植物内生细菌的生物薄膜(biofilm)
专业解读-细菌生物被膜Bacterial biofilm形成、特点预防与控制

细菌生物被膜Bacterial biofilm目录细菌生物被膜Bacterial biofilm (1)定义 (2)形成过程原理 (3)表面特性 (5)特点及耐药性 (6)预防与控制 (7)细菌生物被膜(或称细菌生物膜Bacterial biofilm,BF),是指细菌粘附于接触表面,分泌多糖基质、纤维蛋白、脂质蛋白等,将其自身包绕其中而形成的大量细菌聚集膜样物。
Biofilm formation constitutes an alternative lifestyle in which microorganisms adopt a multicellular behavior that facilitates and/or prolongs survival in diverse environmental niches. Biofilms form on biotic and abiotic surfaces both in the environment and in the healthcare setting. In hospital wards, the formation ofbiofilms on vents and medical equipment enables pathogens to persist as reservoirs that can readily spread to patients. Inside the host, biofilms allow pathogens to subvert innate immune defenses and are thus associated with long-term persistence. Here we provide a general review of the steps leading to biofilm formation on surfaces and within eukaryotic cells, high lighting several medically important pathogens, and discuss recent advances on novel strategies aimed at biofilm prevention and/or dissolution.多糖基质通常是指多糖蛋白复合物,也包括由周边沉淀的有机物和无机物等。
biofilm
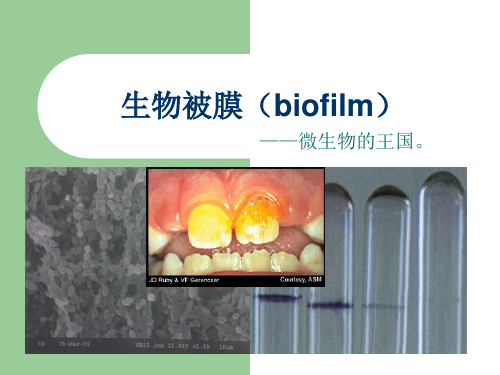
生物被膜(BF)的定义
微生物有组织生长的聚集体。
细菌不可逆的附着于惰性或活性实体的 表面,繁殖、分化,并分泌一些多糖基 质、将菌 体群落包裹其中而形成的细 菌聚集体膜状物。
单个生物被膜可由同种或不同种微生物 形成。
生物被膜(BF)的组成及结构
包括分泌的多糖蛋白、多糖基质、纤维蛋白、脂蛋白 等多糖蛋白复合物。 粘附细胞之间散在着一些“水通道”,即细菌群落之间 存在的充满环境液体的内隙。 成熟生物被模型从外到内包括主体生物膜层、连接层、 条件层、基质层。
三、氧浓度变化对铜绿假单胞菌生物被膜 形成的影响
研究无氧、低氧(10%氧浓度)、正常氧(20%)及高氧(30%、 40%、50%、60%)七个不同氧浓度条件下生物被膜和藻 酸盐的产生情况。 实时定量检测不同氧浓度对Lasl、RhlI、algU表达的影响 建立生物被膜生成量和Lasl、RhlI、algU表达量之间的关 系图。
研究生物被膜(BF)的意义
近年来随着医学界对某些环境中常见细菌所致的一些 慢性和顽固性疾病的深入了解 ,发现生物膜是导致这 些细菌性疾病难以根治的主要原因 。 以生物膜形式存在的细菌不同于浮游细菌 ,它们对抗 生素等杀菌剂、恶劣环境及宿主免疫防御机制有很强 的抗性,生物膜内细菌在生理、代谢、对底物的降解或 利用和对环境的抵抗能力等方面都具有独特的性质
生物被膜(BF)的体外鉴定
一、96孔微量板定量检测法
是目前各实验室广泛应用的定量检测细菌生物被膜的 公认方法。目前几乎所有的细菌都以该方法被报道。 不仅能定性细菌形成生物被膜,而且和不同的染色方 法结合还能定量计算细菌形成生物被膜的能力。
生物被膜形成曲线的测定
生防菌对植物真菌病害作用整理

生防菌对植物真菌病害的作用学院:生命科学学院专业班级:学生姓名:目录摘要 (3)1植物真菌病害 (3)2生防菌的种类及生防机制 (3)2.1 生防菌的种类 (3)2.2 生防菌的生防机制 (4)2.2.1 竞争作用 (4)2.2.2 拮抗作用 (5)2.2.3 诱导抗性作用 (5)2.2.4 促生作用 (6)3 生防菌的筛选与鉴定 (7)3.1 拮抗芽孢杆菌的分离 (7)3.2 芽孢杆菌的分类鉴定 (7)参考文献: (8)生防菌对植物真菌病害的作用摘要:真菌病害是造成作物产量损失的主要原因,作物病害的80%由病原真菌引起,利用微生物及其代谢产物对其进行生物防治,是目前研究的热点。
可用于生物防治的微生物有真菌、细菌、放线菌、病原菌弱致病菌等。
生防菌的生防机制各不相同,主要有竞争作用、拮抗作用、诱导作物抗性和促进作物生长,间接提高作物抗性等作用,许多生防微生物还可通过几种不同机制之间的联合来发挥功能。
本文还对生防菌的分离与分类鉴定进行了简单介绍。
关键词:真菌病害,生物防治,生防机制,木霉菌,芽孢杆菌,放线菌1植物真菌病害植物病害一直是农作物优质高产的重要制约因素之一。
据估计, 全球主要农作物的平均损失约占总产量的10 %~15 %, 每年直接经济损失高达数千亿美元。
在植物病害中,70 %~80 %的病害是病原真菌侵染所引致的。
植物真菌病害不仅直接造成农作物产量下降与品质降低, 而且部分病原真菌在侵染农作物过程中, 可分泌产生多种对人畜有害的毒素与代谢物, 对农产品的安全性构成极大威胁。
此外, 重大农作物真菌病害的控制往往依赖化学防治, 杀菌剂的使用不仅增大生产成本, 而且其反复施用不可避免地带来环境污染与农产品农药残留问题[1]。
因此,近年来世界各国都在努力开发可替代传统化学药剂控制植物病害的新方法。
其中利用微生物及其代谢产物进行生物防治,被公认为是一种环境友好型的选择。
2生防菌的种类及生防机制2.1 生防菌的种类生防菌的种类繁多,生产上广泛应用的有真菌、细菌、放线菌、病毒等。
认识生物被膜

背景介绍
与人类感染有关的生物膜形成菌 Nhomakorabea相关疾病
生物膜形成菌
龋齿 牙周炎 中耳炎 骨骼肌感染 骨髓炎 心内膜炎 隐形眼镜所致感染 缝合部位感染 人工心瓣膜
产酸性G+球菌 口腔G-厌氧菌 嗜血流感杆菌 G+球菌 多种细菌和真菌(混合) 草绿色链球菌 绿脓杆菌和G+球菌 表皮葡萄球菌、金黄色葡萄球菌 金黄色葡萄球菌和表皮葡萄球菌
—J. William Costerton
背景介绍
❖ 细菌生物被膜(biofilm,BF)的存在是细菌为适应环境,有利 于生存而特有的生命现象。细菌吸附于惰性物体如生物医学材 料或机体黏膜表面后, 分泌多糖基质、纤维蛋白、脂蛋白等多 糖蛋白复合物,使细菌相互粘连并将自身克隆聚集缠绕其中形 成膜状物。生物被膜内的细菌对大多数抗生素耐药,能长期存 活,不断释放,成为感染源。
[6] Schinabeck MK, Long LA, Hossain MA, Chandra J,Mukherjee PK, Mohamed S, Ghannoum MA. Rabbit model of Candida albicans biofilm infection: liposomal amphotericin Bantifungal lock therapy[J]. Antimicrobial Agents and Chemotherapy, 2004, 48(5): 1727–1732. [7] Nett J, Lincoln L, Marchillo K, et al. Putative Role of β-1,3 Glucans in Candida albicans Biofilm Resistance[J]. Antimicrob Agents Chemother, 2007, 51(2):510-520.
生物被膜1

金黄色葡萄球菌生物被膜
Lasa I. Int microbiol 2006
表皮葡萄球菌生物被膜
Lasa I. Int microbiol 2006
铜绿假单胞菌生物被膜
方向群, 刘又宁
中华医院感染杂志
2002
生物被膜相关感染
BBF中细菌对抗菌药物高度耐药并可逃避宿主 的免疫作用,导致感染迁延不愈,称为生物被 膜相关感染(Biofilm-related infection), 主要包括生物医学材料相关感染和某些慢性感 染性疾病
Drenkard E, Ausubel FM E, Ausubel FM.. Nature, 2002
表型变异
Drenkard E, Ausubel FM. Nature, 2002
BBF及中性粒细胞的相互作用
补体↓
中性粒细胞
活性氧↓
趋化作用↓
BBF及中性粒细胞的相互作用
6.4 6.2 细菌数(lgCFU/cm2) 6 5.8 5.6 5.4 5.2 5 0MIC 1/4MIC 1/2MIC 1MIC IMI浓度(MIC) OMIC 1/16MIC 1/4MIC
方向群,刘又宁 中华医院感染杂志杂志
2007
亚胺培南与阿齐霉素合用对BF细菌的作用
药物处理前的BF SEM ×4500
细菌生物被膜相关感染
解放军总医院呼吸科 方向群
前言
1 2 3
生物医学材料应用 某些慢性感染
美国CDC统计 60%感染与生物被膜相关
前言
1 2 3
细菌生物被膜的形成 细菌生物被膜的致病机制 细菌生物被膜的药物治疗
细菌生物被膜的定义
细菌生物被膜(Bacterial Biofilm, BBF)是细菌产生多聚复合物基质将自身 包绕,粘附于无活性物体或活体表面,形 成的有一定结构的细菌群体
【word】植物与内生真菌互作的生理与分子机制研究进展

