第六章 基因表达调控
第六章 基因调控1:原核生物基因的表达调控 分子生物学习题

第六章基因表达调控1:原核生物基因表达调控名词解释:操纵子基因表达持家基因正调控和负调控安慰诱导物衰减子(弱化子)魔斑结构基因和调节基因本底水平表达填空1 操纵子的基因表达调节系统属于水平的调节,乳糖操纵子模型由和1961年提出的。
色氨酸操纵子包括和两方面的调控。
2、能够诱导操纵子但不是代谢底物的化合物称为诱导物。
能够诱导乳糖操纵子的化合物就是其中一例。
这种化合物同蛋白质结合,并使之与分离。
乳糖操纵子的体内功能性诱导物是。
3、色氨酸是一种调节分子,被视为。
它与一种蛋白质结合形成。
通过控制起作用。
色氨酸操纵子受另一种系统------ 的调控,它涉及到第一个结构基因被转录前的转录。
4、大肠杆菌乳糖操纵子调节基因编码的与结合,对Lac结合,对Lac表达实施负调控;与复合物结合于上游部分,对Lac表达实施正调控。
5、操纵子中没有基因产物的是和选择题1、下面哪些真正是乳糖操纵子的诱导物?()A.乳糖B.蜜二糖C.O- 硝基苯酚-β-半乳糖苷(ONPG)D.异丙基-β-半乳糖苷E. 异乳糖2、色氨酸操纵子的调控作用是受两个相互独立的系统控制的,其中一个需要前导肽的翻译,下面哪一种调控这个系统?()A.色氨酸B.色氨酰-tRNA TrpC.色氨酰-tRNAD.cAMPE.以上都不正确3、阻遏蛋白(阻抑蛋白)识别操纵子中的()A 启动基因B 结构基因C 操纵基因D 内含子E 外显子4、乳糖、阿拉伯糖、色氨酸等小分子物质在基因表达调控中作用的共同特点是A 与启动子结合B 与DNA结合影响模板活性C 与RNA聚合酶结合影响其活性D 与蛋白质结合影响该蛋白质结合DNAE 与操纵基因结合5.下面那项不属于原核生物操纵元的结构A:启动子B:终止子C:操纵子D:内含子6、下列有关操纵子的论述哪个是错误的()A 操纵子是由启动基因、操纵基因与其所控制的一组功能上相关的结构基因组成的基因表达调控单位B 操纵子不包括调节基因C 代谢底物往往是该途径的可诱导酶的诱导物,代谢终产物往往是可阻遏酶的辅阻遏物D 真核细胞的酶合成也存在诱导和阻遏现象,因此也是由操纵子进行调控的7、操纵子调节系统属于哪一种水平的调节?A 复制水平的调控B 转录水平调控C 转录后加工的调控D 翻译水平的调控8、对调节基因下述哪些论述哪些是对的()A 是编码阻遏蛋白的结构基因B 各种操纵子的调节基因都与启动基因相毗邻C 调节基因是操纵子的组成部分D 调节基因的表达另有转移的调控区9、以下有关阻遏蛋白的哪些是对的()A 阻遏蛋白是调节基因表的的产物B 可诱导操纵子的阻遏蛋白具有直接与操纵子基因结合的活性,与诱导物相互作用后丧失活性C 可阻遏操纵子的阻遏蛋白没有直接与操纵子基因结合的活性,与辅阻遏物结合后才有此活性D 阻遏蛋白可与RNA聚合酶竞争同一结合部位10、关于启动基因的下述论点哪些是错误的()A 启动基因是RNA聚合酶识别并最县结合的一段DNA序列B 启动基因是最先被RNA聚合酶转录的DNA 序列C 启动基因是DNA上富含A-T碱基对的部分D 启动基因是引发DNA复制的特殊序列11、下列有关降解物基因活化蛋白(CAP)的哪些论点是正确的()A CAP-cAMP可专一地与启动基因结合,促进结构基因的转录B CAP可单独与启动子相互作用,促进转录C CAP-cAMP可与调节基因结合,控制阻遏蛋白的合成D CAP-cAMP可与RNA聚合酶竞争地结合于启动基因,从而阻碍结构基因的转录12、与乳糖操纵子操纵基因结合的物质是()A RNA聚合酶B DNA聚合酶C 阻遏蛋白D 反密码子是非题1、葡萄糖和乳糖并存时,细菌优先利用乳糖并启动乳糖操纵子()2、小分子物质如ITPG诱导乳糖操纵子表达时起负调控作用与操纵基因相结合阻抑结构基因的表达()3、色氨酸操纵子中含有衰减子区,其调控作用主要受Trp浓度高低影响()4、色氨酸操纵子(trpoperon)中含有衰减子序列()5、cAMP在laz 操纵子中起正调控作用,其浓度受环境中的葡萄糖影响,与其浓度成正比()6、大肠杆菌乳糖操纵子真正的诱导物不是乳糖,而是它的异构体别乳糖()7、操纵基因又称操纵子,如同启动基因又称启动子一样()8、可诱导操纵子是负责调节糖分解代谢的,可阻遏操纵子是负责调节氨基酸代谢的()问答题:1、试述乳糖操纵子的结构及负控诱导的调控机理2、色氨酸操纵子的结构特点?其弱化子在调控中如何起作用?3、简述色氨酸操纵子中启动子调控作用特点。
分子生物学习题集答案

第六章基因表达调控自测题(一)选择题A型题1.关于基因表达调控的说法错误的是・・A.转录起始是调控基因表达的关键B.环境因素影响管家基因的表达C.在发育分化和适应环境上有重要意义D.表现为基因表达的时间特异性和空间特异性E.真核生物的基因表达调控较原核生物复杂的多2.下列哪项属于可调节基因A.组蛋白编码基因8.5S rRNA编码基因C.异柠檬酸脱氢酶编码基因D.肌动蛋白编码基因E.血红蛋白编码基因3.与a-酮戊二酸脱氢酶系协调表达的是A.肉毒碱脂酰转移酶IB.柠檬酸合成酶C.丙酮酸竣化酶D.葡萄糖-6-磷酸酶E. HMG-CoA合成酶4.乳糖操纵子中,能结合别位乳糖(诱导剂)的物质是A.AraCB.cAMPC.阻遏蛋白D.转录因子E. CAP5.乳糖操纵子模型是在哪个环节上调节基因表达A.复制水平B.转录水平C.转录后水平D.翻译水平E.翻译后水平6.乳糖操纵子的调控方式是A. CAP的正调控B.阻遏蛋白的负调控C.正、负调控机制不可能同时发挥作用D. CAP拮抗阻遏蛋白的转录封闭作用E.阻遏作用解除时,仍需CAP加强转录活性7.与分解代谢相关的操纵子模型中,存在分解代谢物阻遏现象,参与这一调控的主要作用因子是A.阻遏蛋白B. AraCC.衰减子D. cAMP-CAP 复合物E.诱导剂8.原核细胞中,识别基因转录起始点的是A.阻遏蛋白B.转录激活蛋白C.基础转录因子D.特异转录因子E.。
