水稻籼粳杂种育性的QTL分析
水稻品种Lemont抗稻瘟病基因QTL的初步定位
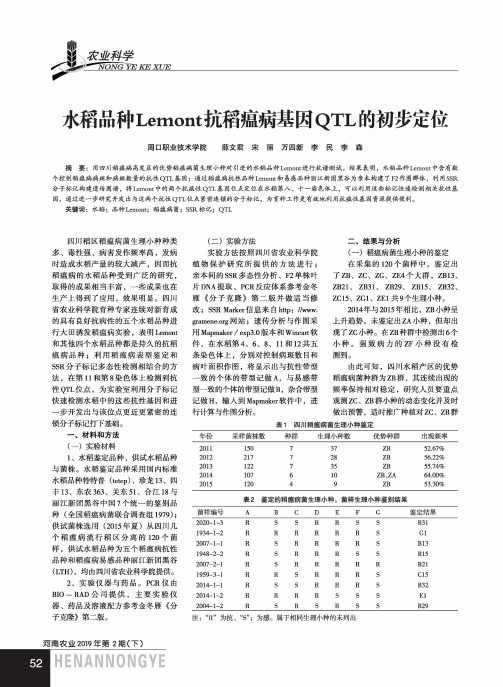
J I輕科学一NONG~Yl水稻品种Lemont抗稻瘟病基因Q T L的初步定位周口职业技术学院薛文君宋丽万四新李民李森摘要:用四川稻瘟病高发区的优势稻瘟病菌生理小种对引进的水稻品种Lemont进行抗谱测试,结果表明,水稻品种Lemont中含有数 个控制稻瘟病病斑和病斑数量的抗性QTL基因;通过稻瘟病抗性品种Lemont和易感品种丽江新团黑谷为亲本构建了 F2作图群体,利用SSR 分子标记构建遗传图谱,将Lemont中的两个抗瘟性QTL基因位点定位在水稻第八、十一染色体上,可以利用这些标记快速检测相关抗性基 因,通过进一步研究开发出与这两个抗性QTL位点紧密连锁的分子标记,为育种工作更有效地利用抗疸性基因资源提供便利。
关键词:水稻;品种Lemont;稻瘟病菌;SSR标记;QTL四川稻区稻瘟病菌生理小种种类多、毒性强、病害发作频率高,发病时造成水稻产量的较大减产,因而抗稻瘟病的水稻品种受到广泛的研究,取得的成果相当丰富,一些成果也在生产上得到了应用,效果明显。
四川省农业科学院育种专家连续对新育成的具有良好抗病性的五个水稻品种进行大田诱发稻瘟病实验,表明Lemont和其他四个水稻品种都是持久的抗稻瘟病品种;利用稻瘟病表型鉴定和 SSR分子标记多态性检测相结合的方法,在第11和第8染色体上检测到抗性QTL位点,为实验室利用分子标记快速检测水稻中的这些抗性基因和进一步开发出与该位点更近更紧密的连锁分子标记打下基础。
一、材料和方法(一)实验材料1、 水稻鉴定品种、供试水稻品种 与菌株。
水稻鉴定品种采用国内标准水稻品种特特普(tetep)、珍龙13、四丰13、东农363、关东51、合江18与丽江新团黑谷中国7个统一的鉴别品种(全国稻瘟病菌联合调查组1979);供试菌株选用(2015年夏)从四川几个稻瘟病流行稻区分离的120个菌样,供试水稻品种为五个稻瘟病抗性品种和稻瘟病易感品种丽江新团黑谷 (LTH),均由四川省农业科学院提供。
水稻种子休眠性及其QTL定位研究进展

2 0 年 第 2 卷 第 5期 07 l
作 物 研 究
水 稻 种 子休 眠性及 其 QTL定 位 研 究 进 展
黄 明 ,陈 立 云
( 南农业大学水稻研究所 , 沙 402 ) 湖 长 1 1 8
摘 要 :简要 介 绍 了 水 稻 种 子 的休 眠 性 及 其 传 统 遗 传 学 研 究 概 况 , 点 对 已报 道 的 种 子 休 眠 性 数 量 性 状 基 因位 点 进 行 了 重
义 ’ 。
原 因所造 成 , 也可 能 由多重原 因所 引起 。 因素之 间的 各 关 系也 比较 复杂 , 时彼 此 间存在 密切联 系 , 有 当某种 因 素 被 消除 , 其 它 因素仍 存 在 时 , 而 种子 依 然 不能 发 芽 。 有时 当一种 因素被 消除 时 , 一种 因素也 随之 消失 , 另 休
休 眠特 性是 长期 系统发 育 的结果 。多数 “ 系” 交稻 三 杂 制 种 的雄 性不 育 恢 复 系种子 的休 眠特 性 属 于这 类 型 。 种 子 收获 后 , 般 品种 的休 眠期 为 2 一 ~3个 月 , 如 明 例
为可 能[ 。迄 今 , 8 ¨ 利用 同工 酶标 记 、 I RF P标 记等 手 段对不 同的 遗传群 体 ,已初 步定 位 了多个与 水稻种 子
T k h s i ] 通过 传 统 的遗 传分 析 提 出 了水 稻种 a a a h 等 人
子休 眠性 的双基 因控 制模 型 。 大多数 的研究 表 明 , 但 种 子休 眠性 的强 弱不仅 决定 于个 体 的基 因 型且 受外 界环 境条 件 的影 响 。 , ]表现 为数 量性状 的遗 传特 性 。随着 分子 标 记技 术 的成 熟 和应 用 , 对水 稻 进行 数 量 性状 基 因座 ( u n i t etatlc s QTI 的定 位 和分 析 成 q a t ai ri ou , t v )
利用大粒籼_小粒粳重组自交系定位水稻生育期及产量相关性状QTL

利用大粒籼/小粒粳重组自交系定位水稻生育期及产量相关性状QTL占小登 于萍 林泽川 陈代波 沈希宏 张迎信 付君林 程式华* 曹立勇*(中国水稻研究所国家水稻改良中心/浙江省超级稻研究重点实验室,杭州310006;*通讯联系人,E-mail:shcheng@mail.hz.zj.cn;caoliyong@caas.cn)QTL Mapping of Heading Date and Yield-related Traits in Rice Using aRecombination Inbred Lines(RILs)Population Derived from BG1/XLJZHANXiao-deng,YUPing,LINZe-chuan,CHENDai-bo,SHENXi-hong,ZHANG Ying-xin,FUJun-lin,CHENGShi-hua*,CAOLi-yong*(National Center for Rice Improvement/Zhejiang Key Laboratory of Super Rice Research,China National Rice Research Institute,Hangzhou 310006,China;*Corresponding authors,E-mail:shcheng@mail.hz.zj.cn;caoliyong@caas.cn)ZHAN Xiaodeng,YU Ping,LIN Zechuan,et al.QTL mapping of heading date and yield-related traits in rice using arecombination inbred lines(RILs)population derived from BG1/XLJ.Chin J Rice Sci,2014,28(6):570-580.Abstract:Heading date and yield-related traits including plant height(PH),number of tillers per plant(NTP),paniclelength(PL),number of filled grains per panicle(NFGP),grain number per panicle(GNPP)and seed setting rate(SR)are important agronomic traits in rice.Detection of new QTL associated with heading date and yield-related traitshave great significance in new genes identification and utilization of rice.We used a recombination inbred lines(RILs)population derived from BG1/XLJ to detect QTL associated with heading date and yield-related traits in twoenvironments,effects of epistatic and interaction with environments were also analyzed.A total of 32QTL associatedwith heading date and yield traits were detected on chromosomes 2,3,4,5,6,7and 8.Furthermore,five pairs ofQTL intervals had significant two-factor epistatic effects,and 5QTL had significant interaction with environments.Many yield related QTL detected were identified near the intervals where yield-related QTL were detected in previousresearches.Among them,the QTL with large effects commonly clustered.Low epistatic and interaction withenvironments effects for yield related QTL,indicated small interactive effects between genes,genes and environments.Key words:rice;recombination inbred lines;heading date;panicle traits;quantitative trait locus占小登,于萍,林泽川,等.利用大粒籼/小粒粳重组自交系定位水稻生育期及产量相关性状QTL.中国水稻科学,2014,28(6):570-580.摘 要:生育期及产量相关性状如株高、单株有效穗数、穗长、每穗实粒数、每穗总粒数和结实率等是水稻重要的农艺性状,发掘新的生育期及产量相关性状QTL,对克隆控制水稻产量性状新基因及利用分子聚合育种改良水稻产量性状具有重要意义。
作物QTL分析的原理与方法

