梨火疫菌(Erwinia amylovora)环腺苷酸受体蛋白基因(crp)的功能分析
群体感应淬灭_防治植物细菌病害的新策略

26(3)241-247 中国生物防治 Chinese Journal of Biological C ontrol 2010年8月群体感应淬灭———防治植物细菌病害的新策略张力群13,田 涛2,梅桂英1(11中国农业大学植物病理系,北京100193;21天津市植物保护研究所,天津300112)摘要:群体感应(quorum sensing,QS)是细菌的一种调控机制,指细菌通过感应特定信号分子的浓度来感知周围环境中自身或其它细菌的数量,并调整相关基因的表达以适应环境的变化。
多种植物病原细菌利用QS系统调控致病因子的表达,因此,QS系统可以作为细菌病害防治的新靶点。
对细菌QS调控机制的干扰和破坏称为群体感应淬灭(quorum quenching)。
本文介绍了QS与植物病原细菌致病性的关系,以及近年来群体感应淬灭研究的新进展。
关 键 词:植物病原细菌;生物防治;群体感应;群体感应淬灭中图分类号:S476;Q93 文献标识码:A 文章编号:100529261(2010)0320241207 Q uorum Q uenching,a N e w Strategy for Controlling Plant B acterial DiseasesZH ANGLi2qun13,TI AN T ao2,MEI G ui2ying1(11Department of Plant Pathology,China Agricultural University,Beijing100193;21Institute of Plant Protection,T ianjin Academy of Agricultural Sciences,T ianjin300112,China)Abstract:Quorum sensing(QS)enables bacteria to m onitor their own population density by means of small,diffusible signals and to coordinate the expression of specialized genes with cell density.Many phytopathogenic bacteria em ploy the QS system to regulate the expression of their virulent factors.This makes QS a very attractive target for the development of novel disease2suppressive strategies.The ability to disrupt QS is known as quorum quenching.This review provides an overview on the relationship be2 tween QS and pathogenicity of phytopathogenic bacteria,and on the progress of the development of the quorum2quenching strategy in plant diseases control during the last decade.K ey w ords:bacterial phytopathogens;biocontrol;quorum sensing;quorum quenching1 细菌的群体感应早在20世纪60年代,研究人员就发现作为单细胞生物的细菌有个体间交流的能力,并能表现出一些多细胞生物的性状,这种细菌间的信号交流方式称作群体感应(quorum sensing, QS)。
检疫性有害生物

49.咖啡潜叶蛾
Leucoptera coffeella Cuerin-Menville
50.稻水象甲
Lissorhoptrus oryzophilus Kuschel
51.黑森瘿蚊
Mayetiola destructor (Say)
• 52.木薯单爪螨 Mononychellus tanajoa (Bonder)
58.欧洲大榆小蠹 Scolytus scolytus Fabricius
59.剑麻象甲 Scyphophorus acupunctatus Gyll.
60.芒果甲象 Sternochetus frigidus (Fabricius) Sternochetus mangiferae (Fabricius) Sternochetus olivieri (Faust)
内发育化蛹。
9.草莓花象 学名:Anthonomus signatus Say 分布:北美。 寄主:草莓、悬钩子、黑莓、露莓及洋莓属植物。 为害部位或传播途径:幼虫取食花芽。
10.点刻长蠹 学名:Apate monachus Fabricius 分布:几内亚、多哥、加纳、象牙海岸、喀麦隆、埃塞俄比亚、肯尼亚、
阿根廷、巴西、秘鲁、乌拉圭和委内瑞拉。 寄主:菊科。 为害部位或传播途径:幼虫潜食叶片。
7.按实蝇 学名:Anastrepha spp.(南美按实蝇、墨西哥按实
蝇、西印度按实蝇和加勒比按实蝇除外) 分布: 南部以南美洲地区。 寄主:许多种瓜、果类植物等。 为害部位或传播途径:以幼虫钻蛀果实。
8.苹象 学名:Anthonomus quadrigibbus Say 分布: 、加拿大。 寄主:苹果、梨、酸苹果、山楂、樱桃等。 为害部位或传播途径:成虫在幼果上产卵、幼虫在果
环磷酸腺苷反应元件

环磷酸腺苷反应元件一、什么是环磷酸腺苷反应元件?环磷酸腺苷反应元件(cAMP response element,CRE)是一种位于基因启动子区域的DNA序列,它可以与激活转录因子结合并调节基因的转录。
CRE由两个部分组成:一个核心序列TGACGTCA和相邻的AT富集区域。
CRE主要通过激活腺苷酸环化酶(adenylyl cyclase)产生的cAMP来调节基因表达。
二、CRE的作用机制是什么?1. 激活转录因子结合cAMP通过激活蛋白激酶A(protein kinase A,PKA)使其进入细胞核,然后与CREB(cAMP response element binding protein)结合形成复合物。
该复合物能够识别和结合CRE,并促进基因转录。
2. 促进染色质重塑除了直接影响基因转录外,CRE还可以通过调节染色质状态来影响基因表达。
研究表明,PKA可以磷酸化组蛋白H3,从而改变染色质的结构和稳定性,促进基因表达。
3. 参与神经传递过程CRE在神经系统中起着重要的作用。
研究表明,PKA/CREB信号通路参与了记忆形成和学习等神经传递过程,对神经元的发育、生长和功能维持有着重要的影响。
三、CRE在哪些生物学过程中起作用?1. 免疫调节CRE通过调节T细胞分化、B细胞增殖和抗体产生等过程参与免疫调节。
研究表明,CREB可以促进T细胞向Th2型分化,并抑制Th1型反应。
2. 肿瘤发生CRE在肿瘤发生和转移中也扮演着重要角色。
许多癌细胞中cAMP/PKA/CREB信号通路异常激活,导致基因表达异常、增殖和转移。
3. 神经系统发育CRE对神经系统发育有着重要的影响。
在大脑皮层的形成中,PKA/CREB信号通路能够促进神经元的迁移和分化,并参与突触形成和功能塑性。
四、如何检测CRE活性?1. 滨藻酸酯(Luciferase)报告基因法将CRE序列与滨藻酸酯基因连接,转染至细胞中,通过检测滨藻酸酯酶的荧光信号来评估CRE的活性。
群体感应淬灭_防治植物细菌病害的新策略
26(3)241-247 中国生物防治 Chinese Journal of Biological C ontrol 2010年8月群体感应淬灭———防治植物细菌病害的新策略张力群13,田 涛2,梅桂英1(11中国农业大学植物病理系,北京100193;21天津市植物保护研究所,天津300112)摘要:群体感应(quorum sensing,QS)是细菌的一种调控机制,指细菌通过感应特定信号分子的浓度来感知周围环境中自身或其它细菌的数量,并调整相关基因的表达以适应环境的变化。
多种植物病原细菌利用QS系统调控致病因子的表达,因此,QS系统可以作为细菌病害防治的新靶点。
对细菌QS调控机制的干扰和破坏称为群体感应淬灭(quorum quenching)。
本文介绍了QS与植物病原细菌致病性的关系,以及近年来群体感应淬灭研究的新进展。
关 键 词:植物病原细菌;生物防治;群体感应;群体感应淬灭中图分类号:S476;Q93 文献标识码:A 文章编号:100529261(2010)0320241207 Q uorum Q uenching,a N e w Strategy for Controlling Plant B acterial DiseasesZH ANGLi2qun13,TI AN T ao2,MEI G ui2ying1(11Department of Plant Pathology,China Agricultural University,Beijing100193;21Institute of Plant Protection,T ianjin Academy of Agricultural Sciences,T ianjin300112,China)Abstract:Quorum sensing(QS)enables bacteria to m onitor their own population density by means of small,diffusible signals and to coordinate the expression of specialized genes with cell density.Many phytopathogenic bacteria em ploy the QS system to regulate the expression of their virulent factors.This makes QS a very attractive target for the development of novel disease2suppressive strategies.The ability to disrupt QS is known as quorum quenching.This review provides an overview on the relationship be2 tween QS and pathogenicity of phytopathogenic bacteria,and on the progress of the development of the quorum2quenching strategy in plant diseases control during the last decade.K ey w ords:bacterial phytopathogens;biocontrol;quorum sensing;quorum quenching1 细菌的群体感应早在20世纪60年代,研究人员就发现作为单细胞生物的细菌有个体间交流的能力,并能表现出一些多细胞生物的性状,这种细菌间的信号交流方式称作群体感应(quorum sensing, QS)。
梨火疫病菌的实时荧光PCR检测

中图分类号 :¥3 . 466
文献标识码 :A
文章编号 :0 1 -94 2 0 )202 - 4 20 1 (0 6 0 -1 30 6
梨 火 疫病 菌 ( rii a yooa 是 我 国一 类 Ewna m lvr)
定 ;) 2 血清学检 测技术 , E IA和免疫荧光等。 如 LS
P t o wh c a o p s , CR n r l t n n e u e a s o i v e u ta d c n a n t n rs s CR meh d。 i h h sn o t P ma i u a i sa d r d c s f l p st e r s l n o tmi ai ik , I o e i o
( 南京农业大学植物保护学 院 农 业部病虫检测与治理重点实验 室 , 南京 209 ) 105
摘要: 根据梨火疫病菌 1S~ 3 6 2S间的 IS保守序列, T 设计并合Байду номын сангаас了一对特异性 引物 R A F A, E / E 应用荧光染料 S B YR
GenI l r , O个梨 火疫的菌株和其它相关参试菌株进行 了检测 。结果 表明 , e 对 l O个梨火疫菌株 都产生荧 光信号而其 它参试 菌株都不 产生荧光信号 , 成功建立了梨火疫病 菌的实时荧光 P R检测方法 。整个检测 过程 只需 3 , 闭管 , 了污染 C 完全 h 降低 的机 会 , P R后处理。检测 的灵敏度是 4 无需 C 个菌体细胞 , 比常规 P R电泳检测提高 了 1 倍 。用该特 异性 引物对梨 枝条 C 0 浸泡 液进行实 时荧 光 P R检测 , C 结果可特异性检测 到 目 标菌 的存 在 , 检测 的灵 敏度是 2 个 菌体 细胞 , 并且 4 比常规 P R电 C
环化核苷酸的功能研究进展

