神经营养素信号通路
神经生长因子

神经生长因子(NGF)成都医学院 2013级临床七班马成杰 1325100381摘要:神经生长因子(NGF)是神经营养因子中最早被发现,目前研究最为透彻的,具有神经元营养和促突起生长双重生物学功能的一种神经细胞生长调节因子,它对中枢及周围神经元的发育、分化、生长、再生和功能特性的表达均具有重要的调控作用。
NGF包含α、β、γ三个亚单位,活性区是β亚单位,由两个118个氨基酸组成的单链通过非共价键结合而成的二聚体,与人体NGF的结构具有高度的同源性,生物效应也无明显的种间特异性。
关键词:神经生长因子(NGF)分子组成生物效应1951年,Levi-M on talc in i和Hamburger首先在培养的小鼠肉瘤细胞内发现NGF存在。
从发现第一个多肽生长因子NGF以来,又相继发现了脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、神经营养素-3(neurotrophic-3,NT-3)、神经营养素-4(neurotrophic-4,NT-4)、神经营养素-5(neurotrophic-5,NT-5),由于它们起源于同一基因家族,故可称为NGF 家族。
此外,还有睫状神经营养因子(c i l i a r y neurotrophic factor,CNTF),胶质细胞源性神经营养因子(g l i a l-derived neurotrophic factor,GDNF),但它们在分子结构和受体类型上与NGF家族不同。
经典的NGF是从小鼠颌下腺中分离所得的分子量为140,000的糖蛋白,由α、β、γ三种肽链构成,肽链间以共价键结合。
其中β亚单位是NGF的活性区。
经过40多年的研究,NGF的蛋白结构已基本了解清楚,三维结构已从其晶体图像证实。
它由两个相同的亚单位组成,亚单位由三个β转角连接的三对反平行β折叠组成。
亚单位间通过“半胱氨酸结点”连接。
目前发现的NGF均为糖蛋白,其构型至关重要的氨基酸在神经营养素家族中保守,不同于NGF的特性由可变区的氨基酸决定,可变区的变化不影响它们的基本结构,但对特异性受体的结合起决定作用。
【国家自然科学基金】_神经营养素_基金支持热词逐年推荐_【万方软件创新助手】_20140801
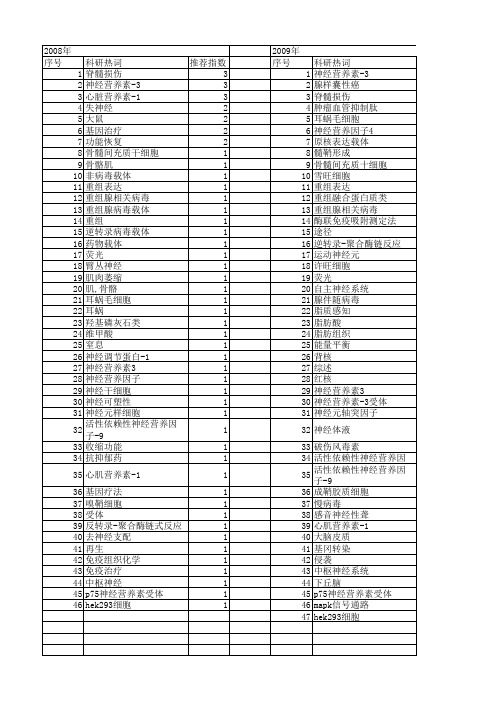
推荐指数 4 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
科研热词 阿尔茨海默病 运动 血管生成 营养素 脑源性神经营养因子 肿瘤 线粒体 神经突起因子 神经干细胞 神经可塑性 树突状细胞 慢病毒载体 存活 基因转染 发育行为儿科学 儿童 信号通路 人神经营养素3 trkc nt-3
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34
2011年 科研热词 神经营养素-3 脊髓损伤 神经纤维再生 基因治疗 骨髓间充质干细胞 额叶皮质 雪旺细胞 重组腺病毒 运动神经元 过度训练 轴突再生 谷氨酰胺 视觉系统 腺病毒 脊髓 移植 神经营养素受体 神经营养素 神经营养因子 神经干细胞 神经再生 神经元 相关因子 相互作用 死亡受体 明胶海绵圆柱体支架 大鼠 坐骨神经 可塑性 信号转导 作用机制 亲和性 trkc p-creb 推荐指数 3 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8
2014年 科研热词 阿尔茨海默病 酪氨酸激酶受体c 记忆 神经营养素-3 推拿 寿命 坐骨神经损伤 卡路里摄入限制 推荐指数 1 1 1 1 1 1 1 1
Slit2在膀胱癌中的表达、信号通路及其与患者预后关系的生物信息分析

