大鼠巨颈囊尾蚴线粒体pcox1基因的分子鉴定
二化螟线粒体cox1基因的克隆、序列测定和分子系统学分析

二化螟线粒体cox1基因的克隆、序列测定和分子系统学分析汪爱民;共桂云;魏兆军【摘要】[目的]克隆并分析二化螟线粒体细胞色素c氧化酶亚基I基因(cox1).[方法]利用PCR方法扩增了二化螟线粒体cox1基因,并测定了其全序列.通过检索GenBank数据库获得了其他21种鳞翅目昆虫的cox1序列,并进行了同源性比较和分子系统学分析.[结果]cox1基因编码框包含1 531个核苷酸,编码510个氨基酸的蛋白;起始密码子为CGA,终止密码子仅由一个T组成.利用ML方法构建了基于cox1基因编码氨基酸序列的鳞翅目昆虫的分子系统树,发现分子系统树与从形态学角度的系统分类在大方面上是基本一致的,但也略有差异.[结论]为进一步研究cox1基因的表达和应用奠定了基础.%[Objective] The research aimed at cloning and analyzing mitochondrial cytochrome oxidase I gene (coxl) of Chilo suppressalis. [ Method] The mitochondrial coxl gene of Chilo suppressalis was cloned with PCR method and then sequenced. Then, coxl sequences of other 21 Lepidopteran species were obtained by blasting the GenBank with coxl gene sequence of C. Suppressalis. Finally, homology comparison and molecular phylogenitic analysis among the 22 Lepidopteran species were conducted. [ Result] The opening reading frame of coxl gene from C. Suppressalis contained 1 531 nucleotides encoding a putative protein of 510 amino acids. The coxl gene used a start codon CGA, and an incom plete termination codon composed of only T. Based on the amino acid sequences of coxl, the molecular phylogenetic tree of Lepidoptera was reconstructed using the maximum likelihood ( ML) method. The molecular phylogenetic tree was similar to the morphologicalphylogenetic tree mainly, but also showed some differences. [ Conclusion] The result will provide reference for further research on expression and application of the coxl gene.【期刊名称】《安徽农业科学》【年(卷),期】2011(039)021【总页数】3页(P12719-12721)【关键词】线粒体DNA;二化螟;cox1基因;系统发育分析【作者】汪爱民;共桂云;魏兆军【作者单位】合肥工业大学生物与食品工程学院,安徽合肥230009;合肥工业大学生物与食品工程学院,安徽合肥230009;安徽建筑工业学院环境工程系,安徽合肥230601;合肥工业大学生物与食品工程学院,安徽合肥230009【正文语种】中文【中图分类】S433.4线粒体(Mitochondrion)是细胞中进行氧化磷酸化和脂肪酸以及某些蛋白质的生物合成的场所[1],并参与细胞的代谢、发育和衰老过程[2]。
应用新一代测序技术测定大室别藻苔虫线粒体基因组全序列

计 划 的基础 , 是 由于 它 测 序 通量 低 、 时费 力 , 但 费 科 学 家们 一直 在努 力寻 求通 量更 高 、 度更 快 、 格更 速 价
便宜的测序技术 。自 20 0 5年 以 来 , Roh 以 c e公 司 的 4 4技 术 、l mia 司 的 S lx 技 术 和 AB 公 5 Iu n 公 l oe a I
1 引 言
诞生于 2 O世 纪 7 0年代 的 S n e 法 是 最 早 被 a gr
广 泛使 用 的 D NA 测 序 技 术 , 是 完 成 人 类 基 因组 也
群 体遗传 分 化的理 想 材料L 。线 粒 体基 因组 的获 2 J
得 传 统上 通 常 是采 取 长 P R 结合 鸟 枪 法 或步 移 法 C 测 序 。虽然 P R方 法 操 作 简 单 , 样 品 需 求量 少 , C 对 但 是花 费时 间较 长 、 工作 量较 大 等问题 , 并且 利用 长 P R扩 增有 时成 功率不 高 , 以获得 目的片段 l 。 C 难 5 ] 苔 藓动 物 ( ro o n ) 为 重 要 的 海 洋 无 脊 椎 by za s 作
关键 词 : 室别 藻苔 虫 ; 粒体基 因组 ; 一代测 序技 术 ; 式 P R; 因排 列 大 线 新 巢 C 基
中 图 分 类 号 : 1. 1 Q9 5 8 5 文 献标 忘码 : A 文 章 编 号 :2 34 9 ( 0 2 0—1 60 0 5 -13 2 1) 20 3 —7
动物 门类 , 存 6 0 余 种 。 目前 国际上较 少 , 完 成 4个苔 藓 动 仅 物物 种 的线 粒体 全基 因组 , 包括 多室 草苔 虫 B g l u ua
n rt a 栉 口 目苔虫 Fls el r ipd 、 ei n 、 i u t l d ah s i a 颈链 血 r i 苔虫 Waes oas boq aa和扇 形管 孔苔 虫 T 一 tri r u tru t p “ b l oaf a elrs6 。大 室别 藻 苔 虫 Me r ui r lbl i¨ p a ] mba
【国家自然科学基金】_cox1基因_基金支持热词逐年推荐_【万方软件创新助手】_20140731
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 科研热词 线粒体基因组 长竹蛏 螽斯科 藻类 纺织娘 直翅目 日本纺织娘 序列分析 大竹蛏 upa dna条形码 cox1 推荐指数 2 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13
科研热词 鳗鲡 鳍足类 酵母双杂交 蛋白质编码基因 线粒体基因组 系统演化关系 系统发育 海豹科 海象科 海狮科 基因易位 cvb3 3a
推荐指数 3 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
科研热词 cox1基因 dna条形码 环介导等温扩增 浮游动物 检测 胶州湾 肌腱病 群落 细粒棘球绦虫 线粒体基因组 磷脂酶a2 环氧合酶 环境 海胆纲 海洋藻类 棘球绦虫 机械拉伸 序列分析 差异位点 四棘食道口线虫 分子标记 upa rbc l cox 1
推荐指数 5 3 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2012年 序号 1 2 3 4 5 6 7 8
科研热词 高磷血症 遗传标记 线粒体基因 甲状旁腺 环氧化酶2 核糖体基因 带绦虫 尿毒症
推荐指数 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12
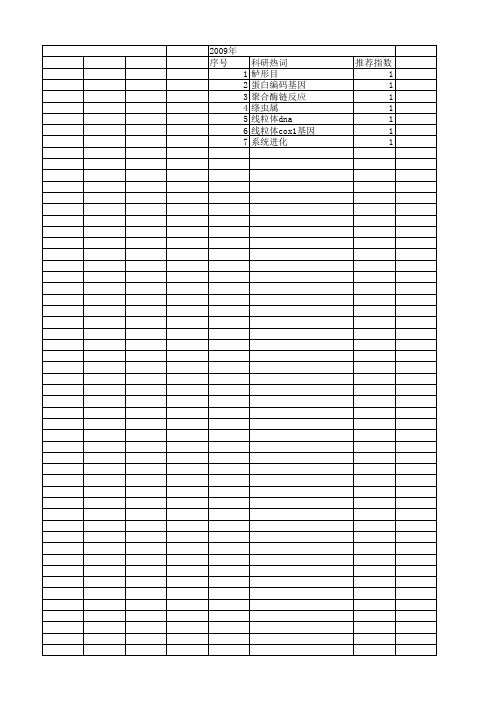
2009年 序号 1 2 3 4 5 6 7
科研热词 鲈形目 蛋白编码基因 聚合酶链反应 绦虫属 线粒体dna 线粒体cox1基因 系统2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31
青海省细粒棘球蚴线粒体cox1基因和nad1基因多态性的研究

1252
中国兽医科学
第 44 卷
G1isolates,5haplotypes occurred in the nad1gene with 5nucleotide mutation sites and 1to 4nucleotide differences or the highest variation rate of 0.45% ,whereas 9haplotypes existed with 27nucleotide muta- tion sites and 1to 11nucleotide differences or the highest variation rate of 0.68% for the cox1gene.Most mutations appeared in the third base of a codon,which hardly leads to the change of their encoded amino acids.Single nucleotide polymorphisms for cox1gene were more than those for the nad1gene,suggesting that the resolution ratio of the cox1gene should be relatively higher than that of the nad1gene in analysis of the nucleotide polymorphisms within E.granulosus G1isolates.In conclusion,the above-mentioned re- sults provide important data for further molecular epidemiological investigation of echinococcosis.
两种野生动物蛔虫线粒体cox1基因序列差异性研究

