2010-膜离子通道总复习汇总
2010膜离子通道考试题

2010生物物理学考试题(3小时)1.名词解释:Desensitization:(2分)Rundown:(2分)Depolarizationr:(2分)Repolarization:(2分)HV A和LV A:(4分)2. 膜片箝放大器的电压箝工作模式有;;和。
(4分)3. 假定阻断剂TEA与K V通道的失活有竞争,当电流减少至原来的五分之一时,失活时间常数应是原来的倍。
(2分)4. 离子通道GV曲线的斜率越表明它对电压的依赖性越强,也意味着其电压敏感器(S4)上的越多。
(4分)5. 影响单通道电导的主要因素是。
(4 分)6. 阻断剂导致通道reopen的原因是:;阻断剂导致电压依赖性阻断的原因是:。
(6 分) 7.在细胞内外溶液浓度分别为[K+]i和[Na+]o条件下,某通道的反转电位为V r,利用两种离子GHK方程,求出该离子通道对上述离子选择性的比率P Na/P K。
(6分)8. 已知浓度为[CTX]蝎毒素阻断BK通道时间常数为τon而回复时间常数为τoff,给出CTX阻断BK通道的结合常数K d的表达式。
(4分)9.为什麽IP3会导致内钙的双相释放?(8 分)10.钾通道过滤器的氨基酸序列特征序列是什麽?(6 分)11.分别画出记录下面曲线所采用的指令电压(Protocol):1)激活(IV)曲线;2)稳态失活曲线;3)恢复曲线。
(6 分)12. 钙离子被称为第二信使是由于它在细胞中的许多重要功能,尽可能多地给出钙离子在细胞中的功能?(10分)13. 假定钾通道遵从二项分布的规律可结合0-4个β亚基,并且α亚基与β亚基的结合几率为p,问结合0,1,2,3和4个β亚基的钾通道在细胞中所占的百分比各为多少?(10 分)k114.假定通道动力学特性可用模型O←--→I来描述。
这里,O和I分别为开放态和失活态;k-1k1为向前速率常数,k-1为向后速率常数。
求稳态时开放态的几率Po?(10 分)15.当激动剂浓度为C时,某受体通道的单通道电流为i(V)、开放几率为P O(V)并已知其激动剂的结合常数为K d。
离子通道药理学

(二)钙通道
钙通道(calcium channels)在正常情况下为细胞外 Ca2+([Ca2+]o)内流的离子通道。它存在于机体各种 组织细胞,是调节细胞内Ca2+([Ca2+]i)浓度的主要 途径。 1.电压门控钙通道(voltage-gated Ca2+ channels):目 前已克隆出L、N、T、P、Q和R 6种亚型的电压依赖性 钙通道。
2.钙依赖性钾通道(Ca2+-dependent K+ channels)
此类钾通道为KCa,其电流为Ik(Ca)。ICa是 一类具有电压和Ca2+依赖性的钾通道。去极化和 提高[Ca2+]i浓度均可激活而使其开放,K+外流使 膜复极化或超极化。分布于血管平滑肌,直接参 与血管张力的调节,具有较大的生理意义。该通 道开放时,K+外流使膜复极化或超极化,同时引 起血管扩张。因此当血管平滑肌细胞去极化和 Ca2+进入细胞时, KCa将起到负反馈调节作用。
2.受体调控性钙通道(receptor-operated Ca2+ channels)
这类通道存在于细胞器如肌质网(sarcoplasmic reticulum, SR)和内质网(endoplasmic reticulum, ER) 膜上,是储钙释放进入胞浆的途径。由于三磷酸肌醇 (inositol triphosphate, IP3)或Ca2+等第二信使激活细 胞器上相应受体而引起通道开放,故称为细胞内受体 门控离子通道。当细胞膜去极化时,电压门控钙通道 开放,Ca2+内流使细胞内Ca2+突然增加而触发Ca2+释 放,从而引起细胞兴奋-收缩耦联等生理活动,这一过 程称为Ca2+诱发Ca2+释放。
离子通道