2021,40(1)河南大学学报(医学版)㊃19㊀㊃文章编号:1672-7606(2021)01-0019-06Slit2在膀胱癌中的表达㊁信号通路及其与患者预后关系的生物信息分析李扬,王云,李遵,朱朝阳∗河南大学淮河医院泌尿外科,河南开封475000摘㊀要: 目的 探讨Slit2在膀胱癌中的表达及其与患者预后关系㊂ 方法 采用生物信息学方法,在GTEx数据库中检索Slit2基因在人体各种组织中的表达水平,在TCGA数据库中分析Slit2多种肿瘤组织及在膀胱癌中的表达㊂采用在线分析工具cBioPortal,分析Slit2基因突变情况㊂根据Slit2表达水平,分为高㊁低表达组,比较两组患者无疾病进展生存(DFS)和总生存(OS)是否存在差异㊂ 结果 Slit2基因mRNA在膀胱癌患者癌组织中的表达水平明显高于癌旁正常膀胱组织(P<0.05),且Slit2表达水平与患者临床分期有关,分期越高表达水平越高(P<0.05)㊂Slit2基因主要突变形式为融合突变㊁扩增突变㊁深度缺失突变和多重突变等㊂其中缺失突变占25.55%,同义突变占8.19%,无意义突变占2.01%㊂单核苷酸突变主要类型为C>T(28.51%)㊁G>T(13.64%)和C>A(9.85%)㊂STRING数据库中构建Slit2相关基因蛋白网络,网络中共有21个相关基因,基因间的联系为92,平均节点相关度为8.76,区域聚类指数为0.776,相关蛋白网络富集明显(P<0.001)㊂蛋白网络相关基因KEGG信号通路主要富集于Ras信号通路㊁T细胞受体信号通路㊁Rap1信号通路和ErbB信号通路等㊂Slit2高低表达膀胱癌患者无疾病进展生存(DFS)和总生存(OS)间均存在明显的差异(P<0.05)㊂无疾病进展生存(DFS)(HR=1.5,P=0.026)和总生存(OS)(HR=1.50,P=0.015),在Slit2高表达组患者显著低于低表达组,提示Slit2高表达是膀胱癌预后不良的因素㊂ 结论 Slit2基因在膀胱癌患者癌组织中表达水平明显上调,高表达与膀胱癌患者的预后不良有关㊂关键词:膀胱癌;生物信息;Slit2基因;预后;信号通路中图分类号:R737.14㊀㊀㊀㊀㊀㊀文献标志码:A㊀收稿日期:2021⁃01⁃15㊀基金项目:河南省科技攻关计划(192102310092)㊀作者简介:李扬(1985⁃),男,硕士研究生,主治医师㊂研究方向:泌尿系肿瘤发生机制㊂㊀∗通信作者:朱朝阳(1968⁃),男,博士,主任医师,教授㊂研究方向:泌尿系肿瘤机制及规范化治疗㊂ExpressionandsignalpathwayofSlit2inbladdercanceranditsrelationshipwithprognosisLIYang WANGYun LIZun ZHUChaoyang∗DepartmentofUrology HuaiheHospitalofHenanUniversity Kaifeng475000 ChinaAbstract Objective ToinvestigatetheexpressionofSlit2inbladdercanceranditsrelationshipwithprognosis. Methods TheexpressionlevelsofSlit2geneinvarioushumantissueswereretrievedfromGTEXdatabasebybioinformatics.TheexpressionofSlit2inbladdercancerwasanalyzedinTCGAdatabase.TheSlit2genemutationwasanalyzedbycbioportal.AccordingtotheexpressionlevelofSlit2 thepatientsweredividedintohighandlowexpressiongroups andthedifferencesofdisease⁃freesurvival DFS andoverallsurvival OS betweenthetwogroupswerecompared. Results TheexpressionlevelofSlit2mRNAinbladdercancertissueswassignificantlyhigherthanthatinadjacentnormalbladdertissues P<0.05 andtheexpressionlevelofSlit2wasrelatedtotheclinicalstageofpatients thehigherthestage thehighertheexpressionlevel P<0.05 .ThemainmutationsofSlit2genewerefusionmutation amplificationmutation deepdeletionmutationandmultiplemutation.Amongthem deletionmutationaccountedfor25.55% synonymousmutationaccountedfor8.19% nonsensemutationaccountedfor2.01%.ThemaintypesofSNPswereC>T 28.51% G>T 13.64% andC>A 9.85% .Therewere21relatedgenesinthenetwork theassociationbetweengeneswas92 theaveragenode㊃20㊀㊃JournalofHenanUniversity(MedicalScience)2021,40(1)correlationwas8.76 theregionalclusteringindexwas0.776 andthecorrelationproteinnetworkwassignificantlyenrichedP<0.001 .KEGGsignalingpathwayismainlyenrichedinRassignalingpathway Tcellreceptorsignalingpathway Rap1signalingpathwayanderbBsignalingpathway.Thereweresignificantdifferencesindisease⁃freesurvivalDFS andoverallsurvivalOS inbladdercancerpatientswithhighandlowexpressionofSlit2P<0.05 .ProgressionfreesurvivalDFS HR=1.5 P=0.026 andoverallsurvivalOS HR=1.50 P=0.015 weresignificantlylowerinpatientswithhighexpressionofSlit2thanthoseinpatientswithlowexpressionofSlit2 suggestingthathighexpressionofSlit2isapoorprognosticfactorforbladdercancer. Conclusion TheexpressionlevelofSlit2geneissignificantlyup⁃regulatedinbladdercancer anditshighexpressionisassociatedwithpoorprognosisofbladdercancerpatients.Keywords bladdercancer bioinformatics Slit2gene prognosis signalingpathway㊀㊀膀胱癌是临床上较为常见的一类泌尿上皮性肿瘤㊂相关流行病学研究显示,全球范围每年膀胱癌新发病例高达45万,因膀胱癌死亡的患者超过15万[1]㊂因此,膀胱癌已成为肿瘤致死的重要原因之一㊂然而,膀胱癌确切发病的分子机制仍不十分清楚[2],且部分患者术后常常多次复发,是导致治疗失败和患者死亡的重要原因㊂因此,在分子机制上探讨膀胱癌发生㊁复发转移和预后分子标志物是目前的研究热点[3⁃4]㊂Slit2基因定位于人4号染色体,既往研究认为其在神经发育中发挥重要作用㊂近年来研究[5⁃8]显示,Slit2在多种实体肿瘤中呈现差异表达,并与其生物学行为如侵袭转移能力增强有关㊂Slit2在人膀胱癌中的研究报道不多㊂因此,本研究采用生物信息学方法探讨Slit2基因在膀胱癌中的表达情况及其与患者预后的关系㊂1㊀资料与方法1.1㊀Slit2表达水平分析采用生物信息学方法首先在GTEx(https://www.gtexportal.org/home/)数据库中检索slit2基因在人体各种组织中的表达水平㊂在TCGA数据库(https://portal.gdc.cancer.gov/)中分析Slit2多种肿瘤组织及在膀胱癌中的表达㊂采用TCGA数据库在线分析软件GEPIA(http://gepia.canc⁃er⁃pku.cn/detail.php)比较膀胱癌患者癌组织和癌旁组织中slit2基因mRNA表达水平及其与患者临床分期的关系㊂1.2㊀Slit2突变分析在用TCGA在线分析工具cBioPortal(http://www.cbioportal.org/)对Robo1的在膀胱癌及多种实体肿瘤中的突变情况进行分析㊂包括突变种类㊁突变频率及对应的突变位置㊂同时,在COSMIC(https://cancer.sanger.ac.uk/cosmic/)数据库中分析突变频率㊁单核苷酸突变位点及频率㊂1.3㊀Slit2相关基因蛋白网络构建及KEGG信号通路分析在STRING数据库中,构建Slit2相关基因表达的蛋白⁃蛋白相互作用网络㊂构建条件为:相互作用来源为实验㊁共表达㊁数据库㊁基因融合等;最小相互作用评分为0.07,相互作用蛋白不超过20个㊂同时在KEGG数据库中(https://www.genome.jp/kegg/pathway.html),对相互左右的蛋白网络进行信号通路富集㊂1.4㊀Slit2表达与患者预后根据Slit2表达水平,分为高低表达组(高表达组:Slit2基因mRNA表达水平>=肿瘤组织中的中未表达数;低表达组:Slit2基因mRNA表达水平<肿瘤组织中的中未表达数),比较两组患者无疾病进展生存(DFS)和总生存(OS)是否存在差异㊂高低表达组生存比较采用log⁃rank建议㊂1.5㊀统计分析数据分析采用STATA12.0软件完成,相关基因表达㊁预后㊁突变等分析采用在线数据分析工具,双侧P<0.05为差异有统计学意义㊂2㊀结果2.1㊀Slit2表达水平分析Slit2基因mRNA在人体各个组织中表达水平存在差异,且组织间表达水平变异程度较大,见图1A㊂TCGA数据库中,Slit2基因mRNA在膀胱癌患者癌组织中的表达水平明显高于癌旁正常膀胱组织(P<0.05),见图1B;且Slit2表达水平与患者临床分期有关,分期越高表达水平越高(P<0.05),见图1C㊂2.2㊀Slit2突变水平分析Slit2基因主要突变形式为融合突变㊁扩增突变㊁深度缺失突变和多重突变等,见图2A㊂具体突变位置,见图2B㊂其中缺失突变占25.55%,同义突变占8.19%,无意义突变占2.01%,见图3A㊂单核苷酸突变主要类型为C>T(28.51%)㊁G>T(13.64%)和C>A(9.85%),见图3B㊂2021,40(1)河南大学学报(医学版)㊃21㊀㊃图1㊀Slit2基因mRNA表达水平分析(A:Slit2基因在多种组织中表达;B:Slit2基因在膀胱癌及其癌旁组织中表达;C:RSlit2基因表达与膀胱癌分期)2.3㊀Slit2相关基因蛋白网络及信号通路富集STRING数据库中构建Slit2相关基因蛋白网络,网络中共有21个相关基因,基因间的联系为92,平均节点相关度为8.76,区域聚类指数为0.776,相关蛋白网络富集明显(P<0.001),见图4㊂蛋白网络相关基因KEGG信号通路主要富集于Ras信号通路㊁T细胞受体信号通路㊁Rap1信号通路和ErbB信号通路等,见表1㊂2.4㊀Slit2表达与患者预后Slit2高低表达膀胱癌患者无疾病进展生存(DFS)和总生存(OS)间均存在明显的差异(P<0.05)㊂DFS(HR=1.5,P=0.026)和OS(HR=1.50,P=0.015)在Slit2高表达组患者显著低于低表达组,提示Slit2高表达是膀胱癌预后不良的因素,见图5㊂3㊀讨论Slit基因于1984年被Jürgens等首次于果蝇胚体中发现,并发现其能够对果蝇幼体角色泽起到作用㊂1988年Rothberg等[9]报道Slit蛋白可以在果㊃22㊀㊃JournalofHenanUniversity(MedicalScience)2021,40(1)蝇中枢神经中线胶质细胞中合成,Slit蛋白减少可导致联合(交叉)神经元轴突和纵行传导通路在中线的聚集出现异常㊂Slit蛋白在哺乳动物体内主要由3个成员组成,分别为Slit1㊁Slit2和Slit3,三种蛋白均可于神经系统中线表达,也可于其他细胞类型中表达[10⁃12]㊂近年来,陆续有研究报道Slit2在多种人实体肿瘤组织中的表达水平明显高于对应的癌旁组织,提示Slit2在肿瘤的发生发展中可能发发挥重要作用[13⁃17]㊂图2㊀Slit2基因突变水平分析(A:Slit2基因主要突变形式;B:Slit2基因突变位置)图3㊀Slit2基因突变频率分析(A:各种突变频率;B:单核苷酸突变频率)2021,40(1)河南大学学报(医学版)㊃23㊀㊃图4㊀Slit2基因相互作用蛋白网络图表1㊀Slit2相关基因蛋白网络KEGG信号通路富集KEGG信号通路基因数背景基因P轴突引导151738.70ˑ10-25致病性大肠杆菌感染4531.02ˑ10-5白细胞跨内膜迁移41120.00012癌症的途径65150.00018趋化因子信号通路41810.00045ErbB信号通路3830.0009Ras信号通路42280.0009T细胞受体信号通路3990.0012神经营养素信号通路31160.0016产生IgA的肠道免疫网络2440.0061癌症中的蛋白多糖31950.0061肌动蛋白细胞骨架的调节32050.0062病毒性心肌炎2560.0074内吞作用32420.0086志贺氏菌病2630.0086黏合连接2710.0094细菌侵袭上皮细胞2720.0094结直肠癌2850.0118流体剪切应力与动脉粥样硬化21330.0263癌症中的MicroRNAs21490.031紧密连接21670.0366病毒致癌作用21830.0414焦点黏着21970.0454Rap1信号通路22030.046图5㊀Slit2基因表达与膀胱癌患者生存期分析(A:高低表达患者无疾病进展生存比较;B:高低表达患者总生存比较)㊃24㊀㊃JournalofHenanUniversity(MedicalScience)2021,40(1)㊀㊀然而,关于Slit2在膀胱癌中的表达及其与患者预后关系的研究报道多,其在膀胱癌中的生物学行为及作为预后分子标志物的可行性亦不明确㊂因此,我们采用生物信息学的方法探讨Slit2在膀胱癌中的表达情况㊁相关信号通路及其与患者预后关系㊂研究结果显示,Slit2基因mRNA在膀胱癌患者癌组织中的表达水平明显高于癌旁正常膀胱组织(P<0.05),且Slit2表达水平与患者临床分期有关㊂分期越高表达水平越高(P<0.05),说明Slit2可能起促癌作用,在膀胱癌的发生中发挥重要作用㊂进一步分析显示,Slit2基因主要突变形式为融合突变㊁扩增突变㊁深度缺失突变和多重突变等㊂其中缺失突变占25.55%,同义突变占8.19%,无意义突变占2.01%㊂上述突变的存在可能进一步影响基因表达产物水平,从而引发肿瘤的发生㊂Slit2相关基因KEGG信号通路主要富集于Ras信号通路㊁T细胞受体信号通路㊁Rap1信号通路和ErbB信号通路等㊂上述小信号通路大多与肿瘤的发生㊁发展及生物学行为有关㊂预后分析显示,Slit2高低表达膀胱癌患者DFS和OS间均存在明显的差异(P<0.05)㊂在Slit2高表达组患者显著低于低表达组,提示Slit2高表达是膀胱癌预后不良的因素㊂因此,Slit2基因在膀胱癌患者癌组织中表达水平明显上调,高表达与膀胱癌患者的预后不良有关㊂但研究本身单纯从生物信息分析水平证实了Slit2在膀胱癌组织中存在差异表达,并与预后有关,但其结论有待进一步相关体内外实验证实㊂参考文献:[1]SIEGELRL,MILLERKD,JEMALA.Cancerstatistics,2019[J].CACancerJClin,2019,69(1):7⁃34.[2]SIEGELRL,MILLERKD,JEMALA.Cancerstatistics,2020[J].CACancerJClin,2020,70(1):7⁃30.[3]季德才,郝斌,姜利宁,等.膀胱癌组织GPR48基因的表达及其临床意义[J].山东医药,2018(4):89⁃92.[4]左玉良,窦中岭.NMP22㊁CA19⁃9㊁CEA在膀胱癌中的表达及临床意义[J].河南科技大学学报(医学版),2015(4):252⁃256.[5]ALVAREZC,TAPIAT,CORNEJOV,etal.SilencingoftumorsuppressorgenesRASSF1A,SLIT2,andWIF1bypromoterhypermethylationinhereditarybreastcancer[J].MolCarcinog,2013,52(6):475⁃487.[6]DONGR,YUJ,PUH,etal.FrequentSLIT2promotermethylationintheserumofpatientswithovariancancer[J].JIntMedRes,2012,40(2):681⁃686.[7]孙瑜,陈公琰.肺癌相关的Slit2-Robo1信号通路的研究进展[J].实用肿瘤学杂志,2014(5):473⁃476.[8]张姣,付丽,谷峰,等.5⁃氮杂⁃2ᶄ⁃脱氧胞苷对MDA⁃MB⁃231乳腺癌细胞SLIT2基因去甲基化作用及细胞运动能力的影响[J].中华实验外科杂志,2012,29(1):70⁃73.[9]ROTHBERGJM,JACOBSJR,GOODMANCS,etal.Slit:anextracellularproteinnecessaryfordevelopmentofmidlinegliaandcommissuralaxonpathwayscontainsbothEGFandLRRdomains[J].GenesDev,1990,4(12A):2169⁃2187.[10]DALLOLA,MORTOND,MAHERER,etal.SLIT2ax⁃onguidancemoleculeisfrequentlyinactivatedincolorectalcancerandsuppressesgrowthofcolorectalcarcinomacells[J].CancerRes,2003,63(5):1054⁃1058.[11]SCHMIDBC,REZNICZEKGA,FABJANIG,etal.TheneuronalguidancecueSlit2inducestargetedmigrationandmayplayaroleinbrainmetastasisofbreastcancercells[J].BreastCancerResTreat,2007,106(3):333⁃342.[12]SHARMAG,MIRZAS,PRASADCP,etal.Promoterhypermethylationofp16INK4A,p14ARF,CyclinD2andSlit2inserumandtumorDNAfrombreastcancerpatients[J].LifeSci,2007,80(20):1873⁃1881.[13]TSENGRC,LEESH,HSUHS,etal.SLIT2attenua⁃tionduringlungcancerprogressionderegulatesbeta⁃cate⁃ninandE⁃cadherinandassociateswithpoorprognosis[J].CancerRes,2010,70(2):543⁃551.[14]LINYY,YANGCH,SHEUGT,etal.Anovelexon15⁃deleted,splicingvariantofSlit2showspotentialforgrowthinhibitioninadditiontoinvasioninhibitioninlungcancer[J].Cancer,2011,117(15):3404⁃3415.[15]黄小莉,曲军英.Slit2/Robo1在宫颈癌中的表达及意义[J].中国临床药理学杂志,2015,31(6):429⁃431,438.[16]黄金,姚珍薇.Slit2与肿瘤[J].重庆医学,2006,35(15):1414⁃1416.[17]李瑜,李启忠,李宏州,等.膀胱副神经节瘤临床探析(附3例报告)[J].河南大学学报(医学版),2019,38(2):126⁃128.[责任编辑㊀陈国剑]。
脑源性神经营养因子的生理作用