因子9.使乳糖操纵子实现高表达的条件是A.乳糖存在,葡萄糖缺乏B.乳糖缺乏,葡萄糖存在C.乳糖和葡萄糖均存在D.乳糖存在E.葡萄糖存在10.大肠杆菌可以采用哪种方式调控转录终止A.阻遏作用B.去阻遏作用C.反义控制D.衰减作用E.降低转录产物的稳定性11.关于色氨酸操纵子错误的描述是・・A.核蛋白体参与转录终止B.衰减子是关键的调控元件C.色氨酸不足时,转录提前终止D.转录与翻译偶联是其转录调控的分子基础E.色氨酸存在与否不影响先导mRNA的转录12.下列哪种因素对原核生物的翻译没有影响• •A. microRNAB.稀有密码子所占的比例C. mRNA的稳定性D.反义RNAE.调节蛋白结合mRNA13.真核生物基因表达调控的关键环节是①染色质活化②转录起始③转录后加工④翻译起始⑤翻译后加工A.①+②+③B.①+②+④C.①+②D.② +③E.②14.下列哪种染色质结构的变化不利于基因表达 .A.组蛋白乙酰化B.核小体解聚C. CpG岛甲基化D.基因扩增E.染色质结构松散,对DNA酶I敏感15.下列哪项不属于真核生物基因的顺式作用元件 .A.激素反应元件B.衰减子C.启动子D.沉默子E.增强子16.与RNA聚合酶相识别和结合的DNA片段是A.增强子B.衰减子C.沉默子D.操纵子E.启动子17.下列哪项不参与调控真核细胞基因的特异性表达 .A.反应元件B.特异转录因子C.增强子D.基础转录因子E.沉默子18.与原核生物相比较,真核生物的基因表达调控包括多个层次,下列哪项不是. 其调控复杂性特有的分子基础A.含有重复序列B.断裂基因C.转录与翻译分离D.细胞内被膜性结构分隔形成多个区域E.染色质结构19.能够与基础转录因子结合的是A.上游启动子元件B. TATA boxC. 增强子D.反应元件E. Pribnow box20.有关基础转录因子的叙述,正确的是A.与非转录核心序列相结合B.决定基因表达的特异性C.其种类和数量在不同组织中差别很大D.辅助RNA聚合酶结合启动子E.在原核生物中的种类比真核生物少21.不属于特异转录因子的是 .A.TF II DB. HSFC. APID.类固醇激素受体E. NF-K B22.关于特异转录因子的说法,正确的是A.调控管家基因的表达B.仅通过蛋白质一蛋白质相互作用进行调控C.仅通过DNA一蛋白质相互作用进行调控D.仅通过RNA一蛋白质相互作用进行调控E.起转录激活或者转录抑制作用23.锌指结构可能存在于下列哪种物质中A.阻遏蛋白B.RNA聚合酶C.转录因子D.端粒酶E.核酶24.下列哪种氨基酸在转录因子的转录激活结构域中含量丰富A. LysB. ArgC. AspD. HisE. Trp25.下列哪种因素不会影响真核细胞中mRNA的稳定性 .A.5'端帽子B.siRNAC.poly A 尾D.去稳定元件E. miRNA26.小干扰RNA调节基因表达的机制是A.封闭mRNA上的核蛋白体结合位点B.特异性降解靶mRNAC.形成局部双链,抑制靶mRNA的模板活性D.使翻译出的蛋白质进入泛素化降解途径E.使翻译提早终止27. eIF-2对翻译起始具有重要的调控作用,下列哪项是它的活性形式A.磷酸化B.脱乙酰化C.乙酰化D.脱磷酸化E. ADP-核糖基化28.不影响真核生物翻译起始的因素是 .A. eIFB.帽子结合蛋白C.RNA编辑D.mRNA非翻译区的二级结构E.miRNA29.原核生物中,某种代谢途径相关的几种酶类往往通过何种机制进行协调表达A.顺反子B.操纵子C.转录因子D.衰减子E. RNAi30.生物体在不同发育阶段,蛋白质的表达谱也相应变化,这主要取决于A.转录调控元件的差异B.翻译调控元件的差异C.基础转录因子的差异D.特异转录因子的差异E.翻译起始因子的差异31.原癌基因通常是A.管家基因B.可调节基因C.突变的基因D.表达增强的基因E.促进凋亡的基因32.管家基因编码的产物不包括 .A.细胞外生长因子B.电子传递链的成员C.细胞的主要结构蛋白D.转录因子E.血红蛋白33.下列哪项不是可调节基因的特点 .A.组织特异性B.阶段特异性C.时间特异性D.空间特异性E.组成性表达34.关于操纵子的说法,正确的是A.几个串联的结构基因由一个启动子控制B.几个串联的结构基因分别由不同的启动子控制C. 一个结构基因由不同的启动子控制D.转录生成单顺反子RNAE.以正性调控为主35.核蛋白体调控转录终止的典型例子是A.乳糖操纵子B.半乳糖操纵子C.阿拉伯糖操纵子D.色氨酸操纵子E.以上都不对36.色氨酸操纵子的显著特点是A.阻遏作用B.诱导作用C.衰减作用D.分解物阻遏作用E.抗终止作用37.关于色氨酸操纵子的调控,正确的说法是A.色氨酸存在时,仅生成前导mRNAB.色氨酸不足时,转录提前终止C.是翻译水平的调控D.具有抗终止作用E.依赖P因子进行转录调控38.细菌优先利用葡萄糖作为碳源,葡萄糖耗尽后才会诱导产生代谢其他糖的酶类,这种现象称为A.衰减作用B.阻遏作用C.诱导作用D.协调调节作用E.分解物阻遏作用39.关于分解物阻遏的作用机制,说法正确的是A.葡萄糖缺乏时,cAMP浓度低B.葡萄糖缺乏时,CAP浓度低C.葡萄糖缺乏时,cAMP不能与CAP形成复合物D.葡萄糖缺乏时,cAMP-CAP复合物浓度高E.葡萄糖缺乏时,cAMP-CAP复合物失去DNA结合能力40.大肠杆菌的乳糖操纵子模型中,与操纵基因结合而调控转录的是A.阻遏蛋白B. RNA聚合酶C.调节基因D. cAMP-CAPE.启动子41.翻译终止阶段,新生多肽链的释放涉及哪种化学键的断裂A.肽键B.磷酸二酯键C.氢键D.疏水键E.酯键42. IPTG诱导乳糖操纵子表达的机制是A.使乳糖一阻遏蛋白复合物解离B.与阻遏蛋白结合,使之丧失DNA结合能力C.与乳糖竞争结合阻遏蛋白D.与RNA聚合酶结合,使之通过操纵序列E.变构修饰RNA聚合酶,提高其活性43.下列哪一项是真核生物可调节基因的表达调控特有的机制A.基础转录因子B.衰减子C. RNA聚合酶D.增强子E.阻遏蛋白44.基础转录因子属于DNA结合蛋白,它们能够A.结合转录核心元件B.结合增强子C.结合5'端非翻译区D.结合3'端非翻译区E.结合内含子45.特异转录因子不能够 .A.结合RNA聚合酶B.结合基础转录因子C.结合其他特异转录因子D.结合转录非核心元件E.结合沉默子 46.基因特异性表达的根本机制是A.顺式作用元件的种类不同B.RNA聚合酶活性的差异C.基础转录因子的质和量的差异D.