Chinese Journal of Veterinary Medicine
两 种 野 生 动 物 蛔 虫 线 粒 体 cox1 基 因 序 列 差异性研究
邱启官1 , 李 庆2ຫໍສະໝຸດ , 许英蕾2(1. 长沙生态动物园 , 湖南 长沙 410119 ; 2. 浙江农林大学动物科技学院 , 浙江 杭州 311300)
摘要:分离到湖南长沙生态动物园白狮和猎豹的两种野生动物蛔虫样本,对这两种蛔虫线粒体 DNA( rDNA) 中细胞色
素 I( cox1) 序列进行扩增测序,并与 GenBank 中收录的猫弓首蛔虫、狸猫弓首蛔虫、犬弓首蛔虫、狮弓蛔虫和猪蛔虫 cox1 序 列进行比较。 结果显示:来自长沙生态动物园的白狮和猎豹蛔虫 cox1 序列长度均为 415 bp;与 GenBank 收录 JF780951. 1 (狮弓蛔虫) 序列用 Blast 软件比较,发现极少数的碱基置换、颠换、缺失及无差异。 利用 DNAStar 5. 0 的 MegAlign 软件差异 性分析显示,白狮蛔虫( WL1、WL2) 基因序列和猎豹蛔虫( CH1、CH2) 基因序列与 JF780951. 1( 狮弓蛔虫) 序列的平均差异分 别为 4. 05% 和 2. 5% 。 而样品序列与其他序列相比差异均在 10. 1% 以上。 因此,这 2 种动物 4 个蛔虫样品种内差异小,种 间差异大,同是狮弓蛔虫。
Key words:white lion ; cheetah ; ascarid ; cox1 sequence ; gene diversity Corresponding author: XU Ying-lei
我国是濒危野生动物大国,野生动物不仅是生 态圈的一个重要部分,同时也是我们全人类的共同 的宝贵财富。 野生动物濒临灭绝大致可以分成两 个因素,第一个因素是我们人类在大自然中的部分 行为所引发的直接的或间接地结果;第二个因素是 野生动物自己本身的基因遗传, 疾病等问题造成 的,在疾病对野生动物的危害中寄生虫危害是其中
犬弓首蛔虫线粒体cox1基因的克隆及序列分析

关键词 1 基 因;种 系发 育关 系
中图分类号 :¥ 8 5 2 . 7 3 1 文献标识码:A 文章编号 :1 0 0 5 . 9 4 4 X( 2 0 1 5 )0 3 - 0 0 6 6 - 0 4
An a l ys i s o f Se qu e n c e Va r i a t i o n i n Mi t o c ho n dr i a l c o xl Ge ne
Ab s t r a c t :T h e o b j e c t i v e o f t h e s t u d y wa s t o a n a l y z e s e q u e n c e v a r i a t i o n i n t h e c y t o c h r o me C o x i d a s e s u b u n i t 1( c o x I )
wo r ms u s i n g he t c o x1 g e n e s e q u e n c i n g . T h e p a r t i a l c o x l( p c o x1 )we r e m p a l i i f e d ro f m i n d i v i d u a l c a n i s s a mp l e s ,
种 系发 育树 。本 实验扩 增所获得 的 p c o x 1 序 列长度 一致 ,均 为 3 9 4 b p ,种 内变异在 0 - 2 . 5 % 之 间,种 间差 异为
8 . 2 %- 1 1 . 6 %。种 系发育分析结 果表 明,1 2个犬 弓首蛔 虫分 离株位于 同一分 支。由于犬 弓首蛔 虫p c o x 1 序 列种 内 相对 保守,种 间差异较 大,故 可作 为种 间鉴定检 测研 究的遗传 标记 ,本研 究结果为犬 弓首蛔 虫的分类 、鉴 定和 群体 遗传 结构 奠定 了基础 。
人体带绦虫的分子鉴定

人体带绦虫的分子鉴定周灵贵;鞠伟;黄江;廖兴江;刘玉江;申萍香;戴佳琳【摘要】目的:对采自云南大理的6条人体带绦虫进行虫种分子鉴定.方法:对云南省大理有排节片史的病人以槟榔-南瓜子法驱虫,对6条人体带绦虫成虫进行形态学鉴定;提取虫体基因组DNA,用亚洲带绦虫和牛带绦虫线粒体cox1基因片段的特异性引物对DNA进行常规PCR扩增,扩增产物经琼脂糖凝胶电泳检测;将虫卵喂食2头幼猪(5×105/头),40d后解剖检查.观察囊尾蚴寄生情况.结果:6条人体带绦虫的形态和牛带绦虫相似,亚洲带绦虫cox1片段PCR为阳性,扩增产物约260 bp,牛带绦虫cox1片段PCR均为阴性;只在感染幼猪肝脏发现有囊尾蚴寄生,其他组织和肌肉内没有囊尾蚴寄生.结论:形态学和分子生物学鉴定6条人体带绦虫标本均属于亚洲带绦虫.【期刊名称】《贵阳医学院学报》【年(卷),期】2014(039)003【总页数】4页(P311-314)【关键词】亚洲带绦虫;牛带绦虫;形态学;分子生物学;鉴定【作者】周灵贵;鞠伟;黄江;廖兴江;刘玉江;申萍香;戴佳琳【作者单位】贵阳医学院多媒体形态学实验室,贵州贵阳550004;贵阳医学院法医学系,贵州贵阳550004;贵阳医学院法医学系,贵州贵阳550004;贵阳医学院物理学教研室,贵州贵阳550004;贵阳医学院多媒体形态学实验室,贵州贵阳550004;贵阳医学院多媒体形态学实验室,贵州贵阳550004;贵阳医学院法医学系,贵州贵阳550004【正文语种】中文【中图分类】R383.32猪带绦虫、牛带绦虫和亚洲带绦虫是寄生于人体引起肠绦虫病的3种带绦虫,在我国分布较普遍,有些地方呈局限性流行[1-2]。
云南大理是多民族聚集地区,当地居民沿袭吃“生皮”、烧烤,生食猪、牛及野生动物肉类的习惯,导致带绦虫病的流行,成为当地严重的公共卫生问题[3]。
2013年3月,在云南省大理白族自治州凤仪镇采集到6条带绦虫。
自然衰老大鼠脑线粒体和血小板膜外周型苯二氮 受体结合活性的变化

ss Th p cfcb n i g a s y o h a ia tv B n a o i tl H PK1 9 o me r n s p ro m e . S a c a d a ay i i. e s e i i d n s a ft e r d o c i e P Rs a t g n s 。 l i 1 5 t mb a e wa e f r d 1 c t h r n l ss
赵悦 , 楠 , 杨 纪超 , 博 , 萍 萍 孙 左
[ 要 ] 目 的 以 3月 龄 青 年 大 鼠 为 对 照 , 察 2 摘 观 4月 龄 自然 衰 老 大 鼠 大 脑 皮 层 线 粒 体 和 血 小 板 膜 外 周 型 二 氮 受 体
( Rs 结 合 数 及 亲 和 力 的改 变 。方 法 雄 性 S 大 鼠断 头 取 脑 , 差 速 离 心 法 提 取 大 脯 皮 层 线 粒 体 , 渗 溶 血 法 制 备 外 周 血 小 板 PB ) D 以 低
b an m io ho i n i p a ee e br ne n agng a s M e h ds r i t c dra a d n l t lt m m a i i r t . t o M ae Spr gu — l a e Dawl y at e e di d d nt 一 a d 4 m o h e r s w r vie i o 3 n 2 一 nt gr oup . A l a m a s we e s rfc d by de a t to nd h r i e e i m e i t l e ov d. M ioc ndra om p e s fom s s 1 ni l r ac iie c pia in a t e b ans w r m d a ey r m e t ho i 1c on nt r di s c e e e alc t r s a e e t d c r br orex we eiolt d. Them e b a e o a e e sf o v no oo s p e ar d by t e ho fby t i m ol — m r n fplt l t r m e usbl d wa r p e hem t d o po on che y
感染高原鼠兔的皮蝇蛆线粒体CO1基因序列分析