脑源性神经营养因子的生理作用脑源性神经营养因子(Neurotrophic factors)是指一类分泌于神经细胞和周围组织中的蛋白质,它能与神经细胞表面的受体特异性结合,并通过细胞内信号转导途径,调控神经元的生长、分化、存活和突触可塑性等生理功能。
这些分子包括神经生长因子(Nerve Growth Factor, NGF)、脑源性神经营养因子(Brain-derived Neurotrophic Factor, BDNF)、神经营养因子(NT)等。
在神经学、生物学和精神医学领域中,研究人员关注到神经营养因子的生理作用,探索如何利用其治疗神经性疾病。
神经营养因子对神经系统具有重要的调节作用,它们能够通过多种途径促进神经细胞的生长和再生,增强神经突触可塑性,改善神经节细胞的代谢和功能。
在许多神经性疾病中,神经营养因子含量减少或缺乏,导致神经元生长受阻、易于损伤,从而引起疾病的发生和进展。
因此,神经营养因子也成为一种研究和治疗神经性疾病的新途径。
一、神经生长因子神经生长因子(Nerve Growth Factor, NGF)是第一个被发现的神经营养因子,它是由目前罕见的先天性感觉神经病人以及高浓度的萎缩性侧索硬化患者分泌。
NGF主要在神经元细胞体和轴突中存在,并调节中枢神经系统、周围神经系统和免疫系统的发育及功能。
NGF受体主要集中于神经系统的神经节细胞和部分非神经系统细胞中,如基底节、纹状体等区域。
NGF与受体结合后,在神经系统中产生一些影响神经生长的效应,包括通过细胞增殖增加神经细胞数量,通过细胞存活增强神经细胞存活率,通过突触传递增强神经细胞与神经元之间的联系,从而使神经细胞生长和发育更为健康。
二、脑源性神经营养因子脑源性神经营养因子(Brain-derived Neurotrophic Factor, BDNF)是目前最为研究的神经营养因子之一,同时也是神经元保护和再生的重要分子。
BDNF主要在大脑皮层、海马、嗅球以及其他神经系统区域表达,参与调节神经元的形态、功能、存活和塑性。
p75神经营养因子受体在脑缺血后神经元凋亡中的作用

