • 与核心启动子元件相互作用 • 与激活因子相互作用 • 与不同激活因子的相互作用需要不同的TAF组合 • TAF1还具有组蛋白乙酰转移酶活性和蛋白激酶活性
11-14
TAF与核心启动子元件相互作用
• TFIID中的TAF因子可以帮助TBP促进聚合酶转录带有起始子和DPE的 启动子:
• 腺病毒E1B和E4启动子: 有TATA box,但无起始 子和下游启动子元件
• 腺病毒主要晚期(AdML) 启动子与果蝇Hsp70启动 子含有TATA box、起始 子和DPE
(upstream-binding factor, UBF)或上游激活因子(upstream activating factor,UAF)
11-28
11.3 III类因子
• III类因子包括结合在5S rRNA基因内部的转录因子TFIIIA以及另外两种 转录因子TFIIIB和TFIIIC
• 所有典型的III类基因的转录都需要TFIIIB和TFIIIC • 5S rRNA的转录需要TFIIIA、TFIIIB和TFIIIC
DABPolF复合物
DABPolFEH复合物
11-8
前起始复合物足迹分析
• DNase I足迹法结果显示DA和DAB保护TATA box
DA和DAB的足迹
DABPolF的足迹 11-9
DABPolF复合物形成的模型
• PolII与F结合 • DAB与TATA结合 • 两个复合物结合,形成DABPolF
• 该磷酸化对于转录起始是必 须的(但体外转录无需CTD 磷酸化)
分子生物学问题汇总

Section A 细胞与大分子简述复杂大分子的生物学功能及与人类健康的关系。
Section C 核酸的性质1.DNA的超螺旋结构的特点有哪些?A 发生在闭环双链DNA分子上B DNA双链轴线高卷曲,与简单的环状相比,连接数发生变化C 当DNA扭曲方向与双螺旋方向相同时,DNA变得紧绷,为正超螺旋,反之变得松弛为负超螺旋。
自然界几乎所有DNA分子超螺旋都为负的,因为能量最低。
2.简述核酸的性质。
A 核酸的稳定性:由于核酸中碱基对的疏水效应以及电荷偶极作用而趋于稳定B 酸效应:在强酸和高温条件下,核酸完全水解,而在稀酸条件下,DNA的核苷键被选择性地断裂生成脱嘌呤核酸C 碱效应:当PH超出生理范围时(7-8),碱基的互变异构态发生变化D 化学变性:一些化学物质如尿素,甲酰胺能破坏DNA和RNA二级结构中的而使核酸变性。
E 粘性:DNA的粘性是由其形态决定的,DNA分子细长,称为高轴比,可被机械力和超声波剪切而粘性下降。
F 浮力密度:1.7g/cm^3,因此可利用高浓度分子质量的盐溶液进行纯化和分析G 紫外线吸收:核酸中的芳香族碱基在269nm 处有最大光吸收H 减色性,热变性,复性。
思考题:提取细菌的质粒依据是核酸的哪些性质?质粒是抗性基因,,在基因组或者质粒DNA中用碱提取法。
Sectio C 课前提问1.在 1.5mL的离心管中有500μL,取出10 μL稀释至1000 μL后进行检测,测得A260=0.15。
问(1):试管中的DNA浓度是多少?问(2):如果测得A280=0.078, .A260/A280=?说明什么问题?(1)稀释前的浓度:0.15/20=0.0075稀释后的浓度:0.0075/100=0.75ug/ml(2)0.15/0.078=1.92〉1.8,说明DNA中混有RNA样品。
2.解释以下两幅图(native:非变性的;denatured:变性的)图一表示dsDNA和RNA的热变性,中间的Tm值表示解链温度。
分子生物学-转录因子

Independence of
DNA-binding domain and
transcription-activation domain
UASG
GAL4 protein :
a yeast activator
• a set of genes responsible for metabolism of galactose
(TBP); needed for RNApol I, II, III
TFIIA;
Binds to TFII D Enhances TFII D binding to TATA box Stabilizing the DNA-TFII D complex
TFII A TBP of D
Wilds minor groove
bridging factor
TFIIH; Large complex made up of > 5 subunits Helicase activity (ATPase, ) kinase activity Phosphorylation of CTD of RNApol IIO DNA repair
+ Cis-factor
4.3.3.2. RNA polymerase in Eukaryotes
RNApol. I, II, III
Including L, L’ subunit & 7-12 small subunits
L’ with 78%±homologous
between 3 RNApol. I, II, III
stimulate only a basal level of transcription.
4.3.3. 与基本转录相关的T.F.