综 述生命科学仪器 2023年第21卷/第5期8作者简介:李钊(1998),男,E -m a i l :3120201405@b i t .e d u .c n㊂通讯作者:高昂(1983),女,博士,北京理工大学,教授,E -m a i l :a n g .g a o @b i t .e d u .c n ㊂基金项目:国家自然科学基金项目(32171219)环化核苷酸的功能研究进展李 钊 高 昂*(北京理工大学生命学院,分子医学与生物诊疗工业和信息化部重点实验室,北京100081)摘要 环化核苷酸是由一个或多个核苷单磷酸通过磷酸二酯键形成的环状二酯小分子,其作为信号转导分子在调节细胞代谢㊁转录调控㊁细胞生长等生理功能中发挥着重要作用㊂最新研究发现多种类型的环化核苷酸分子能够激活后生动物的固有免疫反应和细菌的抗噬菌体防御系统㊂本文总结了目前发现的环化核苷酸种类,对其功能机制的研究进展做了简要综述,并提出了目前所存在的问题以及潜在开发方向㊂关键词 环化核苷酸;信号通路调控;固有免疫;适应性进化R e s e a r c h P r o g r e s s o n t h e F u n c t i o n o f C yc l i c N u c l e o t ide s L I Z h a o ,G A O A n g*(S c h o o l o f L i f e S c i e n c e ,B e i j i n g I n s t i t u t e o f T e c h n o l o g y ,K e y L a b o r a t o r y o f Mo l e c u l a r M e d i c i n e a n d B i o d i a g n o s i s ,M i n i s t r y o f I n d u s t r y a n d I n f o r m a t i o n T e c h n o l o g y ,B e i j i n g ,100081)ʌA b s t r a c t ɔC y c l i c n u c l e o t i d e s a r e c y c l i c d i e s t e r s m a l l m o l e c u l e s f o r m e d b y o n e o r m o r e n u c l e o t i d e m o n o p h o s ph a t e s t h r o u g h p h o s p h o d i e s t e r b o n d s .A s s i g n a l t r a n s d u c t i o n m o l e c u l e ,t h e y p l a y i m p o r t a n t r o l e s i n m a n y p h y s i o l o gi c a l f u n c t i o n s i n c l u d i n g c e l l m e t a b o l i s m ,t r a n s c r i p t i o n r e g u l a t i o n a n d c e l l gr o w t h .T h e l a t e s t r e s e a r c h h a s f o u n d t h a t v a -r i o u s t y p e s o f c y c l i c n u c l e o t i d e m o l e c u l e s c a n a c t i v a t e t h e i n n a t e i mm u n e r e s p o n s e o f m e t a z o a n s a n d t h e a n t i -p h a ge d ef e n s e s y s t e m o f b a c t e r i a .T h i s a r t i c l e s u mm a r i z e s t h e t y p e s o f c y c l i c n u c l e o t i d e s d i s c o v e r e d s o f a r ,b r i e f l yr e v i e w s t h e r e s e a r c h p r o g r e s s o f t h e i r f u n c t i o n a l m e c h a n i s m s ,a n d p u t s f o r w a r d t h e c u r r e n t p r o b l e m s a n d p o t e n t i a l d e v e l o p-m e n t d i r e c t i o n s .ʌK e y wo r d s ɔc y c l i c n u c l e o t i d e s ;r e g u l a t i o n o f s i g n a l p a t h w a y ;i n n a t e i mm u n i t y ;a d a p t i v e e v o l u t i o n 中图分类号:Q 93 文献标识码:A D O I :10.11967/2023211002引言近些年发现,环化核苷酸作为第二信使分子参与调控多项生理活动㊂环化核苷酸主要以环状单磷酸核苷酸和环状寡核苷酸两种形式存在于生物体内㊂目前已知的环状单磷酸核苷酸c AM P ㊁c GM P 在生物体内的代谢㊁转录调控㊁视觉信号等生理过程中起着重要的信号转导作用;3'3'-c -d i -AM P ㊁3'3'-c -d i -GM P ㊁2'3'-c G AM P 等环状寡核苷酸参与调控包括核糖开关㊁基因转录㊁细菌运动性㊁生物膜的形成和固有免疫等生理活动㊂环化核苷酸及其下游信号通路的研究,极大地推动了疫苗等相关临床药物的研发,拓展了免疫疗法的应用㊂例如固有免疫反应中2'3'-c G AM P 的发现,引发了对固有免疫反应激动剂的研究,一系列环化核苷酸类激动剂和纳米药物被研发用于治疗病原体感染和肿瘤等疾病㊂因此,对于环化核苷酸的研究具有重要的理论价值和应用前景㊂本文基于目前对于环化核苷酸的研究,系统总结了其多样性以及在生物体内的调控机制㊂1 环状单磷酸核苷酸环状单磷酸核苷酸通常由核苷酸分子的磷酸部分与核糖部分第三位碳原子的羟基脱水缩合形成一个环状的核苷酸(图1)㊂目前在生物体内发现了四种环状单磷酸核苷酸:c AM P ㊁c GM P ㊁c C -M P ㊁c UM P ㊂其中,c AM P 与c GM P 参与调控体内代谢㊁基因转录等多项生理活动;c C M P 与c UM P 能够激活细菌抗噬菌体系统㊂图1 环状单磷酸核苷酸合成图生命科学仪器 2023年第21卷/第5期综 述9F i g .1S y n t h e s i s o f c y c l i c n u c l e o t i d e m o n o p h o s ph a t e s 1.1 环状单磷酸腺苷 环状单磷酸腺苷(C yc l i c ade n o s i n e m o n o p h o s ph a t e ,c AM P )作为第一个被发现的第二信使,E a r l W.S u t h e r l a n d 在它的基础上提出第二信使学说,更新了对细胞内信号传导系统的理解[12]㊂c AM P 在动植物和微生物中含量很少,主要参与c AM P -P K A 信号转导通路[1]:当细胞中的激素或神经递质作为配体与细胞膜上的G 蛋白偶联受体(G p r o t e i n-c o u p l e d r e c e pt o r s ,G P C R s )结合后,G P C R s 会释放G 蛋白的亚基与腺苷酸环化酶(A d e n y l y l c yc l a s e ,A C )结合㊂A C 被激活后会催化A T P 脱去一个焦磷酸生成c AM P ,c AM P 进一步激活c AM P 依赖蛋白激酶A (P K A )㊂P K A 在未被激活的状态下是由两个催化亚基和两个调节亚基组成的四聚体,c AM P 与调节亚基结合后会改变P K A 的构象,使调节亚基和催化亚基发生解离,催化亚基进而磷酸化不同功能的靶蛋白,进而调控生长发育㊁调节转录调控㊁细胞血糖水平等生理活动(图2),最终c AM P 被磷酸二酯酶(P D E )水解为AM P 而终止c AM P 所传导的生理信号㊂除了c AM P-P K A 途径外,c AM P 直接调控E p a c (E x c h a n g e p r o t e i n d i r e c t l y a c t i v a t e d b y c AM P ),参与调控细胞增殖㊁分化与凋亡[17];c AM P 还可以激活环核苷酸门控离子通道(C yc l i c n u c l e o t ide -g a t e d c h a n n e l ,C N G ),在激活状态时可以允许阳离子非选择性通过,进而调节神经元活动[18]㊂1.2 环状单磷酸鸟苷 环状单磷酸鸟苷(C yc l i c g u a n o s i n e m o n o p h o s ph a t e ,c GM P ),在c AM P 被鉴定的几年后在哺乳动物组织和体液中被证实存在㊂鸟苷酸环化酶分为可溶性鸟苷酸环化酶(S o l -u b l e g u a n y l y l c yc l a s e s ,s G C )和膜结合鸟苷酸环化酶(P a r t i c u l a t e g u a n y l y l c y c l a s e s ,pG C )两种,二者被激活后催化G T P 脱去一个焦磷酸生成c GM P ,不同类型的鸟苷酸环化酶产生的c GM P 决定其调节不同信号通路,c GM P 分子最终被P D E 水解而终止信号㊂细胞内的P D E 有多种亚型,c GM P 可通过激活或抑制P D E 2/3以调节细胞内c AM P 的浓度及c AM P /P K A 信号通路[2],因此c GM P 对c AM P 具有拮抗作用㊂细胞内N O 和利钠肽(N P s )分别激活s G C 和p G C 生成c GM P ,c GM P进一步激活c GM P 依赖蛋白激酶G (P K G ),P K G通过磷酸化下游靶蛋白进而调节血管舒张㊁神经传导等生理功能[20](图2);c GM P 在视觉信号转导中有着重要作用,直接调控C N G ㊂在黑暗中,视网膜的视杆细胞内c GM P 浓度升高,激活C N G ,从而允许大量的胞外N a +与C a 2+通过;当光信号传入时,会激活视杆细胞的视紫红质G P C R ,并通过一系列级联反应激活P D E ㊁水解c GM P ,进而关闭C N G ,导致N a +和C a 2+浓度降低使膜超极化,最终引起大脑对光的视觉反应㊂最新研究发现c GM P 在骨生长㊁心衰竭等方面也发挥着重要作用㊂图2 c AM P 与c GM P 的信号通路F i g .2S i g n a l i n g p a t h w a ys o f c AM P a n d c GM P 注:G P C R s :G 蛋白偶联受体;A C :腺苷酸环化酶;pG C :膜结合型鸟苷酸环化酶;s G C :可溶性鸟苷酸环化酶;P K A :c AM P 依赖蛋白激酶(R :调节亚基;C :催化亚基);P K G :c GM P 依赖蛋白激酶;C N G :环核苷酸门控离子通道;A N P :心房利钠肽;B N P :脑钠肽;U R O :尿钠素㊂1.3 环状单磷酸嘧啶(c C M P ㊁c UM P ) 当c AM P 与c GM P 被鉴定后,环状单磷酸嘧啶的存在及其功能引起了大量关注,但由于技术原因环状单磷酸嘧啶的研究一直停滞不前㊂直至2014年,研究人员在人源H E K 293细胞和鼠源B 103细胞中发现了高浓度的c C M P 和c UM P ,同时发现s A C 与s G C 均可产生c C M P 与c UM P ,且二者的下游靶标为P K A ㊁P K G 等㊂2021年,研究人员在细菌中发现了合成c C M P 和c UM P 的嘧啶环化酶P yc C (图1),并发现在细菌中c C M P 和c UM P 可以作为第二信使激活细菌抗噬菌体系统 P yc s c a r 系统㊂该研究的发现再次引起了人们对这两种环状单磷酸嘧啶的关注[25]㊂2 环状寡核苷酸目前在生物体内发现了多种不同长度㊁不同碱基组成的环状寡核苷酸,并发现其在激活生物体固有免疫方面有着重要作用㊂同时,关于环状寡核苷酸研究还提示细菌和后生动物之间可能存在免疫系统进化关系[26]㊂2.1 环二鸟苷酸 1987年,研究人员在A c e t o -b a c t e r x yl i n u m 中发现了纤维素合成酶的激活剂综 述生命科学仪器 2023年第21卷/第5期10环二鸟苷酸(c -d i -GM P ),并确定了两个GM P 分子之间是以3'-5'磷酯键连接[27]㊂c-d i -GM P 是在所有生物体内发现的第一个环二核苷酸㊂此后在多种细菌中均发现了c -d i -GM P 的存在,证明c -d i -GM P 是一种在细菌中通用的第二信使分子[28]㊂c -d i -GM P 是由二鸟苷酸环化酶(d i g u a n y l a t e c yc l a s e ,D G C )催化两分子G T P 脱去两分子焦磷酸环化生成㊂c -d i -GM P 主要作用于两种效应因子,一种是效应蛋白,例如,含P i l Z 结构域的蛋白:大肠杆菌中的Y c g R 可与c -d i -GM P 结合调控细菌的运动能力;依赖c -d i -GM P 调控的D N A 结合蛋白或转录因子:霍乱弧菌中的c -d i -GM P 可抑制应激反应调节因子R o pS 的表达等[30]㊂另一种是核糖开关,c -d i -GM P 能专一性结合核糖开关G E MM (G e n e s r e -l a t e d t o t h e E n v i r o n m e n t ,M e m b r a n e s a n d M o t i l i -t y),调控蛋白质的翻译㊂目前,c -d i -GM P 被鉴定参与调控细菌运动性㊁生物膜形成㊁多糖形成等多个生理生化过程,同时有研究表明其能够激活后生动物的固有免疫反应[7]㊂c -d i -GM P 的出现,为其他环状寡核苷酸的发现奠定了基础㊂2.2 环二腺苷酸 环二腺苷酸(c -d i -AM P )是生物体内发现的第二个环二核苷酸,它是在解析一种D N A 完整性扫描蛋白(D N A i n t e g r i t y sc a n -n i n g pr o t e i n A ,D i s A )结构时发现存在于D i s A 的晶体结构中,其功能为在细菌D N A 损伤时延迟产孢㊂D i s A 蛋白的N 端结构域能够催化两分子A T P 合成c -d i -AM P ,因此该结构域被命名为D A C 结构域(D i -A d e n y l y l C yc l a s e )㊂c-d i -AM P 在细菌中的受体有两类,一类是蛋白质类受体,例如T e t R 家族的转录因子D a r R 和钾离子转运蛋白家族K t r A 蛋白㊂D a r R 蛋白与c-d i -AM P 结合后会增强自身D N A 结合能力,导致多种基因的表达被抑制,进而影响细菌内脂肪酸的合成等生理活动;K t r A 蛋白由R C K _N 和R C K _C两个结构域组成,其R C K _C 结构域与c -d i -AM P 特异性结合后会引起膜转运蛋白K t r B 的构象或聚集状态改变,进而影响细菌的钾离子转运;另一类是核糖开关类受体,核糖开关y d a O 形成一个 方形 赝对称结构并结合两个c -d i -AM P 分子,yd a O 核糖开关参与调控细菌细胞壁代谢㊁芽孢形成等生理活动[5]㊂目前,c -d i -AM P 已经被鉴定参与细菌多种生理功能,包括监测D N A 损伤㊁控制细胞脂肪酸合成㊁芽孢形成等,同时其在后生动物体内还可诱导宿主细胞产生免疫反应[39]㊂2.3 环状鸟苷酸-腺苷酸2.3.1 3'3'-环状鸟苷酸-腺苷酸 2012年,研究人员在霍乱弧菌V.c h o l e r a e 中发现了一种新型环二核苷酸3'3'-环状鸟苷酸-腺苷酸(3'3'-c G AM P ),它由寡核苷酸环化酶D n c V 催化一分子A T P 和一分子G T P ,通过两个3'-5'磷酯键环化生成(图3)㊂3'3'-c G AM P 起初被发现参与调节霍乱弧菌的毒力㊁趋化性等生理功能,但其结合的效应因子一直未被发现㊂2018年,在霍乱弧菌中发现一种马铃薯糖蛋白样磷脂酶C a p V 可以被3'3'-c G AM P 激活,从而降解细胞膜中的磷脂,并释放游离的脂肪酸㊂后续研究发现D n c V-c G AM P -C a pV 的通路具有抗噬菌体防御功能,并由D n c V ㊁C a pV ㊁泛素化相关结构域E 1㊁E 2以及J A B /J AMM 家族的异肽酶四种基因构成操纵子[43]㊂噬菌体侵染细菌会激活D n c V 生成3'3'-c G AM P 然后激活C a p V 降解细胞膜,导致膜完整性的丧失,因此细菌会在噬菌体完成复制前死亡㊂该通路通过介导顿挫感染从而起到抗噬菌体防御的作用㊂图33'3'-c G AM P 合成图F i g .3S yn t h e s i s o f 3'3'-c G AM P 2.3.2 2'3'-环状鸟苷酸-腺苷酸 2'3'-环状鸟苷酸-腺苷酸(2'3'-c G AM P )是第一个在后生动物中发现的环状寡核苷酸分子,在后生动物中参与了重要的固有免疫反应㊂2013年研究发现后生动物细胞中存在一种寡核苷酸环化酶c G A S (C y c l i c GM P-AM P S yn t h a s e ),当宿主细胞被病原体感染后,c G A S 可以直接识别病原体D N A ,该过程中c G A S 会与d s D N A 结合形成一个2:2的复合物,催化一分子G T P 和一分子A T P 脱去两分子焦磷酸并环化生成第二信使c G AM P ㊂c G AM P 可直接结合并激活接头蛋白S T I N G ,进而招募激酶I K K (I B k i n a s e )和T B K 1(T A N K-b i n d i n g ki n a s e 1),分别激活转录因子N F-κB (N u c l e a r f a c t o r )和I R F 3(I n t e r f e r o n r e g u l a t o r y f a c t o r),最终诱导I 型干扰素和其他促炎细胞因子的表达㊂同年6月,研究人员通过结构和功能实验发现c G A S 产生的c G AM P 与细菌的3'3'-生命科学仪器 2023年第21卷/第5期综 述11c G AM P 并不相同,而是一种包含2'-5'和3'-5'混合磷酯键的新型环二核苷酸:2'3'-c G AM P (图4)㊂c G A S -c G AM P-S T I N G 信号通路的发现加深了人们对后生动物天然免疫信号通路的认识,同时也引发了人们对后生动物中是否存在其他环状寡核苷酸的关注㊂图4 2'3'-c G AM P 合成图F i g .4S yn t h e s i s o f 2'3'-c G AM P 2.3.3 3'2'-环状鸟苷酸-腺苷酸 c G A S在被发现后,研究人员在动物基因组中检测到了多个与c G A S 具有同源性的蛋白质,但这些蛋白质的功能尚不清楚㊂2019年,研究鉴定了一类先天免疫传感器的c G L R s 蛋白(c G A S -l i k e r e c e p-t o r s),能够识别不同的分子模式并催化合成不同的环状寡核苷酸信号分子㊂其中果蝇的c G L R 1能够识别双链R N A 并被激活合成新的环二核苷酸3'2'-c G AM P ,进而与d S T I N G (D r o s o ph i l a S T I N G )结合,发挥其抗病毒免疫作用㊂同年,在不黏柄菌属(A s t i c c a c a u l i s s p.)中发现C d n G 能够在体外合成3'2'-c G AM P ,后者能够结合A s -C a p5使其二聚化并激活其HN H 核酸酶结构域降解D N A ,进而达到抵抗噬菌体的功能[51]㊂这些研究证实了细菌和后生动物在环化核苷酸信号上存在一定的进化关系㊂2.4 其他环状寡核苷酸的发现 继2008年发现第一个环二核苷酸c -d i -GM P ,10年间陆续发现的环状寡核苷酸均由嘌呤核苷酸环化而成㊂2019年,P h i l i p J .K r a n z u s c h 课题组在大肠杆菌(E s c h e r i c h i a c o l i )中发现了一个D n c V 的同源蛋白C d n E ,C d n E 能够催化一分子U T P 和一分子A T P 环化生成嘌呤-嘧啶环二核苷酸3'3'-c UM P -AM P ,其下游效应蛋白为马铃薯糖蛋白样磷脂酶C a p E ,被c -UM P-AM P 激活后可通过顿挫感染的方式抵抗噬菌体的侵染㊂同时,研究人员将c G A S ㊁D n c V ㊁C d n E ㊁O A S 1等能够生成环状寡核苷酸的蛋白质称为c G A S /D n c V 样核苷酸转移酶(c G A S /D n c V -l i k e n u c l e o t i d yl t r a n s -f e r a s e s ,C D-N T a s e s ),并通过对所有C D-N T a -s e s 进行生信分析和大规模的正向生化筛选,鉴定出了七种嘌呤㊁嘧啶㊁嘌呤-嘧啶环二核苷酸:c -d i -GM P ㊁c -d i -AM P ㊁c -d i -UM P ㊁c G AM P ㊁c UM P -AM P ㊁c UM P-GM P ㊁c C M P-UM P ,揭示了C D-N T a s e s 选择核苷酸的分子机制㊂此外,阴沟肠杆菌E n t e r o b a c t e r c l o a c a e 的寡核苷酸环化酶C d n D 能够生成环三核苷酸3'3'3'-c AM P-AM P-GM P (c A A G ),c A A G 激活下游效应蛋白C a p4核酸内切酶活性进而切割D N A ㊂大肠杆菌中的核酸内切酶N u c C 可以被3'3'3'-c AM P -AM P -AM P (c A A A )激活组装为六聚体切割细菌染色质导致细菌死亡,进而抵抗噬菌体的侵染㊂在Ⅲ型C R I S P R-C a s 系统中发现了一种环状寡聚腺苷酸(c O A ),A T P 能够被C a s 10亚基的P a l m结构域催化生成c O A ,c O A 通过二聚化下游C s m 6/C s x 1核糖核酸酶的C A R F 结构域,从而激活其切割R N A 活性,抵抗噬菌体的感染[57]㊂多种不同碱基㊁长度以及磷酯键成键方式的环状寡核苷酸的发现,体现了其在生物体内的多样性,同时环状寡核苷酸参与调节后生动物的固有免疫反应以及细菌中的抗噬菌体感染等重要生理过程,体现了其在生物体免疫系统中的重要性㊂3 环状寡核苷酸在细菌抗噬菌体信号系统中的作用2019年,研究发现3'3'-c G AM P 在细菌体内具有抗噬菌体功能,并发现了一种基于环状寡核苷酸的抗噬菌体信号系统 C B A S S 系统(C y -c l i c o l i g o n u c l e o t i d e -b a s e d a n t i -p h a g e s i g n a l i n gs ys t e m )[43],该系统通过产生环状寡核苷酸信号激活下游效应蛋白以顿挫感染的方式抵抗噬菌体感染㊂C B A S S 系统根据其操纵子的组成分为四类[60],效应蛋白通过多种方式造成细胞死亡来抵御噬菌体的侵染㊂Ⅰ型C B A S S 系统,是最基础的C B A S S 系统,仅由C D-N T a s e 和效应基因两个核心部分组成,例如耶尔森氏鼠疫杆菌Y e r s i n i a a l e k s i c i a e 的2TM (T r a n s m e m b r a n e )蛋白C a p 15可被上游C D-N T a s e 产生的3'3'-c G AM P 激活并在细胞膜上打孔进而杀死细胞㊂Ⅱ型C B A S S系统除核心组分外,还包括编码泛素化相关结构域的辅助基因C a p 2和C a p 3㊂D n c V-C a p V 通路则属于该类系统,3'3'-c G AM P 激活C a pV 降解细胞膜中的磷脂进而造成细胞死亡㊂Ⅲ型C B A S S 系统除核心组分外,包含编码T R I P 13和HO R MA 蛋白的辅助基因C a p 6和C a p 7㊁C a p8㊂大肠杆菌的C d n C 必须结合HO R MA 蛋白才能催化生成环三腺苷酸(c A A A ),N u c C 结合c A A A 后综 述生命科学仪器 2023年第21卷/第5期12发生构象变化由三聚体组装形成六聚体,进而切割细菌染色质导致细菌死亡㊂Ⅳ型C B A S S 系统是一种比较稀有的操纵子组合,主要存在于古细菌中,目前该类型中环化核苷酸的调控通路尚不清楚㊂C B A S S 系统是一个复杂多样的抗噬菌体信号系统,多种环状寡核苷酸在其中起着承上启下的信号传导作用㊂4 以环状寡核苷酸为信号的免疫反应通路之间的进化关系细菌的C B A S S 系统与后生动物的c G A S-S T I N G 通路均通过环状寡核苷酸作为信号分子来激活免疫防御反应,提示了环状寡核苷酸引发的免疫反应可能起源于细菌,并在后生动物中适应性进化㊂最新研究解析了黄杆菌科F l a v o b a c t e -r i a c e a e s p.的S T I N G (F s S T I N G )结合3'3'-c G AM P 的复合物结构,其与人源S I T N G (h S T -I N G )结合2'3'-c G AM P 的复合物结构具有相似的 V 型同型二聚体 和 疏水α螺旋 构型㊂同时,该研究还根据多个种属的S T I N G 蛋白结构分析构建了细菌与后生动物S T I N G 蛋白的系统发育树,确认了固有免疫中环状寡核苷酸分子的进化模型,并表明S T I N G 蛋白在进化过程中存在适应性㊂自此,c G A S-S T I N G 通路中的每个核心部分都在细菌中找到了对应的存在,也基本证实了后生动物的固有免疫反应(c G A S-S T I N G 通路)与细菌的抗噬菌体防御机制之间存在进化关系㊂目前后生动物仅能够自身合成2'3'-c G AM P 和3'2'-c G AM P ,而细菌虽然能够自身合成多种环状寡核苷酸,但并未在细菌中发现2'3'-c G AM P 的存在(表1)㊂同时,在c G A S-S T I N G 通路中2'3'-c G AM P 激活S T I N G 蛋白的能力比细菌的环状寡核苷酸更强,且2'3'-c G AM P 仅能被E N P P 1特异性水解,而细菌中的3'3'-c G AM P 等环化核苷酸却不能被E N P P 1水解,这也体现了2'3'-c G AM P 在免疫反应进化过程中的适应性和独特性㊂目前所有环状寡核苷酸中只有3'2'-c G AM P 同时存在于后生动物和细菌中且均参与自身固有免疫反应,后生动物能否自身合成其他类型环状寡核苷酸成为了目前的一个研究热点㊂表1 目前发现的环状寡核苷酸分子及其种属T a b .1T h e d i s c o v e r e d c y c l i c o l i g o n u c l e o t i d e s a n d t h e i r s pe c i e s 环状寡核苷酸细菌后生动物3'3'-c -d i -GM P A c e t o b a c t e r x yl i n u m -3'3'-c -d i -AM P B a c i l l u s s u b t i l i s-2'3'-c G AM P -H u m a n M o u s e3'3'-c G AM PV.c h o l e r a e-3'2'-c G AM PA s t i c c a c a u l i s s p.D r o s o ph i l a 3'3'3'-c AM P -AM P -AM P E s c h e r i c h i a c o l i -3'3'3'-c AM P -AM P -GM PE n t e r o b a c t e r c l o a c a e-3'3'-c UM P -AM P E s c h e r i c h i a c o l i-3'3'-c -d i -UM P L e g i o n e l l a p n e u m o ph i l a -3'3'-c UM P -GM PB r a d yr h i z o b i u m d i a z o e f f i c i e n s -5 展望自1958年发现了第一个环单磷酸核苷酸c AM P 至今[12],多种环化核苷酸被发现并验证了其在生物体内的功能:从调节生物体内的生理活动到激活后生动物的固有免疫反应以及细菌抗噬菌体防御系统,在生物体内扮演了重要角色㊂针对环化核苷酸的研究,目前仍有一些问题亟待解决㊂例如,后生动物中是否存在其他类型的环化核苷酸及其参与调控的信号通路细菌的寡核苷酸环化酶是否与c G A S 具有完全相同的激活方式,即需要d s D N A 激活才能生成环化核苷酸?环化核苷酸作为S T I N G 蛋白的激活剂,目前在免疫以及炎症领域备受关注,其相关药物的开发具有重要的临床意义㊂此外,细菌中环化核苷酸相关的抗噬菌体免疫的信号通路,参与调控细菌多种重要生命活动,具有被开发为分子生物学工具的生命科学仪器 2023年第21卷/第5期综 述13巨大潜力㊂参考文献[1]S a s s o n e-C o r s i ,P .,T h e c y c l i c AM P p a t h w a y [J ].C o l d S p r i n g H a r b P e r s p e c t B i o l ,2012,4(12).[2]H o f m a n n ,F .,T h e c GM P s y s t e m :c o m po n e n t s a n d f u n c t i o n [J ].B i o l C h e m ,2020,401(4):447-469.[3]O p o k u-T e m e n g ,C .,e t a l .,C yc l i cd i n u c le o t i d e (c -d i -GM P ,c -d i -AM P ,a n d c G AM P )s i g n a l i n gs h a v e c o m e o f a g e t o b e i n h i b i t e d b y sm a l l m o l e c u l e s [J ].C h e m C o mm u n (C a m b ),2016,52(60):9327-42.[4]R y j e n k o v ,D.A.,e t a l .,T h e P i l Z d o m a i n i s a r e c e pt o r f o r t h e s e c o n d m e s s e n ge r c -d i -GM P :t h e P i l Z d o m a i n p r o t e i n Y c g R c o n t r o l s m o t i l i t y in e n t e r o b a c t e r i a [J ].J B i o l C h e m ,2006,281(41):30310-4.[5]G a o ,A.a n d A.S e r g a n o v ,S t r u c t u r a l i n s i g h t s i n t o r e c o gn i -t i o n o f c -d i -AM P b y th e y d a O r i b o s w i t c h [J ].N a t C h e m B i o l ,2014,10(9):787-92.[6]P e n g ,X.,e t a l .,C yc l i cd i -AM P me d i a t e s b i of i l m f o r m a t i o n [J ].M o l M i c r o b i o l ,2016,99(5):945-59.[7]B u r d e t t e ,D.L .,e t a l .,S T I N G i s a d i r e c t i n n a t e i mm u n es e n s o r o f c yc l i cd i -GM P [J ].N a t u re ,2011,478(7370):515-8.[8]W a n g ,J .,e t a l .,P u l m o n a r y s u r f a c t a n t -b i o m i m e t i c n a n o p-a r t i c l e s p o t e n t i a t e h e t e r o s u b t y p i c i n f l u e n z a i mm u n i t y [J ].S c i e n c e ,2020,367(6480).[9]L u o ,Z .,e t a l .,L a n t h a n i d e -N u c l e o t i d e C o o r d i n a t i o n N a n o -p a r t i c l e s f o r S T I N G A c t i v a t i o n [J ].J A m C h e m S o c ,2022,144(36):16366-16377.[10]Z h e n g ,J .,e t a l .,C o m pr e h e n s i v e e l a b o r a t i o n o f t h e c G A S -S T I N G s i g n a l i n g a x i s i n c a n c e r d e v e l o pm e n t a n d i mm u n o -t h e r a p y [J ].M o l C a n c e r ,2020,19(1):133.[11]T a l ,N.,e t a l .,C y c l i c C M P a n d c yc l i c UM P m ed i a te b a c t e -r i a l i mm u n i t y a g a i n s t p h a ge s [J ].C e l l ,2021,184(23):5728-5739e 16.[12]S u t h e r l a n d ,E .W.a n d T.W.R a l l ,F r a c t i o n a t i o n a n d C h a r -a c t e r i z a t i o n o f a C y c l i c A d e n i n e R i b o n u c l e o t i d e F o r m e d b yT i s s u e P a r t i c l e s [J ].J o u r n a l o f B i o l o g i c a l C h e m i s t r y,1958,232(2):1077-1091.[13]T a s k e n ,K.,e t a l .,S t r u c t u r e ,f u n c t i o n ,a n d r e gu l a t i o n o f h u m a n c AM P -d e pe n d e n t p r o t e i n k i n a s e s [J ].A d v S e c o n d M e s s e n g e r P h o s p h o pr o t e i n R e s ,1997,31:191-204.[14]T a s k e n ,K.a n d E .M.A a n d a h l ,L o c a l i z e d e f f e c t s o f c AM Pm e d i a t e d b y d i s t i n c t r o u t e s o f p r o t e i n k i n a s e A [J ].P h ys i o l R e v ,2004,84(1):137-67.[15]S u n ,Z .B .,e t a l .,c AM P S i g n a l l i n g P a t h w a y in B i o c o n t r o l F u n gi [J ].C u r r I s s u e s M o l B i o l ,2022,44(6):2622-2634.[16]T e n g h o l m ,A.a n d E .G y l f e ,c AM P s i g n a l l i n gi n i n s u l i n a n d g l u c a go n s e c r e t i o n [J ].D i a b e t e s O b e s M e t a b ,2017,19S u p pl 1:42-53.[17]L e e ,K.,E p a c :n e w e m e r g i n g c AM P -b i n d i n g pr o t e i n [J ].B M B R e p,2021,54(3):149-156.[18]K a u p p ,U.B .a n d R.S e i f e r t ,C yc l i c n u c l e o t ide -g a t e d i o n c h a n n e l s [J ].P h ys i o l R e v ,2002,82(3):769-824.[19]F r i e b e ,A.,P .S a n d n e r ,a n d A.S c h m i d t k o ,c GM P :a u -n i q u e 2n d m e s s e n g e r m o l e c u l e -r e c e n t d e v e l o pm e n t s i n c GM P r e s e a r c h a n d d e v e l o p m e n t [J ].N a u n yn S c h m i e d e -b e r gs A r c h P h a r m a c o l ,2020,393(2):287-302.[20]K o e s l i n g ,D.,E .M e r g i a ,a n d M.R u s s w u r m ,P h y s i o l o gi c a l F u n c t i o n s o f N O-S e n s i t i v e G u a n y l y l C y c l a s e I s o f o r m s [J ].C u r r M e d C h e m ,2016,23(24):2653-2665.[21]M i c h a l a k i s ,S .,E .B e c i r o v i c ,a n d M.B i e l ,R e t i n a l C yc l i c N u c l e o t ide -G a t e d C h a n n e l s :F r o m P a t h o p h y s i o l o g y to T h e r a p y [J ].I n t J M o l S c i ,2018,19(3).[22]B l a n t o n ,R.M.,c GM P S i g n a l i n g an d M o d u l a t i o n i n H e a r t F a i l u r e [J ].J C a r d i o v a s c P h a r m a c o l ,2020,75(5):385-398.[23]K a l ya n a r a m a n ,H.,N.S c h a l l ,a n d R.B .P i l z ,N i t r i c o x i d e a n d c yc l i c GM P f u n c t i o n s i n b o n e [J ].N i t r i c O x ide ,2018,76:62-70.[24]H a r t w i g,C .,e t a l .,c AM P ,c GM P ,c C M P a n d c UM P c o n -c e n t r a t i o n s a c r o s s t h e t r e e o f l i f e :H i gh c C M P a n d c UM P l e v e l s i n a s t r o c yt e s [J ].N e u r o s c i L e t t ,2014,579:183-7.[25]S e i f e r t ,R.a n d B .S c h i r m e r ,c C M P a n d c UM P c o m e i n t ot h e s p o t l i g h t ,f i n a l l y [J ].T r e n d s B i o c h e m S c i ,2022,47(6):461-463.[26]J e n s o n ,J .a n d Z .J .C h e n ,B a c t e r i a s t i n g v i r a l i n v a d e r s [J ].N a t u r e ,2020,586(7829):363-364.[27]R o s s ,P .,e t a l .,R e g u l a t i o n o f c e l l u l o s e s yn t h e s i s i n A c e t o -b a c t e r x y l i n u m b y c y c l i c d i g u a n yl i c a c i d [J ].N a t u r e ,1987,325(6101):279-81.[28]R o m l i n g ,U.,M.Y.G a l p e r i n ,a n d M.G o m e l s k y ,C yc l i cd i -GM P :t hef i r s t 25y e a r s o f a u n i v e r s a l b a c t e r i a l s e c o n dm e s s e n ge r [J ].M i c r o b i o l M o l B i o l R e v ,2013,77(1):1-52.[29]C h o u ,S .H.a n d M.Y.G a l p e r i n ,D i v e r s i t y o f C yc l i c D i -GM P -B i nd i n g Pr o t e i n s a n d M e c h a n i s m s [J ].J B a c t e r i o l ,2016,198(1):32-46.[30]W a n g ,H.,e t a l .,I n t e r p l a y a m o n g c y c l i c d i g u a n yl a t e ,H a -p R ,a n d t h e g e n e r a l s t r e s s r e s p o n s e r e g u l a t o r (R po S )i n t h e r e g u l a t i o n o f V i b r i o c h o l e r a e h e m a g g l u t i n i n /p r o t e a s e [J ].J B a c t e r i o l ,2011,193(23):6529-38.[31]S u d a r s a n ,N.,e t a l .,R i b o s w i t c h e s i n e u b a c t e r i a s e n s e t h es e c o n d m e s s e n g e r c yc l i cd i -GM P [J ].S c ie n c e ,2008,321(5887):411-3.[32]Z i mm e r ,J .,A M o l e c u l a r D e s c r i p t i o n o f C e l l u l o s e B i o s yn -t h e s i s [J ].B i o p h ys i c a l J o u r n a l ,2015,108(2).[33]L e e ,V.T.,e t a l .,A c y c l i c -d i -GM P r e c e p t o r r e qu i r e d f o r b a c t e r i a l e x o p o l y s a c c h a r i d e p r o d u c t i o n [J ].M o l M i c r o b i -o l ,2007,65(6):1474-84.[34]Y i n ,W.,e t a l .,A d e c a d e o f r e s e a r c h o n t h e s e c o n d m e s -s e n ge r c -d i -AM P [J ].F E M S M i c r o b i o l R e v ,2020,44(6):701-724.[35]W i t t e ,G.,e t a l .,S t r u c t u r a l b i o c h e m i s t r y of a b a c t e r i a l c h e c k p o i n t p r o t e i n r e v e a l s d i a d e n y l a t e c y c l a s e a c t i v i t y r e gu l a -t e d b y DN A r e c o m b i n a t i o n i n t e r m e d i a t e s [J ].M o l C e l l ,2008,30(2):167-78.综 述生命科学仪器 2023年第21卷/第5期14[36]R o m l i n g,U.,G r e a t t i m e s f o r s m a l l m o l e c u l e s :c-d i -AM P ,a s e c o n d m e s s e n ge r c a n d i d a t e i n B a c t e r i a a n d A r c h a e a [J ].S c i S i g n a l ,2008,1(33):pe 39.[37]Z h a n g,L .,W.L i ,a n d Z .G.H e ,D a r R ,a T e t R-l i k e t r a n s c r i p t i o n a l f a c t o r ,i s a c y c l i c d i -AM P-r e s po n s i v e r e -p r e s s o r i n M y c o b a c t e r i u m s m e gm a t i s [J ].J B i o l C h e m ,2013,288(5):3085-96.[38]C o r r i g a n ,R.M.,e t a l .,S ys t e m a t i c i d e n t i f i c a t i o n o f c o n -s e r v e d b a c t e r i a l c-d i -AM P r e c e p t o r p r o t e i n s [J ].P r o c N a t l A c a d S c i U S A ,2013,110(22):9084-9.[39]C o r r i g a n ,R.M.a n d A.G r u n d l i n g ,C yc l i cd i -AM P :a n o t h -e r s e c o n d m e s s e n g e r e n t e r s t h ef r a y [J ].N a t R e v M i c r o b i o l ,2013,11(8):513-24.[40]D a v i e s ,B .W.,e t a l .,C o o r d i n a t e d r e g u l a t i o n o f a c c e s s o r yg e n e t i c e l e m e n t s p r o d u c e s c yc l i cd i -n u c le o t i d e sf o r V.c h o l -e r a e v i r u l e n c e [J ].C e l l ,2012,149(2):358-70.[41]G a o ,J .,e t a l .,I d e n t i f i c a t i o n a n d c h a r a c t e r i z a t i o n o f p h o s -p h o d i e s t e r a s e s t h a t s p e c i f i c a l l y d eg r a d e 3'3'-c yc l i c GM P-AM P [J ].C e l l R e s ,2015,25(5):539-50.[42]S e v e r i n ,G.B .,e t a l .,D i r e c t a c t i v a t i o n o f a p h o s p h o l i pa s eb yc yc l i c GM P-AM P i n E l T o r V i b r i o c h o l e r a e [J ].P r o c N a t l A c ad S c i U S A ,2018,115(26):E 6048-E 6055.[43]C o he n ,D.,e t a l .,C y c l i c GM P-AM P s i g n a l l i n g pr o t e c t s b a c t e r i a a ga i n s t v i r a l i n f e c t i o n [J ].N a t u r e ,2019,574(7780):691-695.[44]S u n ,L .,e t a l .,C y c l i c GM P -AM P s y n t h a s e i s a c yt o s o l i c D N A s e n s o r t h a t a c t i v a t e s t h e t y p e I i n t e r f e r o n p a t h w a y [J ].S c i e n c e ,2013,339(6121):786-91.[45]W u ,J .,e t a l .,C y c l i c GM P -AM P i s a n e n d o ge n o u s s e c o n d m e s s e n g e r i n i n n a t e i mm u n e s i g n a l i n g b y c yt o s o l i c D N A [J ].S c i e n c e ,2013,339(6121):826-30.[46]I s h i k a w a ,H.a n d G.N.B a r b e r ,S T I N G i s a n e n d o pl a s m i c r e t i c u l u m a d a p t o r t h a t f a c i l i t a t e s i n n a t e i mm u n e s i g n a l l i n g[J ].N a t u r e ,2008,455(7213):674-8.[47]H o p f n e r ,K.P .a n d V.H o r n u n g,M o l e c u l a r m e c h a n i s m s a n d c e l l u l a r f u n c t i o n s o f c G A S -S T I N G s i g n a l l i n g [J ].N a t R e v M o l C e l l B i o l ,2020,21(9):501-521.[48]G a o ,P .,e t a l .,C y c l i c [G (2',5')p A (3',5')p]i s t h e m e t a z o a n s e c o n d m e s s e n g e r p r o d u c e d b y D N A-a c t i v a t e d c yc l i c GM P -AM P s yn t h a s e [J ].C e l l ,2013,153(5):1094-107.[49]H o l l e u f e r ,A.,e t a l .,T w o c G A S-l i k e r e c e pt o r s i n d u c e a n t i v i r a l i mm u n i t y i n D r o s o ph i l a [J ].N a t u r e ,2021,597(7874):114-118.[50]S l a v i k ,K.M.,e t a l .,c G A S -l i k e r e c e pt o r s s e n s e R N A a n d c o n t r o l 3'2'-c G AM P s i g n a l l i n g i n D r o s o p h i l a [J ].N a t u r e ,2021,597(7874):109-113.[51]F a t m a ,S .,e t a l .,M o l e c u l a r m e c h a n i s m s o f t h e C d n G-C a p 5a n t i p h a g e d e f e n s e s y s t e m e m p l o y i n g 3',2'-c G AM P a s t h e s e c o n d m e s s e n g e r [J ].N a t C o mm u n ,2021,12(1):6381.[52]W h i t e l e y ,A.T.,e t a l .,B a c t e r i a l c G A S -l i k e e n z y m e s s yn -t h e s i z e d i v e r s e n u c l e o t i d e s i gn a l s [J ].N a t u r e ,2019,567(7747):194-199.[53]G o v a n d e ,A.A.,e t a l .,M o l e c u l a r b a s i s o f C D-N T a s e n u -c l e o t i d e s e l e c t i o n i n C B A S S a n t i -p h a g e d e f e n s e [J ].C e l l R e p,2021,35(9):109206.[54]C h a n g ,J .J .,e t a l .,S p e c i f i c r e c o g n i t i o n o f c y c l i c o l i go n u c l e -o t i d e s b y C a p 4f o r p h a ge i nf e c t i o n [J ].I n t J B i o l M a c r o m o l ,2023,237:123656.[55]L a u ,R.K.,e t a l .,S t r u c t u r e a n d M e c h a n i s m o f a C yc l i c T r i n u c l e o t ide -A c t i v a t e d B a c t e r i a l E n d o n u c l e a s e M e d i a t i n gB a c t e r i o p h a g e I mm u n i t y [J ].M o l C e l l ,2020,77(4):723-733e 6.[56]R o u i l l o n ,C .,e t a l .,A n t i v i r a l s i g n a l l i n g b y a c yc l i c n u c l e o -t ide a c t i v a t e d C R I S P R p r o t e a s e [J ].N a t u r e ,2023,614(7946):168-174.[57]Z h a o ,R.,e t a l .,A M e m b r a n e -A s s o c i a t e d D HH-D H -H A 1N u c l e a s e D e g r a d e s T y p e I I I C R I S P R S e c o n d M e s s e n g e r [J ].C e l l R e po r t s ,2020,32(11).[58]Z h a n g,X.,X.C .B a i ,a n d Z .J .C h e n ,S t r u c t u r e s a n d M e c h a n i s m s i n t h e c G A S-S T I N G I n n a t e I mm u n i t y Pa t h -w a y [J ].I mm u n i t y,2020,53(1):43-53.[59]A t h u k o r a l a g e ,J .S .a n d M.F .W h i t e ,C y c l i c N u c l e o t i d e S i g-n a l i n g i n P h a ge D ef e n s e a n d C o u n t e r -D e f e n s e [J ].A n n u R e v V i r o l ,2022,9(1):451-468.[60]M i l l m a n ,A.,e t a l .,D i v e r s i t y a n d c l a s s i f i c a t i o n o f c yc l i c -o l i g o n u c l e o t ide -b a s e d a n t i -p h a g e s i g n a l l i n g s ys t e m s [J ].N a t M i c r o b i o l ,2020,5(12):1608-1615.[61]D u n c a n -L o w e y,B .,e t a l .,E f f e c t o r -m e d i a t e d m e m b r a n e d i s r u p t i o n c o n t r o l s c e l l d e a t h i n C B A S S a n t i p h a ge d ef e n s e [J ].M o l C e l l ,2021,81(24):5039-5051e 5.[62]Y e ,Q.,e t a l .,HO R MA D o m a i n P r o t e i n s a n d a T r i p13-l i k e A T P a s e R e g u l a t e B a c t e r i a l c G A S -l i k e E n z y m e s t o M e -d i a t e B a c t e r i o p h a g e I mm u n i t y [J ].M o l C e l l ,2020,77(4):709-722e 7.[63]D u n c a n -L o w e y ,B .a n d P .J .K r a n z u s c h ,C B A S S p h a ge d e -f e n s e a n d e v o l u t i o n o f a n t i v i r a l n u c l e o t i d e s ig n a l i n g [J ].C u r r O pi n I mm u n o l ,2022,74:156-163.[64]P a t e l ,D.J .,Y.Y u ,a n d W.X i e ,c G AM P -a c t i v a t e d c G A S-S T I N G s i g n a l i n g :i t s b a c t e r i a l o r i g i n s a n d e v o l u t i o n a r y ad -a p t a t i o n b y me t a z o a n s [J ].N a t S t r u c t M o l B i o l ,2023,30(3):245-260.[65]P a t e l ,D.J .,Y.Y u ,a n d N.J i a ,B a c t e r i a l o r i g i n s o f c yc l i c n u c l e o t ide -a c t i v a t e d a n t i v i r a l i mm u n e s i g n a l i n g [J ].M o l C e l l ,2022,82(24):4591-4610.[66]M o r e h o u s e ,B .R.,e t a l .,S T I N G c y c l i c d i n u c l e o t i d e s e n s i n go r i gi n a t e d i n b a c t e r i a [J ].N a t u r e ,2020,586(7829):429-433.[67]L i ,L .,e t a l .,H y d r o l y s i s o f 2'3'-c G AM P b y EN P P 1a n d d e s i g n o f n o n h y d r o l y z a b l e a n a l o gs [J ].N a t C h e m B i o l ,2014,10(12):1043-8.。
中华人民共和国进境植物检疫性有害生物名录