•综述•p75神经营养因子受体在脑缺血后神经元凋亡中的作用彭林辉李孔平詹丽璇左夏林徐恩广州医科大学神经科学研究所,广州医科大学附属第二医院神经内科510260通信作者:徐恩,Email:enxu@163 .net【摘要】p75神经营养因子受体(p75 neurotrophin receptor, p75NTR)为肿瘤坏死因子受体超家族成员,通过与原肌球蛋白受体激酶(tropomyosin receptor kinase, Trk)受体相互作用或与神经营养因子结合,介导多种复杂的信号转导通路,诱导突触生长和影响细胞存亡。
急性脑缺血后,p75NTR与神经生长因子前体(pro-nerve growth factor, proN G F)、分拣蛋白(sortilin)等多种效应因子结合,进而激活下游凋亡信号分子,导致神经元死亡。
因此,阐明p75NTR在急性脑缺血中介导神经元凋亡的通路及其分子机制对于研发急性脑缺血的新型治疗药物具有重要意义。
【关键词】脑缺血;受体,神经生长因子类;细胞凋亡;神经元基金项目:国家自然科学基金面上项目(81873745,81971124,81971233 );广州市科技计划项目(202002030072)DOI : 10.3760/cma.j.issn.1673-4165.2021.01.011Role of p75 neurotrophin receptor in neuronal apoptosis after cerebral ischemiaPeng Linhui, Li Kongping, Zhan Lixuan, Zuo Xialin, Xu EnInstitute o f Neurosciences, Department o f Neurology, the Second Affiliated Hospital o f Guangzhou MedicalUniversity, Guangzhou 510260, ChinaCorresponding author: Xu En, Email:************【Abstract】p75 neurotrophin receptor (p75N T R) is a member of the tumor necrosis factor receptorsuperfamily, which interacts with tropomyosin receptor kinase (Trk) receptor or binds neurotrophic factors.It mediates a variety of complex signal transduction pathways, induces synaptic growth and affects cellsurvival. After acute cerebral ischemia, p75N rR binds effector factors such as pro-nerve growth factor(proNGF) and sortilin, activating downstream apoptotic signal molecules and leading to neuronal death.Therefore, elucidating the pathways and molecular mechanisms of p75N T R that mediate neuronal apoptosis inacute cerebral ischemia is of great significance for the development of new therapeutic drugs for acutecerebral ischemia.【Key w ords】Brain ischemia; Receptors, nerve growth factor; Apoptosis; NeuronsFund programs : National Natural Science Foundation of China (81873745, 81971124, 81971233);Science and Technology Program of Guangzhou, China (202002030072)DOI : 10.3760/cma.j.issn.1673-4165.2021.01.011急性脑缺血可引起神经细胞凋亡、坏死、自噬、焦亡和坏死性凋亡,作为神经元程序性死亡的一种 方式,凋亡尤为常见。
神经营养学的新研究进展

神经营养学的新研究进展神经营养学是指研究营养对神经系统发育、功能和代谢的影响。
过去几十年中,神经营养学的研究一直处于缓慢的发展状态,但最近几年来,随着科技的不断进步和对大脑和神经系统的认识不断深入,神经营养学的新研究进展正在掀起一股新的浪潮。
一、神经营养素的新认识神经营养素是指对神经系统具有营养作用的物质,包括脂肪酸、氨基酸、维生素等。
最近的研究显示,某些神经营养素不仅能影响神经系统的发育和功能,还能调节细胞信号转导通路、促进细胞存活和减少氧化应激等作用。
例如,多不饱和脂肪酸对神经系统的保护作用已得到广泛研究。
研究发现,多不饱和脂肪酸能够改善神经细胞膜的稳定性,促进神经元的成长和发育。
此外,多不饱和脂肪酸对大脑中的神经递质和神经素的合成和释放也有很大的影响,因此多不饱和脂肪酸摄入不足可能会导致神经系统的功能失调。
二、肠道菌群与神经系统的相互作用近年来的研究显示,肠道菌群与神经系统之间存在复杂的相互作用。
肠道菌群可以通过共生作用、代谢产物和肠道-脑轴向等多种途径影响神经系统的发育和功能。
例如,乳酸菌等益生菌可以通过调节神经递质的合成和释放来影响神经系统的功能。
此外,肠道菌群还会产生多种代谢产物,如短链脂肪酸,这些代谢产物可以对神经系统辅助调节神经元的发育和活动。
最近,一项新的研究发现,肠道菌群中的一种菌群能够分泌翁二酸,这种物质能够穿过血脑屏障,影响大脑的血流和氧气供给,从而调节神经系统的功能。
三、环境因素对神经系统的影响除了营养因素,环境因素也对神经系统的发育和功能具有重要的影响。
最近的研究表明,噪声、电磁辐射、气体污染等环境因素都可能对神经系统产生负面影响。
噪声是一种物理刺激,长时间暴露在噪声环境中会导致神经系统的疲劳和抑郁。
电磁辐射包括电磁波、微波辐射等,长时间暴露在电磁辐射环境中会影响神经元的代谢和信号传导。
气体污染也会对神经系统产生负面影响,例如颗粒物、二氧化硫等气体污染物可经过血脑屏障,直接影响神经元的存活和活动。
脑源性神经营养因子的作用及发挥