真核生物转录元件组成及其分类

真核生物转录元件组成及其分类
真核生物转录元件是指在基因转录过程中与RNA聚合酶和调控因子结合的特定DNA序列,用于调控基因的转录。
它们包括启动子、增强子、沉默区和辅助区等。
1. 启动子:启动子位于基因的转录起始点上游,这是RNA聚合酶与DNA结合的起点。
启动子中一般包含TATA盒子和CAAT盒子等特定DNA序列,它们可以吸引转录因子与RNA聚合酶II结合并激活基因的转录。
2. 增强子:增强子位于启动子上游,它们可以增强启动子的活性,从而提高转录速率。
增强子一般由多个转录因子结合而成,它们可以通过相互作用来协同作用,实现调控基因的转录。
3. 沉默区:沉默区位于基因的转录起始点下游,它们可以阻止RNA聚合酶的结合,从而抑制基因的转录。
沉默区一般包括转录终止位点和多个转录抑制因子结合的DNA序列。
4. 辅助区:辅助区分布在基因的远端、内含子和外显子中,它们可以调节基因的表达水平和组织特异性。
辅助区包括增强子和增强子类似物,它们可以通过转录因子结合来调控基因的转录。
根据其作用和位置的不同,转录元件可以分为启动元件、增强元件和沉默元件等。
启动元件包括启动子和促进元件,主要用于启动基因的转录。
增强元件包括增强子和增强子类似物,主要用于增强基因的表达。
沉默元件包括沉默区和抑制元件,主要用于抑制基因的转录。
转录因子

转录因子基因转录有正调控和负调控之分。
如细菌基因的负调控机制是当一种阻遏蛋白(repressor protein)结合在受调控的基因上时,基因不表达;而从靶基因上去除阻遏蛋白后,RNA聚合酶识别受调控基因的启动子,使基因得以表达,这是正调控。
这种阻遏蛋白是反式作用因子。
而顺式作用因子则指的是基因上与反式作用因子结合的对基因表达起调控作用的基因序列。
转录因子(transcription factor)是起正调控作用的反式作用因子。
转录因子是转录起始过程中RNA聚合酶所需的辅助因子。
真核生物基因在无转录因子时处于不表达状态,RNA聚合酶自身无法启动基因转录,只有当转录因子(蛋白质)结合在其识别的DNA序列上后,基因才开始表达。
转录因子的结合位点(transcription factor binding site,TFBS)是转录因子调节基因表达时,与mRNA结合的区域。
按照常识,转录因子(transcription factor,TF)的结合位点一般应该分布在基因的前端,但是,新的研究发现,人21和22号染色体上,只有22%的转录因子结合位点分布在蛋白编码基因的5'端。
真核生物在转录时往往需要多种蛋白质因子的协助。
一种蛋白质是不是转录机构的一部分往往是通过体外系统看它是否是转录起始所必须的。
一般可将这些转录所需的蛋白质分为三大类:(1)RNA聚合酶的亚基,它们是转录必须的,但并不对某一启动子有特异性。
(2)某些转录因子能与RNA聚合酶结合形成起始复合物,但不组成游离聚合酶的成分。
这些因子可能是所有启动子起始转录所必须的。
但亦可能仅是譬如说转录终止所必须的。
但是,在这一类因子中,要严格区分开哪些是R NA聚合酶的亚基,哪些仅是辅助因子,是很困难的。
(3)某些转录因子仅与其靶启动子中的特异顺序结合。
如果这些顺序存在于启动子中,则这些顺序因子是一般转录机构的一部分。
如果这些顺序仅存在于某些种类的启动子中,则识别这些顺序的因子也只是在这些特异启动子上起始转录必须的。
真核生物基因的转录

(B’’, TBP, BRF)
TF III B
TF III A TF III C
Pol III
四、RNA 聚合酶 II 基因的转录
(一)RNA聚合酶 II 的启动子 1、组成:
核心启动子(core promoter): TATA盒(Hogness box): - 25 ~ -35bp
上游启动子(upstream promoter element,UPE) CAAT盒 :-70 ~ -80区 GC盒:-80 ~ -110区
TF II B —— 覆盖靠近起始点的启动位置,C端与TFIID和DNA 的复合物结合,N-端与TFⅡF协同作用募集RNA聚 合酶II。
TF II F ——结合Pol II并带向启动子;RAP74(ATP依赖性解 旋酶),RAP30(与细菌因子有同源性)
TF II E —— 扩大DNA覆盖区至+30
Module Consensus DNA bound Factor Distribution
TATA box TATAAAA
~10bp
CAAT box # GGCCAATC ~22bp
GC box
GGGCGG
~20bp
Octamer # ATTTGCAT
~20bp
``
``
23bp
B
GGGACTTTCC ~10bp