附件:中华人民共和国进境植物检疫性有害生物名录昆虫1.Acanthocinus carinulatus (Gebler)白带长角天牛2.Acanthoscelides obtectus (Say)菜豆象3.Acleris variana (Fernald)黑头长翅卷蛾4.Agrilus spp. (non-Chinese)窄吉丁(非中国种)5.Aleurodicus dispersus Russell螺旋粉虱6.Anastrepha Schiner按实蝇属7.Anthonomus grandis Boheman墨西哥棉铃象8.Anthonomus quadrigibbus Say苹果花象9.Aonidiella comperei McKenzie香蕉肾盾蚧10.Apate monachus Fabricius咖啡黑长蠹11.Aphanostigma piri (Cholodkovsky)梨矮蚜12.Arhopalus syriacus Reitter辐射松幽天牛13.Bactrocera Macquart果实蝇属14.Baris granulipennis (Tournier)西瓜船象15.Batocera spp. (non-Chinese)白条天牛(非中国种)16.Brontispa longissima (Gestro)椰心叶甲17.Bruchidius incarnates (Boheman)埃及豌豆象18.Bruchophagus roddi Gussak苜蓿籽蜂19.Bruchus spp. (non-Chinese )豆象(属)(非中国种)20.Cacoecimorpha pronubana (Hübner)荷兰石竹卷蛾21.Callosobruchus spp. (maculatus(F.)and non-Chinese)瘤背豆象(四纹豆象和非中国种)22.Carpomya incompleta (Becker)欧非枣实蝇23.Carpomya vesuviana Costa枣实蝇24.Carulaspis juniperi (Bouchè)松唐盾蚧25.Caulophilus oryzae (Gyllenhal)阔鼻谷象26.Ceratitis Macleay小条实蝇属27.Ceroplastes rusci (L.)无花果蜡蚧28.Chionaspis pinifoliae (Fitch)松针盾蚧29.Choristoneura fumiferana (Clemens)云杉色卷蛾30.Conotrachelus Schoenherr鳄梨象属31.Contarinia sorghicola (Coquillett)高粱瘿蚊32.Coptotermes spp. (non-Chinese)乳白蚁(非中国种)33.Craponius inaequalis (Say)葡萄象34.Crossotarsus spp. (non-Chinese)异胫长小蠹(非中国种)35.Cryptophlebia leucotreta (Meyrick)苹果异形小卷蛾36.Cryptorrhynchus lapathi L.杨干象37.Cryptotermes brevis (Walker)麻头砂白蚁38.Ctenopseustis obliquana (Walker)斜纹卷蛾39.Curculio elephas (Gyllenhal)欧洲栗象40.Cydia janthinana (Duponchel)山楂小卷蛾41.Cydia packardi (Zeller)樱小卷蛾42.Cydia pomonella (L.)苹果蠹蛾43.Cydia prunivora (Walsh)杏小卷蛾44.Cydia pyrivora (Danilevskii)梨小卷蛾45.Dacus spp. (non-Chinese)寡鬃实蝇(非中国种)46.Dasineura mali (Kieffer)苹果瘿蚊47.Dendroctonus spp. (valens LeConte and non-Chinese)大小蠹(红脂大小蠹和非中国种)48.Deudorix isocrates Fabricius石榴小灰蝶49.Diabrotica Chevrolat根萤叶甲属50.Diaphania nitidalis (Stoll)黄瓜绢野螟51.Diaprepes abbreviata (L.)蔗根象52.Diatraea saccharalis (Fabricius)小蔗螟53.Dryocoetes confusus Swaine混点毛小蠹54.Dysmicoccus grassi Leonari香蕉灰粉蚧55.Dysmicoccus neobrevipes Beardsley新菠萝灰粉蚧56.Ectomyelois ceratoniae (Zeller)石榴螟57.Epidiaspis leperii (Signoret)桃白圆盾蚧58.Eriosoma lanigerum(Hausmann)苹果绵蚜59.Eulecanium gigantea (Shinji)枣大球蚧60.Eurytoma amygdali Enderlein扁桃仁蜂61.Eurytoma schreineri Schreiner李仁蜂62.Gonipterus scutellatus Gyllenhal桉象63.Helicoverpa zea (Boddie)谷实夜蛾64.Hemerocampa leucostigma (Smith)合毒蛾65.Hemiberlesia pitysophila Takagi松突圆蚧66.Heterobostrychus aequalis (Waterhouse)双钩异翅长蠹67.Hoplocampa flava (L.)李叶蜂68.Hoplocampa testudinea (Klug)苹叶蜂69.Hoplocerambyx spinicornis (Newman)刺角沟额天牛70.Hylobius pales (Herbst)苍白树皮象71.Hylotrupes bajulus (L.)家天牛72.Hylurgopinus rufipes (Eichhoff)美洲榆小蠹73.Hylurgus ligniperda Fabricius长林小蠹74.Hyphantria cunea (Drury)美国白蛾75.Hypothenemus hampei (Ferrari)咖啡果小蠹76.Incisitermes minor (Hagen)小楹白蚁77.Ips spp. (non-Chinese)齿小蠹(非中国种)78.Ischnaspis longirostris (Signoret)黑丝盾蚧79.Lepidosaphes tapleyi Williams芒果蛎蚧80.Lepidosaphes tokionis (Kuwana)东京蛎蚧81.Lepidosaphes ulmi (L.)榆蛎蚧82.Leptinotarsa decemlineata (Say)马铃薯甲虫83.Leucoptera coffeella (Guérin-Méneville)咖啡潜叶蛾84.Liriomyza trifolii (Burgess)三叶斑潜蝇85.Lissorhoptrus oryzophilus Kuschel稻水象甲86.Listronotus bonariensis (Kuschel)阿根廷茎象甲87.Lobesia botrana (Denis et Schiffermuller)葡萄花翅小卷蛾88.Mayetiola destructor (Say)黑森瘿蚊89.Mercetaspis halli (Green)霍氏长盾蚧90.Monacrostichus citricola Bezzi桔实锤腹实蝇91.Monochamus spp. (non-Chinese)墨天牛(非中国种)92.Myiopardalis pardalina (Bigot)甜瓜迷实蝇93.Naupactus leucoloma (Boheman)白缘象甲94.Neoclytus acuminatus (Fabricius)黑腹尼虎天牛95.Opogona sacchari (Bojer)蔗扁蛾96.Pantomorus cervinus (Boheman)玫瑰短喙象97.Parlatoria crypta Mckenzie灰白片盾蚧98.Pharaxonotha kirschi Reither谷拟叩甲99.Phloeosinus cupressi Hopkins美柏肤小蠹100.Phoracantha semipunctata (Fabricius) 桉天牛101.Pissodes Germar木蠹象属102.Planococcus lilacius Cockerell南洋臀纹粉蚧103.Planococcus minor (Maskell)大洋臀纹粉蚧104.Platypus spp. (non-Chinese)长小蠹(属)(非中国种)105.Popillia japonica Newman日本金龟子106.Prays citri Milliere桔花巢蛾107.Promecotheca cumingi Baly椰子缢胸叶甲108.Prostephanus truncatus (Horn)大谷蠹109.Ptinus tectus Boieldieu澳洲蛛甲110.Quadrastichus erythrinae Kim刺桐姬小蜂111.Reticulitermes lucifugus(Rossi)欧洲散白蚁112.Rhabdoscelus lineaticollis (Heller) 褐纹甘蔗象113.Rhabdoscelus obscurus (Boisduval) 几内亚甘蔗象114.Rhagoletis spp. (non-Chinese)绕实蝇(非中国种)115.Rhynchites aequatus (L.)苹虎象116.Rhynchites bacchus L.欧洲苹虎象117.Rhynchites cupreus L.李虎象118.Rhynchites heros Roelofs日本苹虎象119.Rhynchophorus ferrugineus (Olivier) 红棕象甲120.Rhynchophorus palmarum (L.)棕榈象甲121.Rhynchophorus phoenicis (Fabricius) 紫棕象甲122.Rhynchophorus vulneratus (Panzer) 亚棕象甲123.Sahlbergella singularis Haglund可可盲蝽象124.Saperda spp. (non-Chinese)楔天牛(非中国种)125.Scolytus multistriatus (Marsham)欧洲榆小蠹126.Scolytus scolytus (Fabricius)欧洲大榆小蠹127.Scyphophorus acupunctatus Gyllenhal剑麻象甲128.Selenaspidus articulatus Morgan刺盾蚧129.Sinoxylon spp. (non-Chinese)双棘长蠹(非中国种)130.Sirex noctilio Fabricius云杉树蜂131.Solenopsis invicta Buren红火蚁132.Spodoptera littoralis(Boisduval)海灰翅夜蛾133.Stathmopoda skelloni Butler猕猴桃举肢蛾134.Sternochetus Pierce芒果象属135.Taeniothrips inconsequens (Uzel)梨蓟马136.Tetropium spp. (non-Chinese)断眼天牛(非中国种)137.Thaumetopoea pityocampa (Denis et Schiffermuller) 松异带蛾138.Toxotrypana curvicauda Gerstaecker番木瓜长尾实蝇139.Tribolium destructor Uyttenboogaart褐拟谷盗140.Trogoderma spp. (non-Chinese)斑皮蠹(非中国种)141.Vesperus Latreile暗天牛属142.Vinsonia stellifera (Westwood)七角星蜡蚧143.Viteus vitifoliae (Fitch)葡萄根瘤蚜144.Xyleborus spp. (non-Chinese)材小蠹(非中国种)145.Xylotrechus rusticus L.青杨脊虎天牛146.Zabrotes subfasciatus (Boheman)巴西豆象软体动物147.Achatina fulica Bowdich非洲大蜗牛148.Acusta despecta Gray硫球球壳蜗牛149.Cepaea hortensis Müller花园葱蜗牛150.Helix aspersa Müller散大蜗牛151.Helix pomatia Linnaeus盖罩大蜗牛152.Theba pisana Müller比萨茶蜗牛真菌153.Albugo tragopogi (Persoon) Schröter var. helianthi Novotelnova 向日葵白锈病菌154.Alternaria triticina Prasada et Prabhu小麦叶疫病菌155.Anisogramma anomala(Peck)E. Muller榛子东部枯萎病菌156.Apiosporina morbosa (Schweinitz) von Arx李黑节病菌157.Atropellis pinicola Zaller et Goodding松生枝干溃疡病菌158.Atropellis piniphila (Weir) Lohman et Cash嗜松枝干溃疡病菌159.Botryosphaeria laricina (K.Sawada) Y.Zhong落叶松枯梢病菌160.Botryosphaeria stevensii Shoemaker苹果壳色单隔孢溃疡病菌161.Cephalosporium gramineum Nisikado et Ikata麦类条斑病菌162.Cephalosporium maydis Samra, Sabet et Hingorani玉米晚枯病菌163.Cephalosporium sacchari E.J. Butler et Hafiz Khan甘蔗凋萎病菌164.Ceratocystis fagacearum (Bretz) Hunt栎枯萎病菌165.Chrysomyxa arctostaphyli Dietel云杉帚锈病菌166.Ciborinia camelliae Kohn山茶花腐病菌167.Cladosporium cucumerinum Ellis et Arthur黄瓜黑星病菌168.Colletotrichum kahawae J.M. Waller et Bridge咖啡浆果炭疽病菌169.Crinipellis perniciosa (Stahel) Singer可可丛枝病菌170.Cronartium coleosporioides J.C.Arthur油松疱锈病菌171.Cronartium comandrae Peck北美松疱锈病菌172.Cronartium conigenum Hedgcock et Hunt松球果锈病菌173.Cronartium fusiforme Hedgcock et Hunt ex Cummins 松纺锤瘤锈病菌174.Cronartium ribicola J.C.Fisch.松疱锈病菌175.Cryphonectria cubensis (Bruner) Hodges桉树溃疡病菌176.Cylindrocladium parasiticum Crous, Wingfield et Alfenas 花生黑腐病菌177.Diaporthe helianthi Muntanola-Cvetkovic Mihaljcevic et Petrov 向日葵茎溃疡病菌178.Diaporthe perniciosa É.J. Marchal苹果果腐病菌179.Diaporthe phaseolorum (Cooke et Ell.) Sacc. var. caulivora Athow et Caldwell大豆北方茎溃疡病菌180.Diaporthe phaseolorum (Cooke et Ell.) Sacc. var. meridionalis F.A.Fernandez大豆南方茎溃疡病菌181.Diaporthe vaccinii Shear蓝莓果腐病菌182.Didymella ligulicola (K.F.Baker, Dimock et L.H.Davis) von Arx 菊花花枯病菌183.Didymella lycopersici Klebahn番茄亚隔孢壳茎腐病菌184.Endocronartium harknessii (J.P.Moore) Y.Hiratsuka松瘤锈病菌185.Eutypa lata (Pers.) Tul. et C. Tul.葡萄藤猝倒病菌186.Fusarium circinatum Nirenberg et O'Donnell松树脂溃疡病菌187.Fusarium oxysporum Schlecht. f.sp. apii Snyd. et Hans芹菜枯萎病菌188.Fusarium oxysporum Schlecht. f.sp. asparagi Cohen et Heald 芦笋枯萎病菌189.Fusarium oxysporum Schlecht. f.sp. cubense (E.F.Sm.) Snyd.et Hans (Race 4 non-Chinese races)香蕉枯萎病菌(4号小种和非中国小种)190.Fusarium oxysporum Schlecht. f.sp. elaeidis Toovey油棕枯萎病菌191.Fusarium oxysporum Schlecht. f.sp. fragariae Winks et Williams 草莓枯萎病菌192.Fusarium tucumaniae T.Aoki,O’Donnell, Yos.Homma et Lat tanzi 南美大豆猝死综合症病菌193.Fusarium virguliforme O’Donnell et T.Aoki北美大豆猝死综合症病菌194.Gaeumannomyces graminis (Sacc.) Arx et D. Olivier var. avenae(E.M. Turner) Dennis燕麦全蚀病菌195.Greeneria uvicola (Berk. et M.A.Curtis) Punithalingam 葡萄苦腐病菌196.Gremmeniella abietina (Lagerberg) Morelet冷杉枯梢病菌197.Gymnosporangium clavipes (Cooke et Peck) Cooke et Peck 榲桲锈病菌198.Gymnosporangium fuscum R. Hedw.欧洲梨锈病菌199.Gymnosporangium globosum (Farlow) Farlow美洲山楂锈病菌200.Gymnosporangium juniperi-virginianae Schwein美洲苹果锈病菌201.Helminthosporium solani Durieu et Mont.马铃薯银屑病菌202.Hypoxylon mammatum (Wahlenberg) J. Miller杨树炭团溃疡病菌203.Inonotus weirii (Murrill) Kotlaba et Pouzar松干基褐腐病菌204.Leptosphaeria libanotis (Fuckel) Sacc.胡萝卜褐腐病菌205.Leptosphaeria maculans (Desm.) Ces. et De Not.十字花科蔬菜黑胫病菌206.Leucostoma cincta (Fr.:Fr.) Hohn.苹果溃疡病菌207.Melampsora farlowii (J.C.Arthur) J.J.Davis铁杉叶锈病菌208.Melampsora medusae Thumen杨树叶锈病菌209.Microcyclus ulei (P.Henn.) von Arx橡胶南美叶疫病菌210.Monilinia fructicola (Winter) Honey美澳型核果褐腐病菌211.Moniliophthora roreri (Ciferri et Parodi) Evans可可链疫孢荚腐病菌212.Monosporascus cannonballus Pollack et Uecker甜瓜黑点根腐病菌213.Mycena citricolor (Berk. et Curt.) Sacc.咖啡美洲叶斑病菌214.Mycocentrospora acerina (Hartig) Deighton香菜腐烂病菌215.Mycosphaerella dearnessii M.E.Barr松针褐斑病菌216.Mycosphaerella fijiensis Morelet香蕉黑条叶斑病菌217.Mycosphaerella gibsonii H.C.Evans松针褐枯病菌218.Mycosphaerella linicola Naumov亚麻褐斑病菌219.Mycosphaerella musicola J.L.Mulder香蕉黄条叶斑病菌220.Mycosphaerella pini E.Rostrup松针红斑病菌221.Nectria rigidiuscula Berk.et Broome可可花瘿病菌222.Ophiostoma novo-ulmi Brasier新榆枯萎病菌223.Ophiostoma ulmi (Buisman) Nannf.榆枯萎病菌224.Ophiostoma wageneri (Goheen et Cobb) Harrington针叶松黑根病菌225.Ovulinia azaleae Weiss杜鹃花枯萎病菌226.Periconia circinata(M.Mangin)Sacc.高粱根腐病菌227.Peronosclerospora spp. (non-Chinese)玉米霜霉病菌(非中国种)228.Peronospora farinosa (Fries: Fries) Fries f.sp. betae Byford 甜菜霜霉病菌229.Peronospora hyoscyami de Bary f.sp. tabacina (Adam) Skalicky 烟草霜霉病菌230.Pezicula malicorticis (Jacks.) Nannfeld苹果树炭疽病菌231.Phaeoramularia angolensis (T.Carvalho et O. Mendes)P.M. Kirk 柑橘斑点病菌232.Phellinus noxius (Corner) G.H.Cunn.木层孔褐根腐病菌233.Phialophora gregata (Allington et Chamberlain) W.Gams 大豆茎褐腐病菌234.Phialophora malorum (Kidd et Beaum.) McColloch苹果边腐病菌235.Phoma exigua Desmazières f.sp. foveata (Foister) Boerema 马铃薯坏疽病菌236.Phoma glomerata (Corda) Wollenweber et Hochapfel 葡萄茎枯病菌237.Phoma pinodella (L.K. Jones) Morgan-Jones et K.B. Burch 豌豆脚腐病菌238.Phoma tracheiphila (Petri) L.A. Kantsch. et Gikaschvili 柠檬干枯病菌239.Phomopsis sclerotioides van Kesteren黄瓜黑色根腐病菌240.Phymatotrichopsis omnivora (Duggar) Hennebert棉根腐病菌241.Phytophthora cambivora (Petri) Buisman栗疫霉黑水病菌242.Phytophthora erythroseptica Pethybridge马铃薯疫霉绯腐病菌243.Phytophthora fragariae Hickman草莓疫霉红心病菌244.Phytophthora fragariae Hickman var. rubi W.F. Wilcox et J.M.Duncan树莓疫霉根腐病菌245.Phytophthora hibernalis Carne柑橘冬生疫霉褐腐病菌246.Phytophthora lateralis Tucker et Milbrath雪松疫霉根腐病菌247.Phytophthora medicaginis E.M. Hans. et D.P. Maxwell苜蓿疫霉根腐病菌248.Phytophthora phaseoli Thaxter菜豆疫霉病菌249.Phytophthora ramorum Werres, De Cock et Man in’t Vel d 栎树猝死病菌250.Phytophthora sojae Kaufmann et Gerdemann大豆疫霉病菌251.Phytophthora syringae (Klebahn) Klebahn丁香疫霉病菌252.Polyscytalum pustulans (M.N. Owen et Wakef.) M.B.Ellis 马铃薯皮斑病菌253.Protomyces macrosporus Unger香菜茎瘿病菌254.Pseudocercosporella herpotrichoides (Fron) Deighton小麦基腐病菌255.Pseudopezicula tracheiphila (Müller-Thurgau) Korf et Zhuang 葡萄角斑叶焦病菌256.Puccinia pelargonii-zonalis Doidge天竺葵锈病菌257.Pycnostysanus azaleae (Peck) Mason杜鹃芽枯病菌258.Pyrenochaeta terrestris (Hansen) Gorenz, Walker et Larson 洋葱粉色根腐病菌259.Pythium splendens Braun油棕猝倒病菌260.Ramularia beticola Fautr. et Lambotte甜菜叶斑病菌261.Rhizoctonia fragariae Husain et W.E.McKeen草莓花枯病菌262.Rigidoporus lignosus (Klotzsch) Imaz.橡胶白根病菌263.Sclerophthora rayssiae Kenneth, Kaltin et Wahl var. zeae Payak et Renfro玉米褐条霜霉病菌264.Septoria petroselini (Lib.) Desm.欧芹壳针孢叶斑病菌265.Sphaeropsis pyriputrescens Xiao et J. D. Rogers苹果球壳孢腐烂病菌266.Sphaeropsis tumefaciens Hedges柑橘枝瘤病菌267.Stagonospora avenae Bissett f. sp. triticea T. Johnson 麦类壳多胞斑点病菌268.Stagonospora sacchari Lo et Ling甘蔗壳多胞叶枯病菌269.Synchytrium endobioticum (Schilberszky) Percival马铃薯癌肿病菌270.Thecaphora solani (Thirumalachar et M.J.O’Brien) Mordue 马铃薯黑粉病菌271.Tilletia controversa Kühn小麦矮腥黑穗病菌272.Tilletia indica Mitra小麦印度腥黑穗病菌273.Urocystis cepulae Frost葱类黑粉病菌274.Uromyces transversalis (Thümen) Winter唐菖蒲横点锈病菌275.Venturia inaequalis (Cooke) Winter苹果黑星病菌276.Verticillium albo-atrum Reinke et Berthold苜蓿黄萎病菌277.Verticillium dahliae Kleb.棉花黄萎病菌原核生物278.Acidovorax avenae subsp. cattleyae ( Pavarino) Willems et al.兰花褐斑病菌279.Acidovorax avenae subsp. citrulli ( Schaad et al. ) Willems et al.瓜类果斑病菌280.Acidovorax konjaci ( Goto ) Willems et al.魔芋细菌性叶斑病菌281.Alder yellows phytoplasma桤树黄化植原体282.Apple proliferation phytoplasma苹果丛生植原体283.Apricot chlorotic leafroll phtoplasma杏褪绿卷叶植原体284.Ash yellows phytoplasma白蜡树黄化植原体285.Blueberry stunt phytoplasma蓝莓矮化植原体286.Burkholderia caryophylli ( Burkholder ) Yabuuchi et al.香石竹细菌性萎蔫病菌287.Burkholderia gladioli pv. alliicola ( Burkholder ) Urakami et al.洋葱腐烂病菌288.Burkholderia glumae ( Kurita et Tabei ) Urakami et al.水稻细菌性谷枯病菌289.Candidatus Liberobacter africanum Jagoueix et al.非洲柑桔黄龙病菌290.Candidatus Liberobacter asiaticum Jagoueix et al.亚洲柑桔黄龙病菌291.Candidatus Phytoplasma australiense澳大利亚植原体候选种292.Clavibacter michiganensis subsp. insidiosus ( McCulloch ) Davis et al.苜蓿细菌性萎蔫病菌293.Clavibacter michiganensis subsp. michiganensis ( Smith ) Davis et al.番茄溃疡病菌294.Clavibacter michiganensis subsp. nebraskensis ( Vidaver et al. ) Davis et al.玉米内州萎蔫病菌295.Clavibacter michiganensis subsp. sepedonicus( Spieckermann et al. ) Davis et al.马铃薯环腐病菌296.Coconut lethal yellowing phytoplasma椰子致死黄化植原体297.Curtobacterium flaccumfaciens pv. flaccumfaciens( Hedges ) Collins et Jones菜豆细菌性萎蔫病菌298.Curtobacterium flaccumfaciens pv. oortii ( Saaltink et al. ) Collins et Jones郁金香黄色疱斑病菌299.Elm phloem necrosis phytoplasma榆韧皮部坏死植原体300.Enterobacter cancerogenus ( Urosevi ) Dickey et Zumoff 杨树枯萎病菌301.Erwinia amylovora ( Burrill ) Winslow et al.梨火疫病菌302.Erwinia chrysanthemi Burkhodler et al.菊基腐病菌303.Erwinia pyrifoliae Kim, Gardan, Rhim et Geider亚洲梨火疫病菌304.Grapevine flavescence dorée phytoplasma葡萄金黄化植原体305.Lime witches’ broom phytoplasma来檬丛枝植原体306.Pantoea stewartii subsp. stewartii ( Smith ) Mergaert et al.玉米细菌性枯萎病菌307.Peach X-disease phytoplasma桃X病植原体308.Pear decline phytoplasma梨衰退植原体309.Potato witches’ broom phytoplasma马铃薯丛枝植原体310.Pseudomonas savastanoi pv. phaseolicola ( Burkholder ) Gardan et al.菜豆晕疫病菌311.Pseudomonas syringae pv. morsprunorum( Wormald ) Young et al.核果树溃疡病菌312.Pseudomonas syringae pv. persicae ( Prunier et al. ) Young et al.桃树溃疡病菌313.Pseudomonas syringae pv. pisi ( Sackett ) Young et al.豌豆细菌性疫病菌314.Pseudomonas syringae pv. maculicola (McCulloch) Young et al十字花科黑斑病菌315.Pseudomonas syringae pv. tomato ( Okabe ) Young et al.番茄细菌性叶斑病菌316.Ralstonia solanacearum ( Smith ) Yabuuchi et al.(race 2)香蕉细菌性枯萎病菌(2号小种)317.Rathayibacter rathayi ( Smith ) Zgurskaya et al.鸭茅蜜穗病菌318.Spiroplasma citri Saglio et al.柑橘顽固病螺原体319.Strawberry multiplier phytoplasma草莓簇生植原体320.Xanthomonas albilineans ( Ashby ) Dowson甘蔗白色条纹病菌321.Xanthomonas arboricola pv. celebensis ( Gaumann ) Vauterin et al.香蕉坏死条纹病菌322.Xanthomonas axonopodis pv. betlicola ( Patel et al. ) Vauterin et al.胡椒叶斑病菌323.Xanthomonas axonopodis pv. citri ( Hasse ) Vauterin et al.柑橘溃疡病菌324.Xanthomonas axonopodis pv. manihotis ( Bondar ) Vauterin et al.木薯细菌性萎蔫病菌325.Xanthomonas axonopodis pv. vasculorum ( Cobb ) Vauterin et al.甘蔗流胶病菌326.Xanthomonas campestris pv. mangiferaeindicae ( Patel et al. ) Robbs et al.芒果黑斑病菌327.Xanthomonas campestris pv. musacearum( Yirgou et Bradbury ) Dye香蕉细菌性萎蔫病菌328.Xanthomonas cassavae ( ex Wiehe et Dowson ) Vauterin et al.木薯细菌性叶斑病菌329.Xanthomonas fragariae Kennedy et King草莓角斑病菌330.Xanthomonas hyacinthi ( Wakker ) Vauterin et al.风信子黄腐病菌331.Xanthomonas oryzae pv. oryzae ( Ishiyama ) Swings et al.水稻白叶枯病菌332.Xanthomonas oryzae pv. oryzicola ( Fang et al. ) Swings et al.水稻细菌性条斑病菌333.Xanthomonas populi ( ex Ride ) Ride et Ride杨树细菌性溃疡病菌334.Xylella fastidiosa Wells et al.木质部难养细菌335.Xylophilus ampelinus ( Panagopoulos ) Willems et al.葡萄细菌性疫病菌线虫336.Anguina agrostis (Steinbuch) Filipjev剪股颖粒线虫337.Aphelenchoides fragariae (Ritzema Bos) Christie草莓滑刃线虫338.Aphelenchoides ritzemabosi (Schwartz) Steiner et Bührer 菊花滑刃线虫339.Bursaphelenchus cocophilus (Cobb) Baujard椰子红环腐线虫340.Bursaphelenchus xylophilus (Steiner et Bührer ) Nickle 松材线虫341.Ditylenchus angustus (Butler ) Filipjev水稻茎线虫342.Ditylenchus destructor Thorne腐烂茎线虫343.Ditylenchus dipsaci (Kühn ) Filipjev鳞球茎茎线虫344.Globodera pallida (Stone ) Behrens马铃薯白线虫345.Globodera rostochiensis (Wollenweber ) Behrens马铃薯金线虫346.Heterodera schachtii Schmidt甜菜胞囊线虫347.Longidorus (Filipjev) Micoletzky(The species transmit viruses)长针线虫属(传毒种类)348.Meloidogyne Goeldi (non-Chinese species)根结线虫属(非中国种)349.Nacobbus abberans (Thorne ) Thorne et Allen异常珍珠线虫350.Paralongidorus maximus (Bütschli ) Siddiqi最大拟长针线虫351.Paratrichodorus Siddiqi (The species transmit viruses)拟毛刺线虫属(传毒种类)352.Pratylenchus Filipjev (non-Chinese species)短体线虫(非中国种)353.Radopholus similis (Cobb) Thorne香蕉穿孔线虫354.Trichodorus Cobb (The species transmit viruses)毛刺线虫属(传毒种类)355.Xiphinema Cobb (The species transmit viruses)剑线虫属(传毒种类)病毒及类病毒356.African cassava mosaic virus, ACMV非洲木薯花叶病毒(类)357.Apple stem grooving virus, ASPV苹果茎沟病毒358.Arabis mosaic virus, ArMV南芥菜花叶病毒359.Banana bract mosaic virus, BBrMV香蕉苞片花叶病毒360.Bean pod mottle virus, BPMV菜豆荚斑驳病毒361.Broad bean stain virus, BBSV蚕豆染色病毒362.Cacao swollen shoot virus, CSSV可可肿枝病毒363.Carnation ringspot virus, CRSV香石竹环斑病毒364.Cotton leaf crumple virus, CLCrV棉花皱叶病毒365.Cotton leaf curl virus, CLCuV棉花曲叶病毒366.Cowpea severe mosaic virus, CPSMV豇豆重花叶病毒367.Cucumber green mottle mosaic virus, CGMMV 黄瓜绿斑驳花叶病毒368.Maize chlorotic dwarf virus, MCDV玉米褪绿矮缩病毒369.Maize chlorotic mottle virus, MCMV玉米褪绿斑驳病毒370.Oat mosaic virus, OMV燕麦花叶病毒371.Peach rosette mosaic virus, PRMV桃丛簇花叶病毒372.Peanut stunt virus, PSV花生矮化病毒373.Plum pox virus, PPV李痘病毒374.Potato mop-top virus, PMTV马铃薯帚顶病毒375.Potato virus A, PV A马铃薯A病毒376.Potato virus V, PVV马铃薯V病毒377.Potato yellow dwarf virus, PYDV马铃薯黄矮病毒378.Prunus necrotic ringspot virus, PNRSV李属坏死环斑病毒379.Southern bean mosaic virus, SBMV南方菜豆花叶病毒380.Sowbane mosaic virus, SoMV藜草花叶病毒381.Strawberry latent ringspot virus, SLRSV 草莓潜隐环斑病毒382.Sugarcane streak virus, SSV甘蔗线条病毒383.Tobacco ringspot virus, TRSV烟草环斑病毒384.Tomato black ring virus, TBRV番茄黑环病毒385.Tomato ringspot virus, ToRSV番茄环斑病毒386.Tomato spotted wilt virus, TSWV番茄斑萎病毒387.Wheat streak mosaic virus, WSMV 小麦线条花叶病毒388.Apple fruit crinkle viroid, AFCVd苹果皱果类病毒389.Avocado sunblotch viroid, ASBVd鳄梨日斑类病毒390.Coconut cadang-cadang viroid, CCCVd 椰子死亡类病毒391.Coconut tinangaja viroid, CTiVd椰子败生类病毒392.Hop latent viroid, HLVd啤酒花潜隐类病毒393.Pear blister canker viroid, PBCVd梨疱症溃疡类病毒394.Potato spindle tuber viroid, PSTVd 马铃薯纺锤块茎类病毒杂草395.Aegilops cylindrica Horst具节山羊草396.Aegilops squarrosa L.节节麦397.Ambrosia spp.豚草(属)398.Ammi majus L.大阿米芹399.Avena barbata Brot.细茎野燕麦400.Avena ludoviciana Durien法国野燕麦401.Avena sterilis L.不实野燕麦402.Bromus rigidus Roth硬雀麦403.Bunias orientalis L.疣果匙荠404.Caucalis latifolia L.宽叶高加利405.Cenchrus spp. (non-Chinese species) 蒺藜草(属)(非中国种)406.Centaurea diffusa Lamarck铺散矢车菊407.Centaurea repens L.匍匐矢车菊408.Crotalaria spectabilis Roth美丽猪屎豆409.Cuscuta spp.菟丝子(属)410.Emex australis Steinh.南方三棘果411.Emex spinosa (L.) Campd.刺亦模412.Eupatorium adenophorum Spreng.紫茎泽兰413.Eupatorium odoratum L.飞机草414.Euphorbia dentata Michx.齿裂大戟415.Flaveria bidentis (L.) Kuntze黄顶菊416.Ipomoea pandurata (L.) G.F.W.Mey.提琴叶牵牛花417.Iva axillaris Pursh小花假苍耳418.Iva xanthifolia Nutt.假苍耳419.Knautia arvensis (L.) Coulter欧洲山萝卜ctuca pulchella (Pursh) DC.野莴苣ctuca serriola L.毒莴苣422.Lolium temulentum L.毒麦423.Mikania micrantha Kunth薇甘菊424.Orobanche spp.列当(属)425.Oxalis latifolia Kubth宽叶酢浆草426.Senecio jacobaea L.臭千里光427.Solanum carolinense L.北美刺龙葵428.Solanum elaeagnifolium Cay.银毛龙葵429.Solanum rostratum Dunal.刺萼龙葵430.Solanum torvum Swartz刺茄431.Sorghum almum Parodi.黑高粱432.Sorghum halepense (L.) Pers. (Johnsongrass and its cross breeds) 假高粱(及其杂交种)433.Striga spp. (non-Chinese species)独脚金(属)(非中国种)434.Tribulus alatus Delile翅蒺藜435.Xanthium spp. (non-Chinese species)苍耳(属)(非中国种)备注1:非中国种是指中国未有发生的种;备注2:非中国小种是指中国未有发生的小种;备注3:传毒种类是指可以作为植物病毒传播介体的线虫种类。
部分水果细菌病害介绍