D YC Z 一 2 4 B型 电泳 槽 , 北 京 六 一 仪器 厂 产 品 ; My C y —
c l e r TM The r ma l Cy c l e r EN一 6 1 0 1 0 (BI O— RDA ,
畜 牧业 生产 造 成 了严 重 的经 济 损 失 , 其 中 以对 制 革
1 . 1 . 1 虫株 及 收 集 地 区 在 青 海 省 称 多 县 收 集 感
染 高原 鼠兔 的皮蝇 蛆 ( Hy p o d e r ma l a r v a e ) , 将 收 集 的新 鲜幼 虫用 7 0 0 mL / L酒精 固定 , 分别 编号 。
给草地 畜牧 业 带来 很 大 的经 济 损 失 , 而且 是 某 些 人 1 . 1 . 2 主 要 试 剂 DNA 提 取 用 T I AG E N 公 司 生
其 引起 的一 类 慢性 寄生 虫病 , 在 北 半 球 许 多 国 家是 危 害严 重 的动 物 寄 生虫 病 之 一 , 也是 重 要 的 人兽 共 患 寄 生虫病 。在我 国 , 本 病 主要分 布 于西 北 、 东 北 和 华 北地 区 , 其 中青 海 、 内蒙古 、 甘肃 、 新 疆 和 西藏 5 个
动 物 医学进 展 。 2 0 1 4 , 3 5 ( 3 ) : 3 3 - 3 5
Pr o g r e s s i n Ve t e r i na r y Me d i c i ne
感 染 高 原 鼠兔 的皮 蝇 蛆 线粒 体 C O1基 因序 列分 析
朵 红 , 付 永 , 沈 秀英 , 彭 毛 , 郭 志宏 , 李 伟
其 是 滩地 、 河岸 、 山麓 缓 坡 等植 被 低 矮 的开 阔生 境 , 喜 食牧 草 的芽 、 叶、 茎、 花、 种子及根 , 是 草 原上 一 种
胞嘧啶脱氨基酶APOBEC1研究进展

胞嘧啶脱氨基酶APOBEC1研究进展作者:周冰涵严小璇蓝文贤王春喜曹春阳来源:《上海师范大学学报·自然科学版》2020年第03期摘要:為全面理解载脂蛋白B mRNA(ApoB mRNA)编辑酶催化多肽-1(APOBEC1)的作用机制,介绍了APOBEC1和ApoB mRNA的蛋白及核酸序列,总结并绘制了APOBEC1与不同的辅助蛋白的结合模型,阐述了APOBEC1催化ApoB mRNA第6 666位的胞嘧啶(C6666)脱氨基化分子机制.列举了啮齿动物APOBEC1抑制多种逆转录病毒的研究报道,介绍了兔源APOBEC1结合人类免疫缺陷病毒1(HIV-1)的病毒粒子并编辑病毒基因组的机理.同时介绍了APOBEC1通过编辑胞嘧啶或与AU富集元件(ARE)结合来调控癌症等疾病相关的细胞因子表达.关键词:载脂蛋白B mRNA(ApoB mRNA); 载脂蛋白B mRNA编辑酶催化多肽-1(APOBEC1); 胞嘧啶脱氨基化中图分类号: Q-71 文献标志码: A 文章编号: 1000-5137(2020)02-0234-11Research advances on cytosine deaminase APOBEC1ZHOU Binghan1,2, YAN Xiaoxuan2, LAN Wenxian2, WANG Chunxi2, CAO Chunyang2*(1.College of Chemistry and Materials Science,Shanghai Normal University,Shanghai 200234,China; 2.State Key Lab of Bio-organic and Natural Products Chemistry,Shanghai Institute of Organic Chemistry,Chinese Academy of Sciences,Shanghai 200032,China)Abstract: In order to fully understand the mechanisms of Apolipoprotein B mRNA(ApoB mRNA) editing enzyme catalytic polypeptide-1(APOBEC1),this review introduced the amino acid and nucleic acid sequences of APOBEC1 and ApoB mRNA,summarized and mapped the binding models of APOBEC1 with different cofactors to explain the molecular mechanism of APOBEC1 catalyzing the deamination of the 6666 C of ApoB mRNA (C6666).The researches of rodent APOBEC1 inhibiting multiple retroviruses were exemplified here,and the related mechanisms of rabbit APOBEC1 binding to human immunodeficiency virus type 1(HIV-1)and editing the viral genome were discussed.This review also introduced APOBEC1 regulating the expression of cytokines related to cancers and other diseases by deamination editing or combining with AU-rich element(ARE) of RNAs.Key words: apolipoprotein B mRNA(ApoB mRNA); ApoB mRNA editing enzyme catalytic polypeptide-1(APOBEC1); cytidine deamination0 引言载脂蛋白B mRNA(ApoB mRNA)编辑酶催化多肽(APOBEC家族)是一类胞嘧啶脱氨基酶,能催化单链RNA或单链DNA中的胞嘧啶脱氨基转化为尿嘧啶.APOBEC家族由活化诱导胞嘧啶脱氨基酶(AID),ApoB mRNA编辑酶催化多肽-1(APOBEC1),APOBEC2,APOBEC3亚家族(APOBEC3A,APOBEC3B,APOBEC3C,APOBEC3D,APOBEC3E,APOBEC3F,APOBEC3G,APOBEC3H),以及APOBEC4组成.其中APOBEC1与AID串联排列于第12号染色体,APOBEC2位于第6号染色体,APOBEC3亚家族以串联重复的方式排列于第22号染色体[1],APOBEC4则位于第1号染色体[2],如图1(a)所示.APOBEC家族成员脱氨基催化活性由1个或2个锌指结构域提供,位于锌指结构域的氨基酸序列在APOBEC家族中相当保守:His-X-Glu-X23–28-Pro-Cys-X2-4-Cys(其中X表示任何氨基酸);AID,APOBEC1,APOBEC3A,APOBEC3C,APOBEC3H为单锌指催化结构域;APOBEC3B,APOBEC3D,APOBEC3F,APOBEC3G则含有2个锌指催化结构域,如图1(b)所示,而APOBEC2与APOBEC4暂无结构相关报道[2].APOBEC家族中研究最深入的是AID与APOBEC3亚家族,两者都有以DNA为底物的高效脱氨基催化活性,最广为人知的功能是在外源性病毒逆转录过程中对DNA进行编辑,使病毒DNA发生降解以抑制病毒逆转录过程,如人源APOBEC3G编辑人类免疫缺陷病毒1(HIV-1)DNA以抑制HIV-1在人体中的复制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脫氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1与辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],构成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N 端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未经编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512 000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的进攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,編辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脱氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1与辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],构成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N 端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA 的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未经编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512 000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的进攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脱氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1与辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],構成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N 端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA 的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未经编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512 000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的进攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脱氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1与辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],构成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA 的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未經编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的进攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脱氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1与辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],构成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N 端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA 的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未经编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512 000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的進攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动物,尤其兔源APOBEC1蛋白通过RNA/DNA胞嘧啶脱氨基化的机制,抑制某些逆转录病毒的复制;2)随着更多的APOBEC1编辑靶标的鉴定,发现APOBEC1在包括癌症等疾病发生方面具有一定作用;3)APOBEC1也是APOBEC家族中唯一需要与特定的辅助蛋白形成复合物才能进行ApoB mRNA编辑的蛋白[3-4].近年来关于APOBEC1的研究范围越来越广,不再局限于ApoB mRNA的脱氨基化研究.APOBEC1在体内有大量RNA靶标,催化脱氨基化也不是其参与生理过程的唯一机制.为全面了解APOBEC1功能,促进APOBEC1在相关领域的研究,本文作者总结了近年来APOBEC1在生物功能方面的研究进展,介绍了基于同源建模预测的APOBEC1编辑胞嘧啶脱氨基化的分子机制,APOBEC1與辅助蛋白如何形成复合物识别并编辑ApoB mRNA的机制,和APOBEC1在逆转录病毒以及疾病方面的研究成果.1 APOBEC1研究进展1.1 APOBEC1功能域及结构研究APOBEC1最初在ApoB mRNA编辑事件中被发现.人源APOBEC1与兔源APOBEC1包含236个氨基酸(aa),大鼠APOBEC1与小鼠APOBEC1包含229aa,人源APOBEC1与大鼠APOBEC1具有69%的序列相似性[5],构成锌指结构域的脱氨基活性位点H-X-E-X23-28-C-P-X2--C在APOBEC1同源蛋白中也十分保守[4];N端的碱性氨基酸R15,R16,R17,R33和K34被4认为是核定位信号的一部分[6-7],它们对编辑反应很重要[8].MEHTA等[9]发现,APOBEC1的N端区域可能参与辅助蛋白的结合.APOBEC1蛋白均在C端173~210 aa处具有一段保守的亮氨酸富集区域180~196 aa.L180,L182,I185和L189的单突变体,以及P190A/P191A双突变体均导致APOBEC1部分或几乎完全失去编辑活性[8].同位素标记以及高效液相色谱分析显示:APOBEC1可能以同源二聚体的方式存在[10],而APOBEC1的C端残基196~210 aa和221~229 aa对二聚体的形成有很重要的影响[8],如图2所示.其次C端缺失的APOBEC1突变体(APOBEC1截短体1~172 aa和截短体1~196 aa)无法二聚并且无法编辑ApoB mRNA[8,11],IKEDA等[5]发现APOBEC1的二聚结构需要RNA分子的介导.APOBEC1还能对单链DNA 的胞嘧啶进行脱氨基催化[12],基于酵母脱氨酶晶体结构模拟的APOBEC1结构模型支持这一结论[13].IKEDA等[5]发现兔源APOBEC1的C端亮氨酸富集区以及2个二聚体结构域均参与了其包装到HIV-1病毒粒子中的过程,C端结构域同时也是APOBEC1对病毒cDNA和基因组RNA发挥脱氨基活性必不可少的部分.1.2 APOBEC1催化胞嘧啶脱氨基化的可能机制APOBEC1能够特异性催化ApoB mRNA第6666位的胞嘧啶C6666脱氨基化转变为尿嘧啶(U6666),即ApoB mRNA C-to-U编辑,该处密码子则由C6666AA(Q2153)突变为终止密码子U6666AA,经过编辑的ApoB mRNA翻译后得到ApoB蛋白的截短体ApoB48(相对分子质量为241 000),未经编辑的ApoB mRNA则翻译为全长的ApoB100(相对分子质量为512000)[14],如图3(a)所示.ApoB100在血液中运输内源性胆固醇和甘油三酸酯,而截短体ApoB48可代谢膳食脂类[15],但ApoB100结合胆固醇并在血液中运输时有可能增加动脉粥样硬化的风险[16],所以APOBEC1对ApoB mRNA的脱氨基催化产物ApoB48可能降低动脉粥样硬化的风险.APOBEC1催化活性中心是一个锌指结构域,如图1(b)所示,脱氨基化活性位点为His-X-Glu-X23-28-Cys-Pro-X2-4-Cys[4],主要识别底物是RNA,也有研究报道APOBEC1能对DNA胞嘧啶催化脱氨基化[12,17].HARRIS等[1]根据细菌以及酵母胞嘧啶脱氨酶的结构研究预测了APOBEC蛋白对单链DNA脱氨基催化的分子机制,如图3(b)所示.首先,活性位点的组氨酸(His)和半胱氨酸(Cys)与锌离子(Zn2+)配位,此时一个水分子靠近活性位点;随后水分子在Zn2+作用下与谷氨酸(Glu)反应后生成一个氢氧根离子(OH-),激活了锌指结构域;激活后的Glu将胞嘧啶环的N3质子化,导致N3与C4双键不稳定,此时C4易于OH-的进攻;OH-进攻C4后其质子氢被Glu螯合,形成四面体的过渡态;最终,胞嘧啶的氨基侧基(-NH2)接受了被Glu螯合的质子氢,使碳氮键断裂,C4重新与氧原子(O)形成双键并从活性位点处释放尿嘧啶和氨(NH3),如图3(b)所示.APOBEC1催化DNA或RNA胞嘧啶脱氨基化可能也符合这一机制.APOBEC1是一种RNA胞嘧啶脱氨基酶,可特异性编辑ApoB mRNA,编辑DNA不是其主要功能[1].其功能特征具体体现在如下几个方面:1)与AID/APOBEC3相似的是,啮齿动。
线虫线粒体基因组全序列分析研究进展