(2) TFIIA
▪ 含有至少3个亚基 ▪ 与TFIID结合,稳定TFIID-DNA复合体;可能通过解除
TAFs的抑制而激活TBP
TF II A
(3) TFIIB ▪ 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复
合物结合,N-端与TFⅡF协同作用募集RNA聚合酶II
真核生物的转录因子专项文档

真核生物的转录因子专项文档一、转录因子的定义和作用转录因子是一类控制基因表达的蛋白质,它们能够与基因的DNA序列结合,并调控基因的转录过程。
转录因子在真核生物中起着关键的作用,它们能够决定细胞的命运和特化,调控细胞周期和分化,以及响应环境刺激等。
二、转录因子的分类转录因子可分为两大类:顺式调控因子和择性调控因子。
顺式调控因子与DNA的特定序列结合并激活或抑制转录过程,而择性调控因子通过与其他蛋白质和共调控因子的相互作用来调控基因的表达。
顺式调控因子又分为激活基因表达的激活子和抑制基因表达的抑制子。
激活子通过与RNA聚合酶结合来促进转录的发生,而抑制子则与转录复合体的组分相互作用,阻碍转录的进行。
择性调控因子根据其调控机制的不同,可分为组蛋白修饰调控因子、染色体剪接调控因子和基因启动调控因子等。
组蛋白修饰调控因子通过修饰染色体上的组蛋白来调控基因的表达。
染色体剪接调控因子参与基因的剪接过程,影响基因的转录结果。
基因启动调控因子是一类与转录启动复合体相互作用的因子,它们能够促进或抑制转录的发生。
三、转录因子的结构和功能域转录因子的结构通常包括DNA结合结构域、转录激活结构域和转录抑制结构域等。
DNA结合结构域使得转录因子能够与基因的DNA序列结合。
转录激活结构域能够与转录启动复合体相互作用,并激活转录的发生。
转录抑制结构域则可以与其他转录因子或转录启动复合体相互作用,并抑制转录的进行。
四、转录因子的调控机制转录因子的调控机制非常复杂,涉及到信号转导通路、组蛋白修饰、DNA甲基化等多种调控方式。
转录因子的活性和稳定性可以通过磷酸化、去磷酸化、泛素化等化学修饰方式来调节。
此外,染色质的状态和结构也对转录因子的调控起着重要的作用。
五、典型的转录因子家族真核生物中存在许多转录因子家族,其中一些家族在不同物种中高度保守。
例如,家庭基因家族(homeobox gene family)编码一类在胚胎发育过程中起关键作用的转录因子。
真核生物的转录和后加工

– 隔断基因的线性表达而在剪接过程中被除去的 核酸序列。
鸡
卵
鸡卵清蛋
清
白基因
蛋
白
基
hnRNA
因
及
首、尾修饰
其 转
录
、
hnRNA剪接
转
录
后
成熟的mRNA
修
饰
3. 内含子的分类
I:主要存在于线粒体、叶绿体及某些低等真核生物 的 rRNA基因; II:也发现于线粒体、叶绿体,转录产物是mRNA; III:是常见的形成套索结构后剪接,大多数mRNA基
ppi
mRNA鸟苷酰转移酶 5` GpppN
pppG pi
mRNA
甲基化酶
(S-腺苷甲硫氨酸)CH3
5` m7GpppN
mRNA
注:帽子结构中G未甲基化,翻译效果差,但稳定性不变
帽子结构
3`-末端多聚腺苷酸的合成
• 先于剪接加工 • poly A polymerase 催化,转录后修饰
点序列(AAUAAA)提供信号 • 一般长度为100~200个腺苷酸
1. 转录起始前的上游区段
顺式作用元件(cis-acting element)
• 顺式作用元件是指与结构基因串联的特定DNA序列,是转录因子的结合位点,它们通 过与转录因子结合而调控基因转录的精确起始和转录效率。
AATAAA
OCT-1
翻译起始点
外显子
转录起始点
内
含
TATA盒
子
转录终止点
CAAT盒
GC盒
解聚现象。
•
核小体
转
录
延
RNA-Pol
长 转录方向
中
第八章 真核生物的转录

● 真 核 生 物 转 录 起 始 复 合 物
转录因子 TBP
转录复合体
TAFs
TFIIA TFIIB TFIIF Pol II
TFIIE
RNA pol Ⅱ的转录起始
结构基因
DNA
P
O
Z
Y
A
Z: β-半乳糖苷酶 Y: 透过酶
A:乙酰基转移酶
● 5’ 端无“帽子”结构, 3’ 端没有或只有较短
的poly(A )结构。
SD序列:mRNA中用于结合原核生物核糖体的序列。
2、真核生物mRNA的特征 “基因”的分子生物学定义:产生一条多肽链或 功能RNA所必需的全部核甘酸序列。 ● 5’ 端存在“帽子”结构 ●多数mRNA 3’ 端具有poly(A )尾巴(组蛋白除外) ●以单顺反子的形式存在
原核生物和真核生物mRNA结构的比较
五、RNA合成与DNA合成异同点
相同点:
1、都以DNA链作为模板
2、合成的方向均为5’→3’
3、聚合反应均是通过核苷酸之间形成的3’,5’-磷 酸 二酯键,使核苷酸链延长。
不同点:
模板 原料
酶 产物
复制 两条链均复制 dNTP
DNA聚合酶 子代双链DNA (半保留复制) A-T;G-C RNA引物
增强子的作用方式
2. 通用转录因子
能直接、间接辨认和结合转录上游区段
DNA的蛋白质,统称为反式作用因子(transacting factors) ,现已发现数百种 。 反式作用因子中,直接或间接结合RNA
某大学生物工程学院《生物化学》考试试卷(52)

某大学生物工程学院《生物化学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、判断题(95分,每题5分)1. 萌发的油料种子和某些微生物拥有乙醛酸循环途径,可利用脂肪酸α氧化生成的乙酰辅酶A合成苹果酸,为糖异生和其他生物合成提供碳源。
()答案:错误解析:萌发的油料种子和某些微生物拥有乙醛酸循环途径,利用乙醛酸循环将乙酰CoA转化成琥珀酸,由琥珀酸再生成草酰乙酸而进入生糖途径。
2. 脂酸的氧化降解是从分子的羧基端开始的。
()答案:正确解析:3. 线粒体内的翻译系统中,第一个被掺入的氨基酸都是甲酰甲硫氨酸。
()答案:错误解析:哺乳动物线粒体翻译的蛋白质第一个氨基酸不是甲酰甲硫氨酸。
4. 原核生物基因转录时,第一个磷酸二酯键形成以后,α因子与全酶解离,转录从起始状态转入延伸状态。
()答案:错误解析:实际上在合成大约十聚核苷酸以后,σ因子才与RNA pol全酶解离,RNA pol开始移位,转录进入延伸阶段。
5. 葡萄糖磷酸变位酶既参与糖原合成,又参与糖原分解。
()答案:正确解析:6. 脂肪酸彻底氧化产物为乙酰CoA。
()答案:错误解析:脂肪酸彻底分解的过程是先经β氧化作用生成若干个乙酰CoA,生成的乙酰CoA再由三羧酸循环氧化生成二氧化碳和水。
7. 糖酵解过程在有氧无氧条件下都可进行。
()答案:正确解析:8. 脂肪酸活化为脂肪酰CoA时,需消耗两个高能磷酸键。
()答案:正确解析:脂肪酸的活化过程需消耗1molATP,但消耗2个高能磷酸键,也相当于消耗2molATP。
9. 在正常情况下,人体蛋白质的合成与分解处于动态平衡。
()答案:正确解析:10. 反转录病毒都是肿瘤病毒。
()答案:错误解析:HIV是一种反转录病毒,但并不是肿瘤病毒。
11. 当溶液的pH升高时,ATP水解释放的自由能明显增高。
(武汉大学)分子13.转录知识点整理
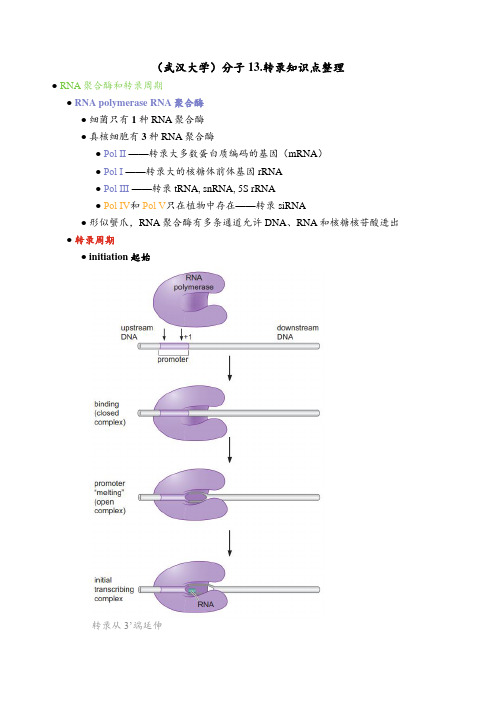
(武汉大学)分子13.转录知识点整理●RNA聚合酶和转录周期●RNA polymerase RNA聚合酶●细菌只有1种RNA聚合酶●真核细胞有3种RNA聚合酶●Pol Ⅱ ——转录大多数蛋白质编码的基因(mRNA)●Pol Ⅰ ——转录大的核糖体前体基因 rRNA●Pol Ⅲ ——转录tRNA, snRNA, 5S rRNA●Pol Ⅳ和Pol Ⅴ只在植物中存在——转录siRNA●形似蟹爪,RNA聚合酶有多条通道允许DNA、RNA和核糖核苷酸进出●转录周期●initiation 起始转录从3’端延伸●promoter 启动子:最初结合RNA聚合酶的DNA序列与所需的转录起始因子一起结合●closed complex 闭合式复合体聚合酶与启动子结合,DNA保持双链状态●open complex 开放式复合体在起始位点周围约14bp处解链形成转录泡(DNA解链)●initial transcribing complex 起始转录复合体聚合酶-启动子复合体,释放出短的(少于10个核苷酸)的转录物●elongation 延伸五个步骤:合成RNA,使前方的DNA解链,使其后的DNA重新复性,逐步解开正在延长的RNA链与模板的配对,校正●启动子逃离:一旦RNA聚合酶合成一小段RNA(10 个碱基左右)转录便进入了延伸阶段●聚合酶的构象进一步改变,使其与模板的结合更加紧密●termination 终止释放RNA产物,同时RNA聚合酶从DNA上释放●细菌的转录周期(原核生物的转录)●E.coli中σ factor识别的启动子(σ^{70} promoter)●-35&-10区域●特征序列:两段6个核苷酸长的保守序列●分隔序列:17~19个核苷酸(最适为17个)●consensus sequence 共有序列:有可能出现的最高频的首选●启动子的强度(其影响因素解释了为什么启动子之间差别如此之大)一个启动子在一定时间内起始转录物的数目●启动子最初与聚合酶的结合程度●对异构化作用的支持效率●此后聚合酶逃离的难易程度●UP-element额外与RNA聚合酶结合的DNA元件●通过提供额外的RNA 