脑源性神经营养因子的作用及发挥脑源性神经营养因子,是指一类由脑细胞合成的生物活性分子,对于维持神经系统健康、发育和修复具有重要作用。
它们能够通过神经元的自身信号通路、血液循环或其他途径,对神经系统的各个方面进行支持和调节,如促进神经元的生长、发育和分化,促进突触形成和神经传递,促进神经元的修复和保护等。
本文将着重阐述脑源性神经营养因子的作用及发挥。
首先,脑源性神经营养因子在神经元生长和发育中发挥重要作用。
神经元的发育过程是一个高度复杂的生物事件,其中涉及到各类细胞因子、生长因子的作用。
脑源性神经营养因子在其中扮演着促进神经元发育和整合的重要角色。
例如,神经营养因子(Neurotrophins)即是最早被发现的脑源性神经营养因子之一,具有促进神经元生长、分化和存活的作用。
而神经生长因子(NGF)更是被认为是神经元发育和维持的关键因子之一。
它通过激活神经元的特定受体,触发神经元突触形成、强化和保护,从而促进神经元与外界的信息交流和适应。
其次,脑源性神经营养因子对神经元突触稳定和塑性具有关键作用。
突触在神经传递中扮演着重要角色,而突触的形成与稳定则需要神经营养因子的支持和调节。
例如,突触连接和塑性的核心分子是神经元钙调蛋白(CaMK)和脑源性BDNF等分子,它们可通过调控神经传递途径、增强认知功能等多种方式,从而维持和提高神经系统的适应性和应激能力。
此外,研究发现,脑源性神经营养因子还能够调控突触间的转运、信号传递和塑性程度等因素,从而为神经系统的各类行为和感觉提供基础支持。
另外,脑源性神经营养因子对神经元修复和保护也具有重要作用。
神经系统受到外界的伤害或病理性损伤,将会导致神经元失去正常功能和结构,从而影响神经系统的整体稳定性和功能。
而许多脑源性神经营养因子能够通过激活细胞内修复机制和促进神经元活性,从而促进神经元的再生、保护和修复。
例如,在中风、脑损伤和退行性神经病等病理刺激下,神经营养因子的表达趋势发生变化,多数情况下是增加。
神经细胞生长和分化的分子机制研究

神经细胞生长和分化的分子机制研究神经细胞是组成我们神经系统的基本单元。
它们能够接收和处理信息,并将信息传递到其他神经细胞,从而实现神经系统的完整功能。
神经细胞的生长和分化是神经系统发育的重要过程,也是理解神经系统功能的基础。
近年来,分子生物学的发展极大地推动了神经细胞生长和分化机制的研究,为我们深入了解神经系统的发育和功能提供了新的思路和方法。
一、神经细胞生长神经细胞的生长包括突触发育、树突、轴突和轴突末梢的生长和分化等过程。
这些过程是由许多基因和分子机制调控的。
其中,神经营养因子和神经生长因子是对神经细胞生长和分化影响最为明显的因子。
神经营养因子是神经系统中的化学信使,它们在细胞外作用,通过与细胞表面的受体结合,调控神经细胞的生长和分化。
目前已经发现了多种神经营养因子,如神经营养因子、乙酰胆碱、ATP等。
这些因子的作用机制复杂,其中部分因子可以直接增强神经细胞的突触效能,从而促进突触的形成和稳定。
此外,一些神经营养因子也可以促进轴突生长和树突分支的形成。
神经生长因子是一类对神经细胞具有专一调节作用的多肽物质。
它们参与调控神经细胞的突触发育、轴突、树突的生长,以及神经元的生存和凋亡。
例如,NGF、BDNF等神经生长因子都能够促进神经突触的形成和增强轴突的生长,同时还能够促进神经细胞的生存和抗氧化能力。
二、神经细胞分化神经细胞分化是指神经干细胞向神经元和神经胶质细胞分化的过程。
它是神经系统发育过程中最关键的一环。
在神经细胞分化过程中,一系列基因和分子机制调控细胞的增殖、分化和修饰。
其中,转录因子和信号通路是神经细胞分化的关键因素。
转录因子是一类能够控制基因表达的分子,它们能够识别DNA上的特定序列,并激活或抑制基因表达。
在神经细胞分化过程中,一些转录因子如Ngn1、Sox2等起到了关键作用。
它们调控神经干细胞的增殖和分化,促进神经元和神经胶质细胞的分化。
信号通路是神经细胞分化中的另一个重要因素。
神经细胞分化所需的信号通路非常复杂,如Notch信号通路、Wnt信号通路等。
神经营养素-3及其在脊髓损伤修复中的作用

神经营养素-3及其在脊髓损伤修复中的作用
张冉
【期刊名称】《中南医学科学杂志》
【年(卷),期】2013(041)002
【摘要】神经营养素-3(NT-3)属于神经营养素(NTs)家族,是神经系统神经元存活、分化和功能维持的重要营养因子,在脊髓损伤后的神经修复方面发挥了重要作
用.NT-3分泌后能通过p75 NTR和Trks受体激活其下游信号通路,促进轴突生长
和血管发生.本文就NT-3及其受体结构、受体信号以及在脊髓损伤修复中的研究
进展作一综述,并对NT-3与其他药物联合治疗脊髓损伤(SCI)的临床应用前景进行
展望.
【总页数】6页(P196-200,210)
【作者】张冉
【作者单位】南华大学医学院组织学与胚胎学教研室,湖南衡阳421001
【正文语种】中文
【中图分类】R329.1
【相关文献】
1.巨噬细胞调节胶质瘢痕在脊髓损伤修复中的作用研究进展 [J], 宋公宇;章涛;张潜
2.神经营养素-3及其在脊髓损伤修复中的作用 [J], 张冉(综述);龙双涟(审校);
3.神经元胞外基质网络在脊髓损伤修复作用中的研究进展 [J], 胡蓉;徐海鹏;何克林;陈怡;吴磊;马睿杰
4.外泌体的神经免疫调节功能在脊髓损伤修复中作用的研究进展 [J], 丁华;张磊;袁
即山;樊晓臣;姚翔;吕斌
5.PD1在脊髓损伤修复中的作用和机制研究 [J], 蔡赟赟;邢玲燕;杨拓;沈卫星;陈罡因版权原因,仅展示原文概要,查看原文内容请购买。
神经系统发育和细胞分化的分子机制和调节