线虫线粒体基因组全序列分析研究进展贾万忠;闫鸿斌;倪兴维;曹平;娄忠子;付宝权;史万贵【期刊名称】《中国农业科学》【年(卷),期】2011(044)006【摘要】线虫(nematode)种类繁多,生活方式多样,一部分线虫可寄生于动物和植物体内,引起线虫病(nematodiasis),其中旋毛虫病、猪蛔虫病等是重要的人兽共患寄生虫病,在中国和世界各地普遍流行,危害严重.本文将对线虫线粒体基因组的研究进展、应用和今后发展方向做一简要综述.迄今,已完成46种线虫的线粒体基因组全序列测定和分析.线虫线粒体基因组的碱基组成、基因结构、基因变异等方面有其特点,这些分析结果为线形动物门线粒体功能基因组学研究、比较基因组学研究、分子分类学研究、虫种(株)鉴定与分类、分子系统发育和进化分析等提供了重要依据和指导作用,为线虫病诊断、分子流行病学调查等分子检测方法的建立提供参考依据.【总页数】11页(P1255-1265)【作者】贾万忠;闫鸿斌;倪兴维;曹平;娄忠子;付宝权;史万贵【作者单位】中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室/农业部兽医公共卫生重点实验室/甘肃省动物寄生虫病重点实验室,兰州,730046;甘肃省动物疫病控制中心,兰州,730046【正文语种】中文【相关文献】1.池蝶蚌线粒体基因组全序列分析 [J], 盛军庆;林巧惠;王军花;彭扣;洪一江2.塔里木兔线粒体基因组全序列分析与系统进化研究 [J], 黄娅琳;黄捷;时玉;徐燕红;周用武;侯森林3.塔里木兔线粒体基因组全序列分析与系统进化研究 [J], 黄娅琳;黄捷;时玉;徐燕红;周用武;侯森林4.哈氏蜈蚣线粒体基因组全序列分析 [J], 孙丽冉;齐迎菊;田晓轩5.药用动物乌梢蛇线粒体基因组全序列分析 [J], 刘杰;田晓轩;崔英;朱彦因版权原因,仅展示原文概要,查看原文内容请购买。
猪囊尾蚴西昌分离株线粒体Cytb和nad4基因的序列测定与种系发育分析

猪囊尾蚴西昌分离株线粒体Cytb和nad4基因的序列测定与种系发育分析郝桂英【摘要】利用PCR技术对2个猪囊尾蚴西昌分离株的线粒体细胞色素b(Cytb)基因全序列和烟酰胺腺嘌呤二核苷酸脱氢酶亚单位4(nad4)基因部分序列(pnad4)进行扩增,分析其遗传变异,并用MEGA 5.0程序NJ法绘制种系发育树,探讨不同地区来源的猪囊尾蚴种系发育关系.测序结果显示,2个猪囊尾蚴分离株的Cytb基因全序列长度均为1 068 bp,nad4基因部分序列长度均为815 bp,核苷酸序列同源性均为99.8%.种系发育分析结果显示,所有猪囊尾蚴分离株形成一个分支,2个西昌分离株均属于猪带绦虫亚洲基因型.结果表明,Cytb和nad4基因均可用于猪带绦虫的分子分类,为猪带绦虫病/猪囊尾蚴病的分子诊断奠定基础.【期刊名称】《动物医学进展》【年(卷),期】2015(036)005【总页数】5页(P59-63)【关键词】猪囊尾蚴;Cytb基因;nad4基因;种系发育【作者】郝桂英【作者单位】西昌学院,四川西昌615013【正文语种】中文【中图分类】S852.734;S858.28猪囊尾蚴(Cysticercus cellulosae)是猪带绦虫(Taenia solium)的中绦期,寄生于猪、人等的肌肉、脑、眼等处,引起囊尾蚴病[1]。
该病呈世界性分布,是一种危害严重的人兽共患寄生虫病,在中国、印度、印尼、泰国、老挝、柬埔寨、尼泊尔、菲律宾、缅甸、越南、韩国、肯尼亚、坦桑尼亚、墨西哥等国家广泛流行[2-3]。
我国东北、西北、东部和南部等地感染率均较高[4]。
猪带绦虫病/猪囊尾蚴病仍然是全世界不可忽视的公共卫生问题[5]。
猪带绦虫及其中绦期在形态上与其他2种带绦虫(牛带绦虫、亚洲带绦虫)很难区分,同时由于受自然、地理等生态环境因素的长期影响,寄生虫种、株间发生不同程度的遗传分化,这些变化很难用形态学方法所区分。
DNA序列分析比形态学方法具有更强的鉴别力,可精确测量生物自然种群的遗传变异程度,是目前进行物种遗传变异、分子分类和分子种系发生等研究的主要手段之一,为寄生虫的准确鉴定和系统学研究提供了有力的工具,弥补了传统分类方法的不足。
基于线粒体COI基因序列的壮真蝎与普洱真蝎的分子鉴定