PDF 文件使用 "pdfFactory Pro" 试用版本创建
3.功能:
L型:兴奋-收缩耦联; T型:对电信号的整合具有重要意义; N型:调节突触前神经递质的释放。
N型钙通道功能
PDF 文件使用 "pdfFactory Pro" 试用版本创建
PDF 文件使用 "pdfFactory Pro" 试用版本创建
4. 非门控和门控离子通道 非门控:总开放
门控:开放或关闭 依据环境因素
PDF 文件使用 "pdfFactory Pro" 试用版本创建
非门控离子通道 —静息膜电位
u
u
PDF 文件使用 "pdfFactory Pro" 试用版本创建
2. 类型:
电压门控钾通道
Ik 型
延迟整流
瞬时钾电流
钙激活
内向整流
介导 M电流
TEA、4AP、Ba2+
TEA
4-AP
TEA:四钇铵
Cs
4-AP: 4-氨基吡啶
PDF 文件使用 "pdfFactory Pro" 试用版本创建
的关系。
PDF 文件使用 "pdfFactory Pro" 试用版本创建
三、离子通道型受体
离子通道型受体在各类受体中反应最为迅 速,受体本身即为离子通道,当配体与受体结 合时,离子通道开放,细胞膜对离子的选择通 透性增加,引起细胞膜兴奋性的快速改变。
PDF 文件使用 "pdfFactory Pro" 试用版本创建
(一)受体分类与特性
受体分类 亚基 数目 5个 亚基跨 膜特点 4TM 亚基N和C末 端定位 N和C末端均 在胞外 配体 来源 举例 n AChR、 胞外 GABAAR、Gly R 、 5-HT3R 胞外 胞内 胞外 NDMAR、 AMPAR、KAR CNG、HCN、 IP3R、RyR ASICs、P2x
离子通道与受体主讲

P2Y受体分子结构及特性: P2Y受体由308-377个氨基酸组成的蛋 白质。 与其它G蛋白耦联受体一样具有7个跨膜 结构域。有较长的胞外N末端,和胞内 C末端。
P2Y受体 胞内转导机制: 大多数P2Y受体经G蛋白耦联,激活PLC, 导致IP3形成和胞内Ca2+动员。少数 P2Y受体经G蛋白耦联,激活AC。
1、膜受体 水溶性物质与膜受体结合 (1)环状受体 即配体门控离子通道 神经递质 (2)七个跨膜α螺旋受体 蛇型受体 其胞浆面第三个环能与鸟苷酸结合 蛋白相偶联
鸟苷酸结合蛋白(简称G蛋白)是一类和 GDP或GTP结合、位于细胞膜胞浆面的 外周蛋白 由三个亚基组成α、β、γ 两种构象 活化型(GDP)、非活化型(GTP) 效应酶 腺苷酸环化酶(AC) 磷脂酶C(PLC)
2、膜片钳构型(configuration) 膜片钳是一种能测定单离子通道(single ion channels)电生理新研究技术 (1)细胞贴附式(cell attached mode): 这种构型适用于在完整的细胞膜上 ( 全细 胞)测定单通道电流,用于研究某些特殊 物质,如神经递质(加入到细胞浸溶液中), 引起通道的改变(调制或调节作用)。
(二)胞内受体 胞内受体多为反式作用因子 类固醇激素、甲状腺素、维甲酸 1. 结构:高度可变区 N端 转录激活 作用 2. DNA结合区 富含半胱氨酸 3. 激素结合区 C端 配体结合区 激活转录
(三)受体作用的特点 1. 结构高度专一性 2. 高度亲和力 3. 可饱和性 4. 可逆性 5. 特定的作用模式 6. 具有放大效应
(4)膜内向外膜片钳 (inside-out patch clamp): 这是膜片与细胞脱离,研究单通道的第二种 模式。通常用于特定细胞研究第二信使参与 通道活性的调制作用,将所要研究的物质加 入浸浴液,直接作用于细胞浆膜的内侧面。
离子通道概论

3、临床应用 (1)高血压 (2)心绞痛和心肌梗死 (3)心肌保护作用 (4)充血性心衰 (5)其他
钾通道
钾通道—选择性允许钾离子跨膜通过的离子通 道,在调节细胞的膜电位和兴奋性以及平滑肌 舒缩活动中起重要作用。 钾通道分类:电压依赖性钾通道(延迟整流钾 通道、瞬时外向钾通道、起搏电流)、钙依赖 性钾通道、内向整流钾通道(内向整流钾通道、 ATP敏感的钾通道、乙酰胆碱激活的钾通道) ATP敏感的钾通道-代谢性调节钾离子外流通道, 分布广泛,受细胞内ATP/ADP的比率、镁离子 和G蛋白的调控,有特异性的开放剂和组滞剂。
钠通道
钠通道—选择性允许钠离子跨膜通过的离子通 道,均为电压门控离子通道,主要功能是维持 细胞膜兴奋性及其传导。 钠通道分类:神经类钠通道、骨骼肌类钠通道、 心肌类钠通道(快钠通道、慢钠通道) 钠通道特征: 1、电压依赖性 2、激活和失活速度快 3、有特异性激活剂和阻滞剂
钙通道
钙通道—在正常情况下为细胞外钙离子内流的离 子通道,是调节细胞内钙离子浓度的主要途径。 钙通道分类:电压门控离子通道(L、N、T、P、 Q、R亚型)、受体调控性钙通道(钙诱发钙释放) 电压门控钙通道特征: 1、电压依赖性 2、激活速度缓慢,失活速度慢于激活 3、对离子的选择性较低
氯通道
氯通道生理作用:在兴奋细胞为稳定膜 电位和抑制动作电位的产生;在非兴奋 性细胞维持其负的膜电位,为膜外钙进 入细胞内提供驱动力;氯通道还在调节 细胞体积、维持细胞的内环境稳定中起 重要作用。 氯通道分类:电压敏感氯通道、囊性纤 维跨膜电导调节体、GABA受体氯通道。
三、离子通道的分子结构及门控机制 1、电压门控离子通道的分子结构 2、内向整流钾通道的分子结构 3、电压门控离子通道的动力学过程: (1)静息态 (2)开放状态 (3)失活状态 (4)复活
2011膜离子通道考试题