作物QTL定位方法与技术作物QTL定位的方法主要有传统连锁分析、基因芯片 技术和深度学习等。连锁分析通过群体遗传学手段,鉴定两个或多个基因位点 间的连锁关系,进而确定控制性状的QTL。基因芯片技术利用基因组wide的标 记分布,对大量基因位点进行同时检测,高效地定位QTL。深度学习则利用神 经网络等算法,自动化学习和识别数据中的特征,实现对QTL的精准定位。
四、自然群体
自然群体是指在没有人为干预下自然形成的群体,如野生种、地方品种、自然 变异群体等。这些群体通常具有丰富的遗传变异和复杂的遗传结构,对于研究 作物的适应性、抗逆性和产量等性状的遗传基础非常有用。此外,自然群体还 可以用于发现和克隆稀有或特殊的QTL。
五、基于基因组的作图群体
随着基因组学技术的发展,基于基因组的作图群体越来越受到重视。这种群体 可以通过重测序技术获得大量的SNP(单核苷酸多态性)标记,并利用这些标 记构建高密度的遗传图谱。这种图谱可以用于精细定位和克隆QTL,以及研究 基因组中的结构变异和非编码区基因组。
2、QTL分析的具体步骤
(1)数据采集:收集作物的基因型和表型数据。基因型数据可以通过高通量 测序技术获得,而表型数据则可以通过田间试验和室内分析等方法获得。
(2)作图:利用作图软件将基因型和表型数据组装成图,以展示它们之间的 关系。常用的作图软件包括QTL Cartographer、QTL IciMapping等。
原理
1、QTL的概念及定义
QTL是指作物基因组中控制数量性状的基因座位,它们可以通过影响表型变异 来影响作物的农艺性状。QTL通常分为两类:主效QTL和微效QTL。主效QTL是 指对表型变异起主要作用的QTL,而微效QTL则是指对表型变异起较小作用的 QTL。
水稻早期育性状与产量性状遗传分析

水稻早期育性状与产量性状遗传分析第一章:引言水稻是世界上最为重要的粮食作物之一,主要分布在亚洲、非洲和南美洲等地。
水稻的生长过程中会受到环境和基因因素的影响,其中育性状和产量性状是非常重要的两个方面。
对这两个性状进行遗传分析可以帮助我们理解水稻的产量和育性状的形成机理。
第二章:育性状的遗传分析育性状是描述水稻生长早期现象的性状,包括种子发芽率、生长势等。
育性状通常由多个基因控制,其中一些基因具有显性基因效应,另一些则具有隐性基因效应。
隐性基因通常被掩盖,直到这些基因的等位基因组合出现变化时才会显现出来。
对育性状进行遗传分析可以通过计算各个基因在遗传中的比重来确定这些基因的作用和相关性。
与其他作物的育性状相比,水稻的育性状遗传分析具有较高复杂性,因此需要进行更为深入的研究。
第三章:产量性状的遗传分析产量性状是描述水稻产量的性状,包括穗数、籽数、粒重等。
产量性状通常由多个基因控制,这些基因也可以具有显性和隐性基因效应。
产量性状遗传分析的方法之一是利用遗传显微镜分析染色体结构和基因组成。
另一个方法是利用统计学方法研究不同基因型之间的差异性。
通常,不同基因型的产量性状差异越大,则这些基因在遗传中的比重越大。
第四章:水稻早期育性状与产量性状的关系早期育性状与产量性状之间存在着密切的关系。
以种子发芽率和产量性状为例,种子发芽率高的水稻将会有更快、更强的生长势,这将带来更高的产量。
因此,研究水稻早期育性状和产量性状之间的关系具有重要的应用价值。
第五章:结论通过对水稻早期育性状和产量性状的遗传分析,可以更好地理解这些性状的形成机理。
同时,也可以为育种研究提供更加有力的工具和方法。
未来,我们需要继续深入研究水稻早期育性状和产量性状之间的关系,为水稻生产提供更加精确、高效的方法。
杂种优势遗传机理研究方法的讨论

杂种优势遗传机理研究方法的讨论(南京农业大学,南京 210095)摘要:主要介绍了分子生物学、细胞生物学、生理生化和数量遗传学等方面对作物杂种优势机理研究的进展及所用的方法,简单讨论了目前研究中存在的问题,同时对杂种优势今后的研究作出展望。
关键词:杂种优势;遗传机理;研究方法;Discussion of Research techniques on Heredity Mechanism of Heterosis Abstract:Advances on heredity mechanism of heterosis and research techniques with molecular biology, cell biology, physiology and biochemistry quantitative genetics is summarized.The main problems in present study were simply discussed and the prospects of future study were reviewed in this paper.Key Word: Heterosi s;Heredity mechanism;Research technique杂种优势是指遗传组成不同的亲本杂交产生的F1比双亲具有强的生活力、生长势、适应性、抗逆性和丰产性的生物学现象, 在水稻、玉米、油菜、棉花等大田作物和蔬菜作物中广泛地应用[1]。
以水稻为例,杂交水稻一般比常规稻增产20 %左右,我国杂水稻自1976 年大面积推广以来, 到2000 年已累计推广37 亿亩,增产稻谷3500亿公斤, 年增稻谷可养活6000万人口。
由于杂种优势是一种复杂的遗传现象, 受诸多因素影响, 自从德国学者Kolreture(1761-1766)在烟草杂交中发现杂种优势,并最早对其进行系统研究以来,各国科学家利用不同手段研究提出了多种假说解释其遗传机制, 如显性假说( dominance hypothesis)、超显性假说(overdominance hypothesis)、核质互作假说( nucleo -cytoplasmic interaction hypothesis) 及遗传平衡假说(genetic equilibrium hypothesis), 基因网络系统假说等[1]。
基于高密度遗传图谱的水稻糙米籽粒大小QTL定位

收稿日期:2023-10-07基金项目:国家自然科学基金(32260497);贵州省基础研究(自然科学)项目(黔科合基础-ZK〔2023〕一般178);贵州省科技计划项目(黔科合平台人才〔2018〕5263);贵州省科技计划项目(黔科合支撑〔2022〕重点028)作者简介:彭强(1986-),男,博士,副研究员,研究方向为水稻分子育种,E-mail:********************通信作者:朱速松(1966-),男,博士,研究员,研究方向为水稻分子育种,E-mail:*****************广东农业科学2023,50(12):96-103Guangdong Agricultural SciencesDOI:10.16768/j.issn.1004-874X.2023.12.009彭强,徐海峰,宫彦龙,吴娴,吴朝昕,吴健强,朱速松. 基于高密度遗传图谱的水稻糙米籽粒大小QTL 定位[J]. 广东农业科学,2023,50(12):96-103.基于高密度遗传图谱的水稻糙米籽粒大小QTL 定位彭 强,徐海峰,宫彦龙,吴 娴,吴朝昕,吴健强,朱速松(贵州省农业科学院水稻研究所,贵州 贵阳 550006)摘 要:【目的】籽粒大小是影响水稻产量的主要农艺性状之一。
采用籼稻品种V20B 与细长型爪哇稻品种CPSLO17衍生的重组自交系开展水稻糙米籽粒大小QTL 定位研究,挖掘遗传稳定的主效QTL,为优质高产稻品种培育提供新的基因资源和科学依据。
【方法】基于V20B/CPSLO17遗传背景的高密度遗传连锁图谱,结合150份重组自交系在4种环境(2019年贵州贵阳、2020年贵州贵阳、2021年贵州贵定、2021年海南三亚)中的糙米籽粒大小表型数据,采用IciMapping 4.0软件的ICIM-ADD 方法进行QTL 扫描。
【结果】亲本V20B 的糙米籽粒大小显著大于CPSLO17,重组自交系的糙米籽粒大小在4种环境间差异显著,均表现出连续的单峰分布。
水稻抽穗扬花期耐热性的QTL分析

摘
要
培育 高产稳产 、 对环 境钝感 的优质 杂交 稻新组合 已成 为 当前 水稻育 种 的重 要 目标 ; 近年 高温造成 水
稻 结实 率显 著下 降而 导致产 量 的严 重损 失 , 水稻耐 热性 的 改 良也 日益 迫切 。本研 究用 一个 籼粳 交重 组 自交
,
g pic dn 5 S resw s eut sr sl so ey l i rcn as T eg n t n a ema l ig 1 7S R makr a e n o o i r i dn y ci nu
合 贡献率 为 2 .1 8 %。 0 关键 词 水稻, S S R标 记, T , Q L 耐热 性
QT p igfr a oeac fh asl eido ie LMapn t lrn eo eT se P r f c o He T t o R
Z a g a ‘ Ya g i Jn af g H a g n S nQ C e nu Z egJ k i‘ hn o。 T n agK i n u n u u L e Mi n h nWef h n a u ’ i
cnt c dw t rc mbnn be n R L p p lt ncn is f n srm cos f rz t a n i o s ut i a e o ia tn rdl e(I ) o uai s to 8 ie o a rs Oya a v d a r e h i i o o s 1 l f 6 o si i c
Z o g o zoNo8 n r ast a a o i e gi, n eq a ta v a s o iQ L a a s nh a l - h n y u a . a dO y i p n aF nj adt u ni t e ri c ( T ) n l i o etoe z av j c n h t i t tl ys t r
籼粳稻问亚种特征表现及其QTL分析