聚合酶特异性相互作用位点增强 RNA 聚合酶与DNA的结合●存在于一些较强的启动子中●直接被RNA聚合酶的αCTD识别●延长的-10区域补偿了-35 domain的缺失●discriminator 鉴别子紧挨着-10 domain的下游DNA元件●鉴别子与聚合酶之间相互作用的强度影响酶与启动子复合物的稳定性●E.coli σ factor——σ^{70}赋予RNA聚合酶识别启动子的特性,全酶=核心酶+σ因子●σ 因子与核心RNA聚合酶结合,与启动子上的不同元件可以发生相互作用●区域4识别-35元件区域4的两个螺旋形成helix-turn-helix螺旋-转角-螺旋结构其中一个螺旋插入到-35 区的大沟中并与该区的碱基相互作用另一个则从大沟的顶部横过,与DNA 骨架接触●区域2识别-10元件●区域3识别延长的-10元件●区域1.2识别鉴别子●UP-element不被σ factor所识别,而是被RNA聚合酶的αCTD结构域识别σ 因子和α 亚基将RNA 聚合酶核心酶招募到启动子上●转录起始●closed complex 闭合式复合体向open complex 开放式复合体的转变涉及和 DNA启动子和 RNA聚合酶的isomerization 异构化●启动子DNA的变化●发生在-11和+3之间的双链打开●聚合酶结构的显著变化●不需要水解ATP获得能量,自发进行●聚合酶前部的β 和β’ 钳子牢固地钳压在下游DNA 上●σ 因子上区域1.1的位置的移动σ_{1.1}是molecular mimic分子模拟物:既可以被区域1.1占据,也可以被DNA占据●未结合DNA 时,σ_{1.1}处于全酶的活性中心裂隙内部●在开放式复合体中,σ_{1.1}出现在酶的外部,使DNA 可以进到裂隙中●要求●不需要引物●起始核糖核苷酸(通常是一个A)被带到活性位点●聚合酶必须与起始核苷酸或第二个核苷酸或两者都建立特异性的相互作用●启动子逃离(起始向延伸的转变)●聚合酶-启动子相互作用的破坏●聚合酶核心- σ相互作用的破坏σ因子常常从延伸复合体中逐出(在延伸阶段不需要σ因子)●流产起始聚合酶重复多次合成并释放短的(9 个核苷酸或更短)RNA 分子●原因:σ因子的3/4 连接区(3.2)堵塞了RNA出口,需要RNA聚合酶尝试多次才能将其逐出●转录延伸●合成过程●双链DNA在两个钳子之间进入聚合酶●在催化裂隙的开口处DNA 两条链分开●核苷酸添加●RNA 产物离开在任何一个特定时刻RNA 链中只有8个或9个核苷酸与DNA模板保持碱基互补配对状态●DNA 两条链在转录后重新形成双螺旋结构●proofreading 校正●校正机制●pyrophosphorolytic editing 焦磷酸化编辑(焦磷酸水解的逆反应)●聚合酶利用它的活性位点在一个简单的逆向反应中通过重新加入PPi,催化错误掺入的核苷酸的去除●可以去除错误和正确的碱基,但更容易去除错误碱基●hydrolytic editing 水解编辑●聚合酶倒退一个或更多个核苷酸并切割RNA 产物,去除包含错误的序列●转录终止●异常终止●损坏的DNA链使RNA聚合酶停滞不前●TRCF蛋白——挪走聚合酶并同时招募修复酶具有ATPase活性,沿着DNA移动,遇到停止的RNA 聚合酶时,碰撞把聚合酶往前推●使聚合酶重新开始延伸●使聚合酶-DNA-RNA转录三重复合体解体结束该转录,给修复酶让位●正常终止在基因末端由terminator 终止子引发的正常过程●Rho-dependent terminator Rho依赖型终止子●需要Rho蛋白诱发终止●具有6个相同亚基的环状蛋白,在单链RNA 离开聚合酶时与之结合●有ATPase 活性,利用水解ATP的能量诱发终止●rut (Rho utilization) 位点Rho的特异性结合位点●几段约 40 nt的序列●不折叠为二级结构(保持单链状态)●富含C残基●无法与任何正在被翻译的转录产物结合(进入核糖体的产物)原核生物中转录和翻译是偶联的,Rho 只终止已超出某个基因或operon 操纵子末端但仍在进行的转录●Rho-independent terminator Rho非依赖型终止子 (intrinsic terminator 固有终止子)以RNA形式起作用(转录后才起作用)●短的反向重复序列(约20个核苷酸)●一段约8个 A-T 碱基对的序列转录为一段后面跟着 A/U 碱基对的发卡结构●发夹的形成破坏延伸复合体-终止●尾部序列使RNA更容易脱离下来●真核生物的转录●转录起始的条件●general transcription factor, GTF 