2011离子通道和细胞电生理试题(3小时)1.名词解释:(14分)开放态寿命:去敏感:可激活细胞:门电流:内向整流:去极化:超极化2.在电生理中,我们规定膜电势等于膜内电势减去膜外电势;电流方向的规定是以膜内流向膜外为正;欧姆定律的形式是。
(8分)3.离子通道从O(开放态)到C(关闭态)的过程叫去激活;从O到I(失活态)的过程叫失活;从C到O的过程叫激活;从I到C的过程叫恢复。
(8分)4.假定细胞外液中,[Na+]o=150 mM和[K+]o=5 mM;内液中有[Na+]i=5 mM而[K+]i=150mM。
已知该离子通道的反转电位在-75.9 mV, RT/F=25.3 mV, 求钠和钾离子对该通道通透性的比值P K/P Na(e-3=0.05),并问这是何种离子通道。
(10分)5.以Kv通道为例,说明HH模型的基本假设。
(10分)6.画图说明以下两实验各自所需的指令电压:激活和稳态失活。
(10分)7.从“目标,酶,荷尔蒙,受体,G蛋白,酶,第二信使”中,正确排出信号转导通路次序。
(5分)8.什麽叫SOC通道(定义)?说出你所知有关SOC通道开放的各种假说。
(10 分)9.α[T]9. 药物动力学模型O←---→B用来描写药物阻断效应。
这里,O和B分别为开放态和阻断β态;α为向前结合速率常数,β为向后解离速率常数,[T]为药物浓度。
若测得该药物的解离平衡常数K d=2 nM和洗脱时间常数τrec=200 s(秒)。
求:(1)α和β(单位:s-1或M-1s-1)(5分)(2)通道电流被阻断3/4时的药物浓度[T] (单位:nM)。
(5分)10.证明在平衡态时,下面动力学模型的顺时针方向的速率常数之积等于其反时针方向的速率常数之积:k1k2k3k411.全细胞记录到的某钾通道的总电流为2500 pA, 已知在40 mV时,该钾通道单通道振幅是25 pA和开放几率0.5,求该细胞中的钾通道的数目?若该钾通道在细胞中的反转电位是-60 mV,求40 mV时的通道电导(假定电导是线性的)?(10分)。
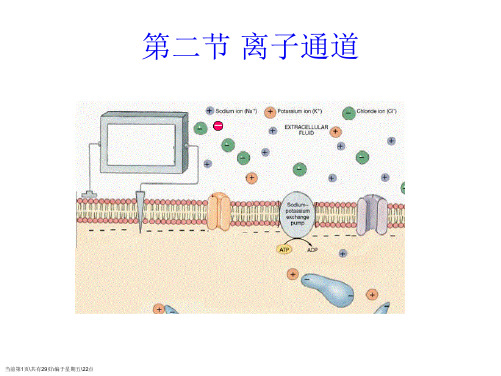
第二节离子通道
2、作用于K+通道的药物
(1)钾通道抑制剂:钾通道抑制剂的种类很多,有无机离子(如Cs+、Ba2+ 等)、有机化合物(如TEA和4-AP等),多种毒素(如蝎毒,蛇毒、蜂毒等), 以及目前临床治疗用药物。无机离子及多数合成的经典钾通道阻断剂多作 为研究工具药使用,它们对钾通道亚型的选择性较差。毒素则对通道亚型 的选择性相对较高,如蛇毒可选择性阻断瞬时外向钾通道;蝎毒可阻断大 电导钙依赖钾通道特异性较高。这些毒素已作为研究钾通道的生化探针而 广泛使用。
1、钾通道的分类及其特性
(1))延迟整流钾通道(Kv):也称IK通道,是第一个被发现的K+通道
,广泛存在于各种组织中。由于电压阶跃引起其电导的改变有一段明
显的延迟而得名,实际上,后来发现的许多其他类型的K+通道也有这 种延迟整流现象。目前所说的Kv主要是指性质与枪乌贼神经轴突 Kv相似的K+通道,是细胞去极化时激活的外向钾电流。主要特 点是电流的激活是电压和时间依赖的,基本上无自动失活。其 主要功能是参与细胞的复极化过程。这类通道对TEA敏感。
Ⅰ类抗心律失常药对钠通道的阻断作用依赖于心率,即当心率快时
阻断作用强,而心率慢时作用不明显或看不出其阻断作用,称之为频率依 赖性。药物与通道间的相互作用及其频率依鞍性阻断与药物对钠通道作用 的状态依赖性有关。处于开放或失活关闭状态的通道对药物亲和力高,而 在静息关闭时通道不与药物结合,或药物只在通道开放时才能进入到其结 合位点。所以高频率电脉冲(如快心率)时通道更多处于开放状态而易被药 物阻断;被阻断的通道在静息时复活减慢,更长时间地处于失活关闭状态 ,更使药物作用加强。因此,药物对钠通道的阻断作用取决于通道进入开 放(使用)状态的频率,故又称为开放状态阻断或使用依赖性阻断。不仅钠 通道阻断剂,钙通道阻断剂如维拉帕米等也具有这一特性。
生物膜中的离子通道与传输机制