神经系统发育和细胞分化的分子机制和调节神经系统发育和细胞分化是一项非常复杂的过程,其发生需要多种因素的共同作用。
在这个过程中,细胞必须遵循一个非常精密的程序,以确保它们能够正确地分化并形成神经系统的不同部分。
在本文中,我们将讨论神经系统发育和细胞分化的分子机制和调节,以加深读者对这个过程的理解。
神经系统分化和发育的机制神经系统的分化和发育是由多个分子机制来控制的,它们依靠基因表达的正规调节和活化而发生。
最初,未分化的神经前体细胞通过神经生长因子(NGF)被激活,然后分化为神经细胞或谷氨酸能神经元。
在分化过程中,细胞通过发送和接收不同类型的信号来调节细胞命运的转变。
这些信号涉及细胞外基质,激素,神经生长因子,脂质分子等。
分子机制的意义在于促进细胞分化,确定其命运,并使其遵循一个连贯的分化过程。
参与神经系统分化和发育的基因神经系统发育和分化涉及基因表达的严格调节,这是由多种因素影响的。
其中一些常见的基因包括:1. Sonic Hedgehog基因(SHH):该基因通过影响细胞分化和增殖,调控中枢神经系统(CNS)和周边神经系统(PNS)的发育。
这个基因是中枢神经系统的建立过程中必不可少的关键因素之一。
2. Neurogenin基因(NGN):该基因控制神经细胞的形成,特别是在中枢神经系统中的神经元、星形细胞和少突胶质细胞的形成,因此也被称为神经分化基因。
3. Beta-catenin基因(CTNNB1):该基因通过对细胞周期调控以及细胞类型命运的决定,参与神经系统的生长和分化。
4. Myelin basic protein基因(MBP):该基因编码髓鞘蛋白的组成部分,对中枢神经系统启动过程的实现非常重要。
神经系统分化和发育中的细胞信号途径神经系统分化和发育中涉及的信号通路是非常显着的。
下面是一些常用的信号通路。
1. Wnt/β-catenin信号途径:该通路可以通过对细胞命运的决定,对神经系统分化和发育进行基因调节。
cKit信号通路在抑郁症致病机理中的作用的开题报告

SCF/cKit信号通路在抑郁症致病机理中的作用的开题报告开题报告:SCF/cKit信号通路在抑郁症致病机理中的作用一、研究背景抑郁症是一种常见的心理障碍,其病因机制至今尚未完全明确。
其中,神经营养因子和神经生长因子的变化被认为是导致抑郁症的重要因素。
已有研究表明,SCF/cKit信号通路(Stem cell factor/c-KIT signalling pathway)对神经元的增殖、分化、迁移和成熟等生命活动发挥重要作用,与神经营养因子和神经生长因子有联系。
因此,探究SCF/cKit信号通路在抑郁症致病机理中的作用,有望为研究抑郁症的发病机制提供新的思路和方向。
二、研究内容本研究将使用抑郁症动物模型,探究SCF/cKit信号通路在抑郁症发病机理中的作用。
主要研究内容包括:1. 构建抑郁症动物模型:使用慢性不可逆性压力法,建立抑郁症动物模型。
2. 观察SCF/cKit信号通路对抑郁症模型大鼠行为学的影响:采用行为学实验,如强迫游泳试验、开放场测试等,观察SCF/cKit信号通路对抑郁症模型大鼠行为学的影响。
3. 检测SCF/cKit信号通路在抑郁症模型大鼠脑组织中的表达:采用Western blot和免疫组化等技术手段,检测SCF/cKit信号通路在抑郁症模型大鼠脑组织中的表达及其影响因素。
4. 探究SCF/cKit信号通路调控神经元幸存和突触可塑性的机制:采用细胞和分子生物学方法,研究SCF/cKit信号通路对神经元幸存和突触可塑性的调控机制。
三、研究意义本研究将有助于深入探究SCF/cKit信号通路在抑郁症致病机理中的作用机制,为进一步研究抑郁症的发病机制提供新思路和方向。
另外,本研究还将有助于拓展抑郁症治疗的新途径,为临床诊断和治疗提供参考。
眼睛的发育和疾病的分子机制

眼睛的发育和疾病的分子机制眼睛是我们日常生活中不可缺少的视觉器官,其发育和疾病的分子机制一直是生命科学的重要研究方向。
本文将从眼睛发育的基本过程入手,探讨分子机制对其发育和疾病的影响。
眼睛的发育一般分为眼前段和眼后段发育两个阶段。
眼前段包括角膜、房水、睫状体等组织的发育,眼后段主要包括玻璃体、视网膜、脉络膜等组织的发育。
这两个阶段的发育均涉及复杂的分子调控机制。
眼前段发育的分子机制主要包括调控角膜、前房形成的基因和信号通路。
关于初始角膜的形成,PAX6是关键的基因,其发挥调控神经、眼睛和胰岛素的功能,而在眼前段的发育过程中则主要调控初生角膜的形成和后续层次的维护和再生。
和PAX6类似,Vsx2是对前房和睫状体发育至关重要的转录因子,并且与肝素生长因子信号、FGF信号等多个重要信号通路互作。
眼后段发育的分子机制关键在于视网膜的发育。
视网膜是视觉摄像的根基,在发育过程中需要调控的基因与信号通路也极为复杂。
多种信号通路包括神经营养因子信号通路、Wnt信号通路、Notch信号通路等均在视网膜的发育中发挥重要作用,信号分子的调控、细胞形态的调整等在其中均扮演着关键的角色。
除了正常发育外,眼睛疾病也是大众关心和关注的问题。
眼病包括青光眼、白内障、角膜移植等多种类型。
不同的眼病因素不同,在治疗方案上也有所不同。
但是,目前越来越多的研究表明,眼病因子与眼球的分子机制密切相关。
青光眼是眼压升高导致视神经受损,进而致盲的一种眼病,其分子机制与角膜、房水体液有关。
TIGR/MYOC信号通路是导致青光眼的重要途径之一。
白内障是晶状体发生混浊,进而致失明的一种眼病。
过氧化氢稳态通路和氧化应激均和白内障相关。
另外,多种免疫相关因子和脂质代谢方面的调控均可能与白内障的发生和发展有关。
角膜移植则涉及到角膜上皮修复分子机制、角膜免疫失调分子机制等。
总的来说,眼睛的发育和疾病的分子机制彼此之间密不可分。
这些分子机制的调控涉及多种信号、基因、细胞等多个方面,是生命科学的热门研究方向之一。
神经营养因子总结

睫状神经营养因子
(Ciliary neurotrophic factor,CNTF)
1980年,Adler等由E8鸡胚眼中分离到CNTF,分子量20kDa的酸性
蛋白,由199aa组成,进化上比较保守,与LIF、IL-6等构成成血细 胞因子(hematopoietic cytokine)超家族
没有直接证据显示NGF是由其靶组织合成,或 者说神经元从靶组织摄取。 神经组织中受体的存在? 特别是,NGF的信号只在生长的神经元轴突接 触到其靶标后才出现
18
NGF summay
NGF可以介导两类特殊的神经细胞类群~交感 神经元和一些感觉神经节细胞)
相关神经元在NGF剥夺的情况下会死亡;而更多的 NGF则可以使更多的神经元存活; 存在于神经元的靶组织; 在相关的投射神经元中存在NGF的受体。 ~~~~~~~~神经营养因子的金标准
43
CNTF的主要生物效应
对中枢和周围运动神经元的营养作用 影响非神经组织
44
45
GDNF的主要生物效应
对多巴胺能神经元的营养支持 促进运动神经元发育、修复损伤后运动神经元 对感觉神经元的营养支持
46
NTF的作用模式
逆行信号转导(靶源性的):经典的NTF作用途径; 靶细胞合成NTF,经轴突末端的受体介导,逆行轴突 转运到胞体
47
48
NTF的作用模式
顺行信号转导:许多靶神经元依赖于传入投射维持其 存活,上游神经元通过大分子物质的顺行轴突转运对 靶神经元起神经营养作用
49
NTF的作用模式
NTF的局部作用(旁分泌方式):靶组织与其支配的
神经元共存于同一组织,来自靶合成的NTF作用于临 近效应神经元
神经营养因子受体的研究进展