试验材料为典型籼稻七山占和粳稻秋光构建 的 F 、 F 重 组 自交系( I) R L 群体 , 共计 15个株 系 , 氏指数测定 参考程 6 程 侃声 的方法 … ; 维管束性状 测定于齐穗期 在每个株 系中选取
2 1 程 氏 指数 及 维 管束 性 状 在群 体 中 的分 布 .
程氏指数及维管束性状 为区分籼 、 粳的主要形 态及解剖
和第二 节问大 、 小维管束数 目, 计算大维管束比和大小维管束
比。选用均匀分布于水稻 1 2条染色体 上的 50对 S R引物 0 S 进行筛 选 , 出在 父母本 上有 差异 并且 扩增条 带较 清 晰的 选 12对引物对群体 进行标记 分析。Q L分析采 用 Mamakr 2 T p re 进行群体遗传 图谱的绘制 , 采用 Q ee2 2 gn .9软件 进行 各性 状的基 因定位分析 , 区间作 图法 , 选用 L D值 3 0作 为阈 O . 值来判断 Q L的存在 , T s T Q L 命名原则遵循 文献 [O 。 1 ]
指数总分值 的热点 区域 ; 在第 1 染色体上检测出的叶毛 、 抽穗
2 2 程 氏指数 、 . 维管束数 目比 Q L分析 T
种特征之 间连锁遗传现象较少 , 籼粳交后代 亚种特征 各性 状已重新组合 , 呈现出一种非随机 分离 现象较少 的状态 。
关键词 : 水稻 ; 重组 自交系 ; 程氏指数 ; 维管束数 目比; T QL 中图分 类号 : 5 102 F 1 .3 文献标 志码 : A 文章编号 :02—10 (0 1 0 04 0 10 3 2 2 1 ) 6— O0— 3
项亚种特征均呈连续分 布, 群体均 呈由籼到粳 的以偏 粳类型
收稿 日期 :0 1 7—2 2 1 —0 7 基金项 目: 国家 自然科学基金 ( 号 : 3 06 ) 教育 部高等学校 博 编 3 7 86 ; 0 士学科专项科研基金( 编号 : 0 0 5 0 1 。 2 5 170 ) 0 作 者简介 : 艇 (9 5 ) 男 , 毛 18 一 , 辽宁沈 阳人 , 硕士研究 生 , 从事水 稻 常规育种及分子育种研究。E—ma:hnmat g9 5 6 . o 。 i cia oi 1 8 @13 cr l n n
优质高产籼粳杂交稻嘉丰优3_号的选育及制种技术