中国柑桔(mandarin)、橘柚(tangelo)、柚子 (grapefruit)、枳橙(citrange)、酸橙(sour orange)、柠
檬(lemon)、山葵(horseradish)等植物。
地理分布: 亚洲: 以色列、土耳其、伊朗等;非洲:摩洛哥、突尼斯、 苏丹等;北美洲:墨西哥、美国(加州、华盛顿、马里兰等州); 南美洲:阿根廷、巴西、秘鲁;欧洲:塞浦路斯、法国、西班牙;
李痘病毒
受李痘病毒侵染的李
受李痘病毒侵染的桃 受李痘病毒侵染的杏
Symptoms of 'stubborn disease' on Navel oranges; note small, asymmetric, unevenly coloured fruits. The largest fruit is healthy
四、葡萄细菌性疫病
病原菌:
Spiroplasma citri Saglio et al., 1973
三、柑橘顽固病螺原体
病原菌:
Spiroplasma citri Saglio et al., 1973
寄主:
中国柑桔(mandarin)、橘柚(tangelo)、柚子 (grapefruit)、枳橙(citrange)、酸橙(sour orange)、柠
檬(lemon)、山葵(horseradish)等植物。
一、梨火疫病
Fire blight canker on apple trunk
Milky ooze containing fire blight bacteria
深圳出入境检验检疫局
SHENZHEN ENTRY-EXIT INSPECTION AND QUARANTINE BUREAU
亚洲梨火疫病致病相关因子的研究进展