神经营养因子受体的研究进展(作者:___________单位: ___________邮编: ___________)【关键词】神经营养因子;受体;信号转导神经营养因子家族在神经细胞的生长发育、保护修复过程中起着极其重要的作用。
而神经营养因子受体是启动信号转导,产生生物学效应的重要物质。
根据同源性大小、基因表达部位和蛋白作用的专一性以及信号传递机制的不同,可将神经营养因子分为三个家族:神经生长因子家族、睫状神经营养因子家族和胶质细胞源性神经营养因子。
本文从结构、功能、信号传递机制等方面,对其相应受体的最新研究进展作一综述。
1 神经生长因子家族受体主要成员为神经生长因子(NGF),脑源性神经营养因子(BDNF),神经营养素3(neurotrophin3,NT3),神经营养素4/5(neurotrophin4/5,NT4/5)。
这些因子最具有代表性的受体为高亲和力受体(Trk)和低亲和力受体(p75NTR),p75NTR受体隶属于肿瘤坏死因子受体家族。
1.1 Trk受体1.1.1 Trk结构 Trk受体家族包括TrkA(p140Trk,主要结合NGF)、TrkB(p145Trk,主要结合BDNF、NT4/5)和TrkC(相对特异的结合NT3) 。
它们在发育的不同时期、不同组织的神经细胞表达不同,而神经生长因子家族的生物学效应主要由高亲和力受体介导,使其表达具有明显阶段特异性和组织特异性。
Trk的细胞膜外结构包括独特的IgG C2区及富含半胱氨酸、亮氨酸的重复结构,以往研究证实生长因子的结合部位位于第二个免疫球蛋白样重复序列上,它的氨基酸排列顺序决定了不同的Trk 受体的特异性及与不同的生长因子的亲和力大小不同。
近期Ultsch 〔1〕已成功探测出各Trk受体上与配体结合部位的晶体结构;此外,Wiesmann等〔2〕也已经公布了NGF与TrkA结合部位的结构,此结构包括两部分:一部分是所有神经营养因子所共有的保守模序,另一部分是TrkA所特有的。
神经营养因子的分泌与调控机制

神经营养因子的分泌与调控机制神经营养因子是一类在神经系统发育和功能维护中起重要作用的分子,包括神经生长因子(NGF)、脑源性神经营养因子(BDNF)等。
它们具有广泛的生物学功能,不仅参与神经细胞的生长分化、突触形成和保护,还对学习记忆、情感行为等高级神经功能发挥作用。
在本文中,我们将讨论神经营养因子的分泌和调控机制。
1. 神经营养因子的分泌神经营养因子在体内的来源种类多样。
最初被发现的NGF主要来源于目标器官,如胰腺、肝脏、肾脏等。
还有一些神经营养因子在分泌前需要经历胞内转运和高级调控,如在细胞质中转变为成熟的蛋白质,此时它们才能顺利地分泌到体外,参与神经系统的发育和维护。
神经营养因子的分泌是由多种信号通路调控的。
经过多年的研究,人们发现神经营养因子的分泌受到多种体内信号的调节,如胆碱能、脂质、氧化应激等。
这些信号调节可以直接或间接地通过转录或后转录调控神经营养因子的基因表达水平,或者通过膜通道和细胞外受体信号转导途径来调节神经营养因子的分泌。
2. 神经营养因子的调控机制神经营养因子的调控机制涉及到许多信号通路和分子机制,主要包括以下几个方面。
2.1 神经元活动神经元的活动是神经营养因子调控的重要环节之一。
一些研究表明,神经元的活动可以促进BDNF等神经营养因子的分泌。
例如,提高神经元活性的方法,如体外电刺激、体外模拟突触传递等,都可使BDNF分泌量显著增加。
这种机制被认为是神经营养因子的回路调节,即神经元活动通过分泌神经营养因子来调节其自身和与之相关的神经元。
2.2 细胞因子细胞因子包括多种介质,如白细胞介素1(IL-1)、白细胞介素6(IL-6)、白细胞介素10(IL-10)等,可以直接或间接地影响神经营养因子的分泌。
有研究表明,IL-1和IL-6均可以促进NGF的分泌,而IL-10则抑制NGF的分泌。
这些分子通过信号转导途径调节神经营养因子表达,从而调控神经系统的功能状态。
2.3 脂质代谢脂质代谢是神经营养因子调控机制的重要组成部分。
神经营养因子对神经元生长发育的调控作用

神经营养因子对神经元生长发育的调控作用作为人体的重要部分,神经系统起着极为重要的调控作用。
而神经元是神经系统的基本单位,它的生长发育与整个神经系统的功能密切相关。
神经元的生长发育受许多因素的影响,其中神经营养因子是调节神经元生长发育的重要因素之一。
本文将从神经营养因子对神经元生长发育的调控作用入手,深入探讨神经营养因子在神经元研究领域中的重要性。
神经营养因子的种类与作用机制神经营养因子是指呈多肽结构的低分子量蛋白质,它们能够调控神经元的凋亡、增殖、分化和迁移。
常见的神经营养因子包括神经生长因子(NGF)、脑源性神经营养因子(BDNF)、神经元生长抑制因子(Nogo)、神经元特异性烷基化因子(MeCP2)等。
这些神经营养因子的作用机制复杂,主要包括长作用的信号途径和短作用的信号途径。
其中长作用的信号途径需要多个步骤的反应过程,例如NGF可使神经元产生突触增长,同时通过PI3K和AKT信号途径阻止神经元凋亡。
而短作用的信号途径则能够快速传递神经元信号,例如BDNF通过TrkB受体介导的神经元突触神经可塑性,从而调控神经元的发育和成熟。
神经营养因子对神经元增殖与分化的调控作用神经元的增殖和分化是神经元生长发育的重要过程。
神经元增殖与分化的失控可导致各种神经系统疾病的产生。
神经营养因子在神经元增殖和分化的过程中起到了关键性的作用。
NGF是神经元增殖的主要因子之一。
它可通过三种通路促进神经元增殖,分别为TRKA / p75NTR / sortilin信号通路。
研究发现,NGF通过TRKA途径激活Akt,促进增殖,同时通过p75减少微管关联蛋白2的稳定性,从而增加神经元的动态不稳定性。
BDNF也能够促进神经元增殖。
BDNF与TrkB结合后激活Ras-Raf-MEK-MAPK信号通路,从而促进神经元的增殖和分化。
神经营养因子对神经元迁移的调控作用神经元迁移是神经元发育过程中难以绕过的重要阶段。
神经元的迁移是由内皮细胞、外细胞基质及积累在神经元胚胎时期的分子通过次级信号途径调节的。
生长因子及其信号通路的研究