特异转录因子的质和量的差异E.表达产物后加工过程的差异47.下列哪一类分子常具有亮氨酸拉链的结构特征A.生长因子B.酪氨酸蛋白激酶受体C.G蛋白D.转录因子E.丝/苏氨酸蛋白激酶B型题A.阻遏蛋白B. AraCC.衰减子D. CAPE. P因子48.葡萄糖缺乏时,细菌中cAMP浓度升高,可以结合49.实验室常使用IPTG作为诱导剂,其作用是结合50.阿拉伯糖操纵子的主要调节因子是51.色氨酸操纵子的调节作用依赖于A.转录起始B.转录终止C.翻译起始D.翻译终止E. mRNA稳定性52.原核基因表达调控的最基本环节是53.真核基因表达调控的最基本环节是A. eIF-2B.特异转录因子C. EF-2D.基础转录因子E. eRF54.真核细胞中管家基因的转录需要55.真核细胞中可调节基因的转录主要取决于56.真核细胞中参与翻译起始复合物形成的是57.绿脓杆菌外毒素抑制真核细胞蛋白质合成的靶点是58.真核细胞中识别终止密码子的是A.阻遏蛋白B.转录因子C. cAMP-CAPD.终止因子E.核蛋白体59.色氨酸操纵子的转录调控依赖于60.乳糖缺乏时,乳糖操纵子不转录,这主要取决于61.葡萄糖缺乏时,乳糖操纵子转录,这主要取决于A.基础转录因子B.特异转录因子C.起始因子D.阻遏蛋白E. P因子62.人血红蛋白表达特异性的决定因素是63.小鼠异柠檬酸合成酶的表达需要哪一类蛋白质因子辅助64.大肠杆菌B一半乳糖甘酶表达的关键调控因素是X型题65.管家基因的转录受哪些因素控制A.基础转录因子B.增强子C.特异转录因子D.启动子E.反应元件66.大肠杆菌乳糖操纵子中,属于调控元件的是A.操纵基因B.调节基因C.启动子D.阻遏蛋白E. CAP结合位点67.色氨酸操纵子模型中,哪些因素参与调控作用A.释放因子B.核蛋白体C. CAP复合物D.阻遏蛋白E.衰减子68.原核生物转录终止的调控机制涉及A. RNA干扰B.分解物阻遏C. P因子D.衰减E.抗终止69.真核生物的基因表达调控表现在A.转录水平B.翻译水平C.染色质水平D.转录后加工E.翻译后加工70.真核生物中,影响RNA聚合酶转录活性的因素包括A.启动子B.增强子C.基础转录因子D.衰减子E.特异转录因子71.真核生物基因表达的空间特异性的机制包括A.特异转录因子的种类不同B.同种特异转录因子的浓度不同C.特定组织的基因中存在组织特异性启动子D.特异转录因子的排列组合不同E.增强子等调控元件在不同组织的基因中分布不同72.转录因子的DNA结合结构域包含哪些结构类型A.螺旋一片层一螺旋B.锌指C.螺旋一转角一螺旋D.亮氨酸拉链E.螺旋一环一螺旋73.与siRNA相比较,miRNA的显著特点是A.单链B.在转录后水平发挥作用C.与靶mRNA碱基互补D.不降解靶mRNAE.个别碱基与靶mRNA序列不完全匹配74.转录因子的作用机制包括A.DNA — DNA相互作用B.DNA — RNA相互作用C.DNA 一蛋白质相互作用D.RNA 一蛋白质相互作用E.蛋白质一蛋白质相互作用75.真核生物的基因转录涉及哪些物质的相互作用A. operatorB.cis-acting elementC.polysomeD.trans-acting factorE.RNA polymerase76.在同一个体的不同组织中A.基因的表达谱不同B.基因组结构不同C.特异转录因子的种类不同D.存在的蛋白质的种类不同E.特异性启动子的种类不同77.真核生物独有的转录调控机制涉及A.启动子B.增强子C.转录因子D.组蛋白E. SD序列78.生物对环境的适应性表现在A.基因变异B.合成不同种类的mRNAC.合成不同种类的蛋白质D.产物的反馈抑制E.蛋白质活性的快速调节79.下列哪些情况对于真核生物的基因转录具有调控作用A.反式作用因子的磷酸化B.类固醇激素与胞内受体结合C.特定DNA序列的甲基化D.组蛋白的乙酰化E.蛋白质因子的羟基化80.管家基因的含义是A.在各组织细胞中都表达B.在特定的组织细胞中表达C.在不同发育阶段都表达D.在特定的发育阶段表达E.表达程度在不同时空条件下差异显著81.关于特异转录因子的描述,正确的是A.在所有组织细胞中组成性表达B.在不同组织细胞中存在的种类不同C.在不同组织细胞中的浓度不同D.调控管家基因的转录E.是真核生物基因表达特异性的根源所在(二)名词解释1.管家基因(housekeeping gene)2.可调节基因(regulated gene)3.顺式作用元件(cis-acting element)4. 反式作用因子(trans-acting factors)/转录因子(transcription factor, TF)5. 基础转录因子(basal/general transcription factor)6. 特异转录因子(special transcription factor)7.操纵子(operon)8. 衰减(2廿0口口2宜。
第六章 原核基因表达调控
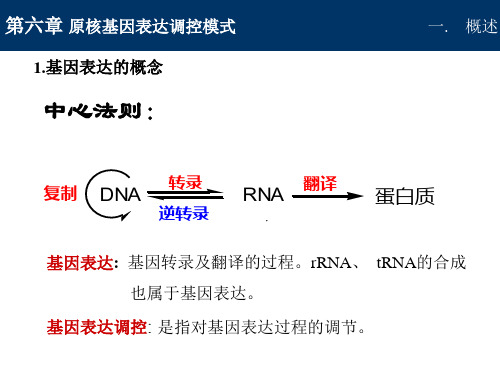
第六章 原核基因表达调控模式
-- 乳糖(+)时, 葡萄糖(+)
二.乳糖操纵子调节机制
i基因
有诱导物时
P
O
Lac Z
Lac Y 转录
lac A
阻遏蛋白 阻遏物 蛋白亚基 无活性的 阻遏蛋白
翻译 转录水平低, 没有乳糖酶的合成 β- 半乳糖苷酶 β- 半乳糖苷通透酶 β- 半乳糖苷乙酰转移酶
mRNA
诱导物
第六章 原核基因表达调控模式
一.
概述
根据细菌酶的合成对环境的反应不同,分为:
组成酶 细菌酶 诱导酶
适Байду номын сангаас酶
阻遏酶
第六章 原核基因表达调控模式
3. 原核基因表达的多级调控
四个基本调控点: 基因活化 转录水平上的调控
最有效的调节环节
一.
概述
转录起始--- DNA元件与调控蛋白相互作用调控 mRNA加工成熟水平的调控 转录后水平上的调控 翻译及翻译后水平的调控
调节基因的产物-阻遏蛋白
负控诱导 负控阻遏 正控诱导
调控机制
负转录调控 (为主) 正转录调控
正控阻遏
调节基因的产物-激活蛋白
第六章 原核基因表达调控模式
负调控 Lac O 正调控 Ara O
一.