780㊀㊀2024年第65卷第4期收稿日期:2024-01-05作者简介:金浩晶,浙江东阳人,主要从事优质㊁高产㊁多抗水稻新品种选育,E-mail:1942152531@㊂通信作者:董国军,浙江龙游人,农艺师,主要从事水稻分子遗传育种研究,E-mail:dongguojun@㊂文献著录格式:金浩晶,曹郁青,郑利欣,等.优质高产籼粳杂交稻嘉丰优3号的选育及制种技术[J].浙江农业科学,2024,65(4):780-784.DOI:10.16178/j.issn.0528-9017.20240021优质高产籼粳杂交稻嘉丰优3号的选育及制种技术金浩晶1,曹郁青1,郑利欣1,厉伟杉1,马强1,董国军2∗(1.浙江可得丰种业有限公司,浙江磐安㊀322300;2.中国水稻研究所水稻生物育种全国重点实验室,浙江杭州㊀310006)㊀㊀摘㊀要:嘉丰优3号是嘉兴市农业科学研究院联合中国水稻研究所㊁浙江可得丰种业有限公司,以嘉禾112A (长粒型粳稻三系不育系)为母本与G1143(中籼迟熟型恢复系)为父本选育的三系杂交水稻新组合㊂该组合具有高产㊁优质㊁中抗稻瘟病等优势,于2021年4月通过浙江审定(浙审稻2021019),2021 2023年先后通过了江西省[(赣)引种 2021 第001号]㊁安徽省(皖引稻2021066)㊁福建省[(闽)引种 2022 第1号]㊁湖北省(鄂引种2022055)的引种备案,适宜在长江中下游地区作中稻种植㊂文章介绍了嘉丰优3号的选育过程㊁特征特性㊁高产栽培及制种技术等㊂关键词:优质高产籼粳杂交稻;嘉丰优3号;选育;制种技术中图分类号:S511㊀㊀㊀文献标志码:A㊀㊀㊀文章编号:0528-9017(2024)04-0780-05Breeding and seed production technology of a indica-japonica hybrid ricevariety Jiafengyou 3with high-yield and high-qualityJIN Haojing 1,CAO Yuqing 1,ZHENG Lixin 1,LI Weishan 1,MA Qiang 1,DONG Guojun 2∗(1.Zhejiang Kedefeng Seed Industry Co.,Ltd.,Panᶄan 322300,Zhejiang;2.China National Rice ResearchInstitute,State Key Laboratory of Rice Biology and Breeding,Hangzhou 310006,Zhejiang)㊀㊀Abstract :Jiafengyou 3is a new hybrid rice combination developed by Jiaxing Academy of Agricultural Sciences,China National Rice Research Institute,and Zhejiang Kedefeng Seed Industry Co.,Ltd.,which using Jiahe 112A (longgrain type japonica rice three line male sterile line)as the female parent and G1143(medium indica late maturing restoringline)as the male parent.This combination has advantages such as high yield,high quality,and moderate resistance to riceblast.It was approved by Zhejiang in April 2021(Zhe Authorized Rice 2021019),and from 2021to 2023,it has successively passed the introduction registration of Jiangxi Province [(Gan )Introduction 2021 No.001],Anhui Province (Wan Introduction Rice 2021066),Fujian Province [(Min)Introduction 2022 No.1],and Hubei Province(E Introduction Rice 2022055),making it suitable for planting medium rice in the middle and lower reaches of the YangtzeRiver.This article introduces the breeding process,characteristic characteristics,high-yield cultivation,and seedproduction techniques of Jiafengyou 3.Keywords :indica-japonica hybrid rice variety with high-yield and high-quality;Jiafengyou 3;breeding;seedproduction technology㊀㊀籼粳杂交稻作为一类新型水稻品种,通常被认为产量相对较高,但米质较差㊂选育单位围绕粳不籼恢这一籼粳亚种间杂种优势进行积极探索,创制粳不籼恢不育系和恢复系新材料,组配了优质高产的新型籼粳杂交稻组合嘉丰优3号,于2021年4月通过浙江审定(浙审稻2021019),2021 2023年先后通过了江西省[(赣)引种 2021 第001号]㊁安徽省(皖引稻2021066)㊁福建省[(闽)引种 2022 第1号]㊁湖北省(鄂引种2022055)的引种备案,适宜在长江中下游地区作中稻种植㊂1㊀亲本来源及选育过程1.1㊀母本嘉禾112A㊀㊀嘉禾112A 是嘉兴市农业科学研究院与中国水稻研究所合作选育的BT 型粳稻三系不育系,2020年5月通过浙江审定[浙审稻(不育系)2020013]㊂2011年嘉兴正季,利用粳型细胞质雄性不育系嘉禾212A 为母本,自育半矮生型长粒粳稻新品系嘉禾112作父本测交配组㊂2012年海南成对种植F 1代,与测交父本嘉禾112成对回交;后经海南 嘉兴多次回交和逐代逐株育性鉴定,2015年海南初定B 6F 1并少量制繁种,定名为嘉禾112A;2015年嘉兴正季继续回交并制繁种㊂转育各个世代进行套袋观察不育系自交结实率,同时对稻瘟病等多种农艺特性进行表现型鉴定㊂该不育系繁茂性佳,开花习性好,配合力较强,异交结实率中等,米质优,中抗稻瘟病㊂1.2㊀父本G1143的选育㊀㊀恢复系G1143是中国水稻研究所选育的偏籼型恢复系㊂2007年在杭州富阳试验基地用IR64与R25(献党/测64-7)进行杂交,通过分子标记辅助选育技术进行单株系统选育,通过富阳㊁海南10代选育而成的偏籼型恢复系㊂1.3㊀嘉丰优3号的选育㊀㊀2013年春,在海南用恢复系G1143分别与嘉禾112A㊁嘉禾212A 等测交配组,次年开展小区品比试验和小面积制种,2015年起由浙江可得丰种业有限公司在浙江嘉兴㊁磐安等地布点试种,观察其抗逆性㊁适应性㊁丰产性,最终表现长势繁茂㊁生育期适中㊁抗性强㊁米质优㊁产量高㊁后期熟色好的嘉禾112A /G1143组合入选,新组合暂定名嘉丰优3号㊂2021年4月通过浙江审定(浙审稻2021019),2021 2023年先后通过了江西省[(赣)引种 2021 第001号]㊁安徽省(皖引稻2021066)㊁福建省[(闽)引种 2022 第1号]㊁湖北省(鄂引种2022055)的引种备案,适宜在长江中下游地区作中稻种植㊂2㊀主要特征特性2.1㊀农艺性状㊀㊀该品种株型适中,植株较高,分蘖力中等偏弱,剑叶内卷㊁长挺㊂叶下禾,着粒中等,穗大粒多,谷壳黄亮,颖尖秆黄色,短顶芒,谷粒椭圆形,结实率较高㊂2.1.1㊀浙江省㊀㊀两年省区试平均全生育期138.2d,比甬优1540(CK)短5.9d㊂表1表明,该品种两年区试每667m 2有效穗数11.9万,株高125.2cm,每穗总粒数306.9粒,实粒数258.8粒,结实率84.5%,千粒重25.65g㊂与甬优1540比较结实率低6.45百分点,千粒重高2.60g㊂表1㊀嘉丰优3号与甬优1540生育期及主要农艺经济性状汇总分析结果(中稻)Table 1㊀Summary analysis results of growth period and main agronomic and economic traits of Jiafengyou3and Yongyou 1540(medium rice )品种名称全生育期/d 每667m 2有效穗/万株高/cm每穗总粒数每穗实粒数结实率/%千粒重/g 嘉丰优3号138.211.9125.2306.9258.884.5025.65甬优1540144.110.5118.6331.9302.190.9523.052.1.2㊀国家统一区试(长江中下游晚籼早熟组)㊀㊀两年嘉丰优3号国家统一区试(长江中下游晚籼早熟组)平均生育期114.3d,生育期比CK 早熟2.7d,表2表明,嘉丰优3号每667m 2有效穗16.5万,株高112.0cm,穗长23.0cm,每穗㊀㊀总粒数212.4粒,结实率79.5%,千粒重27.2g㊂与五优308比每穗实粒数多35.6粒,结实率低2.8百分点,千粒重高3.9g㊂2.1.3㊀国家统一区试(黄淮海粳稻组)㊀㊀2022年生育期154.8d,比CK 晚熟0.3d,每㊀㊀表2㊀嘉丰优3号与五优308生育期及主要农艺经济性状汇总分析结果(连作晚稻)Table 2㊀Summary analysis results of growth period and main agroeconomic traits of Jiafengyou 3and Wuyou 308(continuous late rice )品种名称全生育期/d 每667m 2有效穗/万株高/cm穗长/cm 每穗总粒数每穗实粒数结实率/%千粒重/g 嘉丰优3号114.316.5112.023.0212.4167.979.527.2五优308117.022.6106.221.8160.8132.382.323.3782㊀㊀2024年第65卷第4期667m 2有效穗16.5万,株高118.1cm,穗长24.3cm,每穗总粒数233.8粒,结实率80.3%,千粒重26.3g㊂2.2㊀稻米品质㊀㊀中稻:表3表明,嘉丰优3号平均整精米率50.6%,长宽比2.7,垩白度1.90%,透明度2.0级,胶稠度77.5mm,碱消值5.8级,直链淀粉含量13.6%,与甬优1540比较米质均为二等㊂表3㊀嘉丰优3号与甬优1540米质和抗病分析结果(中稻)Table 3㊀Analysis results of rice quality and disease resistance between Jiafengyou 3andYongyou 1540(medium rice )品种名称精米率/%整精米率/%长宽比垩白度/%透明度/级碱消值/级胶稠度/mm直链淀粉含量/%部标等级稻瘟病综合指数穗瘟损失率级别白叶枯病级嘉丰优3号70.7050.60 2.7 1.90 2.0 5.877.513.60二等 3.457甬优154071.9563.152.31.451.57.077.016.15二等3.957㊀㊀晚稻:表4表明,该品种糙米率81.2%,精米率72.3%,整精米率62.4%,粒长6.7mm,粒型长宽比2.9,垩白度1.5%,透明度2级,碱消值5.3级,胶稠度81mm,直链淀粉含量15.3%,综合评级为部标等级优3,米质优于五优308(普通)㊂表4㊀嘉丰优3号与五优308米质和抗病分析结果(连作晚稻)Table 4㊀Analysis results of rice quality and disease resistance between Jiafengyou 3andWuyou 308(continuous late rice )品种名称精米率/%整精米率/%长宽比垩白度/%透明度/级碱消值/级胶稠度/mm 直链淀粉含量/%部标等级稻瘟病综合指数穗瘟损失率级别白叶枯病级嘉丰优3号72.362.4 2.9 1.52 5.38115.3优3 2.857五优30872.260.72.81.825.86622.6普通5.7992.3㊀抗病性㊀㊀中稻:中抗稻瘟病,稻瘟病综合指数3.4,穗瘟损失率最高5级;感白叶枯病,白叶枯病最高7级;高感褐飞虱,褐飞虱最高9级,抗性和甬优1540不差上下㊂晚稻:稻瘟病综合指数2.8,穗瘟损失率级别5级,白叶枯病级7级,褐飞虱病级9级,抗性优于五优308㊂2.4㊀产量表现(区域试验与生产试验)2.4.1㊀浙江㊀㊀表5表示,2019年浙江省单季籼粳交偏籼稻区试,每667m 2平均产量712.2kg,比甬优1540(CK)增产3.6%;2020年区试每667m 2平均产量639.1kg,比甬优1540(CK)增产0.8%;两年省区试平均产量675.7kg,比甬优1540(CK)增产2.2%㊂2020年生产试验每667m 2平均产量628.9kg,比甬优1540(CK )减产0.9%,产量略高于甬优1540㊂2.4.2㊀国家统一区试(长江中下游晚籼早熟)㊀㊀表6表示,嘉丰优3号2021年初试每667m 2平均产量628.48kg,比五优308(CK)增产4.97%;2022年续试每667m 2平均产量616.43kg,比五优㊀㊀表5㊀嘉丰优3号每667m 2产量(浙江省区域试验和生产试验)Table 5㊀667m 2yield of Jiafengyou 3(regionaland production trials in Zhejiang Province )试验名称嘉丰优3号/kg甬优1540(CK)/kg 比CK 增减产/%2019年区域试验712.2687.5 3.62020年区域试验639.1634.30.82020年生产试验628.9634.3-0.9表6㊀嘉丰优3号每667m 2产量(国家统一区域试验和生产试验)Table 6㊀667m 2yield of Jiafengyou 3(nationalunified regional and production trials )试验名称嘉丰优3号/kg 五优308(CK)/kg 比CK 增减产/%2021年区域试验628.48598.74 4.972022年区域试验616.43579.93 6.292022年生产试验547.07539.171.47308(CK)增产6.29%,两年区试667m 2平均产量622.46kg,比五优308(CK)增产5.62%;生产试验667m 2嘉丰优3号平均产量547.07kg,比五优308(CK)增产1.47%,嘉丰优3号具有明显的产量优势㊂2.4.3㊀国家统一区试(黄淮海粳稻C组)㊀㊀2022年初试每667m2平均产量719.71kg,比CK增产15.86%,增产点比例91.7%㊂3㊀栽培技术要点3.1㊀适时播种,培育壮秧㊀㊀作中稻栽培于4月下旬至6月上旬播种,667m2秧田用种量10~12kg,大盘机插秧每盘50g, 667m2大田用种量0.75~1.5kg㊂3.2㊀适龄移栽,合理密植㊀㊀手插秧龄25d之内,机插20d以内,秧苗二叶一心期和移栽前5~7d,每667m2施尿素5kg,以培育多蘖壮秧,有效穗数13万~15万穗利于高产㊂单季手插密度27cmˑ27cm,双本插,机插密度30cmˑ21cm㊂3.3㊀肥水管理㊀㊀应根据当地的土地肥力水平,合理运筹肥水,实施健身栽培㊂每667m2施复合肥40kg,移栽返青晒田后结合除草,每667m2施尿素7.5kg,隔4~5d后每667m2再施尿素7.5kg加氯化钾10kg;每丛分蘖数15个左右时,排水烤田7d左右,促进根系下扎和稻株基部粗壮,此后灌水2d,搁田5~7d,间歇灌排,直至孕穗;主茎叶枕距为零时灌5~10cm水层,根据苗情667m2施尿素7.5kg加氯化钾7.5kg或复合肥20kg,以提高颖花成花率和结实率㊂如叶片宽长㊁叶色偏深则不可施穗肥,以防贪青和倒伏㊂抽穗扬花结束后间歇灌水,干湿交替,直至收割前7~10d断水,严禁断水过早,直到90%以上谷粒黄熟后收割㊂3.4㊀种子处理和病虫害防治㊀㊀播种前选对药剂,做好浸种,防治恶苗病;稻曲病要在破口前10d和破口期选对口药剂连防两次;水稻抽穗前后慎用咪唑类杀菌剂;严格按规范施用除草剂,严禁在水稻拔节或幼穗分化后施用化学除草剂;结合大田生产和病虫害预报,及时做好稻瘟病㊁纹枯病㊁稻飞虱㊁螟虫㊁白叶枯病等病虫害防治[1-2]㊂3.5㊀风险提示㊀㊀特别注意防治稻瘟病㊁稻曲病㊁白叶枯病㊂4㊀高产制种技术4.1㊀播种期的确定㊀㊀播种期需根据嘉禾112A和G1143的生育期来制定播种差期,父本G1143为中籼材料,花期相遇是杂交稻制种获得高产的前提㊂4.2㊀培育期壮秧㊀㊀父母本种子用浸种灵等抗菌剂浸种24~36h㊂秧田父母本播种量每667m2不超过10~20kg,父本要求稀播,以培育分蘖壮秧,播后及时塌谷可以预防麻雀和鼠害㊂早施断奶肥(二叶一心)和起身肥(移栽前),秧板上水前要求秧沟有水㊁秧板湿润,上水后保持一定水位;秧龄20~25d左右带分蘖移栽㊂4.3㊀合理种植,做好花期预测㊀㊀父母本种植比为2ʒ12较为理想,父本按株行距33.3cm,株距23.3cm,单本或双本插栽㊂母本株行距为16.7cmˑ16.7cm,父母本间距以20cm为宜㊂加强田间栽培管理,促进父母本平衡生长并及时搁田,及时观察父母本幼穗分化进程,使父母本花期处于最佳相遇状态㊂薄水分蘖㊁浅水扬花㊁活水养稻到老㊂4.4㊀肥水管理㊀㊀每667m2施氮㊁磷㊁钾肥比例为1.6ʒ2.0ʒ1.0,底肥主要以氮㊁磷肥为主,适施保花肥,施纯氮2.5~3.0kg,促进大穗,从而提高制种产量㊂4.5㊀授粉㊀㊀授粉以人工辅助授粉方式进行,从而提高不育系异交结实率㊂中午开花后每天赶2~3次粉,旺花期时要求赶3~4次,如遇阴雨天需抢晴赶粉㊂4.6㊀防治病虫害㊀㊀在病虫防治上宜采取化学与生物综合防治的方法,减少化学药剂的施用㊂重点做好纹枯病㊁稻飞虱㊁稻瘟病的预防㊂除采用高效低毒的化学药剂防治病虫害外,还可以采用频振式杀虫灯等方式捕杀二化螟㊁稻纵卷叶螟㊁飞蛾,具体病虫害的预测及防治时间以当地植保部门发布的病虫情报和防治意见为准㊂4.7㊀严把质量关㊀㊀为保证制种纯度,需严把亲本质量关和亲本繁殖田隔离关,繁种田四周要保持150~200m间隔,最好选择天然隔离田块进行繁种,防止不同品种互相串花㊂做好亲本种田间去杂关,父母本移栽后应经常进行田间去杂工作,特别是喷施九二〇前要加大去杂力度,彻底去除异型株及同型可育株㊂收割时严防机械和人为混杂,父本授粉结束后784㊀㊀2024年第65卷第4期及时割去父本,以防父本混进杂交种,同时机器要清理干净㊂种子收割后应及时进行田间纯度鉴定,有杂质的应及时进行加工精选,种子保存期间应经常测定含水量㊁发芽率,以保证大田用种质量㊂5㊀示范推广效果㊀㊀嘉丰优3号适于机械化和大规模生产,米质稳定性好,出米率高,晶莹剔透,米饭有劲道,冷饭不回生㊂2022年因长时间持续罕见高温导致许多水稻品种脱肥早衰,产量严重降低,但嘉丰优3号产量和品质都很稳定,以优异的表现和品质获得了市场的认可㊂参考文献:[1]㊀赵力勤,杨华军,朱东平,等.嘉禾优7245品种特性及机械化高产栽培技术[J].现代农机,2020(1):42-43. [2]㊀张怀杰,宋扬.籼粳杂交稻嘉丰优2号特征特性及栽培技术[J].浙江农业科学,2020,61(10):1977-1978.(责任编辑:董宇飞)。
控制水稻剑叶形态相关性状的数量基因位点(QTL)的定位