植物与内生真菌互作的生理与分子机制研究进展第28卷第9期2008年9月生态ACTAEC0L0GICASINICAV o1.28.No.9Sep.,2008植物与内生真菌互作的生理与分子机制研究进展袁志林,章初龙‟,林福呈,(1.浙江大学生物技术研究所,杭州310029;2.中国林业科学研究院亚热带林业研究所,富阳311400)摘要:在自然生态系统中,植物组织可作为许多微生物定居的生态位.内生真菌普遍存在于植物组织内,与宿主建立复杂的相互作用(互惠,拈抗和中性之间的相互转化),并且存在不同的传播方式(垂直和水平传播).内生真菌通过多样化途径来增强植物体的营养生理和抗性机能.但这种生理功能的实现有赖于双方精细的调控机制,表明宿主和真菌双方都进化形成特有的分子调控机制来维持这种互惠共生关系.环境因子(如气候,土壤性质等),宿主种类和生理状态,真菌基因型的变化都将改变互作结果.此外,菌根真菌和真菌病毒等也可能普遍参与植物一内生真菌共生体,形成三重互作体系,最终影响宿主的表型.研究试图从形态,生理和分子水平阐述内生真菌与植物互作的基础.关键词:内生真菌;植物;互作机制文章编号:10000933(2008)09.4430.10中图分类号:Q143,Q948文献标识码:A Recentadvancesonphysiologicalandmolecularbasisoffungaiendophyte- plantinteractionsYUANZhi—Lin‟一.ZHANGChu.Long一,UNFu—Cheng‟1InstituteofBiotechnology,ZhejiangUniversity,Hangzhou,ZhejiangProvi nce,310008,China2Instituteofsubtropicalforesto”,ChineseAcademyofForestry,Fuyang,Zhe jiangProvince310029,ChinaActaEcologicaSinica.2008.28(9):4430~4439.Abstract:Innaturalecosystems,plantsconstitutesuitablehabitatsandniches forthecolonizationofadiversityofmicroorganisms.Endophyticfungi,livingintheinnerofhealthyplanttissues ubiquitously,exhibitcomplexinteractionswiththeirhosts(acontinuumofmutualism,antagonismandneutralism)anddi fferenttransmissionmodes(horizontalorvertica1).Plantnutrientacquisitionandstresstolerancemaybestrengthenedbythefungalsymbiont.However,a successfulplant—fungalendophyteinteractionrequiresthefinetuningofant agonisticinteractions,whichimpliesthe evolutionofuniqueself—regulatingmechanismsonbothsidestomaintaina mutualisticinteraction.Hostspecies,fungal geuotypeandenvironmentalfactorssuchasclimateandsoilpropertiesaswell asthenutritionalstatusoftheplantwill ultimatelydeterminetheoutcomeoftheinteraction.Additionally,ternaryinte ractionswithmycorrhizalfungiand mycovirusesmayalsobeimportantcomponentsparticipatingintheplant?en dophyteassociationand,thus,influencethehostphenotype,Inthisreview,weattempttooutlinetheinteractivemechanismsof plant—fungalendophyteassociationsonamorphological,physiologicalandmolecularleve1.KeyWords:endophyticfungi;hostplant;interactivemechanisms基金项目:国家自然科学基金资助项目(30600002)收稿日期:2008—0227;修订日期:2008—05—23作者简介:袁志林(1979~),男,苏州人,博士生,从事植物?共生真菌基础理论与应用研究.E-mail:zhi—*********************}通讯作者Correspondingauthor.E—mail:******************.cn;***************.e nFoundationitem:TheprojeetwasfinanciallysupportedbytheNationalNatur alScienceFoundationofChina(No.30600002)Receiveddate:2008—02-27;Accepteddate:2008—05—23 Biography:YUANZhi—Lin,Ph.D.cadidate,mainlyengagedinplant—fun galendophytesinteractions.E-mail:zhi一/in—*****************http://www.ecologica.ca9期袁志林等:植物与内生真菌互作的生理与分子机制研究进展植物与周围环境生物的互作是一种普遍现象,其中植物-微生物的相互作用是重要形式之一.在叶围(phyllosphere)和根围(rhizosphere)区域,植物体时刻与众多的有害,有益和中性微生物同生存,并产生直接或间接的接触.在长期的协同进化过程中,植物对微生物的侵染已经形成一种适应性的机制,既能够识别来自微生物的信号分子并作出相应的生理反应,包括亲和性的互作(compatibleinteractions)和非亲和性的互作(incompatibleinteracti0ns).植物为了适应复杂的生态环境,进化成很多形式的植物?微生物共生体系统.C同位素标记技术显示根系组织中存在相当复杂的植物一真菌,植物一细菌共生体,对提高植物抗性和促进养分吸收起重要作用].现有的一些结果表明,植物与真菌的相互关系可能更为古老,化石证据证实了早在4亿多年前就已经存在植物与真菌的共生体,而且这种共生关系在植物由水生演化到陆生过程中扮演了重要角色口].在自然界中植物与真菌的非致病性的共生关系(nonpathogenicsymbiosis)具有普遍性,而致病性的亲和互作只是个例].因此研究植物.共生真菌的相互关系有助于更好阐明真菌在自然界的生态功能.有两类共生真菌能够侵染并定植在植物组织中,其中菌根真菌(mycorrhizalfungi)只与植物根系建立互惠关系,在根部组织中能形成特定的功能性结构,如AM菌根中存在丛枝和泡囊,外生菌根中可形成菌套(mantle)和哈帝氏网(Hartignet)等,涉及土壤一真菌.植物根系3个界面;而内生真菌(endophyticfungi)普遍存在于植物的地上和地下部分,且只存在植物组织一真菌的互作界面,侵染根系的内生真菌也不形成明显的结构特征.从已积累的研究结果看,内生真菌一植物共生体可能远比菌根共生体复杂.植物一内生真菌共生体是继豆科植物.根瘤共生体,菌根共生体后发现的植物与微生物共生关系的又一种表现形式,业已成为国际研究热点.但从研究历史,深度看,与病原真菌,菌根真菌比较,内生真菌的工作才处于基础探索阶段.在近10a中,在植物内生真菌的研究领域已经取得了很大的进步,从传统的分离培养,类群分析和分类鉴定逐渐过渡到从形态,生理和分子水平阐明互作机理.本文针对这几个方面的问题加以详细叙述和评论.1内生真菌定义及不同类型的生物学特性比较有关内生真菌的定义,国际上一直存有争议,现在普遍接受的是Petrini提出的概念J,即生活史的一部分能侵染并定殖在植物组织器官中,宿主无明显感染症状的一类真菌.绝大部分属于子囊菌和半知菌,担子菌和卵菌作为植物的内生菌也有报道..‟.一般将内生真菌分为两个类群,即禾草内生真菌(grassendophytes orbalansiaceousendophytes)和非禾草内生真菌(non—grassendophytes).它们之间的区别见表1ts,38].表1禾草内生真菌和非禾草内生真菌生物学特性比较[8,38]Table1Comparisonofcharacteristicsoftwogroupsofendophyficfungiinpla nk[8,38]禾草内生真菌Endophytesofgrasshosts非禾草内生真菌Endophytesofnongrasshosts种类较少,主要是麦角菌科的子囊真菌Fewfungalspecies,Clavicipitaceae侵染宿主组织面积广,胞问定殖Extensivecolonizationofgrassleavesandsheath,intercellular与宿主是”组成性互惠关系”,对宿主的增益作用明显“Constitutivemutualism”系统性种子垂直传播,但ichlo#也可水平传播V erticallytransmittedviaseeds绝大多数子囊菌,半知菌和和少数的卵菌,担子菌Manyspecies,taxonomicallydiverse主要以局部组织的定殖侵染为主,胞内或胞问生存Restrictedcolonizationofplanttissues,intracellularorintercellular与宿主是”诱导性互惠关系”,多因素影响互作结果“Inducedmutualism‟‟or”conditionalmutualism”.manyfactors determiningtheoutcomeoftheassociation非系统性孢子水平转播,极少数可垂直传播Horizontallytransmittedviasporesfromplantstoplants2内生真菌侵染宿主的形态学,生理学机制及双方各自的生理反应与菌根真菌和病原真菌相似,内生真菌的侵染过程也经历一系列复杂过程,包括孢子与亲和性宿主的识别,活化,黏附表面基质,萌发直到侵染组织内部_9.在侵染早期,双方各自均释放信息素类似物并被彼此识别,由此在代谢水平上做出相应的调整.在丛枝菌根和外生菌根研究中发现,气生菌丝具有对宿主植物根系定位的能力,在真菌和根系非直接物理接触前就开始了信号的交流”].已经证明根系分泌物中的倍半萜类,黄酮类物质充当了信号分子,促进孢子萌发.在内生真菌与宿主互作中也发现有类似现象,通过建立体外共培养技术(invitrodualculturesystem),观察内生真菌与无菌苗(或愈伤组织)共培养时各自的生理反生态28卷应,研究表明内生真菌菌丝对宿主的某些化学物质具有趋化性(chemotaxis),且生长良好;而在非宿主组织中并无此现象,表明如果在内生真菌一宿主互作中存在明显的趋化信号,该内生真菌并非仅仅是随机性侵染体(incidentalopportunists),而是在进化过程中已经形成对宿主的适应机制|I引.另一方面内生真菌的成功侵染也必须穿透宿主的机械屏障并克服其防御反应.现已发现内生真菌产生的胞外酶系统也相当丰富,如纤维素酶(cellulases),漆酶(1accase),木聚糖酶(xylanase),蛋白质酶(protease)等,在侵染初期破坏植物角质层和皮层细胞的细胞壁,利于菌丝进入组织内部.与菌根真菌相似,植物对于这类共生真菌的侵染所引发的防御反应程度较弱_1,推测一方面内生真菌携带的低毒力因子,菌丝分泌某些糖蛋白或细胞壁组成的改变,从而削弱宿主对其强烈的识别能力8j.最近发现在拟南芥根部细胞内质网中存在一种p一葡萄糖苷酶(PYK10),能限制内生真菌印度梨形孢(Piriformosporaindica)的侵入,从而削弱宿主产生的防御反应.植物组成型次生代谢产物如皂角苷(saponin),精油(essentialoils)等是一种进化形成的抵抗外界病原物的机制.某些病原真菌为了成功侵染产生相应的策略,分泌解毒酶(detoxificationenzyme),能分解利用次生代谢产物,克服宿主的防御系统¨博.内生真菌也有类似的机制,而且内生真菌对次生代谢产物的生物转化能力在一定程度也定了决定其宿主的范围.侵染组织后,为了行使特定的生物学功能,互作双方在形态,生理和分子水平均发生着深刻的变化,从而建立稳定有效的共生体.由于绝大多数内生真菌在细胞间隙繁殖生存的,因此质外体空间(apoplasticspace)是两者信号物质,营养交换的主要场所.对P.indica.大麦共生体系的生理与分子机制研究发现:菌株侵染根系后能削弱根部细胞HvBI一1基因的表达,HvB1—1基因的过表达反能限制菌丝的侵染强度.HvBI一1基因在真核生物中很保守,能抑制细胞程序性死亡,这表明菌丝在宿主体内的生长和繁殖需要植物组织细胞一定程度的死亡,最终两者达到平衡状态.但在侵染过程中,真菌在识别新的环境(如在宿主胞外体空间存在的碳水化合物等)中所发生的一系列生物学行为和反应机制,到目前为止涉及很少.有研究者利用激光共聚焦技术观察了GFP标记的哈茨木霉(Trichodermaharzianum)菌株侵染番茄根系时菌丝形态水平的变化,结果发现共培养2d后菌丝顶端呈现酵母状的乳突型细胞,推测这种特异性的形态变化有助于双方营养的交换.在植物.病原真菌互作研究中,已经发现菌丝的程序性死亡(programmedcelldeath)或自噬(autophagy)对于其成功侵染组织是必需的‟引.研究内生真菌不同侵染时期的基因差异表达,有助于了解其互作的关键调控因子以及分析比较不同植物一真菌相互关系的异同.3内生真菌改变植物生理代谢并增强宿主抗逆性的现象和机制分析过去的研究往往只关注菌根真菌对植物个体,种群和群落结构的影响,而忽视了内生真菌的生理生态功能.自发现禾草地上部分内生真菌Epichlo~/Neotyphodium在增强宿主生物量,抗逆方面具有独特作用,科学家普遍认为植物体进化形成不同类型的真菌共生体对其生存起着关键作用.随着研究的深入,对于水平传播内生真菌的生物学特性也逐渐涉及,尤其在探讨根系内生真菌增强宿主生理功能的机制方面尤为活跃.研究内生真菌自身的生物学特性及其对植物初级代谢和次级代谢的调节有助于我们更好地阐明植物受益的机理.内生真菌赋予植物优良生长性状的特点与菌根真菌类似,如促进植物营养生长,光合作用增强,增加生物量(产量)并提高在逆境中的生存能力.担子菌P.indica能分泌生长素如吲哚乙酸促进植物生长…,内生镰刀菌通过抑制植物体内的乙烯信号途径来提高植物生长活力¨.内生真菌还能通过活化硝酸还原酶,分泌铁载体和磷酸酶等形式促进植物养分吸收,从而更利于植物生长(图1)拍.有些真菌还能分泌多糖类黏液物质,并在根表面形成菌膜(biofilm),协同植物抗旱.Carroll 归纳总结了内生真菌互利共生的5种特性:(1)该内生真菌在特定的宿主植物中普遍存在,地理分布较广,植物不表现任何明显的病症;(2)该真菌能垂直传播或水平传播的效率很高;(3)内生真菌在整株植物组织均能生长定殖,如果只在某一器官中生长,则该组织器官感染内生真菌的强度比较高;(4)内生真菌能分泌毒性或抗生物质;(5)该内生真菌在分类单位上与病原物拮抗菌很接近.因此通过常规的菌株分离,鉴定工作,并结合上述原则,可以筛选出具有9期袁志林等:植物与内生真菌互作的生理与分子机制研究进展特定生物学功能的内生真菌.从热带兰科植物根系中分离出内生真菌,其中有两个菌株Trichodermachlorosporum和Clonostachysrosea,在分类单位上均属于生防菌株,接种试验表明这些菌株能显着提高无菌苗移栽成活率和生物量.植物育种专家通过现代分子生物学技术已经初步阐明植物体存在一些抗逆基因来参与逆境调节,但至少有很多的工作表明植物体的某些抗性特征与内生真菌的存在有关.甚至有专家预测这种内生真菌生物技术可能与传统的抗逆育种和转基因培育技术并驾齐驱.无论是垂直传播内生真菌还是水平传播内生真菌都能增强在生物(病原物)和非生物胁迫(热,盐胁迫等)中的生存能力.内生真菌协同植物适应极端环境有3种假说:①适应性生态位共生(habitat—adapted symbiosis),既一种胁迫环境下植物的内生真菌不能使另一种逆境条件下的植物受益,只能增强原胁迫环境下植物的生理功能;如耐盐植物内生真菌对热胁迫植物无生物学效应;农作物抗病内生真菌对植物耐盐无贡献等.这种植物通过共生真菌的功能来实现抗逆是一种基于基因组间的表观遗传学作用机制(intergenomic epigenetics).②共生体调节(symbioticmodulation),植物,内生真菌双方由于受到周围微环境变化的选择压力,具有选择性地改变另一方的能力.这可能是一种跳一分布于叶片组织间隙的内生真菌菌丝————————卜一EndophyticroyceIiumoccur叶围PhyllospheretInIecellul|dr1yinleavestissues①次生代谢产物②诱导子③铁载体④植物激素⑤侵染定殖…一一一………根-R.ph.陀,根系内生真菌Rootfungalendophytes菌根真菌Mycorrhizalfungi图1植物地上和地下部分组织真菌共生体及可能的生理功能【~]Fig.1Fungalsymbiontsoccurringinabovegroundandbelowground planttissuesandpossiblefunctionstheyplayed①Secondarymetabolites;②Elicitors;③Siderophores;④Phytohormones;⑤Infectionandcolonization;⑥Promotinghost growthandyield;⑦Induceddiseaseresistance;⑧ConferringhostI℃sistancet0abi0ticstress【-36]跃式的协同进化方式,使共生体快速适应环境胁迫.③共生体生活方式的转换(symbioticlifestyleswitching),某些所谓的”病原真菌”,如一些炭疽菌属真菌(Colletotrichumspp.),在一种植物中表现致病,而在另一宿主中却表现互惠共生;病原真菌的单基因突变引起其生活方式的转变,由致病性向互利共生性转化引.但这些假说几乎都基于对现象或通过接种实验来证明,深层次的机理挖掘却很少.在非生物逆境中(如盐胁迫),受内生真菌侵染的植物能更快识别周围的逆境,而且可能通过增强植物组织脯氨酸(proline)的生物合成来消除活性氧的积累_3J.最近有研究者发现一种根际真菌Paraphaeosphaeriaquadriseptata能分泌一种能抑制植物热激蛋白HSP90活力的活性物质,从而提高植物的耐热程度,是否在内生真菌一植物互作体系中也存在类似的作用机制值得深入探讨帅J.目前对于内生真菌的抗病机制的认识还处于起步阶段,但不外乎以下几种途径:(1)分泌抗生物质;(2)生态位竞争;(3)重寄生作用;(4)诱导植物抗性47I引.如禾草内生真菌在离体(invitro)或活体(inplanta,invivo)条件下均能产生一系列生物碱类物质,能有效抗虫;非禾草内生真菌增强植物抗虫能力的发现是源于对虫生真菌的(entom叩athogenicfungi)生态功能的重新认识,某些感染病原虫体的真菌在植物组织中具有内生性特点,对虫体具有很强的抗性.研究最透彻的虫生真菌是Beauveriabassiana,能够与玉米,棕榈,咖啡树和罂粟等共生.B.bassiana能分泌多种代谢产物如bassianin,beauvericin,bassianolide,beauveriolide等,对病原虫体均产生毒害作用.植物体对病原物的抗性有两种不同的机制,即系统获得抗性(systemicacquiredresistance,SAR)和诱导型系统抗病性(inducedsystemicresistance,ISR).SAR反应主要受到病原菌,生防真菌或一些化合物诱导产生,显着特征是水杨酸,茉莉酸和乙烯等物质参与,并伴有病程相关蛋白(pathogenesis—relatedproteins,PR)如几生态28卷丁质酶,葡聚糖酶和多酚氧化酶等的积累,而ISR反应并没有PR蛋白的积累.早前人们对于植物根圈促生细菌(plantgrowthpromotingrhizobacteria,PGPR)引起植物诱导型系统抗病性有深入的认识.近期的研究也表明内生真菌也能诱导植物体产生相似的抗性反应_5卜j,如菌丝分泌的某些小分子蛋白或代谢产物能够作为诱导子引发植物抗性;植物组织内的谷胱甘肽?抗坏血酸代谢途径(glutathione—ascorbatepathway)被激活,从而增强了细胞抗氧化能力引.但内生真菌诱导植物抗性可能有其独特性和复杂性的一面,P.indica和Sebacinavermifel‟a接种烟草的试验表明,这两种内生真菌均能有效促进植物生长,但同时伴随着对烟草天蛾(Manducasexta)抗性能力的下降,结果证明了内生真菌的侵染导致植物体内一种防御蛋白活力的降低一胰蛋白酶抑制剂(trypsinproteinaseinhibitors,TPI)[57j.在镰刀属内生真菌(Fusariumsolani)也发现类似的现象,F.solani接种番茄对病原尖孢镰刀菌(F.oxysporum)的抗性是依赖乙烯合成途径的,但S.vermifera促进烟草生长却是通过抑制乙烯信号转导来实现的.有趣的是,将P.indica 接种大麦后却显现出多重有益效应,能同时抗病,耐盐和提高产量,这项研究结果对传统的观点:”植物抗病,增产不可兼得”提出了挑战.这些有趣的现象提示:在不同基因型的植物一内生真菌互作中所产生的表型可能会发生变化,而且不同的功能性内生真菌进化程度可能存在差别,有些是潜在的病原菌,而有些则是”真正内生菌”(trueendophytes)Lsj.很多报道表明PGPR微生物如假单孢菌属(Pseudomonasspp.)细菌和菌根真菌对根部土传病害的病原菌都具有良好的防治效果,但对植物地上部分病原物的防治能力有一定的局限性引,除非这些微生物能诱导植物系统获得抗性;而内生真菌能稳定存在于植物的整个组织器官中,受到植物体本身机械组织的保护,因此对士传,气传病原物的抗性持久有效.虽然大多数内生真菌增强植物抗病性的报道还多局限于室内盆栽试验,是否在田问试验中也有类似的功效是值得关心的问题.鉴于木本植物内生真菌种群多样性,植物体的抗性反应可能得益于诸多内生真菌发挥生理功能的总和,因此考虑优势内生真菌之问的协同作用可能使植物受益更明显].从大量的研究结果看,植物不同组织器官富含的内生真菌对植物的贡献能力也有差异,这对筛选功能性菌株具有一定的指导意义.根系和叶片组织是植物体受外界生物和非生物因子胁迫最大的部位,因此根系和叶片内生真菌对于保护植物组织免受伤害起着尤为重要的作用.归纳目前的研究结果并加以分析可以看出,内生真菌对植物各组织器官生理效应的影响能力有以下趋势:根系>叶片>茎.植物体根围区域是一个能量和物质交换异常活跃的区域,植物和土壤微生物相互影响,而且根系被认为是一个动态的”碳库(carbonsink)”,营养物质充足,是众多微生物相互竞争的部位j.因此在这种复杂的环境中植物的生存策略之一就是容纳了与之互惠共生的内生真菌;而且根系内生真菌的侵染程度较地上部分要强(extensiveorsystemiccolonization),而叶片内生真菌多以局部侵染为主(1ocallylimitedcolonization).红色不育真菌(SRF,sterileredfungus)和暗色有隔内生菌(DSEs,darkseptateendophytes)是根系非菌根共生真菌的典型代表鲫‟.,某些DSEs甚至能形成类似菌根的侵染结构,国外已经有很多综述文章对这两类真菌的生理生态功能加以评论,虽然存在相矛盾的观点,但不容置疑的是这种在植物根系普遍存在的真菌群体必然扮演着特殊的角色,如Phialocephalasp.能降解根部老化和木栓化的细胞,从而有利于减少植物体能量的耗损.需要指出的是,某些土壤习居菌如一些木霉属(Trichodermaspp.)真菌,镰刀属真菌(Fusariumspp.),也能侵染根系皮层细胞成为无毒”机会性”内生真菌,发挥着与其作为生防菌株类似的功能.可见要严格区分内生真菌和土壤真菌是有一定难度的,长期的选择进化改变了真菌的生活方式.4内生真菌-植物互作体系中的重要分子元件及调控机制当前关于豆科植物菌根共生体和根瘤共生体建立过程中的信号转导研究比较透彻,揭示出植物与微生物形成共生关系具有很大的保守性.研究证实至少有3个植物信号元件参与其中,分别是受体样激酶(DMI2),离子通道(DMI1),依赖钙调蛋白的激酶(DMI3).此外还发现其他重要功能蛋白,如植物质体蛋白和某些核孔蛋白对于真菌和细菌进人根系形成共生关系也至关重要.研究发现日本百脉根(Lotusjaponicus)中存在两种同源的质体蛋白基因(CASTOR和POLLUX)和编码一种核孑L 蛋白的基因NUP85,主要调节质体与胞液9期袁志林等:植物与内生真菌互作的生理与分子机制研究进展之间的离子流量并激活ca信号,被认为是植物与微生物形成内共生体所必需的保守元件..但至今还很少报道有关内生真菌共生体中信号分子的化学本质和维持这种互惠关系的必备元件.在拟南芥突变体Pii.2根部细胞质膜中分离到两个富含亮氨酸重复体(LRR)的蛋白质:At1g13230和A~g16590.其中Atlg13230含有一个内质网滞留信号,A~g16590是一种非依赖磷酸化的信号转导中的受体蛋白,参与识别P.indica的侵染.这两种蛋白对于P.indica发挥生理功能至关重要;但拟南芥的DMI-1突变体却未影响与P.indica的互惠关系,这可能说明植物体已经进化形成多种重要蛋白分子以此来响应不同的微生物类群.与此对应的是,在内生真菌中也存在一些关键的酶基因来执行特定的功能,这是共生体双方相互适应的结果.BarryScott研究小组长期致力于禾草内生菌与宿主的相互关系,阐述了维持这种动态平衡的分子机制,内生真菌Nox基因可以通过调控胞间合成ROS(活性氧)来控制菌丝在宿主中的生物量,从而建立互惠共生关系;Nox的突变体菌株侵染牧草能导致植物严重感病坏死,而且菌丝在组织中生长杂乱无序J.进一步的深入研究发现在E.festucae中的SakA基因编码一种MAP激酶,能调控Nox复合体的活性,一突变体导致ROS增加加;此外在禾草内生真菌中还克隆到一个编码非核糖体多肽合成酶(nonribosomalpeptidesymthetase,NPS)的基因,该基因参与合成铁载体(siderophores),同样NPS基因的突变体导致不能正常合成铁载体,也导致植物的病变坏死,表明内生真菌分泌铁载体能力的缺失改变了共生体铁离子的动态平衡(ironhomeostasis),引起转录水平的重调,最终导致其互惠性向拮抗性转化”J.5内生真菌对宿主表型的可塑性绝大多数内生真菌发挥有益生理功能是在一定条件下实现的,与宿主是诱导型互惠关系.根据内生真菌起源于病原真菌这一观点,Schulz和Boyle等提出了维持植物一内生真菌和谐共生的动态拮抗平衡假说(balanceofantagonisms),既只有内生真菌携带的毒性因子和植物的防御反应处于相平衡状态,才能建立稳定有效的共生体,任何一方的失调就能打破这种平衡关系,导致内生真菌不能成功侵染宿主或使植物感病.诸多因素能够影响内生真菌一植物互作结果,其中宿主基因型和生理状态,真菌基因型,环境因子和土壤的营养水平是最主要的因素.如上所述,Freeman等在1993就发现致病刺盘孢菌(Collectotrichumspp.)的一个单基因突变就能使其生活方式由致病性向互惠内共生性转变.后来证明这个单基因位点编码一种胞外丝氨酸蛋白酶.后来他们又发现,同一种病原菌,接种在不同植物产生的表型却不一样,有的致病,有的却表现互惠共生.特别是在水平传播内生真菌中,表现出了灵活的生活方式,在植物正常生理状态下呈现互惠共生(mutualism),而在宿主遭受逆境胁迫下有表现寄生性(parasitism);在当植物组织衰老死亡时,腐生(saprophytism)生活成为其主要方式.也许正是内生真菌所显现的表型可塑性(phenotypicplasticity)给研究带来了一定难度.6菌根真菌,内生真菌和植物三者之间的关联以上讲述的均是单一的植物一内生真菌互作,但在自然界中,植物体却能够容纳丰富的微生物类群,因此在植物个体组织中也必然存在复杂的植物一微生物,微生物一微生物之间的相互作用.利用免培(cultureindependent)环境PCR方法,提取植株健康根系总基因组DNA,通过真菌特异性引物扩增发现,根系中存在丰。
江南大学微生物名词解释汇总