生物膜中的离子通道与传输机制生物膜是细胞外与细胞内环境之间的物理隔离屏障,对细胞内外物质交换起着重要的作用。
生物膜由磷脂双层以及膜蛋白等多种分子构成,其中离子通道是生物膜中的重要成分之一。
离子通道可以让离子定向地穿越膜,保证了细胞内外的离子浓度差,维持了正常的细胞生理活动。
本文将从离子通道的发现历程、分类及传输机制三个方面,介绍生物膜中的离子通道。
一、离子通道的发现历程早在20世纪50年代,科学家就已经意识到细胞膜是具有选择通透性的物质屏障,但对离子的穿透机制却不甚了解。
1960年代初期,Hodgkin和Huxley对神经元膜电位的研究首次提出了离子通道的概念。
他们通过电生理技术,观察到了离子通道的存在,并系统地研究了通过离子通道的钠离子、钾离子等离子体系的穿透机制。
这项工作为细胞膜电位的传递机制提供了新的认识。
随后,生物学家们不断地发掘各种类型的离子通道。
通过逐一研究它们的通透性、特异性和调节机制等特征,科学家们已经发现了许多种不同类型的离子通道。
二、离子通道的分类根据离子通道的特异性和调控机制,可以将其分为多种类型。
其中,最常见的包括压力调节离子通道、电压门控离子通道、化学门控离子通道、内源锁定离子通道和非门控离子通道等。
这些离子通道有分布在细胞膜内部的学名叫做内向型离子通道;有限制细胞外离子到达细胞内的外向型离子通道;还有分布在细胞内膜的可协同作用的离子通道。
这些不同类型的离子通道的作用可相互协调,维持正常生理活动。
三、离子通道的传输机制离子通道的传输机制是指不同类型、不同调节机制的离子通道穿越细胞膜的方式。
离子通道的传输机制和其类型密切相关。
压力调节离子通道是一种通过压力甚至物理变化进行开关操作的离子通道。
细菌及植物细胞膜上的压力调节离子通道就是通过物理变化感受细胞膜的机械变化,并通过特定的门控机制来让离子定向地穿透膜。
相比之下,电压门控离子通道和化学门控离子通道是通过细胞内外环境的电势或化学信号作为触发信号,在离子通道上开启或关闭,从而把离子从一侧传递到另一侧。
神经元细胞膜间的离子通道机制

神经元细胞膜间的离子通道机制神经元是神经系统中最基本的单位,其主要功能是传递神经信号。
神经传导需要依托离子通道的存在,这些通道控制着神经元膜上的离子通量,决定了神经肌肉系统的信号传递过程。
随着技术的不断发展,对于神经元膜上离子通道机制的研究也在不断深入。
1. 离子通道类型目前,已经发现的离子通道可分为四大类型:钾离子通道、钠离子通道、钙离子通道和氯离子通道,其中前两者常见于神经元膜上,后者则主要存在于肌细胞膜上。
2. 离子通道作用在神经元膜上,离子通道通过控制离子通量来实现以下作用:(1)控制神经元的静息电位:钾离子通道运转较为常规,其打开后可让细胞内外的离子快速平衡,从而维持静息电位。
在人体正常生理状态下,钠离子通道处于关闭状态,细胞内外离子分布不平衡,导致静息电位的建立。
(2)控制神经元的动作电位:当神经元膜电位达到一定阈值时,钠离子通道将发生短暂打开,大量钠离子快速进入细胞内部,导致膜内电位迅速升高,进而产生“动作电位”。
随后,钾离子通道会迅速打开,让大量钾离子迅速离开细胞,从而使膜电位逐渐恢复到静息电位。
3. 离子通道的可塑性神经元膜上的离子通道还具有一定的可塑性,即它们可以根据不同情境下的需求而调整其数量和运转机制。
不同离子通道的调控机制也不尽相同,但总体来说,调控机制可以归纳为以下几个方面:(1)神经递质的影响:某些神经递质(如去甲肾上腺素)可通过作用于神经元膜上的受体来启动一系列离子通道的开/关机制。
(2)调节表面通量密度:神经元膜上的离子通道显示出极高的可塑性,可以通过调节其表面通量密度来影响神经元膜的电生理特性。
(3)磷酸化和脱磷酸化:磷酸化和脱磷酸化是一种重要的调控信号转导机制,它可以通过调控离子通道蛋白的磷酸化水平来直接影响其运转状态。
总之,神经元细胞膜间的离子通道机制是神经传导过程中的核心环节,了解其机制对于神经科学和神经药理学研究具有重要意义。
随着技术的不断进步,我们相信将来会有更多深度研究能够揭示这一过程的更多细节。
离子通道的基础知识及其技术