D nl(9 8 首次提出水稻理想株型模式是叶片小而挺 ; oa 16 ) d 角田重三郎(9 1 认为叶片宜厚而直 18 )
立 ; 岛省 三 (9 1 提 出理 想稻 的上位 叶要 短 小 并 直立 ; 来 , 国学 者提 出超 高产 杂 交 稻理 想 松 18 ) 近 我
维普资讯
第3 第 2期 6卷
2007 年 4 月
上海师范大学学报 ( 自然科学版 )
Ju a o hn hi o lU iesy N mr Sine) o r l f aga r nvri ( a  ̄ c cs n S N ma t e
6 7
杂交产生的回交重组 自交系(akr s eo b atnr ns BL ) bcc s r m i n i e le , Is群体来检测剑叶形态 中 3 o c n b di 个主要 性状的 Q L 期望检测到新的 Q L T, T 遗传位点 , 能用于今后株型分子育种.
1 材料与方法
株型的主要形态特征是上部 3片功能叶要长、 窄、 厚 , 直、 凹、 并提出了具体形态指标 . ’ 在剑叶形态的 遗传研究上, 过去一些学者 研究了籼粳杂种剑叶形态的遗传传递规律及其相关性 , 发现叶长、 叶宽 和叶面积均为多基因控制的数量性状 , 并发现剑叶长宽及叶面积在 F 代呈现较高遗传力, 叶宽基因作 用 主要表 现为加性 效应 , 叶长 、 叶面积则 主要表 现为显 性效应 . 年来 , 近 以分子连 锁 图谱 为基 础 的数 量性 状基 因研究 迅速 发展 , 稻剑 叶形 态相关 性状 的数 量基 因位 点 ( T ) 位 也 有一 些报 道 l 引. 研 水 QL定 3 ’ 本
个 Q L 其两侧的紧密分子标记在水稻株型分子育种上具有一定应用价值. T, 关键 词 : 水稻 ; 关 ; 叶形 态 ; 量性状 位点 ( T ) 相 剑 数 Q L
水稻孕穗期耐冷性QTLs分析

1 材料与方法
1. 1 材料来源与作图群体构建 耐冷基因源“吉冷 1 号”来源于吉林省农业科学 院水稻研究所 ,冷敏基因源“密阳 23”来源于韩国农 村振兴厅作物科学院 。1999 年配制杂交组合获得 F1 代 ,2000 - 2001 年经繁殖加代分别获得 F2 个体 200 个和 F3 家系 200 个 。 1. 2 水稻耐冷性鉴定
(1 中国农业科学院作物品种资源研究所 ,北京 100081 ;2 西北农林科技大学农学院 ,陕西杨凌 712100 ;3 韩国农村振兴厅作物科学院 ,韩国水原 4412100 ; 4 汉城大学校农业生命科学大学农学科 ,韩国汉城 1512742)
摘 要 : 本研究以籼粳交“密阳 23Π吉冷 1 号”的 F2∶3 代 200 个家系为作图群体 ,在韩国春川进行冷水胁迫下水稻耐冷性 鉴定 ,并以利用 SSR 标记构建的分子连锁图谱为基础 ,对水稻孕穗期耐冷性及其相对耐冷性进行数量性状位点 (Q TL s) 分析 。结果表明 ,在第 1 、2 、4 、11 和 12 染色体上检测到与孕穗期耐冷性相关的 Q TL 各 1 个 ,对表型变异的解释率为 5. 6 %~812 % ;在第 1 、3 、4 和 11 染色体上检测到与孕穗期相对耐冷性相关的 Q TL 各 1 个 ,对表型变异的解释率为 5. 9 %~ 1013 %。所检测到的耐冷性 Q TL 的增效等位基因多数来自吉冷 1 号 ,基因作用的方式主要为部分显性 、显性和超显性 。 关键词 : 水稻 ;冷水胁迫 ;孕穗期耐冷性 ;微卫星标记 ;数量性状位点 中图分类号 : S511
水稻籼粳杂种育性QTL定位及其效应分析

Ab s t r a c t T h e F ,p o p u l a t i o n o f Z h e n s h a n 9 7 B /x i u s h u i 1 3 we r e u s e d f o r t h e ma t e r i a l t o t e s t t h e e f f e c t o f t h e
热带 作 物 学报 2 0 1 4 ,3 5 ( 2 ) :2 4 6 — 2 5 2
C h i n e s e J o u r n a l o f T r 0 p i c a 1 C r o
水稻籼粳杂种 育性 QT L定位及 其效应分析
吴建 梅 ,林 荔 辉 ,林 培 清 ,官 华 忠 , 陈 志伟 ,吴 为 人
q P F 5 c o n t r o l l i n g t h e p o l l e n f e r t i l i t y w i t h a v a l u e o f a d d i t i v e e f f e c t一 8 . 6 5 a n d a c o n t r i b u t i o n r a t e o f p h e n o t y p e
WU J i a n me i ,L I N L i h u i ,L I N P e i q i n g ,GUAN Hu a z h o n g ,C HEN Z h i we i ,W U We i r e n
K e y l a b o r a t o r y o fMi n i s t y r o fE d u c t a i o n f o r G e n e t i c s , B r e e d i n g a n d M u l t i p l e U t i l i z a t i o n f o C r o p s ,
水稻抽穗期QTL与环境互作分析