概述
诱
导 阻遏物 阻遏 诱导物 诱导
失活的阻遏物 失活的活性蛋白 阻遏
活化的激活蛋白 诱导物 诱导
Trp O
(1) 葡萄糖(+), 乳糖(–)时,
- 乳糖(–)时, 无别乳糖存在,阻遏蛋白与操纵子上的 O序列结合, 使操纵子处于关闭状态,三个结构基因 不表达。 - 葡萄糖(+)及cAMP浓度低时,CAP 活性低, 无 cAMP- CAP复合物形成。
第六章 原核生物表达调控

第一节概述围绕基因表达过程中发生的各种各样的调节方式都通称为基因表达调控(gene regulation或gene control)。
几个基本概念1、顺式作用元件和反式作用因子:基因活性的调控主要通过反式作用因子(通常是蛋白质)与顺式作用元件(通常在DNA 上)相互作用而实现。
顺式作用元件是指对基因表达有调节活性的DNA序列,其活性只影响与其自身同处在一个DNA分子上的基因;同时,这种DNA序列通常不编码蛋白质,多位于基因旁侧或内含子中,如启动子和终止子,都是典型的顺式作用元件。
反式作用因子是能调节与它们接触的基因的表达的各种扩散分子(通常是蛋白质),如RNA聚合酶、转录因子。
2、结构基因和调节基因:结构基因(structural gene)是编码蛋白质或RNA的基因。
细菌的结构基因一般成簇排列,多个结构基因受单一启动子共同控制,使整套基因或都表达或都不表达。
调节基因(regulator gene)是编码合成那些参与其他基因表达调控的RNA或蛋白质的特异DNA 序列。
调节基因编码的调节物质通过与DNA上的特定位点结合控制转录是调控的关键。
比如:它能使结构基因在需要某种酶时就合成某种酶,不需要时,则停止合成,它对不同染色体上的结构基因有调节作用。
调节物与DNA特定位点的相互作用能以正调控的方式(启动或增强基因表达活性)调节靶基因,也能以负调控的方式(关闭或降低基因表达活性)调节靶基因。
DNA位点通常位于受调节基因的上游,但也有例外.3、操纵基因和阻遏蛋白操纵基因(operator)是操纵子中的控制基因,在操纵子上一般与启动子相邻,通常处于开放状态,使RNA聚合酶能够通过并作用于启动子启动转录。
但当它与调节基因所编码的阻遏蛋白结合时,就从开放状态逐渐转变为关闭状态,使转录过程不能发生。
阻遏蛋白(aporepressor)是负调控系统中由调节基因编码的调节蛋白,它本身或与辅阻遏物(corepressor)一起结合于操纵基因,阻遏操纵子结构基因的转录。
第六章原核基因表达调控模式思考题答案

(完整word版)第六章原核基因表达调控模式思考题答案编辑整理:尊敬的读者朋友们:这里是精品文档编辑中心,本文档内容是由我和我的同事精心编辑整理后发布的,发布之前我们对文中内容进行仔细校对,但是难免会有疏漏的地方,但是任然希望((完整word版)第六章原核基因表达调控模式思考题答案)的内容能够给您的工作和学习带来便利。
同时也真诚的希望收到您的建议和反馈,这将是我们进步的源泉,前进的动力。
本文可编辑可修改,如果觉得对您有帮助请收藏以便随时查阅,最后祝您生活愉快业绩进步,以下为(完整word版)第六章原核基因表达调控模式思考题答案的全部内容。
第七章原核生物的基因调控思考题答案一、填空题1. 能够诱导操纵子但不是代谢底物的化合物称为春基_诱导物•能够诱导乳糖操纵子的化合物IPTG 就是其中一例.这种化合物同塑遏_蛋白质结合.并使之与操纵基因分离。
乳糖操纵子的体内功能性诱导物是异乳糖.2O色氨酸是一种调节分子,被视为辅阻遏物。
它与一种蛋白质结合形成全阻遏物;乳糖操纵子和色氨酸操纵子是两个控制的例子.cAMP-cAP蛋白通过正控制起作用。
色氨酸操纵子受另一种系统弱化作用的调控.它涉及到第一个结构基因被转录前的转录竺止作用。
二、选择题(单选或多选)1o标出以下所有正确表述:(C )(a) 转录是以半保留方式获得序列相同的两条DNA链的过程(b) 依赖DNA的DNA聚合酶是多亚基酶,它负责DNA的转录(c) 细菌的转录物(mBNA)是多基因的(d) 。
因子指导真核生物hnRNA的转录后加工,最后形成mRNA(e) 促旋酶在模板链产生缺口,决定转录的起始和终止2. 下面哪些真正是乳糖操纵子的诱导物? ( (c) (d))(a)乳糖(b) 0—硝基苯酚一B -半乳糖昔(0NPG)(c) 异丙基既基半乳糖昔(d) 异乳糖3. 氨酸操纵子的调控作用是受两个相互独立的系统控制的,其中一个需要前导肽的翻译,下面哪一个调控这个系统?( (b))(a) 色氨酸(b) 色氨酰一tRNA Trp(c) 色氨酰—tRNA(d) cAMP(e)以上都不是三、判断题1. 下面哪些说法是正确的?(a) LacA的突变体是半乳糖昔透性酶的缺陷(b) 在非诱导的情况下,每个细胞大约有4分子的p—半乳糖昔酶(c) 乳糖是一种安慰诱导物(d) RNA聚合酶同操纵因子结合(e)多顺反子mRNA是协同调节的原因(f) Lac阻遏物是一种由4个相同的亚基组成的四聚体(g) 腺昔酸环化酶将cAMP降解成AMP(h) CAP和CRP蛋白是相同的(i) —35和一10序列对于RNA聚合酶识别启动子都是很重要的(j)色氨酸的合成受基因表达、阻遏、弱化作用和反馈抑制的控制(k) Trp的引导mRNA能够同时形成三个“茎一环”结构(l)在转录终止子柄部的A-T碱基对可以增强结构的稳定性(m)真核生物和原核生物的转录和翻译都是偶联的(n)在色氨酸浓度的控制下,核糖体停泊在Trp引导区一串色氨酸密码,但并不与之脱离(o) Ara c蛋白既可作为激活蛋白,又可作为阻遏蛋白起作用(p) Ara c的表达不受调控正确:b, e, f, h, i, j, n, o四、简答题1.为什么只有DNA双螺旋中的一条链能被正常的转录?答:如果两条链都被转录,每个基因就能编码两个不同的多肽。
第六章原核基因表达调控自测题