亚洲梨火疫病致病相关因子的研究进展龙海 杨伟东 陈枝楠 郑耘 王颖(深圳出入境检验检疫局 518001)本课题受科技部国家科技支撑计划课题项目(2006BAK10B01-1-1)资助。
收稿日期:2008-06-10摘要 本文概述了亚洲梨火疫病(E r w inia pyrifoliae )的生物学特性、危害症状、寄主范围、分布情况,重点介绍了该病菌的微生物学、生理生化、分子生物学特点,以及一些致病相关因子的研究进展。
同时,分析比较了亚洲梨火疫病菌和梨火疫病菌以及日本梨枝枯病菌的关系,指出了亚洲梨火疫病菌传入我国的风险以及在检疫上的重要性。
关键词 亚洲梨火疫病 梨火疫病 致病相关因子 过敏反应和致病性 胞外多糖中图分类号 S41-30,S432.4-34 亚洲梨火疫病,又名亚洲枯枝疫病、亚洲梨枯死病、梨枝条黑枯病,是亚洲梨上的一种毁灭性病害,其病原为E r w in ia pyrifoliae ,已被我国列入《中华人民共和国进境植物检疫性有害生物名录》。
该病最早在1995年发现于韩国Chuncheon 地区的亚洲梨(Pyrus pyrifolia )树上,1995~1998年连续发病流行,其病原特点、危害症状等均与梨火疫病(E r 2w in ia am ylovora )类似。
近年来,为了促进经济发展,满足国内市场的需求,我国相继从韩国、日本等地大量引进优良梨、苹果苗木及果实。
因此,亚洲梨火疫病已经对我国果品生产和进出境水果贸易构成潜在威胁。
本文将对亚洲梨火疫病的研究现状进行分析。
1 生物学特征1.1 症状亚洲梨火疫病可以危害梨树的树干、枝条、花序、幼果和叶片,发病迅速,危害严重[1]。
发病梨树主要从花、叶、嫩枝开始发病,叶片上产生褐色至黑色病斑,叶片中脉产生黑至褐色条斑,病斑可扩展至整个枝条,发病枝条几天后迅速变色枯死,一个果园中的大部分果树都会出现相同症状,因此造成严重减产[1]。
亚洲梨火疫病的危害症状与梨火疫病的非常相似,但又不完全相同。
链霉素防治植物细菌性病害的历史与研究现状