江南大学微生物名词解释汇总接合孢子(zygospore):是由菌丝生出形态相同或略有不同的配子囊接合而成的。
是真菌产生的一种有性孢子,二倍体,近圆形,休眠,大,深色。
P108A局限性转导:指通过部分缺陷的温和噬菌体把供体菌的少数特定基因(供体细菌染色体上原噬菌体整合位点附近少数基因)携带到受体菌中,并与后者的基因组整合、重组形成转导子的现象。
P227B碳源物质:凡可以作为微生物细胞结构或代谢产物中碳架来源的营养物质,均可以作为微生物的碳源。
包括糖类及其衍生物、有机酸、脂肪、烃类和CO2、碳酸盐等物质。
P179A芽孢:某些细菌在其生活史的一定阶段(生长发育后期),在营养细胞内形成的一个圆形、卵圆形或圆柱形、厚壁、含水量极低、抗逆性极强的休眠体。
产芽孢的型细菌主要有芽孢杆菌属和梭菌属。
(因为细菌的芽孢都是在内形成的,故又称内生孢子。
基团移位:这是一类既需特异性蛋白载体参与,又需耗能的的一种物质运送方式。
是指被运输的基质分子在膜内经受了共价的改变,以被修饰的形式进入细胞质的运输机制。
由于这种运输是在磷酸进团发生移位的过程中实现的,所以叫基团移位。
P191移码突变:当基因突变时,在DNA序列中由于一对或少数几对核苷酸的插入或缺失(不是三的倍数)而使其后的全部遗传密码的阅读框架发挥僧移动,进而引起转录或翻译错误的突变,它一般只引起一个金银的表达出现错误。
P313 P208B生长因子:某些生生物不能从普通的碳源,氮源合成,而需要另外加入少量来满足生长需要的微量有机物质。
其中包括氨基酸维生素嘌呤嘧啶及其它们的衍生物,脂肪酸及其它的膜成分。
P185准性生殖:(霉菌的基因重组)是一种类似于有性生殖,但比它更为原始的一种两性生殖方式,这是一种在同种而不同君主的体细胞见发生的融合,他可不借减数分裂而导致低频率的基因重组并产生重组子。
因此,可以认为准性生殖是在自然条件下,真核微生物体细胞见的一种自发性的原生质体融合现象,它是某些真菌尤其在还未发现有性生殖的半知菌类中常见。
菌膜名词解释