第六章原核基因表达调控1.(单选题)下列关于“基因表达”概念叙述,错误的是A. 某些基因表达产物不是蛋白质分子B. 基因表达具有组织特异性C. 基因表达具有阶段特异性D. 基因表达都要经历转录及翻译您的答案:D2.(单选题)原核细胞的RNA聚合酶有A. 多种核心酶和多种σ因子B. 多种核心酶和一种σ因子C. 一种核心酶和多种σ因子D. 一种核心酶和一种σ因子您的答案:C3.(单选题)异丙基-β-D-硫代半乳糖苷(IPTG)是乳糖操纵子的A. 诱导物B. 阻遏物C. 辅阻遏物D. 安慰诱导物您的答案:A4.(单选题)成簇有规律间隔短回文重复序列(CRISPR)可以促进细菌A. 生长B. 清除含有相同和相似序列的病毒C. 繁殖D. 抵御一切病毒您的答案:B5.(单选题)下列关于固氮基因调控体系说法正确的是:A. 固氮基因调控体系是一个级联调控体系B. nifL和nifA 基因位于不同的操纵子上C. 在有氧环境下,NifL 可以结合NifA,激活nif 基因转录D. 细胞中氨的浓度不会影响固氮基因的表达您的答案:6.(单选题)关于大肠杆菌CsrAB 调节系统,下列说法不正确的是:A. CsrA 是一个RNA 结合蛋白B. CsrB是一个编码的RNA 分子C. CsrA可以与CsrB 的RNA 分子相结合D. CsrA 蛋白可激活糖酵解过程您的答案:7.(单选题)若维生素B12合成酶的mRNA与维生素B12结合形成核开关,可以维生素B12合成酶基因的A. mRNA分解B. 转录速度下降C. 翻译速度下降D. 转录和翻译均终止您的答案:8.(多选题)辅诱导物A. 可以和诱导物结合使其失活B. 可以和阻遏蛋白结合使其活化C. 可以同诱导物结合使其活化D. 可以和阻遏蛋白结合使其失活您的答案:9.(多选题)当培养基中含有大量葡萄糖时,正确的说法是:A. cAMP水平增高B. cAMP水平降低C. 乳糖操纵子的表达水平增高D. 乳糖操纵子的表达水平降低您的答案:10.(多选题)下列糖代谢中有关酶的调节,哪些属于降解物敏感型操纵子?A. 山梨醇B. 阿拉伯糖C. 乳糖D. 麦芽糖您的答案:11.(多选题)gal 操纵子有哪些特点:A. 有两个启动子B. mRNA 可从两个不同的起始点开始转录C. 它有两个操纵区D. 操纵区在结构基因galE 内部您的答案:12.(多选题)下列哪些操纵子属于多启动子调控A. 核糖体蛋白SI 操纵子B. rRNA 操纵子C. trp操纵子D. DnaQ 蛋白操纵子您的答案:13.(多选题)关于RNA调节基因表达,下列说法正确的是:A. 细菌响应环境压力的改变,会产生长度在20~50 nt 之间的小RNAB. 原核生物sRNA 以反式编码sRNA 为主C. sRNA抑制或促进靶mRNA的翻译D. sRNA加速或减缓靶mRNA的降解您的答案:14.(多选题)核糖体A位点上未装载氨基酸的空载tRNAA. 促进ppGpp的合成B. 使细菌死亡C. 引发细菌的严谨反应D. 使细菌生长缓慢您的答案:15.(判断题)大肠杆菌mRNA的poly(A)缩短会加速mRNA的降解。
第6章原核生物的基因表达调控

一、操纵子(operon)
细菌能随环境的变化,迅速改变某 些基因表达的状态,这就是很好的基因 表达调控的实验模型。人们就是从研究 这种现象开始,打开认识基因表达调控 分子机理的窗口的。
既然从DNA到蛋白质的过程称为基因表达,对这个过程
的调节就称为基因表达调控(gene regulation或gene control)。基因表达调控是现阶段分子生物学研究的中
心课题。
6.1.1基因表达调控的意义
基因组(genome) 是指含有一个生物体生存、发育、活动和 繁殖所需要的全部遗传信息的整套核酸。
一个受精卵含有发育成一个成熟个体的全部遗传信息,在个 体发育分化的各个阶段,各种基因极为有序地表达,一般 在胚胎时期基因开放的数量最多,随着分化发展,细胞中 某些基因关闭(turn off)、某些基因转向开放(turn on), 胚胎发育不同阶段、不同部位的细胞中开放的基因及其开 放的程度不一样,合成蛋白质的种类和数量都不相同,显 示出基因表达调控在空间和时间上极高的有序性,从而逐 步生成形态与功能各不相同、极为协调、巧妙有序的组织 脏器。
组成性基因表达也不是一成不变的,其表达强弱也是受一定机制调控的。
②适应性表达(adaptive expression)指环境的变化容易使其表达 水平变动的一类基因表达。
应环境条件变化基因表达水平增高的现象称为诱导(induction), 这类基因被称为可诱导的基因(inducible gene);
第六章、核酸与蛋白质序列分析2

• 核酸序列中的功能位点包括转录因子结合位点、 转录剪切位点、翻译起始位点等。
• 在蛋白质序列分析中,常使用序列模式这个名
词,蛋白质的序列模式往往与蛋白质结构域或者
作用部位有关。
09.04.2021
根据蛋白质编码基因的一般性质和特征进行识别, 通过统计值区分外显子、内含子及基因间区域 。
• 基于同源序列比较的方法
利用数据库中现有与基因有关的信息(如EST序列、 蛋白质序列),通过同源比较,帮助发现新基因。
09.04.2021
20
第六章、核酸和蛋白质序列分析
四、DNA序列分析方法
一个基本的DNA序列分析方案
9
第六章、核酸和蛋白质序列分析
3、基因识别
• 基因识别是生物信息学领域里的一个重要研 究内容
• 基因识别问题,在近几年受到广泛的重视
—当人类基因组研究进入一个系统测序阶段时, 急需可靠自动的基因组序列翻译解释技术,以处 理大量已测定的但未知功能或未经注释的DNA序 列
09.04.2021
10
第六章、核酸和蛋白质序列分析
5’UTR---基因上游区域的非翻译区域 3’UTR---基因下游区域的非翻译区域
09.04.2021
12
第六章、核酸和蛋白质序列分析
对于任何给定的核酸序列(单链DNA或 mRNA),根据密码子的起始位置,可以按照三 种方式进行解释。
例如,序列ATTCGATCGCAA
(1) ATTCGATCGCAA (2) ATTCGATCGCAA (3) ATTCGATCGCAA
(1)原核基因识别
第七章:原核生物基因表达调控

负转录调控
• 负控诱导 阻遏蛋白不与效应物
结合时基因不转录。
• 负控阻遏 阻遏蛋白与效应物 结合时,基因不转录。