配 体 门 控 性 , 又称化学门控性(chemical gated)离子通道,由递质与通道蛋白质 ligand gated 受体分子上的结合位点结合而开启,以递质受体命名,如乙酰胆碱
受体通道、谷氨酸受体通道、门冬氨酸受体通道等.非选择性阳离 子通道(non-selective cation channels)系由配体作用于相应受 体而开放,同时允许Na+、Ca2+ 或K+ 通过,属于该类.
做出了开创性研究。他们基于电压钳技术,提出 并验证了所谓的Hodgkin—Huxley方程,数学模拟 出和真实状况相符合的神经冲动的传导,由此建
立了细胞动作电位的钠离子学说。离子通道的近
代观念也由此产生。
4.离子通道研究技术
1. 微电极电压钳-电流钳技术
2. 传统膜片钳技术
Kenneth Cole (1900-1984)
E.Neher和B.Sakmann创立了膜片钳技术,使电生理研究空前活跃,并作为重要的里程碑,标志着
电生理分子时代的到来。
膜片钳技术、生物大分子技术以及生物光学技术的结合,使人们对生物电现象的认识水平在分子水 平取得了重大的突破和丰富。
5. 传统膜片钳设备的组成
电学部分
光学部分:
➢ 信号放大器
➢ 荧光显微镜
✓ 高灵敏度:pA级、单通道水平 ✓ 适用范围广:不受样本种类限制
✓ 操作自动化,简单培训即可使用 ✓ 高效率、高通量 ✓ 实现某些传统膜片钳不具有的功能,
例如全细胞模式内液灌流。
操作过程复杂,对实验者要求高
实验数据量低,难以应用于药物筛 选
细胞膜中的离子通道结构和功能研究

细胞膜中的离子通道结构和功能研究细胞膜是细胞的外层保护层,不仅限制了物质进出细胞,还能感知、传递信号。
为了效率和准确性,细胞膜上还存在各种类型的通道分子,负责特定物质或离子的转运。
其中,离子通道是最为重要的一类。
一、离子通道种类细胞膜中的离子通道可以分为多种类型,按照内部结构、离子种类和激活方式等分类。
其中,最常见的有以下几种:1. 钾通道:透过钾离子的离子通道。
可分为电压门控式和内、外源钙激活的两种类型。
在神经递质的传递、心脏节律控制等方面都起着重要的作用。
2. 钙通道:通过钙离子的离子通道。
包括电压门控式、配体门控式等多种类型。
功能非常广泛,参与了神经递质释放、细胞过程调控、心肌收缩等方面。
3. 钠通道:透过钠离子的离子通道。
主要特点是快速响应和活化,通常是电压门控式。
肌细胞、神经元和心肌细胞等组织内都非常重要。
二、离子通道的结构离子通道由多种蛋白质组成,主要包括离子选择性通道蛋白和辅助蛋白等组分。
其中,离子选择性通道蛋白是最主要的成分,决定了通道的选择性和特定的通透性。
一般来说,离子选择性通道蛋白分为两个主要区域:跨膜区域和细胞内外区域。
跨膜区域是最主要的结构部位,是通道内离子通透的主要场所。
细胞内外区域则主要负责信号接受、传递、锚定等作用。
值得注意的是,离子通道的结构是非常灵活多变的,除了基本的通道结构外,延伸出的结构域、融合的功能模块等都起着不容忽视的作用。
三、离子通道的功能离子通道在细胞功能中扮演着非常重要的角色,他们不仅仅是物质通透的通道,更是信号传递的重要分子。
1. 物质转运:离子通道的最基本功能是实现对离子化物质的转运。
通过选择性通透离子的种类和数量,细胞控制物质进出的速率和流量。
2. 信号传导:在许多情况下,离子通道的开启和关闭会引发电位变化或其他信号转导。
这些信号既可以是细胞内外的环境改变,也可以是其他信号调节分子的作用。
这些信号再促进通道开放或关闭,形成不同的电活动和细胞响应。
细胞膜离子通道的结构与功能分析