菌膜名词解释
菌膜(Biofilm)是一种微生物附着在固体表面或其自身相互粘附形成的聚集体。
这个聚集体由一个或多个微生物种类组成,并被一个自我生成的生物膜所覆盖。
这个生物膜是由微生物分泌的胞外聚合物(EPS)组成的,它可以将微生物包裹在其中,并为其提供保护。
菌膜的形成是一个复杂的过程,包括微生物的粘附、生物膜的生长和成熟以及生物膜的脱落等阶段。
在这个过程中,微生物会不断分泌EPS,这些EPS会包裹住微生物并形成一层粘性物质,将微生物聚集在一起形成菌膜。
菌膜在自然界和工业生产中都有广泛的存在,例如在管道、冷却塔、土壤、医疗器械等领域都有可能出现。
由于菌膜的存在可能会影响工业生产的效率和产品质量,因此对菌膜的控制和预防也是非常重要的。
细菌生物膜 口腔 Biofilm

extraradicular factors
there are six biological factors that lead to asymptomatic radiolucencies persisting after root canal treatment.
(i) intraradicular infection persisting in the complex apical root canal system; (ii) extraradicular infection, generally in the form of periapical
生物膜与疾病(总结)
细菌生物膜 进入体内的 可能途径
细菌生物膜的基本性质
生物膜1978年由J. William Costerton等通过电镜 方法首次观察到 Biofilm定义:指细菌自身产生的外部多糖基质、
纤维蛋白质、脂蛋白等包裹着的菌细胞的结构。
• 生物被膜是细菌的一种具有保护性的生长模式,是细胞间 相互协调作用的复杂的多细胞群体,具有结构和代谢复杂性。 • 形成生物被膜的黏附细菌群也可以释放出浮游细菌,是潜 在的持续感染因素。
main and accessory canals, their ramifications and anastomoses where
residual infection can persist.
Further, there are – located within the inflamed periapical tissue – that can interfere with post-treatment healing of apical periodontitis. The causes ……have not been well characterized, but
植物内生细菌的生物薄膜(biofilm)

关 键 词 : 生物 薄 膜 ; 植 物 内生 细 菌 中图分类号 : Q 3 9
引 用 ( n uR vMi oi} 重 要 期 刊 所 归 纳 发 表 ( n e c bo 等 A r l
的定 义 。生 物 薄膜 是 指 微 生 物 菌 体 互 相 黏 附在 一 起 或 附着 到 一 些材 料 表 面上 .并 由胞 外 多 聚物 基
对 遗传 信息 交 换 和菌 体 协 同作 用等 也 有重 大 意 义 :
许 多 细 菌 能 定 殖 在 植 物 的根 、茎 、叶 表 面 或 组 织 内部 .产 生 植 物 微 生 物 相 互 作 用 的适 应 性 体
最 近 .该 种 结 构 才 被 发 现 与某 些 生 物 学 功能 或 属 性 有 关 .进 而 成 为植 物 微 生 物 相互 作 用 研 究 的热 点 .生 物 薄 膜 在 植 物 一 菌 相 互 作 用 中所 扮 演 的 细 角 色 也越 来 越 受关 注 近来 的研 究 又进 一 步 发 现 .许 多 普 通 细 菌 如 大 肠 杆 菌 ( sh r hacl) 枯 草 芽 孢 杆 菌 ( a E c ei i o 和 c i Bc.
别 。生 物 薄 膜 内 的 菌体 细 胞 密 切 协调 。而生 物 薄
膜 体 系 外 的 菌 体 则 不 具 有 这 样 的相 互 作 用 [ 人 2 1 们 最 初 观 测 到 细菌 与植 物 相 互 作 用 形 成 生 物 薄 膜
度依 赖 表 现 型 的表 达 ’ 比如 。对 某 些 有密 度 依 6 ] 。
细菌生物被膜(bacterial biofilm)的研究进展

细菌生物被膜(bacterial biofilm)的研究进展陈维贤;张莉萍【期刊名称】《微生物学杂志》【年(卷),期】2004(024)001【摘要】细菌生物被膜由物体表面集聚生长的细菌群落和细胞外基质构成,植入性医用器械表面较多见,其结构包括主体生物被膜层、连接层、条件层和基质层.细菌之间的信号传导影响着生物被膜的异化形成.生物被膜相关感染治疗较难,易慢性化及反复发作.抗生素或其他化学杀菌剂及金银包裹导管等医用材料表面是常用的预防方法.已形成的生物被膜可用物理方法或某些抗生素清除,而生物学控制是另一可能途径.【总页数】3页(P46-48)【作者】陈维贤;张莉萍【作者单位】重庆医科大学,附属第二医院,重庆,400010;重庆医科大学,附属第二医院,重庆,400010【正文语种】中文【中图分类】Q939.1【相关文献】1.Advances in Research on Resistance Mechanisms of Bacterial Biofilms [J], Yanfei LIU;Jiande YANG;2.Bacterial biofilms in human gastrointestinal tract: An intricate balancebetween health and inflammatory bowel diseases [J], Niharika Chandra; Ankita Srivastava; Sunil Kumar3.Exogenous Protein as an Environmental Stimulus of Biofilm Formation in Select Bacterial Strains [J], Donna Ye;Lekha Bapu;Mariane Mota Cavalcante;Jesse Kato;Maggie Lauria Sneideman;Kim Scribner;Thomas Loch;Terence L. Marsh4.Effect of the Suppression of Cariesitic Bacterial Growth and Biofilm Formation Using Hydrogen/Free Chlorine Mixed Water Produced by an Electrolysis-Type Hydrogen Generator [J], Ryusuke Suzuki;Rin Kaneko;Masaru Nakano;Masato Kanda;Yuji Sebata;Shota Motegi;Tomoko Abe;Kazuaki Muramatsu5.Current Concepts of <i>Gardnerella vaginalis</i>Biofilm: Significance in Bacterial Vaginosis [J], Allison Nicole Kunze;Bryan Larsen因版权原因,仅展示原文概要,查看原文内容请购买。
周质空间蛋白的抗逆机制