链霉素防治植物细菌性病害的历史与研究现状作者:李文红,程英,金剑雪,等来源:《湖北农业科学》 2013年第9期李文红,程英,金剑雪,李凤良(贵州省植物保护研究所,贵阳550006)摘要:综述了近年来国内外链霉素防治植物细菌性病害的主要成果,包括链霉素的发现历史、链霉素在植物细菌性病害防治上的应用及其作用机理、植物病原细菌对链霉素的耐药性以及链霉素的耐药性机制。
关键词:链霉素;植物细菌性病害;化学防治;作用机制;耐药性中图分类号:S482.2;S43文献标识码:A文章编号:0439-8114(2013)09-1985-03HistoryandResearchAdvanceofStreptomycinforControllingPlantBacterialDiseasesLIWen-hong,CHENGYing,JINJian-xue,LIFeng-liang(GuizhouInstituteofPlantProtection,Guiyang550006,China)Abstract:Themajorresultsofstreptomycincontrollingplantbacterialdiseaseswerereviewedfromsuchaspectsasthediscoveryofstreptomycin,theuseofthiscompoundinplantbacterialcontrol,themodeofaction,theantibioticresistancesituation,andalsothemechanismofresistance.Keywords:streptomycin;plantbacterialdiseases;chemicalcontrol;modeofaction;antibioticresistance链霉素(Streptomycin)是从灰链霉菌的培养液中提取的抗生素,属于氨基糖苷碱性化合物。
梨品种‘玉露香’抗火疫病蛋白质组学分析
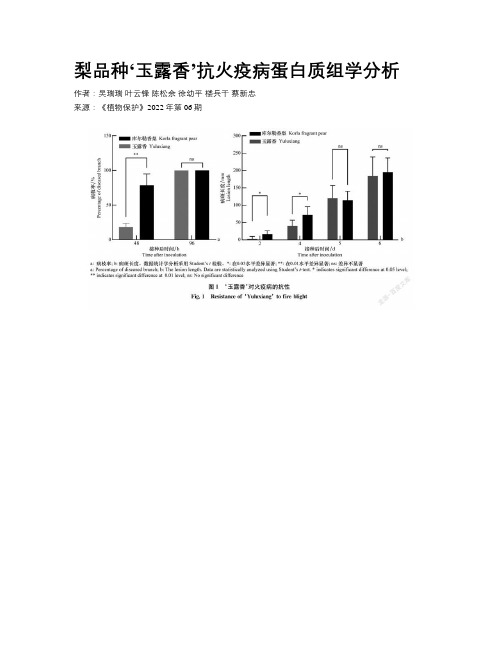
梨品种‘玉露香’抗火疫病蛋白质组学分析作者:吴瑞瑞叶云锋陈松余徐幼平楼兵干蔡新忠来源:《植物保护》2022年第06期关键词梨火疫病;解淀粉欧文氏菌;抗病性;蛋白质组学;铜由革兰氏阴性菌——解淀粉欧文氏菌Eamylovora引起的梨火疫病是世界范围内的毁灭性病害,已被我国列为重要的检疫性有害生物。
梨火疫病菌寄主范围广,主要侵染蔷薇科仁果类果树和其他植物,如梨、苹果和山楂。
其毁灭性还体现在可以侵染寄主植物地上部分所有组织,包括花、嫩枝、树干、主干、果实以及砧木。
该病害起源于北美,但是随着人类活动现已传播扩散至世界60多个国家和地区( https:∥gd. eppo. int/taxon/ERWI-AM/distribution)。
我国果树资源丰富,苹果和梨的栽培面积和总产量均居世界首位。
仅梨年出口创汇就达2亿美元以上,是中国农村和农业经济的支柱产业之一。
如果火疫病在我国大面积发生将造成巨大的经济损失,因此必须进行有效预防和控制。
Eamylovora主要通过自然孔口和伤口侵入植物,进而通过薄壁组织的细胞间隙移至维管束系统,菌体在木质部大量积累阻碍寄主植物的水分传输,从而导致植物枯萎甚至死亡。
Eamylovora致病分子机制已有系统研究,鉴定明确的致病因子包括Ⅲ型分泌系统和胞外多糖。
Ⅲ型分泌系统分泌的效应蛋白包括DspA/E,HrpN,HrpW,Eopl,Eop3和AvrRpt2EA等。
分别与DspA/E和HrpN互作的为苹果感病基因。
植物抗火疫病分子机制近年来取得了重要进展。
已经明确植物对火疫病的抗性是一种数量性状。
目前已经有一些抗火疫病的数量性状位点(quantitative trait loci,QTLs)得到了鉴定,如FB_Mr5,FB_Mfu10以及FB_Mar12。
有些QTL编码典型抗病蛋白(resistance protein,R protein),具有识别Avr等典型基因对基因互作系统中R蛋白的功能。
梨园土壤中梨火疫病菌存活时间研究

um by streak separation method,and the pear fire blight bacterium E.amylovora was confirmed by combination of morphological identification,
in orchards
2019 年重度
2019 年重度
2019 年重度
2019 年重度
2019 年重度
1.2.2 土壤中病原菌分离及纯化。 病原菌分离时间为 2021
28 ℃ 保湿培养,定时观察记录接种叶片和幼果的发病情况。
日、8 月 2 日、9 月 2 日、9 月 25 日、10 月 10 日。
土)、2020040504(砂土)。 2021 年 10 月 10 日(550 d) 时再次
进行病原菌分离,经病原形态鉴定( 图 1)、PCR 检测及致病
性测定确定仍有病原菌存活且具有致病性,但在 2021 年 11
月 9 日(580 d)第 11 次病菌分离时,连续分离了 3 次都未分
离出病原菌,证实该病菌可在土壤中存活 550 d(表 2)。
2 结果与分析
中加入 200 mL 无菌水进行浸泡,静置 12 h 后,各取 2 mL 浸
2021 年 4 月 10 日进行第一次划线分离,检测为阳性的共 3
年 4 月 10 日、4 月 15 日、4 月 22 日、5 月 2 日、6 月 2 日、7 月 2
环腺苷酸受体蛋白基因的过表达对刺糖多孢菌生长和多杀菌素合成的影响

环腺苷酸受体蛋白基因的过表达对刺糖多孢菌生长和多杀菌素合成的影响徐妙;邬洋;杨燕;夏立秋【期刊名称】《中国农业科学》【年(卷),期】2014(000)018【摘要】【目的】利用全局性调控因子环腺苷酸受体蛋白(cyclic AMP receptor protein,Crp)基因在刺糖多孢菌(Saccharopolysporaspinosa)中的过量表达,研究其对刺糖多孢菌生长及形态方面的影响,促进多杀菌素的生物合成。
【方法】PCR 扩增环腺苷酸受体蛋白基因(crp),通过限制性酶切、连接方法构建中间载体pOJ260-cm-PermE-crp,使crp置于红霉素增强型启动子PermE的控制下;PCR 扩增 PermE-crp 基因片段,利用 Red/ET同源重组技术将 PermE-crp 亚克隆至实验室保藏的大肠杆菌-链霉菌穿梭载体 pUC-spn 上,构建成 Crp 表达载体pUC-spn-PermE-crp。
然后,采用接合转移方法将重组载体pUC-spn-PermE-crp导入野生型刺糖多孢菌中,通过单交换同源重组将其整合至刺糖多孢菌染色体上,以染色体上整合了原始质粒pUC-spn的刺糖多孢菌作为对照菌株。
利用PCR 扩增阿伯拉霉素抗性基因及目的基因的方法鉴定阳性接合子;观察工程菌株及对照菌株在不同固体培养基中的生长及菌落形态差异,同时比较工程菌株及对照菌株在液体培养基中的生长情况,测定生长曲线。
借用扫描电子显微镜观察工程菌株及对照菌株的菌丝体形态;采用高效液相色谱检测工程菌株及对照菌株中多杀菌素生物合成情况。
【结果】采用分子生物学方法成功构建了Crp重组表达载体pUC-spn-PermE-crp,并通过接合转移导入到刺糖多孢菌中;PCR检测结果显示,工程菌株作为模板扩增出长约2 kb的cm-PermE-crp目的条带,说明crp已成功整合到刺糖多孢菌染色体上,并且获得的重组工程菌株S. spinosa-Crp遗传性能稳定。
梨火疫病菌两个Sec 依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析