细胞膜离子通道的结构与功能分析细胞膜离子通道是细胞膜上具有选择性通透特定离子的蛋白质通道,它是控制细胞内离子稳态和神经、心肌等细胞兴奋性和肌肉收缩等生理功能的主要机制。
近年来,离子通道的结构和调控机制的深入研究,为了解许多神经系统疾病、代谢性疾病等提供了重要的信息。
下面将从结构和功能两个方面来分析细胞膜离子通道。
结构分析1. 离子通道蛋白的基本组成离子通道蛋白的基本组成由四个同源亚基组成的单体或多聚物,每个亚基包括约300-400个氨基酸残基,其中包含6个跨越膜的α螺旋,它们形成一个α螺旋通道。
离子通道蛋白不仅在氨基酸序列上具有相似性,而且在二级和三级结构上也非常相似。
在细胞膜上的位置不同,离子通道蛋白可分为一类、二类、三类和四类。
2. 离子通道蛋白的跨膜结构离子通道蛋白的跨越膜结构具有各种特殊的性质,如跨膜α螺旋的长度、形状和分布、跨膜α螺旋之间的构象相互依存关系以及跨膜氨基酸的沿膜面分布等。
这些结构特征决定了离子通道蛋白的对特定离子选择通透性。
3. 离子通道蛋白的分子结构离子通道蛋白的分子结构在晶体结构和电镜结构上得到了深入的研究。
以电镜技术为代表的单粒子分析技术在研究离子通道蛋白的分子结构方面发挥了重要作用。
通过单粒子分析,我们可以了解离子通道蛋白的三维结构、不同构象和亚基构成等信息。
功能分析1. 离子通道在调节生理功能中的作用离子通道在调节生理功能中发挥了重要作用。
它们可以控制神经元、心肌细胞和平滑肌细胞等的兴奋性和肌肉收缩,还可以调节胰岛素分泌和肾脏功能,参与酸碱平衡、水盐平衡、细胞体积调节等生理功能。
2. 离子通道的疾病相关性离子通道的异常功能可能导致一系列疾病。
某些遗传病致病基因编码的离子通道蛋白的突变可以导致失调的离子通道功能,从而导致一系列遗传性神经疾病、心脏病和代谢性疾病等。
在神经疾病方面,离子通道的疾病包括癫痫、晕厥、麻痹性疾病等。
3. 离子通道药物治疗的研究离子通道是很多药物作用靶点之一。
生物膜上离子通道的电生理学特性

生物膜上离子通道的电生理学特性生物膜是细胞内外的一个重要屏障,对维持细胞内外环境的稳定性和调节细胞外和内物质的交换具有重要意义。
膜上离子通道作为维持细胞内外环境平衡的主要机制之一,具有重要的生理和病理功能。
离子通道可以通过信号转导、维持细胞电位稳定性、维持细胞内外离子浓度平衡以及调节神经肌肉兴奋等多种生理功能,在多种疾病中的功能异常会导致疾病的发生和发展。
因此,分子生物学、细胞生物学和生物物理学等多个学科对离子通道的研究逐渐深入。
离子通道的特性包括电学本性、氧化还原敏感性、细胞信号转导的作用以及参与疾病等多方面的特性。
其中,电生理学特性是离子通道研究中最基本和重要的特性之一。
下面,我们就来探讨生物膜上离子通道的电生理学特性。
1. 感受器电压门控机制生物膜上离子通道的电生理学最突出的特点之一是感受器门控机制。
这种机制是指离子通道的开放状态受到细胞膜内外电场分布的影响。
因此,这种开关机制是离子通道的一个特殊功能,可被用于细胞膜的神经和肌肉细胞的调节。
感受器电压门控通道是目前发现的最多的离子通道类型。
这种类型的通道有两个感受器作为门控,分别处于通道的内部和外部,内外两者间相距不超过0.33纳米。
当神经脉冲到达细胞膜时,产生的电场可以使得感受器发生构象变化,从而改变离子通道的开放状态。
这样的机制使得离子通道能快速响应外界的刺激,起到紧急反应的作用。
2. 离子选择性离子选性是离子通道的另一个重要电生理学特性,是指离子通道在不同类型的离子之间有不同的选择性。
离子通道是区分离子种类的主要机制,其精准度对于细胞内外环境成分的调节具有重要意义。
离子通道的选择性取决于其结构和特定氨基酸残基的构成。
那些在通道内侧和外侧共同积累的氨基酸残基为其具有特定的选择性。
所以大多数离子通道均能分辨不同类型的离子,如钠、钾、氯等。
这样的选择性机制能够维持细胞膜上离子的平衡,帮助细胞保持正常的生理功能。
3. 原位记录技术生物膜上离子通道的研究需要离子通道的原位记录技术。
2010-膜离子通道总复习汇总