正转录调控
• 正控诱导 有效应物时,激活蛋白 处于活性状态基因转录。
• 正控阻遏 有效应物时,激活蛋 白处于无活性状态基 因不转录。
负控诱导
正控诱导
负控阻遏
正控阻遏
1、原核生物基因调控机制的类型
6、色氨酸操纵子(元)调节过程涉及 D
(A) 转录水平调节 (B) 转录激活调节
(C) 翻译水平调节
(D) 转录/翻译调节
(A) Lac阻遏蛋白 (B) RNA聚合酶
(C) 环一磷酸腺苷
(D) CRP-cAMP 7、与O序列结合
A
8、与P序列结合 B 9、 与CRP结合 C 10、与CAP位点结合 D
境能提供足够浓度的色氨酸时,R与色氨酸结合后而活化,
能够与O结合,阻遏结构基因的转录,开 关。
(二) 弱化子及其作用
当色氨酸达到一定浓度,但还没有高到能够活化 R使其起阻遏作用的程度时,产生色氨酸合成酶 类的量已经明显降低,而且产生的酶量与色氨酸 浓度呈负相关。这种调控现象与色氨酸操纵子特
殊的结构有关。
乳糖存在诱导基因转录
乳糖(lac)操纵子的表达调控
1、阻遏蛋白的负性调控
没有乳糖时,1ac操纵子处于阻遏状态。i 基因在自身的 启动子Pi 控制下,产生阻遏蛋白R。R以四聚体形式与 操纵子o结合,阻碍RNA聚合酶与启动子P的结合。
当有乳糖存在时,乳糖与R结合,使R四聚体解聚成
单体,失去与o的亲和力,与o解离,基因转录开放。
(D) 在生物个体的某一生长阶段持续表达
2、一个操纵子(元)通常含有
第六章 原核生物基因表达调控

图6-7 乳糖操纵子结构模式图
第二节 原核基因表达的调控
乳糖操纵子的上游有一个独立转录的基因lacI,其编码产物LacI 可以结合在乳糖操纵子的操纵基因(lacO)上,即转录控制区,阻抑 下游结构基因的表达。因此,乳糖操纵子是一个负调控系统(图68)。其中,LacI是具有负调控作用的反式作用因子,LacI作用的靶 DNA序列lacO是顺式作用元件。
trpB(UGA处翻译终止) -UGA -GAA-AUC- UGA-UGG-AA A UG-G AAtrpA(AUG处翻译起始)
第二节 原核基因表达的调控
3.稀有密码子对翻译的影响 DNA复制时,引物酶催化一段RNA引物的合成,引物酶 由dnaG编码。rpsU-dnaG-rpoD组成一个转录单位,产生多 顺反子转录物。细胞内三个基因的终产物的浓度相差却很 大,rpsU产物浓度为4×104个/细胞,dnaG产物50个/细胞, rpoD产物2800个/细胞。菌体通过使用稀有密码子,使转 录为一条mRNA链的三个基因的表达产物量可以有很大差异。
第二节 原核基因表达的调控
ቤተ መጻሕፍቲ ባይዱ
图6-10 色氨酸操纵子的负调控
第二节 原核基因表达的调控
4.阿拉伯糖操纵子 阿拉伯糖与乳糖一样,可替代葡萄糖作为碳源物质被 菌体利用。大肠杆菌中,阿拉伯糖(Ara)代谢所需酶的 三个基因分别是:核酮糖激酶基因( araB)、L-Ara异构 酶 基 因 ( araA)、L- 核 酮 糖 - 5 - 磷 酸 差 向 异 构 酶 基 因 ( araD),组成一个基因簇,有共同的启动子 PBAD。与其 它操纵子不同的是,操纵序列位于 PBAD 上游,操纵序列左 端有另一方向转录的启动子 PC,负责调节基因araC的转录, 其产物AraC蛋白有两种活性形式,Pr 对 PBAD 的表达起阻遏 作用,Pi对PBAD的表达起激活作用(图6-11)。
遗传学第六章基因与发育

2
1
3种胚层经过了细胞分化生成各种器官的原基,如肢、眼、心等原基,这是器官生成。
原基先是生成微小而精确的结构,然后逐渐长大,在生物体的各个正确部位长成各种器官和组织,这是形态建成。
神经管胚形成期
器官生成 形态建成 人的发育过程
全能干细胞(totipotent):能够分化产生各种细胞直至个体的细胞。例如胚胎干细胞(embryonic stem cell)。
如果背部蛋白质没有进入细胞核,则细胞核的腹化基因不能激活,背化基因不被抑制——分化出背化细胞。
当合胞体细胞形成许多细胞时,背部蛋白质进入合胞体腹侧的细胞核,细胞核的腹化基因被激活,背化基因被抑制——分化出腹化细胞;
母体背部基因的RNA在卵受精后90min时才翻译产生背部蛋白质。
前端组织中心
后端组织中心
在8个卵裂球期,每个卵裂球在生化、形态和发育潜能上都没有差别,也就是在发育上是全能的。 可是当卵裂球成团结合时,细胞处在外层还是内层,会使以后生成的卵裂球出现了不同的生物学功能。处在外层的细胞生成滋养层,而处在内层的细胞则生成内细胞团而产生胚胎(几个细胞?)。
如果胚泡中只有一个细胞形成以后的胚胎,则生下小鼠的毛色或者全是黑色,或者全是白色。 如果胚泡中有两个细胞参与胚胎的生成,则异决表型小鼠的数目应占子代中的半数(1ww:2wB:1BB), 如果胚泡中的3个细胞生成胚胎,则异决表型小鼠应占75%(1www:3wwB:3wBB:1BBB); 如果胚泡中的4个细胞生成胚胎,则毛色黑白相间小鼠的概率为87.5%。
(2)成对规则基因(pairrule genes)
成对规则基因
2、分节基因
体节极性基因的转录图式是受成对规则基因pair-rule基因所调控,这类基因的功能是保持每一体节中的某些重复结构,体节极性基因在每个体节内部调控其发育模式,包括极性。 当这类基因发生突变后,会使每一体节的一部分结构缺失,而被该体节的另一部分的镜像结构所替代。例如:engrail基因是保持前后体节间的分界,engrail突变型胚胎则出现前后体节融合为一,即每一体节的后半部被后一个体节的前半部的重复结构所替代。
第六章 真核生物基因表达

(一)真核生物基因组DNA甲基化
1.真核生物DNA甲基化位点 真核生物DNA的mCpG是DNA甲基化的主要形式
CpG岛: 由于CpG通常成串在DNA双链对称出现,被称为~, mCpG占全部CpG的70%
76,000 ?
52,000 ?
44,000 ?
?
?
?
?
普遍 淋巴细胞 淋巴细胞 普遍 普遍
▪ 了解启动子及各个元件的特点、信息有 什么作用?
▪ 启动子有没有改造的空间呢?