第18卷 第8期 2008年8月3专题评述3周质空间蛋白的抗逆机制3马元武 陈翠翠 何 晴 冯永君33北京理工大学生命科学与技术学院,北京100081 2007212228收稿,2008201229收修改稿 3国家自然科学基金资助项目(批准号:30400002) 33通信作者,E 2mail :fengyj @摘要 细菌细胞周质空间处于G -细菌与外界相互作用的前沿界面,对生存环境的波动非常敏感,是菌体细胞与外界环境进行相互作用的第一道媒介(趋利作用)或屏障(避害作用),因此,周质空间蛋白及其抗逆作用的相关研究对于探究与理解微生物生存适应性问题具有重要意义.周质空间蛋白在长期进化过程中形成了一套对环境波动的适应机制,文中系统总结了G -细菌细胞周质空间蛋白的分类、定位和抗逆机制,着重介绍了周质空间蛋白在抗酸机理中和RND 转运系统在菌体抗逆生存中所起的重要作用,并讨论了周质空间蛋白通过影响细菌生物薄膜的形成而起到间接抗逆的作用.关键词 周质空间蛋白 抗逆作用 环境胁迫 细菌细胞周质又称膜间质,指位于大肠杆菌(Escherichi a coli )等革兰氏阴性细菌细胞内膜和外膜之间的夹层空间,其大小随环境与胞质间渗透压的变化而改变,约占整个细胞体积的24%—40%[1,2].外膜上存在较多非特异性的孔道蛋白(porins ),能够允许分子质量小于600—1000u 亲水小分子物质的自由扩散,而不需要消耗能量[3—5].由于这种分子筛效应的存在,使得周质空间蛋白对波动的生存环境比较敏感.这些周质空间蛋白在G -细菌长期的进化过程中逐渐形成了多种适应环境波动的机制,在维持周质空间蛋白活性和细胞的生理功能中起重要作用11 周质空间蛋白周质空间呈胶状,主要由一些单糖,寡糖,蛋白质组成,此外还包含一些其他可溶性物质.其中作为周质空间重要组成部分的蛋白质在细菌营养代谢,物质转运,信号传导及能量代谢中起着重要的作用.由于周质空间所处位置的特殊性,使得周质空间蛋白比其他物质更容易受到外界环境的影响,这就要求周质空间蛋白能够适应经常波动的外界环境[6].目前已知的周质空间蛋白按功能大致可以分为以下几种:(1)结合蛋白,该类蛋白作为转运系统的重要组成部分,与底物结合后形成转运复合物,再与内膜转运蛋白发生相互作用,依靠后者的A TP 酶活性亚基水解A TP 释放的能量将底物转运进入胞内.到目前为止,至少发现80种以上的周质空间结合蛋白,其中包括麦芽糖、阿拉伯糖结合蛋白,谷氨酸,亮氨酸,赖氨酸结合蛋白等,这些周质空间蛋白主要转运一些单糖,寡糖,氨基酸,小肽等小分子物质[7].基因组学和蛋白质信息学等的研究发现,在大肠杆菌中这类蛋白与膜结合蛋白形成一个最大的蛋白家族,约占细菌所有ORF (开放阅读框)所编码蛋白的6155%[8].(2)水解酶类,如酸性磷酸酶,碱性磷酸酶,148UDP2葡萄糖水解酶,L2天冬酰胺酶,羧肽酶II等将吸收的复杂大分子降解为简单的小分子物质,有利于营养物质的吸收,有些酶同时也参与胞壁物质的代谢[9,10].(3)合成酶类,如肽聚糖合成酶,SurA,Skp 蛋白等,这些蛋白在肽聚糖,鞭毛形成及细胞壁物质的组装过程中起作用[11].(4)受体酶类,如麦芽糖结合蛋白,半乳糖/葡萄糖结合蛋白等,其本质是作为结合蛋白参与底物的跨膜运输;同时,它们在结合底物后还能与膜上另外的蛋白受体相互作用,介导细胞对底物的趋化性,因此具有双功能性[12,13].这些具有多种功能的周质空间蛋白以及外膜蛋白的前体是在细胞内核糖体上合成后,经与胞内分子伴侣蛋白SecB结合,并在信号肽的引导下,穿过内膜上的Sec YEG孔道(其信号肽在跨膜过程中被膜上信号肽水解酶切掉)进入周质空间,然后在周质空间二硫键异构酶[14],脯氨酸顺反异构酶等的作用下形成有功能的成熟蛋白[11].对于许多外膜蛋白前体在细胞质内合成并通过Sec YEG孔道进入周质空间后,它们会在一些具有分子伴侣功能SurA, PapD,Skp等蛋白的作用下组装成特定的寡聚形式[15],然后整合成为有功能的外膜蛋白寡聚体[16].比如,外膜蛋白LamB就是在细胞质合成后,穿过Sec YEG内膜孔道,然后在SurA,Skp等周质空间蛋白帮助下组装为特定的寡聚形式并整合到外膜上去发挥功能,如图1所示.2 周质空间蛋白的抗逆特性当处于不良的外界环境条件时,G-细菌大都能表现出一定的抗逆特性,以适应生存环境的变化.研究发现,大肠杆菌能够在p H2的较强酸环境条件下生存数小时;一种能够引起囊状纤维病的铜绿假单胞菌(Pseu domonas aeru gi nosa)具有广谱抗生素耐受性[17];此外,还发现伯克霍尔德氏菌(B urk hol deri a cenocep aci a)能够在高温,高渗透压等环境条件下生存[18].周质空间处于细菌与生存环境相互作用的最前沿,对于环境条件的波动更为敏感.研究发现周质空间蛋白质相对于胞内蛋白来说也具有更强的环境适应能力.Liu等分别对全细胞,质膜内组分、周图1 LamB的成熟机制示意图LamB在细胞质内合成后以前体形式存在,并带有信号肽(画有阴影交叉线的条形图),SecB能够稳定并帮助LamB蛋白前体穿越质膜孔道系统(SecA,Sec YEG,SecDF,YajC).在周质空间分子伴侣SurA,Skp和DegP的帮助下转运到Yae T(包含Yae T,Y fgL,Y fiO,NlpB)进而组装到外膜上,成为成熟的外膜蛋白LamB[11]质空间组分分别进行高温,乙醇,盐酸,硫酸铜等逆境处理,发现周质空间蛋白比胞内蛋白具有更强的抗逆特性.对蛋白样品的三级结构和疏水性进行分析,发现周质空间蛋白经逆境处理后,虽然蛋白质的三级结构丧失,并且疏水区域外露,但是却不易聚集生成沉淀[19].相反,如果蛋白聚集就会给菌体带来巨大危害;在另一研究中发现,蛋白聚集会使菌体部分机能减弱/丧失,并伴随有病变产生,严重时会导致细胞死亡[20].正是由于周质空间的这些蛋白虽然丧失了三级结构却不易聚沉的特点,保护了细胞免受因蛋白聚沉所导致的病变乃至死亡[21,22].因此,可以推断周质空间蛋白较强的抗聚沉特性可能是菌体长期进化过程中形成的一种对外界环境的适应策略.研究进一步发现,大部分具有抗逆活性的周质空间蛋白可以在环境信号因子的诱导下产生.如Miyadai等发现,脂蛋白Nlp E的过量表达能够诱导DegP的表达[23],其中DegP既具有分子伴侣活性又具有蛋白酶活性,当变性失活的蛋白较少时,起分子伴侣作用,随着变性蛋白的不断增多,将发挥蛋白酶的活性降解变性蛋白,在维持细胞内环境稳248 第18卷 第8期 2008年8月定中起关键作用[21].当细胞受到外部环境压力影响时,能够诱导一些周质空间蛋白的表达,这些蛋白在细胞的抗逆生存中起重要作用.目前发现所有环境压力信号传导过程都有一个共同的特征:外界环境压力信号(如变性蛋白等)会被周质空间中分子所感应,并通过与内膜受体相互作用释放σE因子,转化成为胞内信号,胞内信号再与特定的基因相互作用诱导一系列具有抗逆活性的周质空间蛋白基因表达,以降低环境波动对细胞造成的伤害[24—26].如环境压力造成的部分周质空间蛋白或膜蛋白的变性会被一些周质空间的信号分子如DegS(一种蛋白酶)所识别,致使DegS蛋白抑制因子释放出来,引起DegS的水解,触发σE介导的信号通路,从而诱导一些在抗逆中起重要作用的周质空间蛋白如SurA, HdeA,DegP等的表达[27].这些周质空间蛋白,或能够在逆境下识别变性蛋白起分子伴侣/蛋白酶活性,或在结合/转运不利环境因子排出细胞中起作用,从而更利于细胞的抗逆过程.目前,对于G-细菌的耐酸机制和对重金属离子的抗逆机制方面的研究最为深入.一些非极端环境微生物如肠道病原菌志贺氏菌(S hi gell a)、大肠杆菌等能够在p H2条件下存活数小时,它们虽然可以在很低的p H条件下存活,但胞内的p H却接近中性,说明这些细菌具有一套机制来适应不良的p H条件.Richard利用微点阵技术研究了大肠杆菌抗酸机理,发现了12个与耐酸有关的基因簇称为A FI (Acid Fit ness Island),其中的四个基因簇ga dA, g ad X,gad W和g ad E通过谷氨酸的脱羧基作用形成一级胺,在脱羧基的同时消耗质子[28];另外h deB,y hi D,y hiU,y hiV四个A FI基因簇编码的蛋白在抗酸机制中起很少或并不起作用;剩余的4个基因簇中Yhi F(常规蛋白)和Slp(脂蛋白)所在系统可能一定程度上抵制酸效应,YhiD和HdeD作为膜组成蛋白可能在抵御质子的跨膜运输中起作用,HdeA(一种周质空间蛋白)在酸变性蛋白的质量控制中起作用[29].Gajiwala等研究发现,作为A FI耐酸基因簇重要成员之一的h deA是一些能够在低p H条件生存的肠道微生物(包括病原菌)所必需的h deA编码周质空间蛋白HdeA,在很低的p H条件下,HdeA 能够由二聚体解离成单体,这些单体具有分子伴侣活性,能与周质空间蛋白或膜蛋白因酸变性产生的疏水面相结合,而防止蛋白聚沉引发的细胞死亡[30].Hong等把HdeA在抵御恶劣酸性环境条件的特性归结为:HdeA具有分子伴侣活性,能够在酸性环境条件下结合变性的蛋白,并在中性条件下释放.这种特性是基于低p H条件下HdeA能够调整为一种有活性,低有序性的球状结构(这种结构未在中性条件下发现).酸性条件下HdeA低有序性的球状结构易于暴露其部分疏水作用面而结合变性蛋白.这些特征保证HdeA能够在酸性环境下结合变性蛋白,防止蛋白聚沉从而使细菌具有一定的耐酸能力[22].除了因环境压力诱导产生的一些周质空间蛋白在菌体的抗逆中起重要作用外,对于每种G-细菌来说还存在多种由底物结合蛋白,转运蛋白和膜蛋白组成的一种转运系统与菌体的抗逆作用有关,该系统由基因组或质粒编码,通常由跨内膜,外膜的蛋白组分以及周质空间蛋白组分3部分构成,该系统具有底物特异性,能结合并转运有害底物(抗生素、重金属盐等)排出细胞,它的存在表现为对重金属盐、抗生素等具有较强的抵抗能力,该类转运系统统称为RND(resistance2nodulation2cell divi2 sion)转运系统[31].其中比较典型的有AcrAB2 TolC,MexAB2OprM和MexXY2Op rM系统等[17].比如,Lee等在大肠杆菌Pco蛋白家族组成的RND 转运系统介导的铜离子的外排研究中发现,周质空间蛋白PcoA能够吸收铜离子,保护周质空间酶免受铜离子造成的损害;Pco E可能在最初的周质空间内铜离子的隔离中起作用;PcoC可能在周质空间结合铜离子,有利于铜离子的外排,PcoD可能在铜离子穿越细胞膜中起作用.该系统通过不同Pco蛋白之间的协调作用,把过量的铜离子排出体外起到抵抗重金属盐离子的作用[32].另外一项关于RND转运系统研究中,Perron 等发现铜绿假单胞菌的CzcR2CzcS转运系统(RND 转运系统中的一种)不仅能够转运重金属盐离子镉和钴等,而且能够转运碳青霉烯(一种抗生素).该系统所表现的多底物适应性是对传统RND转运系统底物特异性认识的一个挑战,同时该系统所具有的多重抗逆效应,对于微生物来说,不仅提高了对348 第18卷 第8期 2008年8月波动环境的适应能力,而且有效地利用了资源,这可能是生物长期进化过程中形成一种适应策略,这对于生物的生存、发展和繁殖都具有非常重要的意义[33].所以,RND转运系统作为G-细菌的一种重要抗逆机制,在抵御重金属盐,抗生素等多个方面起重要的作用.随着对G-细菌抗逆机制研究的不断深入,发现周质空间蛋白不仅自身具有比胞内蛋白更强的抗逆和防聚沉能力,而且许多在G-细菌抗逆机制中起关键作用,上述环境压力信号诱导和σE因子调节周质空间蛋白的表达,表达的周质空间蛋白通过结合变性蛋白,或在RND转运系统的结合/转运下将多种药物、重金属盐离子排出体外,在菌体细胞抗逆中起重要作用[31—33].3 周质空间蛋白通过影响biof ilm形成间接影响抗逆作用 大肠杆菌和铜绿假单胞菌等能够形成一种特殊的生物薄膜(biofilm)[34,35],以适应特定生存环境和生长阶段的需要[36,37].相关研究在植物内生菌方面体现得较为明显[37—39].宋未从水稻品种“越富”中分离到一株植物内生菌成团泛菌(Pantoea a g glo2 merans)YS19[40],我们发现该菌具有固氮和分泌生长素促进水稻生长的能力[40—42],能够形成sym2 plasmata结构(biofilm的一种,菌体生长到一定阶段菌体聚集形成的一种簇凝块状共质体结构),保护细菌细胞免受波动环境条件的影响[43—44].研究发现,许多周质空间蛋白对biofilm的形成而有显著的影响,从而间接的影响了其抗逆作用.Karatan等对biofilm的形成和抑制机制进行的研究发现,一种多胺类周质空间受体蛋白Nsp S,与多胺(如norspermidine)结合形成复合体后影响MbaA蛋白处于周质空间的结构域,而MbaA霍乱弧菌(V ibrio cholerae)的biofilm形成的抑制因子,因此Nsp S对biofilm的形成起间接激活作用,形成后的biofilm保护菌体细胞适应不良的外界环境条件[45].另一大肠杆菌周质空间蛋白SurA属于肽酰脯氨酸异构酶家族,它是一种双功能蛋白,一方面催化肽酰脯氨酸残基的顺式—反式异构,另一方面能影响biofilm的形成.surA基因被敲除后,突变体菌株外膜蛋白OmpA和LamB的合成会剧烈减少,而这两个蛋白是菌体biofilm的形成所必需的,所以周质空间蛋白SurA能促进菌体biofilm的形成,而间接促进抗逆作用[46].不仅如此,还有研究表明周质空间蛋白SurA能参与其他某些外膜蛋白和/或分泌蛋白的成熟,从而促进菌体biofilm的形成,而影响菌体细胞的抗逆作用[47,48].由此可见周质空间蛋白在菌体细胞biofilm的形成/维持起非常重要的作用[45—50].另一方面,biofilm的形成反过来也会影响许多周质空间蛋白的表达,从而进一步调节其抗逆作用.Pysz发现T hermotoga m ariti m a的biofilm在形成前后周质空间蛋白的表达有较明显的差异,如周质空间内的麦芽糖结合蛋白的表达量是形成前的217倍,丝氨酸蛋白酶为原来211倍[51].这样以来,周质空间蛋白可以通过影响biofilm形成而影响菌体的抗逆作用,而biofilm的形成反过来也会影响周质空间蛋白的表达,菌体细胞在这种协同调节中实现对环境的更好适应14 结束语G-细菌是重要的微生物类群,它们在农业生产、工业应用、医学病原分析与治疗、以及环境监测等各方面都具有非常重要的地位,G-细菌抗逆机制的研究对于阐释它们在上述领域研究中的相关问题意义重大.周质空间处于G-细菌与外界相互作用的最前沿,对于生存环境的波动非常敏感.本文简要综述了G-细菌细胞周质空间蛋白的分类、定位和抗逆机制,着重介绍了目前研究较多周质空间蛋白在抗酸机理中和RND转运系统在菌体抗逆生存中所起的重要作用等.从中可以看出,关于细菌周质空间蛋白的研究主要集中于这些蛋白在菌体抗逆生存的生理功能确认方面,但对具体发挥抗逆作用的分子生物学机制方面研究不多,比如已知某蛋白具有抗酸活性(生理功能),但问题是它是如何抗酸(分子机制)的却研究得不够清楚.另一方面,值得注意的是,许多周质空间蛋白(如SurA,Mal E等)都具有双功能特性的,它们本身是一些酶蛋白,然而又具有显著的抗逆作用,它们在结构与功能的关系上是如何协调这些双功能特性的,这些问题都有待于进一步深入研究.448 第18卷 第8期 2008年8月因此,作为菌体细胞与外界环境进行相互作用的第一道媒介(趋利作用)或屏障(弊害作用),周质空间蛋白及其抗逆作用的相关研究无疑将为探究与理解微生物之间的相互作用及生存适应性乃至微生物进化等重要问题提供依据,具有广阔的研究前景.参 考 文 献1 Bladen HA,Waters J F.Electron microscopic study of some strains of B acteri des.J Bacteriol,1963,86:1339—13442 Stock JB,Rauch B,Roseman S.Periplasmic space in S al monella t y phi muri um and Escherichia coli.J Biol Chem,1977, 252(21):7850—78613 Decad GM,NIkaido H.Outer membrane of Gram2negative bac2 teria.XII.Molecular2sieving function of cell wall.J Bacteriol, 1976,128(1):325—3364 Schulz GE.Porins:General to specific,native to engineered pas2 sive pores.Curr Opin St ruc Biol,1996,6:485—4905 Terry J.Beveridge structures of gram2negative cell walls and t heir derived membrane vesicles.J Bacteriol,1999,181:4725—47336 Adams MD,Dale L Oxender.Bacterial periplasmic binding pro2 tein tertiary structures.J Biol Chem,1989,264(27):15739—157427 Richarme G,Caldas TD.Chaperone properties of t he bacterial periplasmic substrate2binding proteins.J Biol Chem,1997, 272(25):15607—156128 Blattner FR,Plunkett G,Bloch CA,et al.The complete genome sequence of Escherichia coli K212.Science,1997,277: 1453—14629 #sgeirsson B,Adalbjornsson BV,Gylfason GA.Engineered di2 sulfide bonds increase active2site local stability and reduce cata2 lytic activity of a cold2adapted alkaline phosphatase.Biochim Bio2 phys Acta,2007,1774:679—68710 Beck BD,Park J T.Basis for t he observed fluctuation of car2 boxypeptidase II activity during t he cell cycle in BU G6,a tem2 perature2sensitive division mutant of t he Escherichia coli.J Bac2 teriol,1977,130(3):1292—130211 Ureta AR,Endres R G,Wingreen NS,et al.Silhavy.K inetic analysis of t he assembly of t he outer membrane protein LamB in Escherichia coli mutant s each lacking a secretion or targeting fac2 tor in a different cellular compart ment.J Bacteriol,2007,189: 446—45412 Boos W,Shuman H.Maltose/Maltodextrin system of Esche2 richia coli:Transport,metabolism,and regulation,microbiol.Mol Bio Rev,1998,62:204—22913 Wassenberg D,Lieb W,J aenicke R.Maltose2binding proteinfrom t he hypert hermophilic bacterium T hermotoga mariti ma: Stability and binding properties.J Mol Biolog,2000,295(2): 279—28814 Missiakas D,Raina S.Protein folding in t he bacterial periplasm.J Bacteriol,1997,179:2465—247115 Sch fer U,Beck K,Müller M.Skp,A molecular chaperone of Gram2negative bacteria,is required for t he formation of soluble periplasmic intermediates of outer membrane proteins.J Biol Chem,1999,274(35):24567—2457416 Behrens S.Periplasmic chaperones2preservers of subunit folding energy for organelle assembly.Cell,2003:556—55717 Eda S,Maseda H,Nakae T.An elegant means of self2protection in Gram2negative bacteria by recognizing and extruding xenobiot2 ics from t he periplasmic space.J Biol Chem,2003,278(4): 2085—208818 Sarter S,Nguyen HN,Hung L T,et al.Antibiotic resistance in Gram2negative bacteria isolated from farmed catfish.Food Con2 trol,2007,18:1391—139619 Liu Y,Fu XM,Shen J,et al.Periplasmic proteins of Escherich2 ia coli are highly resistant to aggregation:Reappraisal for roles of molecular chaperones in periplasm.Biochem and Bioph Res Co, 2004,316:795—80120 Liu J,Tang M,Mestril R,et al.Aberrant protein aggregation is essential for a mutant desmin to impair t he proteolytic function of t he ubiquitin2proteasome system in cardiolyocytes.J Mol Cell Cardiol,2006,40:451—45421 Pogliano J,Lynch AS,Belin D,et al.Regulation of Escherichia coli cell envelope proteins involved in protein folding and degra2 dation by t he Cpx two2compont system.Genes Dev,1997,11: 1169—118222 Hong WZ,Jiao WW,Hu J C,et al.Periplasmic protein HdeA exhibit s chaperone2like activity exclusively wit hin stomach p H range by transforming into disordered conformation.J Biol Chem,2005,280(29):27029—2703423 Miyadai H,K imie TM,Mat suyama SI,et al.Effect s of lipopro2 tein overproduction on t he induction of DegP(HtrA)invovlved in quality control in t he Escherichia coli periplasm.J Biol Chem, 2004,279(38):39807—3981324 Raivio TL.Periplasmic stress and ECF sigma factors.Annu Rev Microbiol,2001,55:591—62425 Raivio TL.Envelope stress responses and Gram2negative bacteri2 al pat hogenesis.Mol Microbiol,2005,56:1119—112826 Duguay AR,Silhavy TJ.Quality control in t he bacterial periplasm.Biochim Biophys Acta,2004,1694:121—13427 Y oung J C,Ulrich HF.A stress sensor for t he bacterial periplasm.Cell,2003,113:1—228 Richard H,Foster J W.Escherichia coli glutamate2and arginine2 dependent acid resistance systems increase internal p H and548 第18卷 第8期 2008年8月reverse transmembrane potential.J Bacteriol2004,186:6032—604129 Mates A K,Sayed A K,Foster J W.Product s of t he Escherichia coli acid fitness island attenuate metabolite stress at extremely low p H and mediate a cell density2dependent acid resistance.J Bacteriol,2007,189:2759—276830 Gajiwala KS,Burley SK.HDEA,a periplasmic protein t hat sup2 port s acid resistance in pat hogenic enteric bacteria.J Mol Biol, 2000,295:605—61231 Tikhonova EB,Wang Q,Zgurskaya H I.Chimeric analysis of t he multicomponent multidrug efflux transporters from Gram2 negative bacteria.J Bacteriol,2002,184:6499—650732 Lee SM,Grass G,Rensing C,et al.The Pco proteins are in2 volved in periplasmic copper handling in Escherichi a coli.Bio2 chem and Bioph Res Co,2002,295:616—62033 Perron K,Caille O,Rossier C,et al.CzcR2CzcS,a two2compo2 nent system involved in heavy metal and carbapenem resistance in Pseu domonas aeru ginosa.J Biol Chem,2004,279(10):8761—876834 Reisner A,H ller BM,Molin S,et al.Synergistic effect s in mixed Escherichia coli bioflims:Conjugative plasmid transfer drives biofilm expansion.J Bacteriol,2006,188:3582—3588 35 Zambrano MM,K olter R.Mycobacterial biofilms:A greasy way to hold it toget her.Cell,2005,123:762—76436 Zhang XS,Rodolfo GC,Wood T K.Ycf R(BhsA)Influences Escherichia coli biofilm formation t hrough stress response and surface hydrophobicity.J Bacteriol,2007,189(8):3051—3062 37 冯永君,何 晴.植物内生菌的生物薄膜(biofilm).生命的化学,2007,27(1):87—8838 冯永君,宋 未.植物内生菌.自然杂志,2001,23(5):249—25239 卢镇岳,杨新芳,冯永君.植物内生细菌的分离、分类、定殖与应用.生命科学,2006,18:90—9440 冯永君,宋 未.水稻内生优势成团泛菌GFP标记菌株的性质与标记丢失动力学.中国生化与分子生物学报,2002,18(1): 85—9141 沈德龙,冯永君,宋 未.内生成团泛菌YS19对水稻乳熟期光合产物在旗叶、穗分配中的影响.自然科学进展,2002,12(8): 863—86542 Feng Y J,Shen DL,Song W.Rice endophyte Pantoea ag glomer2 ans YS19promotes host plant growt h and affect s allocations of host photosynt hates.J Appl Microbiol,2006,100:938—945 43 Feng Y J,Shen DL,Dong XZ,et al.I n vit ro symplasmata for2 mation in t he rice diazotrophic endophyte Pantoea ag glomerans YS19.Plant Soil,2003,255:435—44444 Duan J Y,Y i T,Feng Y J,et al.Rice endophyte pantoea ag glo2 merans YS19forms multicellular symplasmata via cell aggrega2 tion.FEMS Microbiol Lett,2007,270:220—22645 Karatan E,Duncan TR,Watnick P I.NspS,a predicted poly2 amine sensor,mediates activation of V ibrio cholerae biofilm for2 mation by norspermidine.J Bacteriol,2005,187(21):7434—744346 Lazar SW,K olter R.SurA assist s t he folding of Escherichia coli outer membrane proteins.J Bacteriol,1996.178(6):1770—177347 Hunstad DA,J ustice SS,Hung CS,et al.Suppression of blad2 der epit helial cytokine responses by uropat hogenic Escherichia coli.Infect Immun,2005,73:3999—400648 J ustice SS,Lauer SR,Hultgren S J,et al.Maturation of intra2 cellular Escherichia coli communities requires SurA.Infect Im2 mun,2006,74(8):4793—480049 Schembri MA,Kjaergaard K,K lemm P.G lobal gene expression in Escherichia coli biofilms.Mol Microbiol,2003,48(1):253—26750 Beloin C,Valle J,Patricia LL,et al.G lobal impact of mature biofilm lifestyle on Escherichia coli K212gene expression,Mol Microbiol,2004,51(3):659—67451 Pysz MA,Conners SB,Montero CI,et al.Transcriptional anal2 ysis of biofilm formation processes in t he anaerobic,hypert her2 mophilic bacterium T hermotoga mariti ma.Appl Environ Mi2 crob,2004,70(10):6098—6112648 第18卷 第8期 2008年8月。
生物学科名词解释