2010总复习一.基本定义和概念:欧姆定律:I=G×(E-E rev);膜电势=膜内-膜外;电流方向规定从胞内流向胞外为正;膜等效电路;离子选择性. E rev=RT/zF*ln(P A[A]o/P B[B]i)E rev=RT/F*ln((P K[K]o+P Na[Na]o+P Cl[Cl]i)/ (P K[K]i+P Na[Na]i+P Cl[Cl]o))动作电位产生原因:Na/Ca的内流, K的外流产生动作电位1.动作电位产生于Na通透性的增加:电击后,膜电位首先从V rest增加并超过0 mV达到最大值(叫去极化)之后开始下降至V rest或更低时叫复极化或超极化。
AP:All-or-none;有阈值(去极化10-15 mV);动作电位是一种脉冲式的电信号(常见于CNS中)。
见图。
有关动作电位的名词:去极化: 极性程度的减弱;复极化;超极化;动力学名词:激活activation:电导在一个短时间延时后急剧增加(去极化过程);失活:电导在一个短时延时后急剧增加后掉到低值(去极化过程中通道关闭);去激活deactivation:电导回到通道关闭的水平;尾电流:驱动力、通道开放;门电流/门电荷:位移电流,通道上电荷在电场力的策动下运动所致。
一般在10ms内消失,电流很小,可以通过阻断所有通道电流测得。
计算门电荷数/有效电荷:利用GV曲线的Slope得到门电荷或用方差法从电流中得到通道数或从门电流中得到总电荷数;O/C=exp(-(w-z g q e E)/k B T)O/(O+C)=1/(1+exp((w-z g q e E)/k B T))) 这里z g是等效电荷数。
,k为gv曲线中参数bQ10定义:会利用已知的Q10,计算速率常数随温度的变化;生物学中定义:Q10= k(T+10o C)/k(T),这里κ代表速率常数。
Eyring 速率定律:k(T) =k(0)exp(-∆G/RT). 用近似T=273>>10.任意温度间隔∆T的系数:Q∆T=(Q10)∆T/10.其它术语:Rundown:全细胞模式下,由于胞内的小分子能量物质可以通过电极流失而导致通道电流在很短时间内减小或消失,且不可恢复,可以在电极中加入ATP, Mg,cAMP等物质消除Rundown现象;Desensitization:加入Ach的几秒钟后,宏观终板电导减小或微观Ach电流减小。
离子通道课件讲解

全细 胞钾 离子 通道 电流
单通 道记 录钙 离子 通道 电流
抛光仪
拉制仪
显微镜操作系统及可见光源
冷 光 源
膜片钳放大器、操作系统
低 温 水 浴 摇 床
材料的 好坏直 接影响 膜片钳 实验的 成败!
要根据不同的实验目的来栽培材料。 根细胞:通常根长度在2厘米左右,但突变体要 根据该基因起作用的发育阶段来确定; 保卫细胞:通常在土壤中生长4-6周,但绝不能 抽苔;叶肉细胞对光照要求较高,因为光照直 接影响叶绿体的活动。
3) 由于电压钳微电极尖端很细( 电阻大) , 会影响记录电流的能力。
膜片钳技术是用微玻管电极膜片电极或膜片吸管接触细胞膜, 以千兆 欧姆以上的阻抗使之封接, 使与电极尖开口处相接的细胞膜的小区域膜片 与其周围在电学上分隔, 在此基础上固定电位, 对此膜片上的离子通道的 离子电流(pA级)进行监测记录的方法.
( 3) 第二信使激活的离子通道, 包括由细胞内Ca2+、 IP3、G 蛋白及蛋白激酶激活的离子通道。
质膜上的离子通道
植
阴离子通道
物
细
胞
液泡膜上的离子通道
离
子
通 道
阳离子通道
其他细胞器膜上的离子通道
自从在蚕豆保卫细胞膜上发现植物离子通道以来,人们对植物细 胞乃至作为细胞器的液泡膜上离子通道的认识迅速深入,到目前为 止,已经发现植物细胞及其内膜上存在多种离子通道。
植物细胞离子通道
植物根系不仅从土壤中吸收水分,还要吸收各种矿质 元素,以维持正常的生命活动,促进植物的生长发育目 的获得高产优质。如:小麦、水稻、玉米、烟草、蔬菜 、水果等 植物吸收矿质元素首先细胞从环境中吸收,这个环境可 以是植物的生存外部环境,如土壤,也以是植物本身的 内部环境。 肥料→离子→吸附→交换→细胞 被动吸收:外液浓度>细胞浓度 吸收 主动吸收:外液浓度<细胞浓度 吸收 积累离子
离子通道概述