基于线粒体COI基因序列的壮真蝎与普洱真蝎的分子鉴定李蓉;郑雨田;李春青;陈艳艳;杨振升;陈善元;肖蘅【摘要】对蝎类物种的传统分类主要依靠形态和行为特征,但由于该类群种间形态特征极为相似,物种的划分和鉴定困难.为弥补传统分类方法的不足,本研究以线粒体细胞色素氧化酶亚基Ⅰ (COI)基因作为分子标记,对形态相似的壮真蝎Euscorpiops validus和普洱真蝎E.puerensis进行分子水平的物种鉴定.采用PCR 扩增测序获得壮真蝎与普洱真蝎共24个样本的COI基因部分片段序列(660 bp),进行了遗传距离、系统发育及单倍型网络图分析.结果显示:壮真蝎15个样本中共检测到4个单倍型,单倍型之间的相似度为99.3%~99.8%;普洱真蝎9个样本中共检测到4个单倍型,单倍型之间的相似度为99.6%~99.8%;2种蝎的种间序列相似度为90.1%~90.6%,单倍型间的稳定差异核苷酸位点数为61个.壮真蝎与普洱真蝎种内平均遗传距离分别为0.0040、0.002 3,种间平均遗传距离为0.103 9,且种间遗传距离为种内的34.6倍.此外,分子系统发育树显示壮真蝎与普洱真蝎的单倍型序列各自聚为2个单系枝,且具有很高的分枝自举值(100%).单倍型网络图结果也显示壮真蝎与普洱真蝎8个单倍型明显分为2大类群,且壮真蝎的单倍型HAP2与普洱真蝎的单倍型HAP7之间的突变步数高达62步.上述结果不仅进一步确认壮真蝎与普洱真蝎为2个不同的物种,且表明线粒体COI基因可用于开展真蝎属Euscorpiops物种的分子鉴定.%The traditional classification and taxonomy of scorpiones are mainly based on morphological and behavioral characteristics.However,due to the similar morphological characteristics among species,it is difficult to classify and identify distinct species among scorpions.To compensate for the insufficiency of traditional taxonomic methodology,this study used mitochondrial cytochrome oxidase subunit I(COI) gene as molecular marker to conduct molecular identification of 2 morphologically similar scorpion species Euscorpiops validus andE.puerensis.The partial COI gene sequences (660 bp) from 24 samples of E.validus and E.puerensis were amplified by PCR followed by gene sequencing.The genetic distances,phylogenetic and haplotype network analyses were then carried out.The results showed that:4 haplotypes with similarity of 99.3%-99.8% were detected in 15 individuals of E.validus,and4 haplotypes with similarity of 99.6%-99.8% were found in 9 individuals ofE.puerensis;the interspecific similarity was 90.1%-90.6% and the number of stable differentiated nucleotide sites between the haplotypes of 2 species was 61.The intraspecific average genetic distances among E.validus haplotypes and among E.puerensis haplotypes were 0.004 0 and 0.0023,respectively,while the interspecific average genetic distance between E.validus and E.puerensis was O.103 9,being 34.6 times higher than that of intraspecific values.In addition,molecular phylogenetic tree clearly showed that the haplotype sequences of E.validus and E.puerensis clustered as 2 reciprocally monophyletic clades with high bootstrap values (100%).The haplotype network also showed that 8 haplotypes of E.validus andE.puerensis can be clearly divided into 2 clades,and the mutation steps between HAP2 haplotype of E.validus and HAP7 haplotype of E.puerensis reached 62.These results further confirmed that E.validus and E.puerensis were 2 distinct species,and indicated that mitochondrial COI gene was suitable for molecular species identification of Euscorpiops species.【期刊名称】《四川动物》【年(卷),期】2017(036)002【总页数】6页(P139-144)【关键词】壮真蝎;普洱真蝎;COI基因;分子鉴定【作者】李蓉;郑雨田;李春青;陈艳艳;杨振升;陈善元;肖蘅【作者单位】云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091;云南大学生命科学学院,昆明650091【正文语种】中文【中图分类】Q953;Q38壮真蝎Euscorpiops validus和普洱真蝎E. puerensis均属节肢动物门Arthropoda螯肢动物亚门Chelicerata蛛形纲Arachnida蝎目Scorpiones真蝎科Euscorpiidae真蝎属Euscorpiops,为真蝎属中等大小的物种(Kovaík et al.,2015)。
基于线粒体COI基因的17种菱蜡蝉亚科昆虫DNA条形码研究(半翅目:蜡蝉总科:菱蜡蝉科)

基于线粒体COI基因的17种菱蜡蝉亚科昆虫DNA条形码研究(半翅目:蜡蝉总科:菱蜡蝉科)肖永刚;陈祥盛【摘要】通过对菱蜡蝉科Cixiidae菱蜡蝉亚科Cixiinae 9属17种昆虫的mt DNA COI基因序列进行研究,探讨DNA条形码在菱蜡蝉亚科昆虫中快速识别和准确鉴定的可行性.采用MEGA 5对序列进行比对和遗传距离分析,基于COI基因序列构建ML、MP、NJ、ME系统发育树.结果显示:属间平均遗传距离为0.133,介于0.102 ~0.147之间;属内种间平均遗传距离为0.047,介于0.025~ 0.079之间;地理种群间平均遗传距离为0.039;介于0.024~0.064之间.系统发育树显示:同属物种聚为一小支,分支置信度高达97%~ 100%;同一地理类群聚为一支,分支置信度高达98%~100%.结果表明应用基于COI基因片段的DNA条形码对菱蜡蝉亚科昆虫分类鉴定是可行的.【期刊名称】《山地农业生物学报》【年(卷),期】2014(033)002【总页数】7页(P44-50)【关键词】菱蜡蝉亚科;DNA条形码;mt COI基因;遗传距离;系统发育树【作者】肖永刚;陈祥盛【作者单位】贵州大学昆虫研究所,贵州贵阳550025;贵州昆虫资源开发利用省级特色重点实验室,贵州贵阳550025;宜宾学院实验与教学资源管理中心,四川宜宾645300;贵州大学昆虫研究所,贵州贵阳550025;贵州昆虫资源开发利用省级特色重点实验室,贵州贵阳550025;贵州大学动物科学学院,贵州贵阳550025【正文语种】中文【中图分类】Q969.35线粒体细胞色素氧化酶第一亚基DNA(mt DNA)被认为是动物界中最适合的DNA 条形码标准基因[1-2]。
COI基因包含丰富的遗传信息,既相对保守又能保证足够变异,进化速率适中,可作为属种系统进化研究的良好标记,目前已广泛应用于昆虫分子系统发育分析[3-6]。
DNA条码技术已为研究和利用地球上众多的生物资源,鉴定生物多样性提供了强大的工具,并在各物种的鉴定与分类中发挥重要作用[5-8]。
用cox1基因片段的PCR鉴别亚洲带绦虫和牛带绦虫

用cox1基因片段的PCR鉴别亚洲带绦虫和牛带绦虫陈峥宏;郎书源;吴晓娟;包怀恩【期刊名称】《贵阳医学院学报》【年(卷),期】2009(034)003【摘要】目的:用cox1基因片段的PCR技术对亚洲带绦虫和牛带绦虫的快速鉴别.方法: 用亚洲带绦虫和牛带绦虫线粒体cox1基因片段的特异性引物以及带绦虫cox1基因片段的通用引物,对贵州省都匀地区和从江地区的带绦虫虫卵和节片DNA进行常规PCR扩增,扩增产物经琼脂糖凝胶电泳检测和核酸序列测定,并用NCBI Blast 对扩增产物的核酸序列与NCBI 数据库中带绦虫的cox1基因进行比对.结果: 5株都匀带绦虫DNA样品经亚洲带绦虫cox1基因片段的特异性引物PCR,均扩增出约260 bp的产物,PCR产物的核酸序列与NCBI 数据库中亚洲带绦虫线粒体cox1基因片段的同源性为100%;2株从江带绦虫的DNA 样品用亚洲带绦虫特异性引物和牛带绦虫特异性引物的PCR均未见扩增产物,而通用引物PCR则扩增出约1 000 bp的产物,核酸序列与NCBI 数据库中牛带绦虫线粒体cox1基因片段的同源性为99%,在牛带绦虫特异性引物的结合位点存在一个碱基差异.结论: 常规PCR扩增特异性的cox1基因片段可快速鉴定亚洲带绦虫;从江牛带绦虫株的cox1基因特异性片段部分存在变异,导致与特异性引物结合能力的差异,可采用cox1基因通用引物PCR结合测序进行鉴定.【总页数】4页(P239-241,261)【作者】陈峥宏;郎书源;吴晓娟;包怀恩【作者单位】贵阳医学院,微生物学教研室,贵州,贵阳,550004;贵阳医学院,寄生虫学教研室,贵州,贵阳,550004;贵阳医学院,微生物学教研室,贵州,贵阳,550004;贵阳医学院,寄生虫学教研室,贵州,贵阳,550004【正文语种】中文【中图分类】Q789;R383.32【相关文献】1.南瓜子槟榔驱除亚洲绦虫、猪带绦虫和牛带绦虫的效果比较 [J], 李树林;杨益超;谢祖英;吴钦华;区方奇;许洪波;商少明;黄铿凌;麦富珍2.传统方法和改进的PCR-RFLP鉴别猪带绦虫和牛带绦虫的比较研究 [J], 季旻珺3.兰州兽医所成功解析牛带绦虫和亚洲带绦虫基因组序列 [J], ;4.猪带绦虫和牛带绦虫卵的超微结构鄄观察 [J], 田喜凤;张运鹏5.亚洲带绦虫和牛带绦虫实验感染乳猪后肝脏细胞凋亡的研究 [J], 牟荣;包怀恩;张科;吴家红;郎书源因版权原因,仅展示原文概要,查看原文内容请购买。
基于cox1基因的棕熊锡兰钩虫分子鉴定与种系发育分析