生物学科类名词解释A1.乙酸营养型(Acetotrophic):能利用、消耗乙酸的营养型。
2.乙酰辅酶A途径(Acetyl-CoA pathway):一种自养营养型固定CO2的途径,广泛存在于专性厌氧的产甲烷细菌、同型乙酸和硫酸盐还原细菌中。
3.酸性矿排出的水(Acid mine drainage):微生物氧化硫铁矿产生的含有H2SO4的酸性水。
4.嗜酸菌(Acidophile):生物体能在较低pH下生长得最好。
5.活化能(Activation energy):使底物达到反应状态所需的能量。
6.激活蛋白(Activator protein):一种调控蛋白,在正调控中,它与DNA上专一位点结合,激活转录。
7.急性(Acute):短期感染,通常是以剧烈发作为特征。
8.好氧菌(Aerobe):生物体在呼吸中利用O2,一些生长时需要O2。
9.耐氧厌氧菌(Aerotolerant):不能呼吸氧气但是有氧气存在时也不会影响生长的微生物。
10.藻类(Algae):光合营养型真核微生物。
11.嗜碱菌(Alkliphile):能在高pH下生长得最好的生物体。
12.变构酶(Allosteric enzyme):具有两个结合位点的酶。
这两个位点分别为活性位点(底物结合位点)和变构位点(效应物结合位点)。
13.阿米巴样运动(Amoeboid movement):生物体以细胞质流动进行移动的一种运动方式。
14.氨酰-tRNA合成酶(Aminoacyl-tRNA synthetase):一组酶,每一种酶能催化一种正确的氨基酸与tRNA连接。
15.氨基糖苷类抗生素(Aminoglcoside):同链霉素相同的一类抗生素,含有糖苷键连接的氨基糖基团。
16.合成代谢(Anabolism):细胞内所进行的全部生物合成反应。
17.厌氧菌(Anaerobe):生物体在呼吸中不需O2,它们的生长可以被O2抑制。
18.厌氧呼吸(Anaerobic respiration):由SO42-或NO3-等物质代替O2作为最终电子受体的呼吸作用。
细菌生物被膜