生物技术进展2020年㊀第10卷㊀第5期㊀517~523CurrentBiotechnology㊀ISSN2095 ̄2341研究论文Articles㊀收稿日期:2020 ̄07 ̄24ꎻ接受日期:2020 ̄08 ̄05㊀基金项目:新疆生产建设兵团重点领域科技攻关计划项目(2018AB038)ꎮ㊀联系方式:严海璘E ̄mail:yanhailin@ms.xjb.ac.cnꎻ∗通信作者李为民E ̄mail:liweimin01@caas.cn梨火疫病菌两个Sec依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析严海璘1ꎬ2ꎬ3ꎬ㊀朱宗财1ꎬ2ꎬ3ꎬ㊀张王斌1ꎬ2ꎬ㊀杜培秀3ꎬ㊀张超3ꎬ㊀赵文军4ꎬ㊀李为民3∗1.塔里木大学植物科学学院ꎬ新疆阿拉尔843300ꎻ2.塔里木大学ꎬ南疆农业有害生物综合治理重点实验室ꎬ新疆阿拉尔843300ꎻ3.中国农业科学院生物技术研究所ꎬ北京100081ꎻ4.中国检验检疫科学研究院ꎬ北京100176摘㊀要:利用生物信息学技术结合基于大肠杆菌的碱性磷酸酯酶(PhoA)检测体系ꎬ在梨火疫病菌(Erwiniaamylovora)全基因组序列中筛选鉴定了2个依赖Sec途径分泌的纤维素酶:内切葡聚糖酶EAMY_3602和β ̄葡萄糖苷酶EAMY_1236ꎮ将梨火疫病菌DSM17948接种酥梨幼果ꎬ分别于接种后0㊁8和24h定量检测基因表达ꎬ结果显示EAMY_3602编码基因(Ea3602)在接种后8和24h的mRNA表达量逐渐降低ꎬ但与0h相比无显著差异ꎻEAMY_1236编码基因(Ea1236)在接种后8和24h的mRNA表达量逐步提高ꎬ较0h分别提高了2.43和3.26倍ꎬ差异具有统计学意义(P<0 01)ꎮ由研究结果推测Sec依赖分泌的内切葡聚糖酶EAMY_3602可能与梨火疫病菌侵染无关ꎬ而β ̄葡萄糖苷酶EAMY_1236可能是梨火疫病菌的毒力因子ꎬ在侵染寄主植物过程中发挥作用ꎮ关键词:梨火疫病菌ꎻSec依赖分泌蛋白ꎻ纤维素酶ꎻ致病机理DOI:10.19586/j.2095 ̄2341.2020.0084IdentificationofTwoSec ̄dependentCellulasesinErwiniaamylovoraandGeneExpressionAnalysisDuringInfectionofImmaturePearFruitYANHailin1ꎬ2ꎬ3ꎬZHUZongcai1ꎬ2ꎬ3ꎬZHANGWangbin1ꎬ2ꎬDUPeixiu3ꎬZHANGChao3ꎬZHAOWenjun4ꎬLIWeimin3∗1.CollegeofPlantScienceꎬTarimUniversityꎬXinjiangAlar843300ꎬChinaꎻ2.SouthernXinjiangKeyLaboratoryofIPMꎬTarimUniversityꎬXinjiangAlar843300ꎬChinaꎻ3.BiotechnologyResearchInstituteꎬChineseAcademyofAgriculturalSciencesꎬBeijing100081ꎬChinaꎻ4.ChineseAcademyofInspectionandQuarantineꎬBeijing100176ꎬChinaAbstract:InthisstudyꎬbyusingbioinformaticsanalysisandEscherichiacoli ̄basedalkalinephosphatase(PhoA)genefusionsystemꎬweidentifiedtwoSec ̄dependentsecretedcellulasesꎬanendoglucanaseEAMY_3602andaβ ̄glucosidaseEAMY_1236ꎬfromthewholegenomeofErwiniaamylovora.ThecellsuspensionofE.amylovorastrainDSM17948wasinoculatedintotheyoungpearfruitsꎬquantitativedetectionofgeneexpressionat0ꎬ8and24hpi(hourspost ̄inoculation)ꎬrespectivelyꎬwhichshowedthatthemRNAexpressionlevelsofEa3602ꎬthecodinggeneofEAMY_3602ꎬwerecontinuouslydecreasedat8and24hpiꎬwithnosignificantdifferencewhencomparedwiththatat0hpi.ThetranscriptlevelsofEa1236ꎬthecodinggeneofEAMY_1236ꎬat8and24hpiwere2.43and3.26foldhigherthanthatat0hpi.ResearchresultsshowedthattheSec ̄dependentsecretedendoglucanaseEAMY_3602hadnoimplicationinplantinfectionꎬbutβ ̄glucosidaseEAMY_1236probablyactedasavirulencefactorthatplaysroleintheprocessofE.amylovorainfectingtheimmaturepears.Keywords:ErwiniaamylovoraꎻSec ̄dependentsecretionꎻcellulaseꎻpathogenicmechanism. All Rights Reserved.㊀㊀梨火疫病(fireblight)是一种毁灭性的植物病害ꎬ主要危害仁果类果树和蔷薇科植物ꎮ该病害自19世纪末首次发现于美国ꎬ现已蔓延至欧洲㊁北非㊁大洋洲和亚洲等50多个国家ꎬ严重威胁全球苹果和梨产业的发展[1 ̄2]ꎮ梨火疫病的病原为梨火疫病菌(Erwiniaamylovora)ꎬ是一种革兰氏阴性细菌ꎬ隶属于肠杆菌科欧文氏菌属ꎮ因其危害严重ꎬ梨火疫病菌名列全球十大植物病原细菌之一[3]ꎬ在许多国家和地区被列为重要的检疫性病原[4]ꎮ梨火疫病菌一般通过自然孔口和伤口作为感染途径ꎬ首先侵入树木皮层薄壁组织的细胞间隙ꎬ在后期进入木质部导管ꎬ并能在植物体内快速迁移ꎻ感病植株最初症状表现为水浸状ꎬ随后出现萎蔫和坏死ꎬ受感染的部位变成棕色或黑色ꎬ类似火烧一样ꎬ最终导致整棵树的死亡[5 ̄6]ꎮ病原菌分泌的致病相关蛋白也称为效应因子ꎬ在与宿主植物互作过程中发挥重要作用[7 ̄8]ꎮ在革兰氏阴性菌中ꎬIII型分泌系统㊁IV型分泌系统㊁Sec依赖分泌途径和双精氨酸移位酶途径(Tat途径)是效应因子输出的主要通道[9]ꎮ其中ꎬSec分泌途径包括由3种跨膜蛋白SecY㊁SecE和SecG组装的SecYEG通道ꎬ在外周膜ATP酶SecA的协助下转运包括毒力因子在内的各种蛋白ꎻ此外ꎬSec途径还包含SecD㊁SecF和YajCꎬ由这3种膜蛋白参与调控蛋白质的高效输出[10 ̄11]ꎮSec分泌蛋白在细胞质中以未折叠前蛋白的形式存在ꎬ氨基末端为信号肽(signalpeptideꎬSP)ꎬ指导前体蛋白经由Sec系统转运ꎮ在革兰氏阴性菌中ꎬ前体蛋白跨越细胞内膜ꎬ转移至周质ꎬ由胞外信号肽酶I(LepB)或脂蛋白信号肽酶II(LspA)切割SPꎬ并折叠为成熟蛋白ꎻ成熟蛋白或者停留在周质中ꎬ或者转移到次级分泌途径ꎬ如V型分泌系统(typeVsecretionsystemꎬT5SS)㊁脂蛋白(Lol)途径或Ⅱ型分泌系统(typeⅡsecretionsystemꎬT2SS)ꎬ并进一步转运为外膜或胞外蛋白[7 ̄8ꎬ11]ꎮ植物病原菌分泌的细胞壁降解酶是一类重要的致病因子[12 ̄14]ꎮ细胞壁降解酶包括纤维素酶㊁果胶酶㊁半纤维素酶㊁木聚糖酶等ꎬ在病原菌消解植物抗侵染机械屏障㊁入侵寄主等方面发挥着重要作用ꎮ纤维素酶是与致病相关的主要酶类之一[15 ̄18]ꎬ已知许多黄单胞菌属(Xanthomonas)㊁欧文氏菌属㊁青枯菌属(Ralstonia)的植物病原细菌ꎬ以及丝核菌属(Rhizoctonia)㊁稻瘟菌属(Magnaporthe)的植物病原真菌均分泌纤维素酶[19 ̄22]ꎬ以此协助病原自身的入侵与扩散ꎮ迄今为止ꎬ关于梨火疫病菌是否编码具有分泌特征的纤维素酶类尚不清楚ꎮ本研究利用生物信息学技术结合大肠杆菌(Escherichiacoli)碱性磷酸酯酶(phosphataseꎬPhoA)检测体系ꎬ从梨火疫病菌全基因组序列中筛选出2种依赖Sec途径分泌的纤维素酶EAMY_3602(GenBank:FN434113 1)和EAMY_1236(GenBank:FN434113.1)ꎬ并对其进行反转录实时荧光定量PCR(reversetranscriptionquantitativereal ̄timePCRꎬRT ̄qPCR)分析ꎬ以期发现梨火疫病菌的毒力因子ꎬ为梨火疫病的防治提供参考ꎮ1㊀材料与方法1.1㊀材料与试剂梨火疫菌株DSM17948由中国检验检疫科学研究院保存ꎬpET ̄30a质粒㊁大肠杆菌DH5α和BL21(DE3)均由本实验室保存ꎮ试验所用试剂盒均购自TaRaKa㊁TIANGEN㊁全式金公司ꎻ其他试剂均为国产分析纯ꎮ1.2㊀方法1.2.1㊀生物信息学分析㊀分泌蛋白信号肽的生物信息学分析基于SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)[23]㊁Lipopversion1.0(http://www.cbs.dtu.dk/services/LipoP/)[24]和Phobius(http://phobius.sbc.su.se/)[25]完成ꎻSec依赖分泌蛋白中纤维素酶的筛选参照梨火疫病菌CFBP1430编码蛋白的功能注释[26]ꎮ1.2.2㊀基因克隆㊀根据梨火疫病菌CFBP1430(GenBank:FN434113.1)㊁EAMY_3602和EAMY_1236编码基因(分别命名为Ea3602和Ea1236)的核酸序列ꎬ设计两对引物3602 ̄F/3602 ̄R和1236 ̄F/1236 ̄R(引物序列见表1)ꎮ利用CTAB法提取梨火疫菌株DSM17948的基因组DNAꎬ以此为模板ꎬ利用上述引物分别扩增Ea3602和Ea1236ꎬPCR扩增㊁产物回收㊁连接㊁转化均参照严海璘等[27]的方法进行ꎮ1.2.3㊀PhoA检测㊀基于大肠杆菌的PhoA检测体系由本研究室构建[28]ꎮ设计两对引物3602SP ̄F/3602SP ̄R和1236SP ̄F/1236SP ̄R(引物序列见表1)ꎬ分别以pMD ̄Ea3602和pMD ̄Ea1236质粒815生物技术进展CurrentBiotechnology. All Rights Reserved.表1㊀本研究所用引物序列Table1㊀Primersusedinthisstudy基因名称引物名称引物序列(5ᶄң3ᶄ)EAMY_36023602 ̄F5ᶄ ̄ATGCTGTTGGCGCTCATAATG ̄3ᶄ3602 ̄R5ᶄ ̄TTAGCGGTCTTTAAACGCCAG ̄3ᶄ3602SP ̄F5ᶄ ̄AAGGAGATATACATATGCTGTTGGCGCTCATAATG ̄3ᶄ3602SP ̄R5ᶄ ̄AGGCATTTCTGGAAGCTTCCAGCCATCTGCGGCCAT ̄3ᶄ3602 ̄qF5ᶄ ̄GGTTTATTTCAACGACCGCA ̄3ᶄ3602 ̄qR5ᶄ ̄TTTCTCCCGCCTTCAGTAGC ̄3ᶄEAMY_12361236 ̄F5ᶄ ̄ATGAATTGGATTTGTTCTGTCA ̄3ᶄ1236 ̄R5ᶄ ̄CTACAGATAATCAAATTCGGC ̄3ᶄ1236SP ̄F5ᶄ ̄AAGGAGATATACATATGAATTGGATTTGTTCTGTCA ̄3ᶄ1236SP ̄R5ᶄ ̄AGGCATTTCTGGAAGCTTGATGATATCGGCGGCGAAA ̄3ᶄ1236 ̄qF5ᶄ ̄TTTGGCGAAGATACCTGGTT ̄3ᶄ1236 ̄qR5ᶄ ̄ATACGGCGGCAGATAGTCCT ̄3ᶄrecArecA525F5ᶄ ̄GCGTATGATGAGCCAGGCA ̄3ᶄrecA716R5ᶄ ̄ACCACTTCGTCGCCCTCTT ̄3ᶄ㊀注:CATATG表示NdeⅠ酶切位点ꎬAAGCTT表示HindⅢ酶切位点ꎮ为模板ꎬPCR扩增Ea3602和Ea1236基因的信号肽编码序列ꎬ长度分别为42和60bpꎬmphoA载体构建和检测均参照严海璘等[27]的方法进行ꎮ1.2.4㊀接种梨幼果㊀梨幼果接种参照严海璘等[27]的方法进行ꎮ接种后分别在0㊁8㊁24h取样ꎬ用无菌刀片刮取大约0.8g的剖面组织ꎬ于-80ħ冰箱中保存备用ꎮ1.2.5㊀总RNA提取与RT ̄qPCR分析㊀提取样品总RNAꎬ反转录获得cDNAꎮ以梨火疫菌recA(GenBank:FN434113.1)为内参基因[29]ꎬ利用引物3602 ̄qF/3602 ̄qR㊁1236 ̄qF/1236 ̄qR(引物序列见表1)分别进行Ea3602和Ea1236基因表达的qPCR分析ꎮ具体参照TBGreen PremixExTaqTMII(TliRNaseHPlus)说明书进行ꎮ数据分析采用2-ΔΔCT(Livak)方法计算相对表达量ꎬΔΔCT=(CT目的基因-CT内参基因)试验组-(CT目的基因-CT内参基因)对照组ꎬ显著性差异分析采用Student st ̄test[30]ꎬP<0.05和P<0.01表示差异具有统计学意义ꎮ2㊀结果与分析2.1㊀筛选梨火疫病菌Sec依赖分泌纤维素酶前期研究表明ꎬ梨火疫病菌CFBP1430编码3393个蛋白ꎬ其中168个蛋白N末端为SPsꎬ推测其可能通过Sec系统分泌[31]ꎮ参照CFBP1430编码蛋白的功能注释[26]ꎬ我们从168种Sec依赖分泌蛋白中进一步筛选出2种具有纤维素酶功能的蛋白:EAMY_3602和EAMY_1236ꎬ其中前者为内切葡聚糖酶ꎬ后者为β ̄葡萄糖苷酶ꎮ目前ꎬ已有CFBP1430[26]㊁ATCC49946[32]和E ̄2[33]共3个梨火疫病菌菌株完成了全基因组序列测定ꎮBlast分析表明EAMY_3602和EAMY_1236在所有3个菌株中的同源性均为100%ꎮ利用SignalP4.1㊁Lipopversion1.0和Phobius分析进一步证实EAMY_3602和EAMY_1236N末端为信号肽序列ꎬ且均由信号肽酶LepB切割ꎬ切割位点分别位于第14个和20个氨基酸(表2)ꎮ2.2㊀克隆Ea3602和Ea1236基因以梨火疫菌株DSM17948的基因组DNA为模板ꎬ利用3602 ̄F/3602 ̄R和1236 ̄F/1236 ̄R扩增EAMY_3602和EAMY_1236的编码基因ꎬ分别获得975和2298bp的PCR产物(图1)ꎬ与预期结果大小一致ꎮ将目的片段回收㊁连接ꎬ测序结果确证所扩增片段为Ea3602和Ea1236ꎮ2.3㊀验证EAMY_3602和EAMY_1236的分泌特性为了验证EAMY_3602和EAMY_1236的分泌特性ꎬ我们分别将EAMY_3602和EAMY_1236信号肽(3602SP和1236SP)的编码序列插入pET ̄mphoAꎬ与mphoA融合ꎬ构建pET ̄3602SP ̄mphoA915严海璘ꎬ等:梨火疫病菌两个Sec依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析. All Rights Reserved.表2㊀梨火疫病菌Sec依赖分泌纤维素酶Table2㊀TheputativeSec ̄dependentsecretedcellulaseidentifiedfromE.amylovora蛋白名称GenBank登录号信号肽酶切割位点功能注释EAMY_3602WP_004161055.1LepB14~15内切葡聚糖酶EAMY_1236WP_004156773.1LepB20~21β ̄葡萄糖苷酶㊀注:参照梨火疫菌株CFBP1430(GenBank:NC_013961.1)ꎮM DL5000Markerꎻ1 Ea3602的PCR产物ꎻ2 Ea1236的PCR产物图1㊀Ea3602和Ea1236基因的PCR扩增Fig.1㊀PCRproductofEa3602andEa1236gene和pET ̄1236SP ̄mphoA(图2A)ꎬ转化至大肠杆菌BL21细胞ꎮPhoA检测结果显示:含有pET ̄3602SP ̄mphoA或pET ̄1236SP ̄mphoA转化子在指示培养基培养6h后ꎬ呈现蓝色ꎬ与阳性对照(含有pET ̄phoA的BL21细胞)结果相似ꎬ而阴性对照(带有pET ̄mphoA的BL21细胞)仍保持白色(图2B)ꎮ这一结果表明3602SP和1236SP能够引导mphoA从细胞质转移到周质ꎬ由此证实EAMY_3602和EAMY_1236可通过梨火疫病菌的Sec途径进行分泌ꎮ2.4㊀Ea3602和Ea1236在梨火疫病菌侵染幼梨过程中的表达分析为了揭示Ea3602和Ea1236在病原菌侵染过程中的潜在功能ꎬ我们用梨火疫病菌接种酥梨幼果ꎬ分别在接种后0㊁8㊁24h提取总RNAꎬ利用RT ̄qPCR分析这两种纤维素酶编码基因的表达水平ꎮ结果显示:Ea3602在8和24h的表达量逐渐降低ꎬ但与0h相比无显著差异(图3)ꎻ相反ꎬEa1236在8和24h的表达量逐步提高ꎬ分别较0h提高2.43和3.26倍ꎬ差异具有统计学意义(P<0.01)ꎮ由此推测β ̄葡萄糖苷酶EAMY_1236可能是梨火疫病菌的毒力因子ꎬ在侵染寄主植物过程中发挥重要作用ꎬ而内切葡聚糖酶EAMY_3602则与梨火疫病菌侵染无明显关联ꎮA:PhoA检测体系的载体示意图ꎻB:信号肽3602SP和1236SP引导mphoA分泌图2㊀利用phoA体系检测EAMY_3602和EAMY_1236的信号肽Fig.2㊀ThesignalpeptidesofEAMY_3602andEAMY_1236weredetectedbyphoAsystem025生物技术进展CurrentBiotechnology. All Rights Reserved.注:∗∗ 与同组0h相比ꎬ差异有统计学意义(P<0.01)ꎮ图3㊀Ea3602和Ea1236基因在梨火疫病菌侵染过程中的表达分析Fig.3㊀AnalysisontheexpressionofEa3602geneandEa1236geneintheprocessofinfectionofE.amylovora3㊀讨论在细菌中ꎬ大约20%~30%的编码蛋白定位于胞质外ꎬ蛋白的胞质外转运对于维持细菌生存至关重要[9ꎬ34]ꎮSec分泌途径在细菌中广泛存在ꎬ是蛋白胞质外转运主要的跨膜运输系统[35]ꎬ包括毒力因子在内的很多蛋白均通过Sec途径分泌到胞质外环境中[10 ̄11]ꎮ关于梨火疫病的研究已有百余年历史ꎬ目前已明确胞外多糖(exopolysaccharideꎬEPS)能够保护梨火疫病菌有效躲避植物防御系统ꎬ以及在干旱条件下免受水分和营养损失[36 ̄38]ꎻ此外发现DspA/E㊁HrpN㊁Eop1和AvrRpt2EA等III型分泌系统(TypeIIIsecretionsystemꎬT3SS)分泌蛋白对于病原菌入侵及其在寄主植物中定殖同样具有重要作用[39 ̄41]ꎮ最新研究显示ꎬ梨火疫病菌可能利用Sec途径分泌蛋白酶介导病原侵染[31]ꎮ本研究利用生物信息学分析结合PhoA检测体系ꎬ从梨火疫病菌Sec分泌蛋白中筛选鉴定出2个纤维素酶:EAMY_3602和EAMY_1236ꎮ已有研究表明许多植物病原细菌和病原真菌分泌的纤维素酶是其完全毒力发挥所必需的[19 ̄21]ꎻ同时ꎬ纤维素酶也是激发植物防御反应的有效诱导物ꎬ例如ꎬ用纤维素酶处理甜椒叶片ꎬ能够诱导水杨酸水平显著增加[42]ꎮ纤维素酶根据结构㊁功能和作用方式ꎬ可分为外切葡聚糖酶㊁内切葡聚糖酶和β ̄葡萄糖苷酶[43 ̄44]ꎮ参照梨火疫病菌编码蛋白的功能注释[26]ꎬ本研究筛选的两个纤维素酶EAMY_3602和EAMY_1236分别为内切葡聚糖酶和β ̄葡萄糖苷酶ꎮ关于病原菌内切葡聚糖酶已有大量研究ꎬ如:水稻黄单胞菌(X.oryzaepv.oryzae)㊁菊欧文氏菌(E.chrysanthemi)㊁柑橘黄单胞菌亚种(X.citrisubsp.citri)的内切葡聚糖酶均为其重要毒力因子ꎬ参与病原菌的致病过程[21ꎬ45 ̄46]ꎮ但是ꎬ本研究发现梨火疫病菌在侵染幼梨时ꎬEa3602的mRNA表达量没有显著变化ꎬ因而推测Sec分泌内切葡聚糖酶EAMY_3602可能与梨火疫病菌侵染无密切关联ꎮ与之相反ꎬEa1236基因在病原侵染过程中mRNA表达量显著提高ꎬ暗示β ̄葡萄糖苷酶EAMY_1236可能是梨火疫病菌潜在的毒力因子ꎮ但根据早期报道ꎬ梨火疫病菌在致病过程中没有造成寄主植物细胞壁多糖的降解[47]ꎬ暗示纤维素酶没有参与该病原对寄主的侵染ꎻ此外ꎬ另有文献指出在梨火疫病菌中表达大肠杆菌β ̄葡萄糖苷酶ꎬ对其毒力没有显著影响[48]ꎮ因此ꎬ我们在后续研究中将制备Ea1236基因的敲除突变体ꎬ确证EAMY_1236是否参与病原侵染以及在不同时空维度的作用ꎬ以期揭示梨火疫病菌致病的新机制ꎮ参㊀考㊀文㊀献[1]㊀CHATTERJEEA.Fireblight:thediseaseanditscausativea ̄gentꎬErwiniaamylovora[J].Eur.J.PlantPathol.ꎬ2001ꎬ107(5):569-569.[2]㊀ZHAOYꎬTIANYꎬWANGLꎬetal..Fireblightdiseaseꎬafast ̄approachingthreattoappleandpearproductioninChina[J].J.Integr.Agric.ꎬ2019ꎬ18(4):815-820. [3]㊀MANSFIELDJWꎬGENINSꎬMAGORISꎬetal..Top10plantpathogenicbacteriainmolecularplantpathology[J].Mol.PlantPathol.ꎬ2012ꎬ13(6):614-629.[4]㊀胡白石.梨火疫病菌的风险分析及检测技术研究[D].南京:南京农业大学ꎬ博士学位论文ꎬ2000.[5]㊀VANNESTEJLꎬEDENGREENSꎬVANNESTEJL.MigrationofErwiniaamylovorainhostplanttissues[M].Blight:TheDiseaseanditsCausativeAgentꎬ2000.[6]㊀MALNOYMꎬMARTENSSꎬNORELLIJLꎬetal..Fireblight:appliedgenomicinsightsofthepathogenandhost[J].Ann.Rev.Phytopathol.ꎬ2012ꎬ50(1):475-494. [7]㊀TSIRIGOTAKIAꎬDEGEYTERJꎬŠOšTARICNꎬetal..Pro ̄125严海璘ꎬ等:梨火疫病菌两个Sec依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析. All Rights Reserved.teinexportthroughthebacterialSecpathway[J].Nat.Rev.Microbiol.ꎬ2017ꎬ15(1):21-36.[8]㊀SMETSDꎬLOOSMSꎬKARAMANOUSꎬetal..ProteintransportacrossthebacterialplasmamembranebytheSecpath ̄way[J].ProteinJ.ꎬ2019ꎬ38(3):262-273. [9]㊀HOLLANDIB.Theextraordinarydiversityofbacterialproteinsecretionmechanisms[J].MethodsMol.Biol.ꎬ2010ꎬ619(619):1-20.[10]㊀訾祯祯ꎬ杨志伟.细菌蛋白分泌途径的研究进展[J].生物技术通报ꎬ2011(8):44-50ꎬ54.[11]㊀CRANFORDSMITHTꎬHUBERD.Thewayisthegoal:howSecAtransportsproteinsacrossthecytoplasmicmembraneinbacteria[J/OL].FemsMicrobiol.Lett.ꎬ2018ꎬ365(11):fny093.https://doi.org/10.1093/femsle/fny093.[12]㊀HAVEATꎬBREUILWOꎬWUBBENJPꎬetal..Botrytisci ̄nereaendopolygalacturonasegenesaredifferentiallyexpressedinvariousplanttissues[J].FungalGenet.Biol.ꎬ2001ꎬ33(2):97-105.[13]㊀MANTEAUSꎬABOUNASꎬLambertBꎬetal..DifferentialregulationbyambientpHofputativevirulencefactorsecretionbythephytopathogenicfungusBotrytiscinerea[J].FEMSMi ̄crobiol.Ecol.ꎬ2003ꎬ43(3):359-366.[14]㊀REIGNAULTPꎬVALETTECOLLETOꎬBOCCARAMꎬetal..Theimportanceoffungalpectinolyticenzymesinplantinvasionꎬhostadaptabilityandsymptomtype[J].Eur.J.PlantPathol.ꎬ2007ꎬ120(1):1-11.[15]㊀OKADAG.PurificationandpropertiesofacellulasefromAs ̄pergillusniger[J].Agric.Biol.Chem.ꎬ1985ꎬ49(5):1257-1265.[16]㊀WALKERDꎬREEVESPJꎬSALMONDGPꎬetal..ThemajorsecretedcellulaseꎬCelVꎬofErwiniacarotovorasubsp.carotovoraisanimportantsoftrotvirulencefactor[J].Mol.PlantMicrob.Interact.ꎬ1994ꎬ7(3):425-431.[17]㊀DEESEDCꎬSTAHMANNMA.FormationofpecticenzymesbyVerticilliumAlbo ̄atrumonsusceptibleandresistanttomatostemtissuesandonwheatbran[J].J.Phytopathol.ꎬ1963ꎬ46(1):53-70.[18]㊀李永胜.柑桔溃疡病菌(Xanthomonasaxonopodispv.citri)纤维素酶的分析[D].重庆:西南大学ꎬ硕士学位论文ꎬ2011. [19]㊀GOUGHCLꎬDOWJMꎬBARBERCEꎬetal..CloningoftwoendoglucanasegenesofXanthomonascampestrespv.Campestris:analysisoftheroleofthemajorendoglucanaseinpathogenesis[J].Mol.PlantMicrob.Interact.ꎬ1988ꎬ1(7):275-281.[20]㊀ROBERTSDPꎬDENNYTPꎬSCHELLMA.CloningoftheeglgeneofPseudomonassolanacearumandanalysisofitsroleinphytopathogenicity.[J].J.Bacteriol.ꎬ1988ꎬ170(4):1445-1451.[21]㊀TIANXꎬYANJIAOLꎬDONGLINGSꎬetal..Correction:i ̄dentificationofanextracellularendoglucanasethatisrequiredforfullvirulenceinXanthomonascitrisubsp.Citri[J/OL].PLoSONEꎬ2016ꎬ11(5):e0156458.https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0156458[22]㊀李宝聚ꎬ周长力ꎬ赵奎华ꎬ等.黄瓜黑星病菌致病机理的研究Ⅱ细胞壁降解酶及其在致病中的作用[J].植物病理学报ꎬ2000(1):13-18.[23]㊀PETERSENTNꎬSØRENBRUNAKꎬHEIJNEGVꎬetal..SIGNALP4.0:discriminatingsignalpeptidesfromtransmem ̄braneregions[J].Nat.Methodsꎬ2011ꎬ8(10):785-786. [24]㊀JUNCKERASꎬWILLENBROCKHꎬHEIJNEGVꎬetal..PredictionoflipoproteinsignalpeptidesinGram ̄negativebac ̄teria[J].ProteinSci.ꎬ2003ꎬ12(8):1652-1662. [25]㊀KäLLLꎬKROGHAꎬSONNHAMMERELL.Advantagesofcombinedtransmembranetopologyandsignalpeptideprediction ̄thePhobiuswebserver[J].Nucl.AcidsRes.ꎬ2007ꎬ35:429-432.[26]㊀SMITSTHꎬREZZONICOFꎬKAMBERTꎬetal..CompletegenomesequenceofthefireblightpathogenErwiniaamylovoraCFBP1430andcomparisontootherErwiniaspp.[J].Mol.PlantMicrob.Interact.ꎬ2010ꎬ23(4):384-393.[27]㊀严海璘ꎬ朱宗财ꎬ张王斌ꎬ等.梨火疫病菌Sec依赖分泌蛋白EAMY_3046的鉴定及其原核表达[J].植物检疫ꎬ2020ꎬ34(4):1-6.[28]㊀LIUXꎬFANYꎬZHANGCꎬetal..Nuclearimportofasecreted Candidatusliberibacterasiaticus proteinistempera ̄turedependentandcontributestopathogenicityinNicotianabenthamiana[J/OL].Front.Microbiol.ꎬ2019ꎬ10:1684.ht ̄tp://doi.org/10.3389/fmicb.2019.01684.[29]㊀KALUZNAMꎬKURASAꎬPULAWSKAJꎬetal..ValidationofreferencegenesforthenormalizationoftheRT ̄qPCRgeneexpressionofvirulencegenesofErwiniaamylovorainappleshoots[J].Scient.Rep.ꎬ2017ꎬ7(1):2034-2034. [30]㊀施雨ꎬ李耀武.概率论与数理统计应用[M].西安:西安交通大学出版社ꎬ2005.[31]㊀ZHANGWꎬYANHꎬZHUZꎬetal..Genome ̄wideidentifica ̄tionoftheSec ̄dependentsecretoryproteasegenesinErwiniaamylovoraandanalysisoftheirexpressionduringinfectionofimmaturepearfruit[J].J.ZhejiangUniv.Sci.Bꎬ2020ꎬ21(9)ꎬdoi:10.1631/jzus.B2000281(inpress).[32]㊀SEBAIHIAMꎬBOCSANCZYAMꎬBIEHLBSꎬetal..Com ̄pletegenomesequenceoftheplantpathogenErwiniaamylovorastrainATCC49946[J].J.Bacteriol.ꎬ2010ꎬ192(7):2020-2021.[33]㊀LAGONENKOALꎬKOMARDINAVSꎬNIKOLAICHIKYꎬetal..FirstreportofErwiniaamylovorafireblightinbelarus[J].J.Phytopathol.ꎬ2008ꎬ156(10):638-640.[34]㊀PUGSLEYAP.Thecompletegeneralsecretorypathwayingram ̄negativebacteria[J].Microbiol.Rev.ꎬ1993ꎬ57(1):50-108.[35]㊀WILSONJW.BacterialProteinSecretionMechanisms[M]//NickersonCAꎬSchurrMJ.MolecularParadigmsofInfectiousDisease:ABacterialPerspective.Boston.MA:SpringerUSꎬ2006ꎬ274-320.[36]㊀BELLEMANNPꎬBERESWILLSꎬBERGERSꎬetal..Visual ̄izationofcapsuleformationbyErwiniaamylovoraandassaystodetermineamylovoransynthesis[J].Int.J.Biol.Macromol.ꎬ1994ꎬ16(6):290-296.[37]㊀MAESMꎬORYEKꎬBOBEVSꎬetal..InfluenceofamylovoranproductiononvirulenceofErwiniaamylovoraandadifferentamylovoranstructureinE.amylovoraisolatesfromRu ̄225生物技术进展CurrentBiotechnology. All Rights Reserved.bus[J].Eur.J.PlantPathol.ꎬ2001ꎬ107(8):839-844. [38]㊀KOCZANJMꎬMCGRATHMJꎬZHAOYꎬetal..ContributionofErwiniaamylovoraexopolysaccharidesamylovoranandlevantobiofilmformation[J].Phytopathologyꎬ2009ꎬ99(11):1237-1244.[39]㊀NISSINENRMꎬYTTERBERGAJꎬBOGDANOVEAJꎬetal..AnalysesofthesecretomesofErwiniaamylovoraandse ̄lectedhrpmutantsrevealnoveltypeIIIsecretedproteinsandaneffectofHrpJonextracellularharpinlevels[J].Mol.PlantPathol.ꎬ2007ꎬ8(1):55-67.[40]㊀CASTIBLANCOLFꎬTRIPLETTLRꎬSUNDINGWꎬetal..RegulationofeffectordeliverybytypeIIIsecretionchaperoneproteinsinErwiniaamylovora[J].Front.Microbiol.ꎬ2018ꎬ9:146-146.[41]㊀SCHROPFERSꎬBOTTCHERCꎬWOHNERTꎬetal..AsingleeffectorproteinꎬAvrRpt2EAꎬfromErwiniaamylovoracancausefireblightdiseasesymptomsandinducesasalicylicacid ̄dependentdefenseresponse[J].Mol.PlantMicrob.Inter ̄act.ꎬ2018ꎬ31(11):1179-1191.[42]㊀SATOCꎬOKANꎬNABETAKꎬetal..Cellulaseappliedtotheleavesofsweetpepper(CapsicumannuumL.var.grossum)upregulatestheproductionofsalicylicandazelaicacids[J].Biosci.Biotechnol.Biochem.ꎬ2011ꎬ75(4):761-763.[43]㊀SINGHANIARRꎬPATELAKꎬSUKUMARANRKꎬetal..Roleandsignificanceofbeta ̄glucosidasesinthehydrolysisofcelluloseforbioethanolproduction[J].Bioresour.Technol.ꎬ2013ꎬ127:500-507.[44]㊀TAYILꎬKUMARSꎬNATHAWATRꎬetal..AmutationinanexoglucanaseofXanthomonasoryzaepv.oryzaeꎬwhichconfersanendomodeofactivityꎬaffectsbacterialvirulenceꎬbutnottheinductionofimmuneresponsesꎬinrice[J].Mol.PlantPathol.ꎬ2018ꎬ19(6):1364-1376.[45]㊀HUJꎬQIANWꎬHEC.TheXanthomonasoryzaepv.oryzaeeg ̄lXoBendoglucanasegeneisrequiredforvirulencetorice[J].FEMSMicrobiol.Lett.ꎬ2007ꎬ269(2):273-279.[46]㊀BOCCARAMꎬAYMERICJLꎬCamusC.Roleofendoglu ̄canasesinErwiniachrysanthemi3937virulenceonSaintpauliaionantha[J].J.Bacteriol.ꎬ1994ꎬ176(5):1524-1526. [47]㊀SEEMüLLEREA.Absenceofcellwallpolysaccharidedegra ̄dationbyErwiniaamylovora[J].Phytopathologyꎬ1976ꎬ66(4):433-436.[48]㊀KERPPOLATKꎬSERWOLD ̄DAVISTꎬGROSSDCꎬetal..EffectofIncreasedbeta ̄GlucosidaseActivityonVirulenceofErwiniaamylovora[J].Appl.Environ.Microbiol.ꎬ1987ꎬ53(4):677-682.325严海璘ꎬ等:梨火疫病菌两个Sec依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析. All Rights Reserved.。
梨火疫病抑菌制剂的室内筛选