基于cox1基因的棕熊锡兰钩虫分子鉴定与种系发育分析贺名叶;谢旖【期刊名称】《中国动物传染病学报》【年(卷),期】2022(30)3【摘要】本次研究旨在利用线粒体细胞色素I(cox1)部分基因(pcox1)对长沙市动物园棕熊粪便分离的钩虫进行种类鉴定,并分析其基因多态性。
用PCR技术扩增从棕熊粪便分离的钩虫长沙分离株的pcox1基因,并对其基因序列进行分析。
结果显示7条钩虫分离株的pcox1序列片段大小均为423 bp,种内遗传差异为0.0%~0.5%,与GenBank中收录的锡兰钩虫(Ancylostoma ceylanicum)分离株同源性为93.8%~94.8%,与其他钩虫(犬钩虫、巴西钩虫、十二指肠钩虫、猫钩虫和美洲钩虫等)序列相似性低于88.7%;系统发育分析显示,本次研究获得的棕熊钩虫分离株与已知锡兰钩虫聚为一支,与其他钩虫相隔较远。
结果表明,长沙市动物园棕熊粪便分离的钩虫为锡兰钩虫,线粒体pcox1基因在锡兰钩虫种内相对保守,与其他钩虫种类序列差异较大,可以作为熊锡兰钩虫分子鉴定及遗传变异研究。
【总页数】5页(P196-200)【作者】贺名叶;谢旖【作者单位】湖南省益阳职业技术学院【正文语种】中文【中图分类】S852.7【相关文献】1.基于16S rRNA基因的鼠Mycoplasma haemomuris的分子鉴定和种系发育分析2.基于线粒体cox1和nad1基因对青海牦牛种系发育关系的研究3.基于cox1基因序列分析山羊褐黄血蜱种系发育关系4.黑龙江省不同地区东方次睾吸虫线粒体cox1和nad1基因序列分析及种系发育研究5.福建省蛇源蛔虫cox1基因扩增与种系遗传发育分析因版权原因,仅展示原文概要,查看原文内容请购买。
基于线粒体COⅠ~COⅡ序列的黑带食蚜蝇(Episyrphus balteatus)遗传多样性初探