第28卷第6期作 物 学 报V o l.28,N o.6 2002年11月 771~776页A CTA A GRONOM I CA S I N I CA pp.771~776 N ov.,2002水稻抽穗期QT L与环境互作分析Ξ李泽福1,2 周 彤1 郑天清1 罗林广1 夏加发2 翟虎渠3 万建民1,3(1南京农业大学作物遗传与种质创新国家重点实验室,南京农业大学水稻研究所,江苏南京210095;2安徽省农业科学院水稻研究所,安徽合肥230031;3中国农业科学院,北京100081)摘 要 本文利用由98个家系组成的N i pponbare Kasalath N i pponbare回交重组自交系(backcro ss inbred lines,B I L s)作图群体(BC1F9)和混合线性模型的Q TL定位方法,联合分析南京、合肥和海南3个不同地点的水稻抽穗期Q TL及Q TL与环境互作。
检测到8个抽穗期Q TL,分别位于第1、2、3、4、6、7、8染色体上,其中,第3染色体上有2个Q TL。
单个Q TL的加性效应大小范围为1.46天~4.58天,对性状的贡献率在1.49%~12.2%之间,加性效应总的贡献率为34.13%。
有6个Q TL与环境存在显著互作,单个Q TL与环境互作效应的贡献率在2.18%~15.84%之间,互作效应总的贡献率为41.11%,大于加性效应的总贡献率。
位于第4、6和7染色体的3个Q TL在3个不同地点均与环境存在显著互作,并且,在南京和合肥的互作效应均使抽穗期延长,在海南的互作都使抽穗期缩短,表现感光性特征。
对抽穗期Q TL检测的稳定性进行了分析。
关键词 水稻,抽穗期,混合线性模型,Q TL与环境互作中图分类号:S512 文献标识码:AAna lysis of QT L×Env i ronm en t I n teracti on s for Headi n g Da te of R i ce(O ryza sa t i-va L.)L I Ze2Fu1,2 ZHOU Tong1 ZH EN G T ian2Q ing1 LUO L in2Guang1 X I A J ia2Fa2 ZHA I H u2Q u3WAN J ian2M in1,3(1S tate K ey L aboratory of C rop Genetics and Ger m p las m E nhance m ent,R ice R esearch Institute,N anj ing A g ricultural U niversity,N anj ing210095, China;2R ice R esearch Institute,A nhui A cad e m y of A g ricultural S ciences,H ef ei230031,China;3Chinese A cad e m y of A g riculture&S ciences,B ei2 j ing100081,China)Abstract Q uan titative trait L ocus(Q TL)con tro lling heading date of rice acro ss th ree environm en ts w as ana2 lyzed using98backcro ss inbred lines derived from a subs pecific cro ss of N i pponbare Kasalath N i pponbare by m ixed linear model app roach.8Q TL fo r heading date w ere m apped in the ch romo s om es of1,2,3,4,6,7 and8w ith2Q TL in ch romo s om e3.T he general con tributi on of additive Q TL w as34.13%,the p ropo rti on of pheno typ ic variati on exp lained by individual additive Q TL ranged from1.49%to12.2%,and additive effects ranged from1.46~4.58days.6Q TL w ere founded to be affected sign ifican tly by environm en ts,the p ropo r2 ti on of pheno typ ic variati on exp lained by individual Q TL×environm en t in teracti on ranged from2.18%~15. 84%,and the general con tributi on w as41.11%w h ich w as larger than that of additive Q TL.3Q TL in ch ro2 mo s om e4,6and7had sign ifican t in teracti on s acro ss all th ree environm en ts,and the Q TL×environm en t in ter2 acti on in N an jing and H efei delayed the heading date and that in H ainan advanced the heading date,w h ich i m2 p lied that the3Q TL w ere sen sitive to pho toperi od.T he stability of heading date Q TL detecti on w as discussed.Key words R ice;H eading date;M ixed linear model;Q TL×environm en t in teracti on基金项目:农业部“948”项目(201002A)和“教育部优秀骨干教师基金”资助。
全基因组关联分析定位水稻分蘖角度QTL

中国水稻科学(Chin J Rice Sci), 2024, 38(3): 266-276 266 DOI: 10.16819/j.1001-7216.2024.230904全基因组关联分析定位水稻分蘖角度QTL朱裕敬#桂金鑫#龚成云 罗新阳 石居斌 张海清*贺记外*(湖南农业大学农学院, 长沙 410128;*通信联系人,email:**********************;*****************.cn)QTL Mapping for Tiller Angle in Rice by Genome-wide Association AnalysisZHU Yujing#, GUI Jinxin#, GONG Chengyun, LUO Xinyang, SHI Jubin, ZHANG Haiqing*, HE Jiwai*(College of Agronomy, Hunan Agricultural University, Changsha 410128, China; *Correspondingauthor,email:**********************;*****************.cn)Abstract:【Objective】Tiller angle is a critical agronomic trait influencing rice yield. Identifying rice tiller angle QTL (genes) and detecting their elite haplotypes can be beneficial for developing ideal rice varieties. 【Method】333 core germplasms from the rice 3K resources were utilized as research materials. These germplasms were cultivated in Yunyuan and Chunhua of Hunan Agricultural University in 2020 and 2022, respectively. Tiller angles of various germplasms were measured during the heading stage. Genome-wide association analysis was conducted using the MLM model of TASSEL 5.2, combined with the genotypes of the germplasms. 【Results】Six QTL for tiller angle were identified on rice chromosomes 2, 5, 6, 9, and 12, designated as qTA2, qTA5, qTA6.1, qTA6.2, qTA9, and qTA12, respectively. These QTL explained phenotypic variation ranging from 6.23% to 16.22%. Notably, qTA9 co-localized with the major QTL TAC1 for tiller angle, while the other five QTL were newly discovered. Candidate gene analysis was conducted for these five QTL. The candidate genes for qTA2 and qTA6.1 were identified as Os02g0817900 and Os06g0682800, respectively. Os02g0817900 encodes a rice cytochrome P450 family protein, while Os06g0682800 encodes a zinc finger domain protein.【Conclusion】This study successfully identified new QTL for tiller angle in rice and analyzed candidate genes, offering valuable insights for the cloning of tiller angle QTL (genes) and genetic improvement of tiller angle in rice.Key words: rice (Oryza sativa L.); tiller angle; QTL; candidate gene; haplotype摘 要:【目的】水稻分蘖角度是影响水稻产量的关键农艺性状,挖掘水稻分蘖角度QTL(基因)及其优势单倍型,有助于构建水稻理想株型。
水稻颖花退化的遗传研究进展

水稻颖花退化的遗传研究进展作者:戚华雄,杜雪树,李进波来源:《湖北农业科学》 2014年第24期戚华雄,杜雪树,李进波(湖北省农业科学院粮食作物研究所,武汉430064)摘要:水稻(OryzasativaL.)颖花数是产量构成的重要因子之一,每穗颖花数的多少取决于幼穗分化前期颖花的分化数和幼穗分化后期颖花的退化数两个方面。
水稻颖花退化现象在水稻生产中普遍存在,它包括穗基部颖花退化和穗顶部颖花退化两种。
水稻颖花退化性状是复杂的数量性状,可能受基因型、栽培条件以及生态因子等影响。
利用不同的遗传作图群体,目前定位了18个水稻颖花退化QTL,这些QTL分布在第1至第11号染色体上,但贡献率均不大。
利用2个颖花退化的突变体确定了2个候选基因,但还有待于进一步研究。
并对育种及生产中水稻颖花退化的问题进行了初步分析。
关键词:水稻(OryzasativaL.);颖花退化;遗传研究;QTL中图分类号:Q75;S511文献标识码:A文章编号:0439-8114(2014)24-5905-03DOI:10.14088/ki.issn0439-8114.2014.24.001收稿日期:2014-10-21基金项目:湖北省农业科技创新中心资助项目(2011-620-002-001)作者简介:戚华雄(1964-),男,湖北仙桃人,研究员,主要从事水稻遗传育种研究,(电话)027-87389224(电子信箱)qi_huaxing@163.com。
水稻(OryzasativaL.)单位面积颖花数是产量形成的重要基础,它由单位面积的有效穗数与每穗颖花数的乘积构成。
不同品种之间每穗颖花数差异较大,同一品种在不同栽培管理条件下每穗颖花数的变幅也较大,每穗颖花数的多少取决于幼穗分化前期颖花的分化数和幼穗分化后期颖花的退化数两个方面。
水稻颖花退化现象在水稻生产中普遍存在,生产上的一些主栽品种例如两优培九、扬两优6号等在穗分化阶段遭遇不适气候条件(例如低温、干旱等)会出现颖花退化现象,在颖花退化严重的情况下水稻产量会大幅降低。
水稻叶片气孔性状QTL的初步定位

水稻叶片气孔性状QTL的初步定位郭志富;李晓林;赵明辉;马殿荣;马慧;徐海;徐正进【摘要】利用气孔性状差异明显的粳稻和籼稻品种构建的Sasanishiki/Habataki//Sasanishiki///Sasanishiki回交重组自交系群体为试验材料,根据已有遗传连锁图谱选定236个RFLP分子标记,对水稻齐穗剑叶气孔性状进行QTL分析。
共检测到4个气孔性状相关的QTL,包括3个控制气孔密度的QTL(qSD2、qSD4和qSD12),分布在第2、4和12号染色体上,贡献率分别为13.11%,11.76%,22.70%;1个控制气孔宽度的QTL(qSW3),位于第3号染色体上,贡献率为10.04%。
气孔密度和气孔宽度的增效等位基因分别来源于粳稻和籼稻。
气孔密度与气孔长度存在极显著负相关(r=-0.509**),气孔长度与气孔宽度呈极显著正相关(r=0.302**)。
4个QTL位点除了qSW3与前人研究的控制全氮含量的qTLN-3a重叠外,其他QTL均为未被检测到的位点,反映了气孔性状遗传机理的复杂性。
为揭示水稻气孔性状的遗传机理以及籼粳稻气孔性状差异的分子机制提供重要参考。
%Using backcross recombinant inbred lines derived from a cross between a tropical japonica,Sasanish-iki and an indicavariety,Habataki,quantitative trait loci( QTLs) controlling the stoma related traits of flag leaf by 236 RFLP molecular markers from genetic linkage map in rice were analyzed. A total of four QTLs for stoma related traits were identified. Three putative QTLs (qSD2,qSD4 and qSD12) for stomatal density were mapped on chromo-some 2,4 and 12 with the contribution of each QTL was 13. 11%,11. 76% and 22. 70%,respectively. On chromo-some 3 of rice,one QTL(qSW3)for stomatal width was detected,with the explained variance of 10. 04%. The alleles for stomatal density andstomatal width were contributed by Sasanishiki and Habataki,respectively. Significant nega-tive differences were found between stomatal density and stomatal length(r= -0. 509A),while significant positive correlations were observed between stomatal length and stomatal width(r=0. 302A). All the three QTLs controlling stomatal density were not previously reported,but qSW3 controlling stomatal width was overlapped with qTLN-3a controlling total nitrogen content from other study. It reflected the genetic complexity of stomatal traits. These results will help to reveal the genetic mechanism of stomatal trait in rice and provide vital reference for the molecular mech-anism of difference of stomatal traits between indica and japonica rice.【期刊名称】《华北农学报》【年(卷),期】2014(000)003【总页数】5页(P1-5)【关键词】水稻;剑叶;气孔性状;数量性状位点【作者】郭志富;李晓林;赵明辉;马殿荣;马慧;徐海;徐正进【作者单位】沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866; 沈阳农业大学生物科学技术学院,辽宁沈阳110866;沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866;沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866;沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866;沈阳农业大学生物科学技术学院,辽宁沈阳 110866;沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866;沈阳农业大学水稻研究所,农业部东北水稻生物学与遗传育种重点实验室,辽宁沈阳 110866【正文语种】中文【中图分类】S511.03在陆生植物的进化过程中,形成了一种叫气孔的表皮结构,在调节植物光合作用、蒸腾作用以及水分利用中扮演着至关重要的角色,是物质生产能力、水分胁迫和温度胁迫抗性的重要因素[1-6]。
水稻籼粳杂交不育机理研究进展-