通用转录因子真核生物中有效的、启动子特异的转录起始所需的因子●体外:通用转录因子和RNA聚合酶Ⅱ构成,直接从DNA模板转录●体内:除GTF外,还需额外的因子参与例如mediator complex 中介蛋白复合体、DNA调节蛋白●启动子进行有效转录的序列元件(顺式作用元件)●core promoter 核心启动子在体外检测时,Pol Ⅱ 机器精确起始转录所需的最少一组序列元件●TFⅡB识别元件(TFⅡB recognition element, BRE)●TATA元件●起始子 (initiator, Inr)●下游启动子元件 (downstream promoter element, DPE)包括DPE下游启动子元件, DCE下游核心元件, MTE基序十元件●特征:通常一个启动子只含有这些元件之中的几个,比如启动子通常含有TATA元件或DPE,而不是两者兼备●regulatory sequence 调节序列在核心启动子之外的(通常是在其上游)且在体内进行有效转录所需的其他序列元件●增强转录——promoter proximal element 启动子最近元件、upstreamactivator sequence, UAS 上游激活物序列、enhancer 增强子●抑制转录——silencer 沉默子、isulator 绝缘子●所有这些元件都与调控蛋白结合以发挥其功能●转录起始●pre-initiation complex 前起始复合体的形成TFⅡD中的TBP结合于TATA 元件,TFⅡA、TFⅡB 被招募 RNA聚合酶Ⅱ-TFⅡF 被招募TFⅡE、TFⅡH(介导ATP 水解产生的能量使启动子解旋)被招募,前起始复合体形成 CTD 尾巴磷酸化后逃离启动子●定义:结合在启动子上准备开始转录的一套完整的通用转录因子和聚合酶形成的复合体●形成位置:TATA元件●识别和建立前起始复合体的必需因子——TFⅡD●TATA binding protein, TBP TATA元件结合蛋白●能结合DNA,使TATA元件扭曲●TBP-DNA 复合物招募其他通用转录因子和聚合酶●TBP 相关因子——TAF,识别其他核心启动子元件●其他通用转录因子●TBP-associated factor, TAFTBP 与大约10 个TAF 结合●2个TAF结合启动子处DNA元件 (Inr, DPE)●几个TAF以类似组蛋白的方式结合DNA●1个TAF 调控TBP 与DNA 的结合●TFⅡB单链●在TBP 之后进入前起始复合体形成TFⅡB-TBP-DNA complex●与上游BRE 的大沟和下游TATA 的小沟的碱基特异性相互作用●与TBP-TATA 复合物的非对称结合,导致后续前起始复合体的不对称组装,继而导致单向转录●与Pol Ⅱ 接触,是连接结合了TATA的TBP 与聚合酶之间的桥梁●TFⅡF双亚基因子,与Pol Ⅱ 结合并一起被招募到启动子上●稳定DNA-TBP-TFⅡB 复合物●TFⅡE:招募和调控TFⅡH两个亚基●TFⅡH含有10个亚基,其中两个是ATPase,一个是蛋白激酶,是最大的通用转录因子●控制ATP 依赖的前起始复合体向开放式复合体的转变●对于启动子解链和启动子逃离有重要作用●启动子逃离——RNA聚合酶Ⅱ的尾巴CTD磷酸化(起始向延伸的转变)包含多个重复的七肽序列:Tyr-Ser-Pro-Thr-Ser-Pro-Ser 可以由TFⅡH 的一个亚基作为激酶而诱导●摆脱大部分起始因子(GTF 通用转录因子、中介蛋白)●招募延伸因子(TFⅡS 和hSPT5)●招募RNA 加工因子●体内转录需要额外蛋白质原因:DNA 模板在细胞内包装在核小体和染色质内通过activator 激活因子(转录调控蛋白)招募蛋白质●中介蛋白复合物(激活因子招募蛋白复合物)●核小体修饰和重塑因子●转录激活因子●转录延伸●延伸RNA聚合酶在行进过程中必须处理组蛋白●facilitates chromatin transcription, FACT 促进染色体转录因子●Spt16:结合H2A-H2B●SSRP1:结合H3-H4●SPT6:与H3结合,辅助装载组蛋白●RNA加工●RNA capping 5’ 端加帽●保护mRNA 不被核酸外切酶降解,提高转录效率●帮助mRNA 运输到细胞质●通过5’ 端帽子剪接第一个内含子●RNA polyadenylation 3’ 端多聚腺苷酸化与转录终止密切相关●保护mRNA 不被核酸外切酶降解,提高转录效率●聚合酶的CTD 尾巴参与招募多聚腺苷酸化必需的酶●Poly-A信号序列的转录使得多聚腺苷酸化发生CPSF(切割和聚腺苷酸化特异性因子)和CstF(切割刺激因子)结合导致mRNA切割Poly-A Polymerase (PAP) 以ATP 为底物,在mRNA 3’端添加约200 个A 被Poly-A结合蛋白结合并稳定转录终止●RNA splicing 剪接去除内含子(分子14.