基于线粒体COⅠ~COⅡ序列的黑带食蚜蝇(Episyrphusbalteatus)遗传多样性初探张卓;胡建楠;王晓昕;周婵;刘广纯【摘要】利用COⅠ的通用引物,采用PCR法对黑带食蚜蝇(Episyrphus balteatus)线粒体DNA序列进行扩增、测序及比对;4个不同地区的种群分析结果表明:①共获得1041个bp的DNA序列,A、T、G、C的比例分别为40.2%、30.1%、13.1%、16.6%,表现出明显的A+T偏倚性;②经过与GENEBANK已知序列比对,该片断位于线粒体的COⅠ~tRNALEU~COⅡ区域;③共检测到8处(N=20)碱基变异,已经产生了遗传差异,形成1个共有单倍型、7个特有单倍型;④NETWORK分析表明,已经产生明显的遗传分化.【期刊名称】《沈阳大学学报》【年(卷),期】2015(027)002【总页数】4页(P116-119)【关键词】黑带食蚜蝇;遗传多样性;COⅠ~COⅡ;种群;生物地理【作者】张卓;胡建楠;王晓昕;周婵;刘广纯【作者单位】沈阳大学城市有害生物治理与生态安全重点实验室,辽宁沈阳110044;沈阳大学城市有害生物治理与生态安全重点实验室,辽宁沈阳 110044;沈阳大学城市有害生物治理与生态安全重点实验室,辽宁沈阳 110044;辽宁大学生命科学院,辽宁沈阳 110036;沈阳大学城市有害生物治理与生态安全重点实验室,辽宁沈阳 110044【正文语种】中文【中图分类】Q95黑带食蚜蝇(Episyrphus balteatus)隶属于双翅目(Diptera)、环裂亚目(Cyclorrhapha)、无缝组(Aschiza),蚜蝇科(Syrphidae).蚜蝇科又称为食蚜蝇科,是双翅目中较大的一个类群,全世界约230属,6 000余种[1].黑带食蚜蝇是蚜蝇中较为常见的一个种类,其对蚜虫的捕食力极强,研究表明黑带食蚜蝇整个幼虫期可捕食棉蚜(Aphisgossypii Glover) 840~1 500头,是蚜虫的重要天敌[2];其次,黑带食蚜蝇成虫是仅次于蜜蜂的重要授粉昆虫,具有访花的习性,对个别植物而言,其作用大于蜜蜂.国外曾报道在欧芹(Petroselinum crispum)传粉研究中,食蚜蝇传粉种子产量是蜜蜂的近3倍(蜜蜂传粉地块的种子产量为617 kg/hm2,蚜蝇传粉地块的种子产量为1 620 kg/hm2)[3].线粒体是动物细胞核外唯一具有DNA的细胞器,并与细胞核进行一定的遗传物质交换,又保持相对独立的遗传体系,具有结构简单、进化速度快、母系遗传、基因重组率小等特点,其复制、转录和翻译在线粒体内,受自身和核基因的双重控制等特点[4-6].线粒体DNA被广泛用于种群遗传多样性的检测和研究当中去[7-10].国内外对于黑带食蚜蝇的研究仅停留在野外识别[11-12]、新种描述[13]、新记录种分布[14]和物种修订[15]等方面,对于部分种类基于形态和分子数据进行了少量研究[16],而对于黑带食蚜蝇的遗传多样性研究还未见涉及.本研究以黑色食蚜蝇线粒体DNA中的COⅠ~COⅡ片断为目的片断,进行遗传变异检测,探讨该片断在黑带食蚜蝇种群遗传差异检测的可行性和有效性,为黑带食蚜蝇的遗传多样性分析、生物地理演化和分子钟假设检验等相关问题奠定前期实验基础.本实验有利于对黑带食蚜蝇的认知,并为今后进行生物利用和生物防治做好准备. 样品采集地点(见表1).采用网捕法进行食蚜蝇的采集.收集到的样本放在无水乙醇中,-20 ℃冷冻保存至DNA提取前.Agilent 8800梯度PCR扩增仪,STR16R小型台式高速冷冻离心机,海尔冰箱、KD-S24恒温水浴锅、BIO-RAD水平电泳仪、BIO-RAD GelDoc 2000凝胶成像系统、FA2004N电子分析天平、北京六一高压灭菌锅.TaqDNA聚合酶、dNTPs,10×Reaction buffer缓冲液,Mg2+、6×Loading buffer均购于鼎国科技有限公司;1000 bp DNA Mark和蛋白酶K购于上海生工;0.5×TBE电泳缓冲液、琼脂糖、EB, SDS,NaCl、Tris-饱和酚、氯仿、异戊醇、异丙醇和无水乙醇均为国产分析纯.引物由上海生工合成,DNA序列测定由鼎国科技有限公司北京总部测试完成.1.4.1 全基因组提取剪取黑带食蚜蝇的3条腿,置于灭菌的2.0 mL离心管中;总量DNA采用常规酚—氯仿法进行,无水乙醇脱水晾干后,溶于100 μL的TE缓冲液中,抽取5 μL进行电泳检测,其余基因组样品放入-20 ℃冰箱保存到PCR前.1.4.2 引物选择及目标片段PCR扩增根据双翅目COI的通用引物[17]选择LCO1490~HCO2198及LCO1490~HCO3014形成两对嵌套的引物对,引物序列如下:LCO1490(5′-GGTCAACAAATCATAAAGATATTGG-3′)、HCO2198(5′-TAAACTTCAGG GTGACCAAAAAATCA-3′)和HCO3014(5′-TCCAATGCACTAATCTGCCATATTA-3′).PCR(聚合酶链式反应)体系如下:10×buffer—5 μL,Mg2+(25 mmol)—1.8μL,dNTP(2.5 mmol)—1 μL,上游引物(LCO1490,15 mmol)—1 μL,下游引物(HCO2198或HCO3014,15 mmol)—1 μL,Taq DNA聚合酶1 μL(2U/μL)—1 μL,模板DNA—3 μL,灭菌双蒸水补足至50 μL.PCR扩增反应程序分为如下三个阶段:①预变性阶段,94 ℃,7 min;②扩增阶段,94 ℃变性60 s,55~59 ℃复性30 s,72 ℃延伸60 s,共运行35~40个循环;③后延伸阶段,72 ℃延伸7 min.PCR产物用1.5%琼脂糖凝胶进行电泳检测(见图1).PCR产物送鼎国科技有限公司北京总部进行测定.1.4.3 DNA序列测定与比对采用SEQUENCE 4.0.5软件对序列测定结果进行人工较对;利用GENEBANK中的BLAST功能进行序列比对与分析;采用Clastal W 1.83软件进行DNA序列对齐;采用NETWORK 4.6.1.2软件进行网络进化分析;采用MEGA 6.06软件进行系统发育进化树的构建.(1) PCR产物经ABI377进行序列测定后,经SEQUENCE 4.0.5软件进行两端DNA 序列拼接及人工较对,共获得1 041个bp的基因片断,经过与GENEBANK已经序列比对,该片断位于线粒体的COⅠ~tRNALEU~COⅡ区域;(2) 对1 041个bp DNA碱基序列分析表明: A,T,G,C比例分别为40.2%,30.1%,13.1%,16.6%,A+T=70.3%,表现出明显的A+T偏倚性;(3) 定义了8个单倍型(N=20), 包括7个特有单倍型和1个共有单倍型[包括13个个体(其中SYBN 3个、SXFY 5个、GSLX 2个、BXYHG 3个)];(4) 与共有单倍型相比较,检测到8处碱基变异(n=20),分别处于第4 bp、26 bp、65 bp、148 bp、166 bp、290 bp、899 bp和1 008 bp处;(5) 在COⅠ~tRNALEU~COⅡ基因片断上,黑带食蚜蝇20个个体中有7个产生了突变,其比例达到35%(7/20),已经产生了明显的分化.因此,这一区段的突变比例达到检测种群内遗传多样性的要求.在NETWORK 4.6.1.2软件中,以Median Joining法进行网络进化分析,结果表明,7个特有单倍型通过1到2步突变与共有单倍型聚在一起(见图2).已经形成明显的网格状分化.但并没有形成明显的地理分化.利用MAGE 6.06软件进行UPGMA、NJ、ME、ML和MP树的构建,获得类似的拓扑结构树(以UPGMA树为例),如图3所示.结果表明,本研究的4个样点,20个个体的样本库并没有形成明显的地理结构,这与取样个体过少有关,当增大取样量后有可能会形成一个明显的地理谱系.分析的片断位于线粒体的COⅠ~tRNALEU~COⅡ区域,A+T达70.3%,表现出明显的A+T偏倚性,符合昆虫纲线粒体DNA的总体规律;网络进化分析表明,已经产生明显的地理分化.因此,以本研究中的两对嵌套引物,对黑带食蚜蝇线粒体COⅠ~tRNALEU~COⅡ区域进行PCR扩增并进行DNA序列的测定、比对,适于进行黑带食蚜蝇的种群遗传多样性研究.本研究为黑带食蚜蝇的种群线粒体DNA遗传多样性研究奠定了前期理论基础.【相关文献】[ 1 ] 霍科科,任国栋,郑哲民. 秦巴山区蚜蝇区系分类[M]. 北京:中国农业科学技术出版社, 2007:146-150.(Huo Keke, Ren Guodong, Zheng Zhemin. Fauna of Syrphidae from Mt. Qinline-Bashan in China (Insecta: Diptera)[M]. Beijing: China’s Agricultural Science and Technology Press, 2007:146-150.)[ 2 ] 曾强,陈乾锦,何榕宾,等. 黑带食蚜蝇对烟蚜的捕食功能反应及寻找效应[J]. 武夷科学, 2003,19(1):274-278.(Zeng Qiang,Chen Qianjin,He Rongbin,et al. The Predatory Functional Response and Searching Efficiency of Episyrphus balteatus (Degeer) on Myzus persicae (Sulzer)[J]. Wuyi Science Journal, 2003,19(1):274-278.)[ 3 ] 浦子钢,迟莉. 食蚜蝇(Syrphidae)生物学特性及人工室内饲养技术的研究进展[J]. 黑龙江农业科学, 2006(5):120-123.(Pu Zigang,Chi Li. Study on Biological Characters of Syrphidae and the Advances of Manpower Rearing Technique Indoor[J]. Heilongjiang Agricultural Sciences, 2006(5):120-123.)[ 4 ] 殷玲,吉挺,陈晶,等. 蜜蜂线粒体DNA在遗传分析中的应用[J]. 昆虫知识, 2008,45(5):708-712. (Yin Ling,Ji Ting,Chen Jing, et al. Applications of mtDNA in Honeybee Genetic Analysis[J]. Chinese Bulletin of Entomology, 2008,45(5):708-712.)[ 5 ] Capps G J,Samuels D C,Chinnery P F.A Model of the Nuclear Control of Mitochondrial DNA Replication[J]. Journal of Theoretical Biology, 2003,221(4):565-583. [ 6 ] 郭松长,祁得林,陈桂华,等. 家牦牛线粒体DNA(mtDNA)遗传多样性及其分类[J]. 生态学报, 2008,28(9):4286-4294.(Guo Songchang, Qi Delin, Chen Guihua, et al. Genetic Diversity and Classification of Domestic Yaks Inferred from Mitochondrial DNA Sequences[J]. Acta Ecologica Sinica, 2008,28(9):4286-4294.)[ 7 ] 徐国威,周丹银,刘意秋,等. 基于线粒体DNA(mtDNA)COⅠ~COⅡ分析四川省东方蜜蜂遗传多样性[J]. 云南农业大学学报, 2014,29(1):63-70.