水稻籼粳杂交 不育
×
籼粳杂种不育控制基因
1
S5基因
三等位基因系统座模型
2
Sa基因
两基因/三元件互作模型
S5基因的发现
1984年,日本的水稻育种家池 桥宏认为籼粳杂种的不育性主要受 第6染色体S5座位的等位基因控制。 籼稻、粳稻的等位基因分别是S5-i
和S5-j,S5-n被称为广亲和基因,
将具有S5-n基因的品种称之为广 亲和品种,它与籼稻和粳稻杂交的 后代都表现比较正常的育性
互作造成。
一些研究者发现正反交差异不大。因此,可能
在一些特殊的籼粳杂交组合存在这种质核不协调。
×
籼粳杂交——染色体差异
籼粳杂种不育是由于籼、粳两者染色体间在结构 上存在微小差异,使得F1染色体配对不正常导致不育。 一些研究者发现籼粳杂种染色体配对是正常的。 因此,染色体行为异常与杂种不育无直接关系。
籼粳杂交:
1928年日本学者Kato 报道: 不育 或半 不育
引起籼粳杂种不育的原因
1、雄配子败育
2、雌配子败育
3、花药开裂障碍 4、花粉在柱头上萌发障碍 5、雌雄异熟 6、环境条件的影响
籼粳杂种不育性的遗传机理
1
三 种 假 设
质核不协调
2
染色体结构差异
3
不育基因控制
籼粳杂交——质核不协调
细胞质内蛋白质与酶的平衡在籼粳杂合核基因 组作用下发生波动,从而引起孢子发育的变化,导 致不育或半不育。可通过正反交来判定是否是质核
籼粳杂种花粉不育基因的定位

籼粳杂种花粉不育基因的定位陈静;江玲;王春明;池桥宏;翟虎渠;万建民【期刊名称】《作物学报》【年(卷),期】2006(32)4【摘要】To estimate QTL for the pollen sterility, an F2 population of 180 plants from an indica / japonica hybrid was tested with 110 molecular markers. By the genome wide screening, three QTLs were identified for the pollen sterility on chromosomes 3, 5, 7, respectively. In addition, a total of nine QTL for segregation distortion via male gametes were mapped on rice chromosomes, of which seven QTLs were suggested to be the same type of gamete abortion as found at ga14 and ga-11. The distortion data seemed to be more sensitive to detection of pollen abortion genes in the F1 hybrid than the morphological inspection of pollen. Two QTLs for the segregation distortion on chromosomes 5 and 6 indicated an abnormally high frequency of heterozygous genotype. At a locus indicated by qHPS-5, japonica-type homozygote showed a lower fertility level than the heterozygotes and indica-type homozygote. The molecular markers identified here would be useful in introducing a set of new and known sterility-neutral alleles to solve the pollen sterility problem in subspecificF1 hybrids. Further, the present results could be helpful to combine as many neutral alleles for distortion as possible on whole genome basis to stabilize the pollen fertility as a whole.%以IR36(indica)和热研2号(japonica,广亲和品种)为亲本,构建了包含180个单株的F2群体及包括110个标记的分子连锁图谱.利用该F2群体,进行了水稻花粉不育数量性状基因座(quantitative trait locus,QTL)的检测和遗传效应分析,共检测到3个花粉不育QTL,分别位于第3、5、7染色体上,此外,共检测到9个由雄配子引起的偏分离QTL,其中7个与ga-14和ga-11位点的配子败育类型相同.与花粉形态鉴定相比,偏分离的数据对检测F1杂种花粉败育基因更为敏感.在第5、6染色体上控制偏分离的2个QTL位点,其杂合基因型出现的频率偏高.在qHPS-5位点,粳型纯合子表现出比杂合子和籼型纯合子更低的育性水平.本研究获得的分子标记将有助于聚合尽可能多的中性亲和基因以解决亚种间F1杂种的花粉不育性问题.【总页数】7页(P515-521)【作者】陈静;江玲;王春明;池桥宏;翟虎渠;万建民【作者单位】南京农业大学作物遗传与种质创新国家重点实验室,江苏省植物基因工程技术研究中心,江苏,南京,210095;南京农业大学作物遗传与种质创新国家重点实验室,江苏省植物基因工程技术研究中心,江苏,南京,210095;南京农业大学作物遗传与种质创新国家重点实验室,江苏省植物基因工程技术研究中心,江苏,南京,210095;日本大学生物资源学部,日本国神奈川县,252-8510;中国农业科学院,北京,100081;南京农业大学作物遗传与种质创新国家重点实验室,江苏省植物基因工程技术研究中心,江苏,南京,210095;中国农业科学院作物科学研究所,北京,100081【正文语种】中文【中图分类】S511【相关文献】1.水稻籼粳亚种间杂种广亲和基因与不育基因 [J], 李万昌;王永飞;马三梅;李景原;王太霞2.籼粳杂种不育的遗传模式及亲和基因的研究进展 [J], 丁效华3.根据杂种不育性位点S—5n等位基因的PCR标记预测水稻籼/粳杂交的成功[J], Will.,CE;谢国禄4.豫粳6号保持系与恢复系TR2604间杂种花粉不育基因的定位 [J], 张宏根;孙一标;封智蔷;钱凯;裴艳;李闯;汤述翥;梁国华;顾铭洪5.水稻籼粳亚种间杂种低温花粉不育的QTL分析 [J], 杨杰;翟虎渠;王才林;仲维功;邹江石;池桥宏;万建民因版权原因,仅展示原文概要,查看原文内容请购买。
栽培稻与普通野生稻BC2F2群体产量相关性状的QTL分析