▪ 双向启动子
▪ 组织特异性启动子
▪ 诱导性启动子
(二)增强子
增强子(enhancer):又称为远上游序列,位于转录起始位点
人类基因组中免疫球蛋白基因主要片段的数量比较
V、C和J基因片段在胚胎细胞中相隔较远。编码产生免疫球蛋白的细胞,发育 分化时,通过染色体内DNA重排把4个相隔较远的基因片段连接在一起,产生 具有表达活性的免疫球蛋白基因。
▪ 酿酒酵母接合型:
▪酵母细胞通 过交换型转 换过程改变 自己的性别。 MATa或 MATα基因 座位两侧分 别存在两个 MAT样基因 HMLα和 HMRa沉默 交配型盒。
(3)DNA去甲基化位点的特点
① DNA去甲基化位点范围和DNA酶I优先敏感区域十分 吻合
②只有一小部分CG二核苷酸对的去甲基化与基因激活有关, 它们位于对基因表达关系十分密切的序列中
▪ (4) DNA甲基化/去甲基化对基因活性调控的相对性 ① DNA甲基化程度因物种而异
DNA甲基化随进化程度的提高而增强
的上游,它们不是启动子的一部分,但能增强或促进转录的
分子生物学第六章真核生物基因表达调控

教案首页课程名称分子生物学任课教师李市场第6 章真核生物的表达调控计划学时 6教学目的和要求:a)真核生物的基因结构与转录活性b)真核基因的转录c)反式作用因子*d)真核基因转录调控的主要模式e)其他水平上的基因调控重点:真核生物翻译调控的影响因素及其转录调控的主要模式。
难点:真核生物翻译调控的影响因素及其转录调控的主要模式。
思考题:1真核基因转录调控的主要模式有哪些?第六章真核基因表达调控原核生物的调控系统就是要在一个特定的环境中为细胞创造高速生长的条件,或使细胞在受到损伤时,尽快得到修复,所以,原核生物基因表达的开关经常是通过控制转录的起始来调节的。
真核生物(除酵母、藻类和原生动物等单细胞类之外)主要由多细胞组成,每个真核细胞所携带的基因数量及总基因组中蕴藏的遗传信息量都大大高于原核生物。
人类细胞单倍体基因组就包含有3×109bp总DNA,约为大肠杆菌总DNA 的1000倍,是噬菌体总DNA的10万倍左右!真核基因表达调控的最显著特征是能在特定时间和特定的细胞中激活特定的基因,从而实现"预定"的、有序的、不可逆转的分化、发育过程,并使生物的组织和器官在一定的环境条件范围内保持正常功能。
真核生物基因调控,根据其性质可分为两大类:第一类是瞬时调控或称可逆性调控,它相当于原核细胞对环境条件变化所做出的反应,包括某种底物或激素水平升降及细胞周期不同阶段中酶活性和浓度的调节。
第二类是发育调控或称不可逆调控,是真核基因调控的精髓部分,它决定了真核细胞生长、分化、发育的全部进程。
转录水平调控(transcriptional regulation);转录后水平调控(post transcriptional regulation);翻译水平调控(translational regulation);蛋白质加工水平的调控(regulation of protein maturation)等。
《分子生物学》教案

《分子生物学》教案第一章:分子生物学概述1.1 分子生物学的定义和发展历程1.2 分子生物学的研究内容和方法1.3 分子生物学的重要性和应用领域第二章:DNA与基因2.1 DNA的结构和功能2.2 基因的概念和作用2.3 基因的表达和调控第三章:RNA与蛋白质3.1 RNA的结构和功能3.2 蛋白质的结构和功能3.3 蛋白质合成和调控第四章:酶与催化作用4.1 酶的定义和特性4.2 酶的分类和作用机制4.3 酶的研究方法和应用第五章:分子生物学实验技术5.1 分子克隆与基因工程5.2 PCR技术及其应用5.3 蛋白质分离和鉴定技术5.4 生物信息学在分子生物学中的应用第六章:基因表达调控6.1 基因表达的转录和翻译过程6.2 真核生物的转录调控机制6.3 翻译调控和后修饰机制第七章:蛋白质结构与功能7.1 蛋白质结构的基本层次7.2 蛋白质功能的多样性7.3 结构决定功能的原则第八章:信号传导与细胞代谢8.1 细胞信号传导的基本概念8.2 细胞信号传导的主要途径8.3 信号传导与细胞代谢的调控第九章:基因组学与遗传变异9.1 基因组学的基本概念和方法9.2 基因组结构和变异类型9.3 遗传变异在疾病和进化中的作用第十章:分子生物学在生物技术与医学中的应用10.1 基因克隆与基因治疗10.2 重组蛋白药物的开发与应用10.3 分子诊断与个性化医疗10.4 生物芯片技术及其应用第十一章:分子生物学实验设计与分析11.1 实验设计的原则和方法11.2 实验数据的收集与分析11.3 实验结果的验证与解释第十二章:蛋白质相互作用与网络12.1 蛋白质相互作用的机制12.2 蛋白质相互作用网络的构建与分析12.3 蛋白质相互作用在生物学中的意义第十三章:RNA干扰与基因沉默13.1 RNA干扰机制及其作用13.2 基因沉默技术在研究中的应用13.3 RNA干扰在医学和生物技术领域的应用第十四章:病毒分子生物学14.1 病毒的基本结构与生命周期14.2 病毒基因组的复制与表达14.3 病毒与宿主细胞的相互作用第十五章:分子生物学在生物技术与医学中的应用案例分析15.1 基因治疗与基因编辑技术的应用15.2 生物制药与重组蛋白的应用15.3 分子诊断与个性化医疗的实践案例重点和难点解析第一章:分子生物学概述重点:分子生物学的定义和发展历程,研究内容和方法,重要性和应难点:分子生物学研究方法的理解和应用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
6.2 操纵子
• 又称操纵元,是一组核苷酸序列,包括了一个操纵序 列,一个普通的启动子,及一个或以上的结构基因被 用作生产信使RNA(mRNA)的基元。操纵子主要在 原核生物及线虫动物门出现 。 • 把生物学活性相关的基因放在一起,便于管理,一开 俱开,一关俱关。 I P O X Y Z
Operon control model
High trp
Trp is inserted at the trp codons
Translate to the end of leader message Ribosome occlude sequence 2
Terminate transcription at (3:4 form terminator)
• 大肠杆菌半乳糖操纵子(galactose operon)包括3个结构 基因,一个调节基因:
• • • • 异构酶(UDP-galactose-4epimerase,GalE); 半乳糖-磷酸尿嘧啶核苷转移酶(galactose transferase, GalT); 半乳糖激酶(galactose kinase, Galk)。 阻遏蛋白(GalR)
gal操纵子的特点
• 它有两个相距仅5 bp启动子,其mRNA可从两个不同 的起始点开始转录; • P1起始(强启动):需要半乳糖+CAP+cAMP • P2起始(弱启动):低浓度cAMP-CAP(即存在葡萄 糖)。
总结起来: 1)无glu有gal,P1高效起始; 2)有glu无gal,P2低效起始。 问题: 1)有glu有gal,如何起始? 2)无gal时,阻遏蛋白仍然结合O位点,RNApol 如何从P2起始呢?
• 半乳糖的代谢步骤:
• • • • Gal+ATP Gal-P+ADP (Galk) Gal-P+UDPGlu UDPGal+Glu-p (GalT) UDPGal UDPGlu (GalE) 总反应式: Gal+ATP =ADP+Glu-p
GalR阻遏机理
无半乳糖: GalR接合两个调控区域,环化, RNApol无法正确启动; 有半乳糖:半乳糖结合GalR使其失去阻遏作用。
CAP结合位点
•CAP为二聚体, 45KD ,被cAMP 激活; •结合位点~22bp I -70 ~ -50 II -50 ~ -40
CAP的结合对DNA构型的影响
• DNA弯曲 • 弯曲点位于CAP结合位点二重对称的中心 • 弯曲使CAP能与启动子上的RNA pol 接触
6.4 半乳糖操纵子
衰减子普遍的生物学意义
6.6 噬菌体基因表达的调控
• λ噬菌体进入细菌细胞以后,可以按裂解或建立溶原两 种生活方式生存和繁殖; • 在每个菌体内繁殖100个左右的新噬菌体,并裂解细菌 释放出来,再度感染其它活菌,称为裂解; • λ噬菌体把自身DNA整合进细菌染色体DNA中,作为 细菌基因的一部分,随菌的细胞分裂而增殖,这种生 活状态称为溶原;
2. 基因表达的调控涉及 3. 原核生物与真核生物基因表达调控机制具有 惊人的相似性,共同的起源与共同的分子基 础
4.