※RNA剪接)●转录终止●鱼雷模型最后转录出没有帽子的RNA 分子被高延伸性的RNase(Rat1/ hXm2,鱼雷)识别和降解,使得PolⅡ 从DNA 模板上解离下来●变构模型PolⅡ诱导构象改变,使得其延伸性降低,最终自发终止●由RNA聚合酶Ⅰ和Ⅲ 催化的转录●RNA聚合酶Ⅰ——转录 rRNA前体基因●启动子●core promoter 核心启动子●UCE 上游控制元件●起始需要Pol Ⅰ、UBF和SL1●UBF 和UCE 结合,将 SL1 带到核心启动子区●SL1 通过招募Pol Ⅰ ,从核心启动子激发转录●RNA聚合酶Ⅲ——转录 tRNA基因、snRNA基因和 5S rRNA基因●核心启动子(Box A、B或 A、C)位于转录起始位点下游tRNA需要转录因子TFⅢC和TFⅢB 5S rRNA还需TFⅢA(结合在Box A上)●本章名词●RNA聚合酶和转录周期●ribonucleotide●transcription●initiation●closed complex●open complex●initial transcribing complex●elongation●termination●promoter●细菌的转录周期●起始●consensus sequence●discriminator●helix-turn-helix●isomerization●molecular mimic●延伸●pyrophosphorolytic editing●hydrolytic editing●终止●intrinsic terminator●operon●真核生物的转录●general transcription factor, GTF●core promoter●regulatory sequence●TFIIB recognition element, BRE●initiator, Inr●downstream promoter element●pre-initiation complex●TATA binding protein, TBP●TBP-associated factor, TAF●carboxy-termical domain, CTD●activator●elongation factor●facilitates chromatin transcription, FACT●RNA capping●RNA splicing●RNA polyadenylation●由RNA聚合酶I和III催化的转录●snRNA●重点知识点●转录与复制的区别①转录得到的新链是由核糖核苷酸组成的,而不是脱氧核糖核苷酸;②RNA 聚合酶不需要引物,能够从头起始转录,但必须在特定的序列上才能起始;③RNA 产物不与模板DNA 链的碱基保持互补状态;④转录不如复制更为精确;⑤转录仅是有选择性地复制基因组的特定部分,并从任何特定的部分产生一个到几百个,或者甚至上千个拷贝。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
另一锌指结构是锌离子与4个半 胱氨酸结合,它出现在一百多种类 固醇激素受体转录因子中。这些因 子由同型或异型的二聚体组成,其 中每一单体包含2个C4锌指结构。 两个单体通锌离子稳定折叠成更 复杂的构象,再把每个单体的α-螺 旋插入到DNA的连续大沟中。
13
14
15
3. 碱性结构域
在许多DNA结合蛋白中都发现 了碱性结构域,它通常是与亮氨酸 拉链或HLH基序中的一个联合在一 起的,结果被称做碱性亮氨酸拉链 (bZIP)或碱性HLH蛋白。蛋白的二 聚作用使二个碱性结构城相邻,进 而可与DNA发生作用。
domain: A recognition α-helix interacts with the DNA and is separated from anotherαhelix by a characteristic right angle β-turn. The zinc finger domain exists in two forms. The C2H2 zinc finger has a loop of 12 amino acids anchored by two cysteine and two histidine residues that tetrahedrally co-ordinate a zinc ion. Usually three or more C2H2 zinc fingers are required for DNA binding. C4 zinc finger in which the zinc ion is co-ordinated by four cysteine residues. The basic domain is generally associated with one or other of two dimerization domains, the leucine zipper or the helix-loophelix(HLH) motif)