(Xu Guowei,Zhou Danyin,Liu Yiqiu,et al. Genetic Diversity of Apis cerana in Sichuan Province Based on Mitochondrial DNA Sequence COI~COⅡ[J]. Journal of Yunnan Agricultural University, 2014,29(1):63-70.)[ 8 ] 管成林,李继姬,郭宝英,等. 基于线粒体COⅠ基因比较分析野生与养殖厚壳贻贝种群的遗传多样性[J]. 安徽农业科学, 2012(13):7660-7662.(Guan Chenglin, Li Jiji, Guo Baoying, et al. Genetic Diversity Evaluation by the Comparative Analysis on Mitochondrial COⅠ Gene between Wild and Cultured Populations of Mytilus coruscus[J]. Journal of Anhui Agricultural University, 2012(13):7660-662.)[ 9 ] 屠云洁,高玉时,周新民,等.我国6个地方鸡品种线粒体COⅠ基因遗传多样性分析[J]. 扬州大学学报:农业与生命科学版, 2007,28(3):31-33.(Tu Yunjie,Gao Yushi,Zhou Xinmin,et al. The Genetic Diversity Analysis of mtDNA COⅠ Genes in Six Indigenous Chicken Breeds in China[J]. Journal of Yangzhou University: Agricultural and Life Science Edition, 2007,28(3):31-33.)[10] 姜石生. 基于线粒体DNA的16S rRNA、COⅠ和COⅡ基因的黄脊竹蝗5地理种群遗传多样性研究[D]. 长沙:中南林业科技大学, 2011:1-75.(Jiang Shisheng. Study on Genetic Diversity of Rammeacris kiangsu of 5 Geographical Populations Based on Partial Sequences of 1 6S rRNA Gene,COⅠ Gene,and COⅡ Gene[D]. Changsha: Central South University of Forestry and Technology, 2011:1-75.)[11] 霍科科,任国栋. 陕西秦岭太白山北坡食蚜蝇科昆虫区系调查[J]. 昆虫知识, 2006,43(5):700-705.(Huo Keke,Ren Guodong. Faunal Study on Syrphids of the North Slope of Taibai,Qinling Mountains of Shaanxi[J]. Chinese Bulletin of Entomology, 2006,43(5):700-705.)[12] 高欣,张春田. 辽宁食蚜蝇科昆虫名录(双翅目:食蚜蝇科)[J]. 四川动物, 2006,25(1):114-115,127. (Gao Xin, Zhang Chuntian. Note on Ayrphidae from Liaonin Province (Diptera: Syrphidae)[J]. Sichuan Journal of Zoology, 2006,25(1):114-115,127.)[13] 白素芬,张国威,李欣,等. 河南省食蚜蝇科3个新记录属和12个新记录种[J]. 河南农业科学, 2011,40(7):86-89.(Bai Sufen,Zhang Guowei,Li Xin,et al. Three Genera and Twelve Species of Syrphid F1ies(Diptera:Syrphidae) Newly Recorded from Henan Province[J]. Journal of Henan Agricultural Sciences, 2011,40(7):86-89.)[14] 陈玉宝,高文韬,张义涛,等. 食蚜蝇科中国一新纪录种(双翅目:食蚜蝇科)[J]. 北华大学学报:自然科学版, 2014,15(2):238-239.(Chen Yubao, Gao Wentao, Zhang Yitao, et al. A New Record Species of Hoverfly in China (Diptera: Syrphidae)[J]. Journal of Beihua University (Natural Science), 2014,15(2):238-239.)[15] 张宏杰,霍科科. 食蚜蝇科的分类系统及其研究进展[J]. 昆虫知识, 2005,42(2):132-138. (Zhang Hongjie, Huo Keke. The Classification Systems of Syrphidae and Advance in the Related Researches[J]. Chinese Bulletin of Entomology, 2005,42(2):132-138.)[16] 成新跃,吕静,黄春梅,等. 基于形态和分子数据确定缩颜蚜蝇族的系统分类地位[J]. 中国科学:C辑, 1999,29(6):645-654.(Cheng Xinyue, Lyu Jing, Huang Chunmei. System Classification Status of Pipizini Family Based on Morphological and Molecular Data[J]. Science in China:Series C, 1999,29(6):645-654.)[17] Folmer O, Black M, Hoen W, et al. DNA Primers for a Mplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates[J]. Molecular Marine Biology and Biotechnology, 1994,3(5):294-299.。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
引物 的核 苷酸序 列 如下 [ 5 1 :
J : ’TI[ TGGGCA CC GAGG1丌 AT 3 B3 5一 1兀 t T T T 一’ J 4 5 5一 AAAGA B .: ’T AAGAACAT T AA GAAAAT
G一3 ’
2 2 第5 ( 第1 期) 0 年 期总 7 1 1
说 明兔 也可 作为 巨颈 绦虫 的 中间宿 主。研究 表 明 , 从 不 同 中 间宿 主获 得 的链尾 蚴 感染 终 末 宿 主猫 后 所 获得 的虫 卵 , 中间宿主 的感染 性存 在差 异 。巨 对 颈 绦虫 的节 片具 有很 强 的运 动 性 ,可 自行 蠕 动很 长 的距 离 。因此 , 果受 感染 的猫 与作 为宠物 的啮 如 齿类 动 物 同居 一 室 ,即可对 这 些 动物具 有 感染 的 危 险 。巨颈绦 虫也偶 然感 染人 类 , 并作 为 中间宿主 和终 末 宿主 。阿根廷 、 日本 等 国 已报 道从 人体 内取 出巨颈 绦 虫 的成虫 ;我 国 台湾也 曾报道 一 例 巨颈 绦虫 感染 4 ] 。 本 文应 用 D A序 列 分 析 技 术 对 巨颈 囊 尾 蚴 N 的线粒 体 po 1 因进 行分 析与研 究 。o l 因属 cx 基 cx 基 于 线粒 体 基 因组 ,从 而 它 的每 一个 序 列都 是个 单 模标 本 【 o l 因具 有母 系遗传 为主 、 化速 度 5 x基 ] 。e 进 快和不存在双亲遗传等遗传差异特点 ,可以避免 基因重组 , 易位 、 倒位等畸变 的影响。与核基因相 比来 说 ,线粒 体 基 因进化 如 同一 条 简捷 的途径 发 展 ,研究者认为线粒体基 因更有利于研究分子进 化 的基 因 更适 用 于研 究 种 内遗 传差 异 。 , 本 研 究 选用 了一对 保 守 引物 ,对 来 自湖 南农 业 大 学 自繁 S D系大 鼠巨颈 囊尾 蚴 样 品用 P R方 C 法 扩增 pol 分序 列 ,并 与 G n ak M 收录 的 cx 部 eB nT 绦 虫 pol cx 序列进 行 了 比较 。结 果发 现 , 与收 录 的 巨颈 绦虫 报道 的序 列 同源性 很 高 。与 其 它 的绦 虫 序 列还 有一定 差 异 。本 试验 在 国 内首次 P R扩增 C 了大 鼠肝 脏 内巨颈 囊尾 蚴 的分 子分 类 学 和分 子遗 传 学进 一步研 究 奠定 了基础 。 口
参考文献 :
引 物 由上 海 生 工 生 物 技 术 有 限 公 司进 行 合 成。 扩增 体 系为 5 L 依 次加 入 双蒸水 1.5 I 0 。 52 、 1 XP R uf 25 L Mg 1 ( 5 0 C b f r . e 、 C2 2 mM) . L 1 、 5 d T s (5 NP 2 mM) L Pi e 1O M) . L T q 3 ,r r(O p O5 , a m plm rs / L 02 L模 板 D A2 L 反 应 o eae U ). y 5 5 , N ; 在 Bo e a i t 循环反应仪上进行, mr 扩增条件为:4 9 预 变 性 5m n 然后 9 性 3 、5退火 3 、 i, 4变 0S5 0S72延 伸 3 s共 3 0, 5个 循 环 , 后 7 最 2延 伸 1mi。 取 0 n 6 LP R产物在 1 T E琼 脂糖 凝 胶 电泳 ,用 溴 C % A 化 乙锭 染 色 , 外 投射 仪 下 观察 结 果 , 胶成 像 系 紫 凝 统 摄像 。
本实 验样 品成 功扩 增 出 4 7 p的 p o l 因 片 9b cx 基 段 , 除 引物 后 , 得 到 49 p的序 列 。将样 品序 剔 均 4b 列 与 G n ak M 收 录 的 巨 颈 绦 虫 、 阔节 裂 头 绦 eB nT 虫 、棘 球 绦虫 、多 头绦 虫 及 泡状 带 绦虫 相应 序 列 ( eB nT 登 录 号 分 别 为 :F 9 621 G n ak M E 00 1.、 AF 9 2 0 AB 0 0 41 E 3 3 2 .、 Q 2 S 91 0 6 4 、 2 8 6 .、 F 9 6 01 G 2 S 1. ) 进 行 比较 ,经 Cutl20Bo dt MeAi . ls x .、 i i与 a E g ln41 g 软 件 分 析 显 示 ,大 鼠肝 脏 分 离 的 巨 颈 囊 尾 蚴 与 G n ak M 中 巨颈 绦 虫 相 似 性 最 高 是 9 .%, e Bn T 93 与 泡 状 带绦 虫相 似性 最 低是 8.%。采 用 N 法 建树 25 J 方 法 构 建 的 系统 发 生树 显 示 ,种 系 发育 树 的拓 扑 结 构 基 本 一 致 ( 3, 沙 大 鼠肝 脏 分离 的 巨颈 囊 图 )长 尾 蚴 序 列 与 G n a k M 中报 道 的 巨颈 绦 虫 位 于 eB nT 同一分枝 , 系统发生树 中的 B o t p 均较高 , otr 值 sa 长沙 分 离株所属分枝与其它 的绦虫所属分枝相 隔较远 。
1 . DNA测 序及 序 列分析 5
根 据 P R扩增 的结 果 ,将 阳性产 物 直接 送南 C 京 金思 瑞有 限公 司测 序 。使用 D A t ( .1 的 N Sa V5 ) r 0 E i e MeA i 程序对测序结果进行序列分析 。 dt q和 g Lg S n
2 结 果
el e n ai egi.g , 1 i e ul , ee pa s dD ne H gl A e r e D z a l n