• • •
调控机理
核酸分子间的互作 核酸与蛋白质分子间的互作 蛋白质分子间的互作
5.
• • • •
调控层次
transcriptional level post—transcriptional level translational level post—translational level
噬菌体是如何进行选择,是进入裂解 还是溶原途径呢?
OR3 OR2 OR1
cro
cⅡ
PE tR1
O P Q
tR2 tR3 PR’
cⅢ
tL1
N
PLOL
cⅠ
PM
PROR
OL1 OL2 OL3
N CⅢ
Cro
早期 CⅡ
O P Q
晚早期
CⅠ CⅠ
PE PM
晚期
λ噬菌体基因调控过程
• 早期 • 由PL转录翻译出抗终止蛋白N,由PR转录出调节蛋 白Cro; • 晚早期 • 在N蛋白的作用下,使得RNApol 越过tL1、tR1、 tR2,从而使蛋白CⅢ、CⅡ、复制相关蛋白O、P, 抗终止Q等表达; • CⅢ与CⅡ共同作用,启动cro与cⅡ基因之间的启动 子PE,使得CⅠ的得以表达; • 转录出来的RNA含有cro基因的反义mRNA,后者抑 制了cro基因的翻译; • 由于PR和PE的RNApol会碰头, 负责PR的RNApol的 活性会被PE的RNApol排挤;
• 细胞外研究诱导作用时 ,很少使用乳糖 • 替代物:
• IPTG,异丙基半乳糖苷 • TMG ,巯甲基半乳糖苷 • ONPG, O-硝基半乳糖苷
6.3.3 阻遏蛋白的作用机制
• 38KD,4 聚体; • lacI,组成型转录 ,Pi 弱启动子,5 -10个/cell; • 具有二重性:
• 阻止转录(与lacO 结合) • 开始转录(与诱导 物结合)
Chapter 6
原核生物基因表达的调控
本章内容
• • • • • • • • 概述 操纵子 乳糖操纵子 半乳糖操纵子 色氨酸操纵子 噬菌体基因表达的调控 信号转导系统 真核基因的表达调控
6.1 概述
1. 基因调控存在的生物学意义
• • • • 原核生物对环境的适应,相关的应答; 真核生物的细胞分化, 组织特化 , 个体发育. RNA转录的开/关,数量,选择性加工 蛋白质翻译.1 转录水平的调控
通过阻遏物结合操纵子,转录效率降低 至1/70;然而细胞中,在阻遏状态下, trp酶系统的酶活却只有1/700,那么,是 不是还存在另外一个阻遏系统呢?
6.5.2 翻译水平的调控
前导肽的前后序列结构
4个片段进行配对,形成不同的二级结构
Lack of trp Lack of aminoacyl tRNA Ribosome pause at trp codons, occluding sequence 1 Form 2:3 hairpin antiterminator Transcription into trpE and beyond
• λ噬菌体进入溶原周期 • CⅠ蛋白与操纵子的亲和力顺序: OR3<OR2<OR1, OL3<OL2<OL1;Cro蛋白与操纵子的亲和力顺序: OR3>OR2>OR1, OL3>OL2>OL1; • 当CⅠ蛋白表达以后,迅速占据OR1与OL1位点,由 于这两个操纵子被占据后,RNApol无法起使转录, PL与PR转录被迫终止; • PR不能转录, CⅡ就不能合成,那么CⅠ的合成也 将被关闭;这时,结合上OL1的CⅠ会促进PM的转录 ,当CⅠ过量时, CⅠ便会结合到OL3,而与后者的 结合则会影响PM的转录,因此CⅠ始终维持平衡; • CⅡ除了促进CⅠ蛋白的表达,还促进PI启动子的转 录,转录翻译出来的整合酶,促进λ噬菌体整和进宿 主的染色体DNA,完成溶原过程。
6.3.4 CAP-cAMP复合物在乳糖操纵子表达 中的作用----乳糖操纵子的正调控
葡萄糖耗尽
ATP
腺苷环化酶 促进RNApol 与启动子结合 ,起始转录
cAMP
cAMP+CAP
与CAP接受位点结合
CAP-cAMP正调空转录的机制
• 大多数原核启动子的-10区序列为TATAAT; • 而乳糖操纵子的-10区碱基序列为TATGTT,由于存在 G:C碱基对,使得RNA聚合酶无法解开双链进行转录 起始; • CAP-cAMP复合物的结合,使得CAP附近的G:C稳定 性降低,使得RNA聚合酶能顺利解开DNA双链,起始 转录; • 如果把-10区的碱基序列从TATGTT突变成TATAAT, 细胞将能同时利用葡萄糖和乳糖。
阻遏物
OFF
ON
6.3.2 诱导底物
别乳糖(诱导物)
β-半乳糖苷酶(LacZ)来源?
本底组成型表达
• 阻遏蛋白对lacO的结合并不是经久不变的,它有10 -20分钟的半衰期; • lacO处于游离状态的时间很短,新的阻遏蛋白会立即 结合上去; • 但有时RNA聚合酶会抓住机会,进行一次转录,保 证每个细胞周期有一个lac mRNA分子,产生几个β-半 乳糖苷酶来巡逻执勤。
6.3 乳糖操纵子 (负调控模式)
• 1940年, Monod发现:细菌在含葡萄糖和乳糖的培养 基上生长时,细菌先利用葡萄糖,葡萄糖用完后,才 利用乳糖;在糖源转变期,细菌的生长会出现停顿。 即产生“二次生长曲线”; • 1951年,Monod与Jacob合作,发现两个基因: • Z基因:与合成β-半乳糖苷酶有关; • I基因:决定细胞对诱导物的反应。 • Jacob:结构基因旁有开关基因(操纵基因),阻遏物 通过与开关基因的结合,控制结构基因的表达; • 1961年, F. Jacob & J.Monod提出乳糖操纵子概念; • 1965年诺贝尔生理学和医学奖。
• λ噬菌体进入裂解周期 • 宿主中存在hfl基因,它的表达产物是蛋白CⅡ的水 解酶,水解了CⅡ, CⅠ将得不到表达…… • 而hfl基因的表达受到cAMP-CAP的负调控,存在 cAMP-CAP时, hfl基因关闭; 缺乏cAMP-CAP时 , hfl基因打开; • 没有CⅠ蛋白, Cro蛋白将得到大量积累,而Cro结 合于OR3与OL3位点,不影响PL与PR转录的继续进行 ; • 当细胞中有足够的复制蛋白O、P,抗终止蛋白Q以 后,噬菌体结构基因A ……J表达,溶菌酶基因得已 表达,A蛋白切割cos位点,噬菌体包装后释放出来 。
两个转录起始位点的生物学意义
• 半乳糖不仅作为碳源供细胞生长,另一方面尿苷二磷酸 半乳糖(UDPgal)是大肠杆菌细胞壁合成的前体; • 在半乳糖存在的情况下,由半乳糖操纵子合成UDP-gal ;在没有半乳糖的情况下,UDP-gal是通过半乳糖异构 酶的作用由UDP-Glu合成的,该酶是GalE基因的产物 ;因此,细菌必须以低水平不断合成GalE; • 若只有P1,当Glu存在时,无法合成GlaE,若只有P2 ,当Glu不存在而Gla存在时,无法高水平合成Gal酶系 统。