梨火疫病抑菌制剂的室内筛选梁慧敏;刘君;王希东;杨治月;李敬桦【摘要】[目的]筛选抑制梨火疫菌(Erwinia amylovora)的药剂,为防控梨火疫病提供参考与依据.[方法]采用最低抑制浓度和抑菌圈比较的方法,对30种抑菌制剂进行筛选.[结果]内吸性抑菌制剂1.8%辛菌胺醋酸盐,有机类抑菌制剂3%噻霉酮(ME)、3%噻霉酮(WP)和30%壬菌铜,植物源类型制剂80%乙蒜素以及微生物制剂荧光假单胞杆菌3 000,是室内试验对梨火疫菌抑菌效果较好的制剂.[结论]不同药剂之间、同一种药剂不同剂型之间抑菌效果差异较大,研究筛选得到的6种抑菌制剂可为梨火疫的防治提供参考.【期刊名称】《新疆农业科学》【年(卷),期】2019(056)002【总页数】12页(P333-344)【关键词】梨火疫病;病害防控;抑菌制剂;抑菌圈【作者】梁慧敏;刘君;王希东;杨治月;李敬桦【作者单位】新疆农业大学林学与园艺学院,乌鲁木齐830052;新疆农业大学林学与园艺学院,乌鲁木齐830052;新疆农业大学农学院,乌鲁木齐830052;新疆农业大学农学院,乌鲁木齐830052;新疆农业大学农学院,乌鲁木齐830052【正文语种】中文【中图分类】S480 引言【研究意义】梨火疫(Fire blight)是引起梨、苹果等蔷薇科植物发病的最具毁灭性的细菌性病害之一,梨火疫病[1]是我国对外重大检疫性病害之一。
在1951~1960 年间,该病害造成美国平均每年因该病经济损失400万美元[2]。
该病侵染情况复杂[3]且市场需求种植的品种多为感病品种,目前该病害还没有被有效地控制,防治上较为困难,仍然是大多数梨和苹果种植国主要关注的问题[4]。
虽然目前我国还没有梨火疫病发生的报道,但梨火疫病的人为和自然传播已对我国构成了潜在戚胁[5]。
仍需严格地做好疫情调查和植物检疫,筛选高效防病药剂,科学防控该病的发生与为害,对我国植物检疫及梨和苹果产业的健康发展具有重要意义。
亚洲梨火疫病菌入侵中国的风险分析

亚洲梨火疫病菌入侵中国的风险分析
商明清;赵文军;李志红;杨伟东;朱水芳
【期刊名称】《植物检疫》
【年(卷),期】2009(23)6
【摘要】亚洲梨火疫病(Erwinia pyrifoliae)是一种严重为害梨树的毁灭性病害,本文从其分布状况、寄主、危害及经济重要性、寄主植物经济重要性、传入与定殖可能性和风险管理难度等方面进行了综合分析评估,其综合风险值R为2.52,属于特别危险的有害生物,并据此提出了防止其入侵为害的风险管理措施。
【总页数】3页(P37-39)
【关键词】亚洲梨火疫病;Erwinia;pyrifoliae;风险分析
【作者】商明清;赵文军;李志红;杨伟东;朱水芳
【作者单位】中国检验检疫科学研究院;山东省植物保护总站;中国农业大学;深圳出入境检验检疫局
【正文语种】中文
【中图分类】N945.24;S41
【相关文献】
1.梨火疫病菌在中国的潜在分布及入侵风险分析 [J], 陈晨;陈娟;胡白石;白艺珍;姜英华;刘凤权
2.梨火疫病菌两个Sec依赖的外泌纤维素酶鉴定及在侵染梨幼果过程中的基因表达分析 [J], 严海璘;朱宗财;张王斌;杜培秀;张超;赵文军;李为民
3.我国20个梨品种(种质)对国外梨火疫病菌的抗病性评价 [J], 李洪涛;张静文;盛强;唐章虎;张祥林;张春竹;罗明
4.用离体巴梨枝条测定梨火疫病菌的致病性 [J], 张乐;石秀丽
因版权原因,仅展示原文概要,查看原文内容请购买。
免疫吸附PCR技术提高梨火疫病菌检测灵敏度

免疫吸附PCR技术提高梨火疫病菌检测灵敏度
苏梅华;吴建波;李秋英;卢玲;胡白石
【期刊名称】《植物检疫》
【年(卷),期】2010(24)3
【摘要】本研究采用已经发表的引物和制备的梨火疫病菌抗血清,研制了免疫吸附-PCR技术,使其检测梨火疫病菌纯菌的灵敏度比标准PCR技术提高10倍;检测模拟样品中的梨火疫病菌灵敏度提高了1000倍;从相关混合菌液中能够更加灵敏和准确地检测出梨火疫病菌。
该方法简单易行,准确灵敏,具有广阔的应用前景。
【总页数】3页(P8-10)
【关键词】梨火疫病菌;免疫吸附技术;抗血清;PCR
【作者】苏梅华;吴建波;李秋英;卢玲;胡白石
【作者单位】太仓出入境检验检疫局;新疆林业学校;南京农业大学农业部病虫监测与治理重点开放实验室
【正文语种】中文
【中图分类】S432
【相关文献】
1.梨火疫病菌的免疫检测技术(简报) [J], 谢洪芳;沈秀萍
2.免疫捕获PCR检测进境苹果果实中梨火疫病菌 [J], 何丹丹;周国梁;陈仲兵;印丽萍;徐殿胜;易建平
3.梨火疫细菌实时荧光PCR和诱捕PCR-ELISA检测方法的建立 [J], 朱建裕;廖晓
兰;高必达;朱水芳;陈红运;戴富泉
4.利用间接免疫荧光染色和协同凝集技术检测梨火疫病菌 [J], 胡白石;卢玲;刘凤权;许志刚;陈建东;焦国尧
5.梨火疫病菌的实时荧光PCR检测 [J], 钱国良;胡白石;卢玲;刘凤权;许志刚
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
R c tr rt nGe e c )rm Ew naa yooa epo oe n (r f r ii m l r P i p o v
.
h mo o f r e e d s n t d Ea r wa d n i e n l n d i r i i my o o a A r dir p i n mutn o l g o p g n , e i ae cp, si e t d a d c o e E w n a a lv r c g i f n cp s to u at
农业 生物 技 术 学 报 ,00年 , 1 , 5 , 8 7 8 5页 21 第 8卷 第 期 第 2 ~ 3
J u a f r u t r l o e h o o y 2 1 , 11 , . , 2 - 3 o r l Ag i l a t c n l g , 0 0 Vo . 8 No 5 8 7 8 5 n o c u Bi
rcpo rti, R ) 果胶 酶基 因 的表达 调控 和菌株 致病 性起 着重 要 的作用 。 eetr oe C P 对 p n 本研 究 首次鉴 定 并克 隆出梨
火疫 菌 中的 c r p同源基 因, 命名 为 E cp 并通过 同源 重组 的方 法 , 建 了梨火 疫菌 的 c ar , 构 r 因突变 体 E A r p基 ac p 以及互 补子 , 行 了致 病 性 、 敏性 反应 、 进 过 胞外 多糖 、 毛运动 等 一系列 相关 表型 的鉴 定 。结果表 明,r 因 鞭 c p基 影响着 梨火 疫菌 的致病 性 、 胞外 多糖 、 动性 、 游 生长 情况 等 多种 生物 学特 性 , 然而 , a c E A r 能 引起烟 草过 敏 p仍
摘
要
梨 火 疫 菌 (rii m lvr) 引起 梨 、 Ewnaa yooa可 苹果 等 蔷 薇 科 ( oaee 植 物 的火 疫 病 。在菊 欧文 氏菌 R sca)
(r ii hyate )中 , 由 c E wnac r nh mi s r p基 因编 码 的环 腺 苷 酸 受 体 蛋 白 ( c ca eo ie n h sht c c l d n s p op ae( yi n mo AMP )
研 究 论 文
An Aril tc e
梨 火 疫菌 ( r iia yooa 环 腺 苷 酸受 体 蛋 白基 因 (r) E wna m l r) v c 的功 能分析 p
刘倩 倩 于洋 洋 宋俊 贤 胡 白石 ‘ 范加 勤 刘凤 权
南 京 农业 大学 农 业 部 病 虫 监 测 与 治 理 重 点 开 放 实 验 室 , 京 2 0 9 南 10 5 通 讯作 者 ,b@玎a. uc h s jue . d a
e pr s i n oft c i o y i e sa d r gu ai n o e p t g ni i n Er na c r s nt e x e so pe tn l ssg ne n e lto ft aho e ct i wi i h y a h mi I t i t d ,t e he h y n h ssu y h
性 反应 , 并且在 过 氧化 氢 敏感度 以及沉 降性 、 生物膜 和 AI . 号分子 的生成 方 面与 野 生型 菌 株相 比差 异 明 2信 显。 本研 究结果 说 明, 火疫 菌 c 梨 r 因对 病菌 的胞外 多糖 分泌 、 p基 生长 、 动性 以及致病 性方 面具有 关键作用 。 游 关键 词 梨火 疫 菌 ,r 因 , 病 性 , c p基 致 胞外 多糖
LuQi qa Y n y n S n n i H asi F nJ q LuF n q a i a i n n uYa g a g o gJ xa u n uB i a a i h i n i egun
Ke a r t r f M o i rn nd M a a e e t o l t Di e s s a d I s c s y L bo a o y o n t i g a n g m n f P a s a e n n e t,M i s y o rc l r o n nit f Ag u t e r i u
Abs r c Er n a a v r a e r i h f p a ,a pl ,a d ma y oft e Ros c a a l Th y lc ta t wi i myl o a c us s f e blg to e r p e n n h o i a e e f miy e c ci
.
a e oiemo p op a (A ) ee tr rt n ( R ) w ihi e c d db p gn , l s rc loei dn s n h sh t c MP rcpo oe C P , hc n o e yc ee pa cui l n n e p i s r ya ar
,
Naj gAgi lr iesy Naj g ni r ut eUnv rt, ni n c u i n
21 0 5 Ch n 0 9 , ia
C r so dn to, b@na . uc or p n iga h rh s jue . e u d a DO : 03 6 ̄i n17 —9 82 1 .5 0 I 1.9 9 .s . 47 6 .0 00 . 1 s 6 0