生长因子及其信号通路的研究生长因子是一类能够促进细胞生长和增殖的分子,它们通过与细胞表面的受体结合,在细胞内部启动一系列信号传递途径,使细胞进入增殖状态,同时也会调节细胞形态、分化、迁移和细胞死亡等生命过程,从而对组织和发育过程发挥非常重要的作用。
生长因子是一类非常广泛的分子,目前已发现了上百种,包括成纤维细胞生长因子(FGF)、神经营养因子(NGF)、表皮生长因子(EGF)、人类生长激素(HGH)等,它们在不同细胞类型和生命阶段发挥着不同的作用。
生长因子信号通路是一系列分子级联反应的组合,被称为“生长因子信号通路”,其主要作用是将生长因子的信号转导至细胞内部,从而促进细胞生长和增殖。
生长因子信号通路可以大致分为三个部分:第一个部分是生长因子与其受体的结合和激活,这个过程会使受体二聚化,并开始激活其下游的信号分子;第二个部分是信号分子的级联反应,包括了各种酶、激酶和蛋白质等,在这个过程中,信号会从受体传递到细胞的细胞质或细胞核区域,从而启动各种信号途径;第三个部分是转录因子的激活和基因表达的调控,使细胞进入生长状态,从而完成细胞分化、增殖和形态的改变等生命过程。
对于生长因子及其信号通路的研究非常重要,它可以帮助我们理解一些疾病的发生机制,并开发出更有效的治疗方法。
例如,某些肿瘤具有特定的生长因子异位表达或受体过度表达,这些因子和受体的异常表达会导致细胞生长和增殖的异常,从而发生恶性肿瘤。
现在已有一些针对这些生长因子或受体的抑制剂或抗体药物被开发出来,它们可以有效地抑制肿瘤的生长和扩散,是目前治疗肿瘤的重要药物之一。
在生长因子信号通路的研究中,还涉及到一些非常重要的分子,例如:Ras、MAPK、Akt、PI3K等。
这些分子在生长因子信号通路中扮演着非常重要的角色,它们可以调节细胞的生长、分化,维持细胞的正常功能,同时也参与了一些疾病的发生和发展。
其中,Ras基因突变是导致多种类型恶性肿瘤的重要原因之一,因此,研究Ras在生长因子信号通路中的作用对于治疗某些肿瘤非常重要。
细胞内信号通路对神经发育的调节

细胞内信号通路对神经发育的调节神经发育是一种复杂而严密的过程,它涉及许多因素的相互作用。
细胞内信号通路是神经发育中重要的调节因素之一。
细胞内信号通路从细胞表面的受体开始,通过一系列的途径,最终使得细胞做出相应的反应。
在神经发育中,细胞内信号通路主要发挥着调节神经系统发育与分化、神经元迁移、网络形成和突触可塑性等多方面作用。
一、细胞内信号通路对神经元分化与迁移的调节神经元的生成和分化是神经系统发育的关键步骤。
在神经发育早期,一定的信号通路可以诱导神经干细胞向神经元方向分化。
其中,Wnt信号通路、Notch信号通路和Shh信号通路是最为经典和重要的信号通路之一。
Wnt信号通路通过强烈激活β-catenin的下游分子,增加神经元前体细胞的增殖和分化,促进神经系统的发育成熟。
Notch信号通路则通过抑制Ngn1和Ngn2等神经元特异性转录因子的表达,在神经元分化方面发挥着负调控作用。
Shh信号通路能够调节神经干细胞的增殖和分化,同时指导神经元前体细胞分化成各种具体类型的神经元和神经胶质细胞。
除了分化,神经元生成后的迁移也需要特定的信号通路调控。
宋教授团队在2007年发现,Drag1可以通过调控Cdc42和Pak1等蛋白的表达,促进胚胎期间颅神经神经元的迁移。
以Drag1为例,它在神经元迁移加速期表达量高,而在迁移减缓期表达量时低。
这说明Drag1是神经元的一个关键调控分子,可揭示线粒体运动与神经元迁移联系的分子机制。
二、细胞内信号通路对神经网络形成的调节神经网络形成是神经系统发育中的另一重要过程。
神经元通过突触连接形成复杂的神经网络,在此过程中细胞内信号通路也扮演着重要的角色。
下面以铁元素为例来简单介绍一下。
铁元素在神经网络形成中的调节作用:铁元素参与许多细胞代谢过程,如DNA复制、蛋白质合成、电子传递、信号转duction等。
一项研究表明,与含铁蛋白质结合的铁元素与神经营养因子的交互作用能诱导神经元生长锥的扩散和生长,同时还能加强突触形成和突触前细胞的活动,从而促进神经网络形成。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Nykjaer et al. 2004 & 2008
• The function of p75NTR depends on the cellular context; activation of p75NTR promotes death in numerous cells, including injured neurons, but promotes migration, growth and survival in other cells.
Chao MV. 2004
• The ratio of receptors is important in dictating the numbers of surviving cells.
• Interactions between p75 and Trk receptors provide greater discrimination between different neurotrophins.
• To transmit the signal over a long distance, neurotrophins and activated Trks are transported together in endocytic vesicles to the cell soma.
TRAF6 (TNF receptor associated factor) Ub (Ubiquitin) – Proteasome pathway of protein degradation
Thangiah Geetha, Jianxiong Jiang, Marie W. Wooten. 2005
• Internalization and signaling is regulated by polyubiquitination of Lys485 of TrkA.
Ubiquitin properties (human)
• b, In contrast, when bound to unprocessed NGF (proNGF), a complex of p75NTR and sortilin induces cell death.
Nykjaer et al. 2004 & 2008
• How could NGF promote survival through TrkA and death through p75NTR — sometimes in the same cell type?
• The mature proteins:
– about 12 kDa in size, form stable, non-covalent dimers, – normally expressed at very low levels during development. – The amino-terminal half (or pro-domain) of the pro-
•1
Nykjaer et al. 2004 & 2008
• The interaction of p75NTR with TrkA enhances the responsiveness of TrkA to nerve growth factor (NGF) to promote survival and growth.
Secondary structure
Symmetry
Models of Trk and p75 receptor
• Trk receptors contain extracellular immunoglobulin G (IgG) domains for ligand binding and a catalytic tyrosine kinase sequence in the intracellular domain.
• However, in reality, the binding of NGF to TrkA, and of BDNF to TrkB is of low affinity, but it can be regulated by receptor dimerization, structural modifications or association with the p75 receptor.
• Growth factors such as neurotrophins that promote the survival of neurons are known as neurotrophic factors.
• Secreted by target tissue and act by preventing the associated neuron from initiating programmed cell death(PCD) - thus allowing the neurons to survive.
• The extracellular portion of p75 contains four cysteine-rich repeats, and the intracellular part contains a death domain. Neurotrophin binding to the p75 receptor mediates survival, cell migration and myelination through several signalling pathways.
NGF – Nerve growth factor BDNF – Brain-derived neurotrophic factor NT3 – Neurotrophin 3 NT4 – Neurotrophin 4
Function
• A class of growth factors, secreted proteins that are capable of signaling particular cells to survive, differentiate, or grow.
– Trk receptor tyrosine kinases – p75 neurotrophin receptor (p75NTR).
• Transmembrane receptor, also known as TNFRSF16. • a member of the tumour necrosis factor receptor (TNFR) death-
neurotrophin is believed to be important for the proper folding and intracellular sorting of neurotrophins.
Neurotrophin Receptors
• Each neurotrophin can signal through two different types of cell surface receptor
• Neurotrophins also induce differentiation of progenitor cells, to form neurons.
PDB 1SG1
Chao MV. 2004
Biogenesis
• The neurotrophins are initially synthesized as precursors or pro-neurotrophins, which are cleaved to produce the mature proteins. (ProNGF -> NGF)
receptor family.
Models of Trk and p75 receptor
Chao MV. 2004
• Different neurotrophins show binding specificity for particular receptors.
• These interactions have generally been considered to be of high affinity.
Lys-11 or Lys-48: marking the substrate for ubiquitin–proteasomemediated degradation. Lys-63: Marking substrates for endocytosis or a role in intracellular signaling such as NF-κB activation. Lys-6, Lys-27, Lys-29 and Lys-33: have not been elucidated.
• As a result, increased ligand selectivity can be conferred on the Trk receptors by the p75 receptor.
Crystal Structen TrkA and p75
Neurotrophin Signaling (Trk Signaling Pathway)
Neurotrophins
• The neurotrophins are a family of proteins that are essential for the development of the vertebrate nervous system.
Chao MV. 2004
• The p75 receptor can bind to each neurotrophin, and also acts as a co-receptor for Trk receptors.