栽培稻与普通野生稻BC2F2群体产量相关性状的QTL分析栽培稻与普通野生稻BC2F2群体产量相关性状的QTL分析以广陆矮4号(Oryza sativa ssp.indica)为母本及轮回亲本,普通野生稻(Oryza rufipogon)为父本,分单株连续回交2次,构建BC2F2群体.首先用241对具有双亲多态性的SSR标记对BC2F1单株进行代换片段分析,在此基础上,根据BC2F1的表型选产量较优的单株自交获得BC2F2群体,用代换片段上具有双亲杂合型基因型的24对SSR标记进行QTL定位.在所选的BC2F1单株上,共检测到分布于7条染色体上的20个野生稻的代换片段,平均每条染色体上有2.86个;代换片段长度最小为0.55 cM,最大为33.00 cM,平均长度为12.36 cM,总覆盖长度为247.20 cM,覆盖率为16.21%.利用BC2F2群体对14个产量相关性状进行QTL定位,共检测到控制8个性状的20个QTL.对性状表型值起增效作用的有11个,占总检出数的55%.控制产量相关性状的QTL存在簇状分布现象,这与表型相关分析结果相符合.作者:程桂平冯九焕梁国华刘向东李金泉 CHENG Gui-ping FENG Jiu-huan LIANG Guo-hua LIU Xiang-dong LI Jin-quan 作者单位:程桂平,冯九焕,刘向东,李金泉,CHENG Gui-ping,FENG Jiu-huan,LIU Xiang-dong,LI Jin-quan(华南农业大学,广东省植物分子育种重点实验室,广东,广州,510642)梁国华,LIANG Guo-hua(扬州大学,植物功能基因组学教育部重点实验室,江苏,扬州,225009)刊名:中国水稻科学 ISTIC PKU英文刊名:CHINESE JOURNAL OF RICE SCIENCE 年,卷(期):2006 20(5) 分类号:Q943 S511.032 关键词:栽培稻普通野生稻 BC2F2群体产量相关性状数量性状座位。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
关 键 词 :水 稻 ;籼 粳 杂 种 ;数 量 性 状 基 因 座 ;花 粉 育 性 ;小 穗 育 性
中 图 分 类 号 :Q943.2;S511.032
文 献 标 识 码 :A
文 章 编 号 :1001-7216(2011)06-0594-05
Байду номын сангаас
水稻籼粳亚种间杂 种 优 势 可 达 30% ~50%,利 用其杂种 优 势 已 成 为 当 前 水 稻 遗 传 育 种 的 研 究 重 点 。 [1] 但籼粳交后代存在结实率低、籽粒不 饱 满、生 育 期 偏 长 、植 株 偏 高 和 米 质 分 离 等 缺 点 ,成 为 籼 粳 亚 种间杂 种 优 势 利 用 主 要 技 术 难 题,其 中 育 性 是 关 键[2]。1986 年,Ikehashi等 首 [3] 次 报 道 了 广 亲 和 基 因S-5n。以该理论为 依 据,水 稻 育 种 家 在 对 水 稻 不 同生态型和品种间 的 杂 种 不 育 性 的 研 究 中,相 继 发 现了 S-7、S-8、S-9、S-15、S-16、S-17 和S-p(t)等 广 亲 和 基 因 ,并 拟 通 过 上 述 基 因 的 重 组 聚 合 ,创 制 广 亲 和育种 材 料,以 克 服 籼 粳 杂 种 的 育 性 障 碍。Chen 等 成 [4] 功克隆了水稻广亲和基因S-5n。杨杰等 对 [5] S-5n的 功 能 标 记 进 行 了 开 发 与 应 用。 与 此 不 同,张 桂权等 在 [6] 对水 稻 籼 粳 杂 种 花 粉 育 性 研 究 基 础 上, 提出了“特异亲和 基 因 理 论”,认 为 亚 种 间 杂 种 花 粉 不育受6个特异亲 和 基 因 S-a、S-b、S-c、S-d、S-e 和
应)QTL(qPF3、qPF5 和qPF6)和4个 控 制 小 穗 育 性 的 主 效 应 QTL(qSF3、qSF5、qSF6 和qSF8),其 中qPF5 与qSF5、qPF6 与qSF6 均 为 同 一 个 QTL,为 重 要 的 育 性 QTL。另 外 ,检 测 到3对 花 粉 育 性 和4对 小 穗 育 性 QTL 间 互 作 ,说 明 上 位 性 效 应 对 籼粳杂交后代的育性具有重要作用。
花粉育性和小穗育性的百分数值经反正弦转换 后 用 于 统 计 分 析。 利 用 MapMaker/Exp3.0 软 件 构 [20] 建 分 子 标 记 遗 传 图 谱。 利 用 QTLNetwork 2.0 软 件[21]进 行 QTL 定 位,总 体 显 著 水 平 设 为 5%,通 过 置 换 测 验(permutationtests)估 计 相 应 的 显著阈值,当 F 统 计 量 峰 值 超 过 阈 值 时,即 认 为 峰 值 对 应 的 染 色 体 位 置 存 在 1 个 QTL。 采 用 McCouch等 的 [22] 方法对 QTL 命名。
595
改良,这是亚种间杂 种 优 势 利 用 的 一 项 重 要 基 础 工 作。而且,尽管已经 发 现 了 许 多 影 响 籼 粳 杂 种 育 性 的 基 因 ,仍 有 待 发 掘 未 知 的 有 关 基 因 。
为了 选 育 亲 籼 偏 粳 型 不 育 系 种 质,我 们 以 一 些 籼稻保持系和粳稻保持系为亲本进行了杂交。本研 究以其中一个杂交组合的 F2群 体 为 材 料,在 3 种 环 境条 件 下 对 籼 粳 杂 种 育 性 进 行 QTL 分 析,旨 在 揭 示其杂种不育的遗 传 基 础,为 克 服 该 杂 交 组 合 后 代 的 育 性 障 碍 ,选 育 亲 籼 偏 粳 型 不 育 系 提 供 理 论 依 据 。
S-f 的控制,其中S-a、S-b 和S-c 已被精细定位或克 隆 。 [7-9] 近年来,一 些 新 的 亲 和 基 因 S26、S27、S28、 S29、S30、S31、S32、S33 和S34 等被陆续发现 。 [10-17]
由于 存 在 籼 粳 杂 种 不 育 的 现 象,为 了 利 用 籼 粳 亚种间杂种优势,必 须 选 育 亲 籼 偏 粳 型 或 亲 粳 偏 籼 型的杂交稻亲本。这为利用分子标记辅助育种技术 选育此类亲本,解决 籼 粳 杂 种 不 育 提 供 了 重 要 的 理 论 依 据 。 然 而 ,这 些 研 究 结 果 也 说 明 ,控 制 籼 粳 杂 种 育性的遗传基础 十 分 复 杂。 因 此,不 同 的 籼 粳 交 组 合所涉及的杂种育性基 因 可 能 存 在 很 大 的 差 异 。 [18] 在 实 际 育 种 中 ,必 须 针 对 具 体 的 籼 粳 交 组 合 ,分 析 其 育性遗 传 基 础,才 能 有 效 地 进 行 育 种 亲 本 的 遗 传
如 下 :在 成 熟 期 每 株 取 5 穗 考 查 结 实 率 ,取 平 均 值 。 1.3 分子标记检测
采用 SDS方法提 取 亲 本 及 上 述 176 个 F2单 株 的 DNA。SSR 标记引 物 由 上 海 生 工 生 物 工 程 技 术 服务有限公司合成。依据 McCouch等 的 [19] SSR 标 记连锁图,筛选在亲 本 间 具 多 态 性 且 覆 盖 水 稻 基 因 组的SSR 引物,检测各 F2单株的基因型。将亲本辽 91B 的带 型 记 为 1,珍 汕 97B 的 带 型 记 为 2,杂 合 带 型 记 为 3,缺 失 数 据 记 为 0。 1.4 数据分析
(1 福建农林大学 作物科学学院,福建 福州 350002;2福建农林大学 生命科学学院,福建 福 州 350002;* 通 讯 联 系 人,E-mail:wrwu2005@ )
QTL AnalysisforFertilityofIndica-JaponicaHybridsinRice
indica-japonicahybrids. Keywords:rice;indica-japonicahybrid;quantitativetraitlocus;pollenfertility;spikeletfertility
林荔辉,王爱梅,周元昌,等.水稻籼粳杂种育性的 QTL 分析.中国水稻科学,2011,25(6):594-598.
LIN Lihui,WANG Aimei,ZHOU Yuanchang,etal.QTLanalysisforfertilityofindica-japonicahybridsinrice.ChinJRice Sci,2011,25(6):594-598. Abstract:AnF2populationconsistingof176individualswasdevelopedfromacrossbetweenanindicamaintainerlineZhenshan
摘 要 :以 籼 稻 保 持 系 珍 汕97B 和 粳 稻 保 持 系 辽91B 为 亲 本 ,构 建 了 一 个 包 含176个 单 株 的 F2群 体 及 123 个 SSR 标 记 的 遗 传 图 谱 ,在3种 环 境 下 联 合 检 测 控 制 花 粉 育 性 和 小 穗 育 性 的 QTL。共 检 测 到3个 控 制 花 粉 育 性 的 主 效 应(加 性 效 应 和 显 性 效
1 材料与方法
1.1 水稻材料 供 试 材 料 包 括 籼 稻 保 持 系 珍 汕 97B(母 本 )和 粳
稻保持系辽91B(父本)及其 F1和 F2。 1.2 材料种植和性状调查
2007年早季在福建福州 种 植 亲 本、F1、F2群 体, 其中亲本与 F1 各 种 植 49 株,F2 群 体 种 植 2000 株。 6月15日开始抽穗后,对亲本和 F1的所有植株进行 花粉育性和小穗育性(结实率)调查(E1),同 时 在 F2 群体中随机挑取 176 个 单 株,逐 株 调 查 花 粉 育 性 和 小 穗 育 性 。 随 后 ,逐 株 割 蔸 离 蘖 再 生 ,对 再 生 植 株 进 行了两次性状调查。第1次:8 月 20 日 调 查 花 粉 育 性 ,9 月15 日 调 查 小 穗 育 性 (E2);第2 次 :9 月15 日 调查花粉育性,10 月 11 日 调 查 小 穗 育 性 (E3)(注: 虽然 F2 单 株 的 抽 穗 期 存 在 变 异,但 由 于 割 蔸 离 蘖 后 ,各 株 再 生 的 分 蘖 穗 表 现 为 连 续 抽 穗 ,使 得 各 株 的 抽穗时间存在重叠,因而对 F2群 体 的 花 粉 育 性 和 小 穗 育 性 的 取 样 调 查 皆 可 分 别 在 同 一 天 完 成 )。 显 然 , 3次性状调查可视为3个环境下的试验。花粉育性 观 察 具 体 方 法 如 下 :在 抽 穗 期 ,于 8:00 左 右 ,每 株 取 3穗,每 穗 取 当 日 开 花 的 颖 花 5 朵,混 合 制 片,然 后 用 1%I2-KI染 色 法 镜 检。 花 粉 育 性 用 可 育 花 粉 的 百分率表示,取3 穗 平 均 值。 小 穗 育 性 的 调 查 方 法
97Bandajaponica maintainerlineLiao91Bandagenetic mapcontaining123SSR markers wasconstructedbasedonthe population.Pollenfertilityandspikeletfertility wereinvestigatedinthreeseasons.Three main-effect (additiveeffectand dominanteffect)quantitativetraitloci(QTLs)forpollenfertility (qPF3,qPF5 andqPF6)andfourmain-effectQTLsfor spikeletfertility (qSF3,qSF5,qSF6 andqSF8)weredetected.Amongthem,qPF6 andqSF6,andqPF5 andqSF5, importantQTLsforricefertility,weremappedonthesamelocations.Inaddition,threepairsofQTLinteractionforpollen fertilityandfourpairsforspikeletfertilityweredetected,suggestingthatepistaticeffectsplayimportantrolesinthefertilityof