2
反式作用因子是能直接或 间接地识别或结合在各类顺式 作用元件核心序列上,参与调 控靶基因转录效率的蛋白质。
3
转录因子结构域结构
转录因子结构域结构,转录因子能特异性结合 DNA并激活转录,由两个结构域:DNA结合结构 域和激活结构域。有些转录因子还有二聚体结构域 (Transcription factor domain structure, transcription factors have two distinct activities and two domains, They bind specifically to their DNA-binding site (DNA-binding domains) and activate transcription. (activation domains). Some transcription factors have dimerization domains)
4
转录因子结构域结构 DNA结合结构域 转录激活结构域 二聚体结构域
5
Eukaryotic activators (真核激活蛋白) have separate DNA binding and activating functions
6
Eukaryotic regulators use a range of
9
10
11
2. 锌指结构域
型锌这指种通结过构2个域半有胱两氨种酸形和式2,个“组C氨2H酸2” 残基固定,这四个残基与锌离子在空间 上形成一个四面体结构。这种锌指折叠 形成一个紧密的结构,由二条β链和一 个α-螺旋组成,α-螺旋与DNA大沟结 合。该α-螺旋区域上含有保守的碱性氨 基酸,负责与DNA的结合。
7
The helix-turn-helix domain The zinc finger domain The basic domain:Leucinezipper(bZIP) or
helix-loop-helix(HLH)
8
DNA结合结构域
1. 螺旋-转折-螺旋(helix-turn-helix, H-T-H)结构 这一类蛋白质分子中有至 少两个α螺旋,中间由短侧链氨基酸残 基形成“转折”,近竣基端的α螺旋中 氨基酸残基的替换会影响该蛋白质在 DNA双螺旋大沟中的结合。与DNA相 互作用时,同源域蛋白的第一、二两个 螺旋往往靠在外侧,其第三个螺旋则与 DNA大沟相结合,并通过其N-端的多余 臂与DNA的小沟相结合。
DNA binding domains(DNA结合结构域)
DNA结合结构域,螺旋-转角-螺旋结构域: 两个α-螺旋由一个β-转角 分开,识别的α-螺旋与DNA发生作用。锌指结构域,有两种:C2H2 型锌指有一个12氨基酸的环,通过2个半胱氨酸和2个组氨酸残基固 定与锌离子形成4面体结构,一般需要3个或更多的C2H2型锌指才能 与DNA结合;C4锌指结构由4个半胱氨酸与锌离子结合而成。碱性 结构域,通常与亮氨酸拉链或HLH基序中的一个联合一起,又称碱 性亮氨酸拉链或碱性HLH 蛋白(DNA-binding domains, The helix-turn-helix
16
二聚体结构域
二聚体结构域, 亮氨酸拉链和螺旋-环-螺 旋结构域(Dimerization domains include leucine zippers and the helix-loop-helix domain)
17
二聚体结构域
1. 亮氨酸拉链 亮氨酸拉链的肽链上每 相隔七个残基就会有一个疏水的亮氨酸 残基,这些残基位于DNA结合域的C端α螺旋上,这样α-螺旋的侧面每两圈就会 出现一个亮氨酸,形成一个疏水的表面。 结果在α-螺旋的疏水表面间就可以互相 作用,形成二聚体。这种相互作用形成 一个卷曲的卷曲结构 (coiledcoilstructure)。
Eukaryotic transcruption factors(真核生物的转录因子)
1
真核生物启动子和增强子是由若干 DNA序列元件组成的,由于他们常与特 定的功能基因连锁在一起,因此被称为顺 式作用元件。这些序列组成基因转录的调 控区,影响基因的表达。在转录调控过程 中,除了需要调控区外,还需要反式作用 因子