细菌⽣物被膜细菌⽣物被膜(或称细菌⽣物膜Bacterial biofilm,BF),根据《Annu Rev Microbiol》等权威期刊所归纳发表的定义,⽣物薄膜是指细菌粘附于接触表⾯,分泌多糖基质、纤维蛋⽩、脂质蛋⽩等,将其⾃⾝包绕其中⽽形成的⼤量细菌聚集膜样物。
多糖基质通常是指多糖蛋⽩复合物,也包括由周边沉淀的有机物和⽆⽣物被膜是微⽣物为适应⾃然环境⽽形成的。
例如:河流中的微⽣物就能吸附在岩⽯表⾯,这种吸附作⽤更有利于⾃⾝的⽣存。
对于细菌来说,它可以分泌多糖蛋⽩复合物(glycocalyx)将⾃⾝粘附于各种物体的表⾯,细菌在所吸附的物体表⾯不断分裂就形成了细菌⽣物被膜。
使病原菌可以在体内插管表⾯及粘膜表⾯形成⽣物被膜[1]。
三、细菌⽣物被膜的致病特点⼀般来说细菌⽣物被膜导致的难治性细菌感染性疾病有以下特点:1.病灶局部的炎症反应不很强烈,感染有相互转化的静⽌期和发作期;2.抗菌药物治疗起初可能有效,但以后治疗常常失败;3.致病菌主要是来⾃⽪肤和周围环境中的致病菌如铜绿假单胞菌,⾦黄⾊葡萄球菌[1]。
铜绿假单胞菌是慢性呼吸道感染的重要致病菌之⼀,它的粘液型菌株可以产⽣藻酸盐,⽽⾮粘液型菌株可以产⽣其它种类的多糖蛋⽩复合物形成细菌⽣物被膜。
⽐较典型的病例是肺囊性纤维化合并肺部感染,虽然抗菌药物有⼀定的临床疗效,但是铜绿假单胞菌总是难以彻底清除,电镜观察可见病变部位有细菌⽣物被膜形成。
其中藻酸盐是重要的组成成分,它可以使细菌牢固地粘附于肺上⽪表⾯形成⽣物被膜,⼀⽅⾯可以抵御单核-巨噬细胞的吞噬作⽤,另⼀⽅⾯可以抵制抗菌药物的杀灭作⽤。
进⼀步的研究表明,铜绿假单胞菌藻酸盐的合成是由细菌alC和alD基因控制的。
有实验表明,铜绿假单胞菌和硅胶膜表⾯接触后,可以激活控制藻酸盐合成的基因组,促使细菌合成⼤量的藻酸盐。
所以细菌⽣物被膜的形成是受严密的基因调控的[12]。
1、⽣物膜的研究历史:1676 年Antony⽤⾃制的显微镜从⽛菌斑中观察到了微⽣物的存在,为⽣物膜的研究奠定了基础。
细菌生物被膜检测与清除方法研究进展

食品研究与开发F ood Research And Development圆园20年10月第41卷第20期DOI :10.12161/j.issn.1005-6521.2020.20.036基金项目:国家自然科学基金项目(31972170)作者简介:张君怡(1994—),女(汉),硕士研究生,研究方向:食品安全。
*通信作者:郝建雄(1979—),男,教授,博士,研究方向:食品安全与控制。
细菌生物被膜检测与清除方法研究进展张君怡,王静怡,巴巨伟,崔雪岩,崔佳琪,郝建雄*(河北科技大学生物科学与工程学院,河北石家庄050000)摘要:开展食源性致病菌的防控研究以保障食品安全,具有显著的实践意义。
作为细菌为适应不良环境所表现出的一种自我保护机制,食源性致病菌在食品生产加工过程中,可以黏附在食品原材料表面以及各种食品加工器械表面形成生物被膜,增大了消毒清洗的难度,给食品安全带来了严重的隐患和危害。
因此,如何有效地清除细菌生物被膜成为食品安全领域的研究热点。
结合细菌生物被膜形成过程和调控机制,本研究系统分析总结了细菌生物被膜的检测与清除方法的现状,为实现细菌生物被膜的有效控制,消除由生物被膜造成的食品安全隐患提供了综合性的认识,可为研究人员解决细菌生物被膜带来的食源性微生物污染提供参考,对保障食品安全有重要意义。
关键词:细菌生物被膜;检测方法;清除;微生物污染;食品安全Development on Detection and Removal of Bacterial BiofilmZHANG Jun-yi ,WANG Jing-yi ,BA Ju-wei ,CUI Xue-yan ,CUI Jia-qi ,HAO Jian-xiong *(College of Life Science and Engineering ,Hebei University of Science and Technology ,Shijiazhuang 050000,Hebei ,China )Abstract :It is of great practical significance to carry out research on prevention and control of foodbornepathogenic bacteria to ensure food safety.As a self -protection mechanism for bacteria to adapt to the bad environment ,food borne pathogenic bacteria can adhere to the surface of food raw materials and various food processing equipment in the process of food production and processing to form biofilm ,which increases the difficulty of disinfection and cleaning ,and brings serious hidden danger and harm to food safety.Therefore ,how to effectively remove bacterial biofilm has become a research hotspot in the field of food bined with the formation process and regulation mechanism of bacterial biofilm ,this study systematically analyzed andsummarized the current situation of detection and removal methods of bacterial biofilm ,which provided a comprehensive recognition for the effective control of bacterial biofilm and elimination of food safety risks caused by biofilm.This study could provide a reference for researchers to solve the food borne microbial pollution caused by bacterial biofilm ,which was of great significance to ensure food safety.Key words :bacterial biofilm ;detection method ;removal ;microbial contamination ;food safety引文格式:张君怡,王静怡,巴巨伟,等.细菌生物被膜检测与清除方法研究进展[J].食品研究与开发,2020,41(20):218-224ZHANG Junyi ,WANG Jingyi ,BA Juwei ,et al.Development on Detection and Removal of Bacterial Biofilm[J].Food Research and Development ,2020,41(20):218-2241978年,英国学者J.Willam Costerton 首次提出生物被膜的概念。
细菌生物被膜在生物防治中的作用

细菌生物被膜在生物防治中的作用凡肖;束长龙;关雄;黄天培【期刊名称】《中国生物防治学报》【年(卷),期】2018(034)005【摘要】细菌生物被膜(bacterial biofilm,BBF)是在固—液和气—液表面上生长并封闭在胞外多糖基质中的细胞群,能调控细菌对环境胁迫的适应能力,提高细菌对紫外线(UV)胁迫等环境的抗逆性,是细菌的一种保护性生长方式.生防菌剂(biological control agents,BCAs)因其能以环境友好的方式长期控制病害虫等优点,一直是生物农药领域研究的热点.本文简介了细菌生物被膜及其应用,以苏云金芽胞杆菌(Bacillus thuringiensis,Bt)和枯草芽胞杆菌(Bacillus subtilis,Bs)为核心综述了BBF对生防活性的影响、生防细菌生物被膜形成、解离的表达、调控机制以及生物被膜调控基因的鉴定方法,并指出未来BBF在生防应用方面主要的研究方向,旨在为解决BCAs田间持效期较短等问题提供新的思路.【总页数】11页(P791-801)【作者】凡肖;束长龙;关雄;黄天培【作者单位】福建农林大学生命科学学院/植物保护学院闽台作物有害生物生态防控国家重点实验室/生物农药与化学生物学教育部重点实验室,福州350002;中国农业科学院植物保护研究所/植物病虫害生物学国家重点实验室,北京100193;福建农林大学生命科学学院/植物保护学院闽台作物有害生物生态防控国家重点实验室/生物农药与化学生物学教育部重点实验室,福州350002;福建农林大学生命科学学院/植物保护学院闽台作物有害生物生态防控国家重点实验室/生物农药与化学生物学教育部重点实验室,福州350002【正文语种】中文【中图分类】S476.1【相关文献】1.细菌生物被膜在不可吸收内植物感染中的作用 [J], 刘航涛;王万明2.细菌生物被膜在生物防治中的作用 [J], 凡肖; 束长龙; 关雄; 黄天培3.林业有害生物防治在林业生态环境建设中的作用 [J], 李伦兵4.有害生物防治在农林业中的作用分析 [J], 刘冰;田妮娟5.探讨林业有害生物防治在林业生态环境建设中的作用 [J], 李玲因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
9DE )$ 在 甘 蔗 叶 细 胞 间 也 被 包 埋 于 一 种 糖 蛋 白 形 成的纤维状基质中 4#5 & 89C<D>F 等 4G5 报道的成团肠 杆 菌 (’H;:?<=>9;:? >IIJ<K:?>HE )$ 在 水 稻 根 面 定 殖
过程中形成的菌体细胞簇连而成的凝块状结构$ 有对宿主水稻的定 殖 专 一 性 ’ 检 测 表 明 $ 在 该 结 构表面形成胞外多糖 和 蛋 白 质 的 包 被 & 我 们 从 水 稻分离的固氮成团 泛 菌 (L>H;<:> >IIJ<K:?>HE ) 是 另 一重要内生细菌 4M1$N5 $ 其中对其聚集成团的共质体 结构进行的研究表明 $ 该 结 构 的 表 面 基 质 成 分 主 要 为 蛋 白 质 $ 并 分 离 得 到 了 一 种 !$O! FP 胞 外 基 质蛋白 &
#" 生物薄膜结构与功能 生物薄膜的形成会对细菌生态学和生物学作
用产生重要影响 ’ 生 物 薄 膜 有 助 于 菌 体 细 胞 抵 抗 干燥( 紫外线( 被捕食和杀菌剂等的胁迫作用* 对遗传信息交换和菌体协同作用等也有重大意义 * 某些对于单个菌体来 说 难 以 代 谢 的 化 合 物 可 以 被 生物薄膜利用 * 生 物 薄 膜 还 可 能 引 起 某 些 细 菌 密 度依赖表现型的表达 X!1516Y ’ 比如 % 对某些有密度依 赖行为的细菌基因 的 表 达 % 可 以 通 过 群 体 感 应 系 统 $TP=ZP@ [HI[<IJ & 的可扩散分子如 酰 基 高 丝 氨 酸 内酯来诱导 % 生物 薄 膜 基 质 有 助 于 这 类 分 子 的 积 累 ’ 上述这些功 能 的 获 得 % 很 多 直 接 得 益 于 生 物 薄膜的理化结构 % 特 别 是 生 物 薄 膜 所 形 成 的 屏 障 作用 ’ 再比如 % 由 于 生 物 薄 膜 中 的 细 菌 彼 此 很 接 近 % 且结构内的 基 质 有 某 些 保 护 作 用 % 可 以 促 进 生物薄膜中进行的细菌之间遗传信息的交换 X!Y’ 那么% 生物薄膜的存在会成为水分和营养物
!" 生物薄膜的概念 为了准确说明生物薄膜的概念% 我们有必要
引 用 !]IIP ,H^ (<QZ=;<=? " 等 重 要 期 刊 所 归 纳 发 表 的定义 % 生物薄膜是 指 微 生 物 菌 体 互 相 黏 附 在 一 起或附着到一些材料 表 面 上 % 并 由 胞 外 多 聚 物 基 质对其包埋的生长状 态 * 一 个 生 物 薄 膜 就 意 味 着 是一个微小生境% 该生境与周围环境可能有区 别 % 生物薄膜内的 菌 体 细 胞 密 切 协 调 % 而 生 物 薄 膜 体 系 外 的 菌 体 则 不 具 有 这 样 的 相 互 作 用 X!Y ’ 人 们最初观测到细菌与植 物 相 互 作 用 形 成 生 物 薄 膜 可 追 溯 到 !9 世 纪 69 ( #9 年 代 * 但 长 期 以 来 % 相 关研究主要集中在对 其 形 态 的 观 察 与 描 述 ’ 直 到
!! !! !! !! !! !
收 稿 日 期 # !996C44C"B 国 家 自 然 科 学 基 金 资 助 项 目 7D=E 59F""""!3 作者简介# 冯 永 君 74B#5 ) 3 % 男 % 博 士 % 副 教 授 % 联 系 作 何 晴 2 $B:5 ) 8 % 女 % 硕 士 生 % 者 % ’C@G<? # >HIJKLM;<NEHOPEQI *
4!5
种 特 殊 的 QAJK $ 但 是 在 微 生 物 和 生 物 化 学 领 域 $ =A<QAJK 绝 对 不 是 =A<K:K=?>H: & 因 此 $ 将 =A<QAJK 译为 * 生物膜 + 不 但 从 本 质 上 曲 解 了 它 本 身 的 原 意 $ 而 且 会 干 扰 生 物 膜 原 本 概 念 ( 也 即 =A<K:K !
有氧条件下 $ 在生 物 薄 膜 内 可 以 形 成 缺 氧 区 $ 有 利于固氮作用 $ 因为 固 氮 酶 发 挥 催 化 活 性 需 要 低 氧 环 境 ’ 在 中 性 6& 下 生 长 的 菌 体 细 胞 $ 在 生 物 薄膜内可以形成局部 酸 性 区 $ 有 利 于 代 谢 某 些 化 合物 ’ 在生物薄膜表 面 形 成 阻 止 分 子 量 较 大 的 化 合物扩散的区域 $ 或 表 现 出 对 特 定 化 合 物 选 择 性 吸附作用 $ 有利于 趋 利 避 害 & 在 这 种 对 各 类 物 质 的传质限制和选择的 平 衡 状 态 下 $ 生 物 薄 膜 内 的 菌体细胞表现出独特的环境生存适应性 & 生物薄膜的胞外多聚物基质主要成分是多糖 $ 和7或 蛋 白 质 $ 和7或 糖 蛋 白 & 曾 有 报 道 $ 甘 蔗 内 生 固 氮 菌 重 氮 营 养 醋 杆 菌 (89:;<=>9;:? @A>B<;?<6CA!
=?>H: ) 的正常使用 &
事实上$ 随着近年来人们对植物微生物相互 作用研究的日益重视 $ 该 领 域 的 研 究 已 经 迅 速 发 展起来 $ 我们发现对于 =A<QAJK 不规范甚至不正确 的翻译或提法也广泛 存 在 & 这 种 不 当 的 翻 译 方 法 主要表现为两个方面 $ 一 是 国 内 有 些 期 刊 在 发 表
植物微生态环境的波 动 会 产 生 各 种 生 存 压 力 % 菌 体的这种聚集行为可 能 是 一 种 重 要 适 应 策 略 % 其 所表现的生物学功能往 往 是 未 聚 集 的 单 个 细 胞 所 不具有的 ’
X5Y
?P[ [P;N<?<[ &等都可以形成生物薄膜结构 % 以适应特
定生存 环 境 或 生 长 阶 段 的 需 要 XF1‘Y ’ 这 些 研 究 大 大 拓宽了有关生物薄膜结构与功能的研究内容与 价值’ 但本文主要从植物内生细菌与植物相互 作用的角度% 探讨植物<IJUVPW=IJM;<NEHOPEQI
万方数据
! GG !
! 生命的化学 "!""# 年 !# 卷 $ 期
%&’()*+,- ./ 0)/’ !""#1 !#2$3
!!"#$%&’ ()*+,-
质的传质障碍吗 # 研 究 发 现 $ 尽 管 生 物 薄 膜 的 包 埋基质多为亲水性物 质 $ 但 相 对 于 在 水 中 的 自 由 扩散来说 $ 氧气 % 营 养 成 分 以 及 其 他 物 质 的 扩 散 还是受到一定的限制 & 但是由于这种限制$ 在
许多细菌能定殖在植物的根( 茎( 叶表面或 组织内部 % 产生植物 微 生 物 相 互 作 用 的 适 应 性 体 系 ’ 尽管细菌是单细胞微生物% 但它们在植物
X4Y
最近 % 该种结构才 被 发 现 与 某 些 生 物 学 功 能 或 属 性有关 % 进而成为 植 物 微 生 物 相 互 作 用 研 究 的 热 点% 生 物 薄 膜 在 植 物C细 菌 相 互 作 用 中 所 扮 演 的 角色也越来越受关注 ’ 近来的研究又进一步发现% 许多普通细菌如 大肠杆菌 $’[QSHZ<QS<G Q=?< & 和 枯 草 芽 孢 杆 菌 $_GQ<?"
!" 关于生物薄膜 !#$%&$’(" 与生物膜 !#$%()(#*+,)" 关于 =A<QAJK 的中文译名 $ 科学出版社出版 的 ! 英汉生物学词汇 "( 第二版 ) 给出的 翻 译 是 * 生 物
膜 +$ 但 容 易 产 生 误 解 & 众 所 周 知 $ 中 文 的 生 物 膜概念实际上早有所 指 $ 也 已 经 为 广 大 从 事 生 命 科学与技术研究的工 作 者 所 接 受 $ 生 物 膜 原 本 指 的是细胞的外周膜 ( 质膜 )% 核 膜 和 各 种 细 胞 器 膜 的总称 $ 各生物膜 的 骨 架 是 一 样 的 $ 主 要 是 由 磷 脂类构成的双分子层 ( 脂双层 )$ 这 个 名 称 来 源 于 英文词 =A<K:K=?>H: & 在生命科学的领域内 $ 生物 膜 % 膜和膜系统等概 念 往 往 都 是 指 这 种 磷 脂 双 分 子层结构 ’ 虽然 $ 有 些 膜 结 构 可 能 镶 嵌 有 蛋 白 质 和 7 或糖蛋白等 $ 但其骨架结构依然是磷脂双分子 层 & 然 而 $ 与 之 大 相 径 庭 $ =A<QAJK 却 主 要 是 由 多 糖 $ 和7或 蛋 白 质 $ 和7或 糖 蛋 白 等 多 聚 物 的 黏 稠 基质组成的 $ 未必是 ( 或者可 能 完 全 不 是 ) 磷 脂 双 分 子 层 结 构 & 虽 然 可 以 认 为 =A<K:K=?>H: 属 于 一
!!"#$
文 章 编 号 # 4"""! 45567!""#894!99:#! 95
! 生命的化学 "!""# 年 !# 卷 $ 期
%&’()*+,- ./ 0)/’ !""#1 !#2$3
" !" "
植物内生细菌的生物薄膜 !!"#$"%& "
冯永君 何 晴
$ 北 京 理 工 大 学 生 命 科 学 与 技 术 学 院 % 北 京 $999:$ &
=A<QAJK 相 关 研 究 论 文 以 及 综 述 时 经 常 将 其 字 面 化
为生物膜$ 这其中包括一些学报级的重要刊物& 另一方面 $ 有很多硕 7 博士研究生在向本文第一作 者请教或交流植物微 生 物 相 互 作 用 的 问 题 时 $ 经 常提到 * 内生菌 的 生 物 膜 + 如 何 如 何 $ 常 常 使 人 被搞糊涂 ’ 直到再 问 及 他 们 英 文 表 述 后 $ 才 明 白 原 来 他 们 心 里 指 的 是 * :H@<6CR;A9 =A<QAJK + $ 而 不 是 他 们 口 中 说 的 * =A<K:K=?>H: <Q >H :H@<6CR;: + & 造 成 这 种 不 规 范 甚 至 不 正 确 的 翻 译 或 使 用 =A<QAJK 概念的原因 $ 很可 能 是 某 些 中 文 期 刊 中 * 以 讹 传 讹 + 造成误导 $ 也 可 能 是 部 分 使 用 者 本 人 没 有 严 格对待 & 由此可见 $ 规范 =A<QAJK 的中文名称已经 十分必要 & 根 据 上 面 所 引 用 的 !8HHD ,:S LCR;<6>;C<J " 和 !8HHD ,:S (A9?<=A<J " 等期刊对 =A<QAJK 的归 纳 $ 以 及我们对 =A<QAJK 结构形成 % 生理生化性质 % 以及 生物学功能研究的回 顾 和 总 结 $ 结 合 我 们 对 细 菌