离子通道概述离子通道是神经、肌肉、腺体等许多组织细胞膜上的基本兴奋单元。
它们产生和传导电信号,具有重要的生理功能。
由于生物物理学和分子生物学的迅速发展,新的研究技术包括膜片钳技术、分子克隆及基因突变技术等的广泛应用,人们开始从分子水平来解释离子通道的孔道特性、动力学过程结构与功能的关系以及功能的表达和调节等。
第一节离子通道的分类离子通道必须能够开放和关闭才能实现其产生和传导电信号的生理功能。
至尽为止离子通道还没有一个系统的分类法。
1、按激活机制划分:①.电压门控性通道(Voltage-gated channel)或电压敏感性通道、电压依赖性通道、电压操作性通道。
其开、关一方面由膜电位(电压依赖性)所决定,另一方面与电位变化的时间有关(时间依赖性)。
这类通道在维持兴奋细胞的动作电位方面起重要作用。
如Na、K、Ca、Cl 通道等。
②.化学门控性通道或递质敏感性通道(Transmitter-sensitive channels)、递质依赖性通道、配体门控性通道(Ligand-gated channel)其开、关取决于与该通道相耦联的受体的状态,直接受该受体的配体的调控。
如Ach激活的K+通道,突触后膜的受体离子通道,谷氨酸受体、甘氨酸受体、Υ-氨基丁酸受体等。
③.感觉受体通道(Sensory-receptor channels)分布于精细的膜结构上或神经末梢上。
许多感觉末梢很小,故任何对代谢或细胞外介质产生的微小干扰都会很快导致膜内物质浓度的变化。
这类通道无特异阻断剂,对离子选择性很差,阳离子或阴离子均可通过。
感觉受体有两类:一类是受刺激后受体本身作为通道直接开或关。
另一类则要经过第二信使,才能使通道开或关。
某些神经递质可以影响电压门控性通道,而某些化学门控性通道也受膜电位的影响,形成离子通道的“双闸门机制”。
2、按门控的特点来划分①三门控性通道、②双门控性通道(I Na、I to、I si)、③单门控性通道(I k、I f)、④无门控性通道(I k1、I b)。
生物膜离子通道[整理版]
![生物膜离子通道[整理版]](https://img.taocdn.com/s3/m/b90e5e5af6ec4afe04a1b0717fd5360cba1a8db0.png)
生物膜离子通道生物膜离子通道示意图生物膜离子通道(ion channels of biomembrane)是各种无机离子跨膜被动运输的通路。
生物膜对无机离子的跨膜运输有被动运输(顺离子浓度梯度)和主动运输(逆离子浓度梯度)两种方式。
被动运输的通路称离子通道,主动运输的离子载体称为离子泵。
生物膜对离子的通透性与多种生命活动过程密切相关。
例如,感受器电位的发生,神经兴奋与传导和中枢神经系统的调控功能,心脏搏动,平滑肌蠕动,骨骼肌收缩,激素分泌,光合作用和氧化磷酸化过程中跨膜质子梯度的形成等。
细胞膜离子通道细胞膜上离子通道的功能,除了可以调节细胞内外的渗透压,也是维持细胞膜电位的重要分子,而神经细胞要进行讯号传导,便是靠离子的进出以造成膜电位的变化。
虽然科学家对于细胞膜上离子通道已有相当程度的了解,对于离子通道所具有的特殊选择性,也从能蛋白质的结构大略获得解释,但是一直缺乏一套完整详细的分子作用机制。
原因是,要做出膜蛋白三维结构的高解析度影像,非常不容易。
1998年,麦金农做出了链霉菌的离子通道蛋白质KcsA的高解析三维结构影像,并首度从原子层次去了解离子通道的作用方式。
KcsA离子通道中有一种“滤嘴”,能让钾离子(K+)通过,却不允许同族元素中体积更小的钠离子(Na+)通过,这令科学家百思不得其解。
但是麦金农根据KcsA的立体结构,发现离子通道中“滤嘴”边上的四个氧原子的位置,恰好跟钾离子在水溶液中的情况一样,亦即滤嘴边上的氧与水分子的氧距离相同,所以钾离子能够安然通过通道,一如在水中一样;但钠离子尺寸较小,无法顺利接上滤嘴边上的四个氧原子,因此只能留在水溶液,而无法轻易穿过通道。
而离子通道的开关会受到细胞的控制,麦金农发现,离子通道的底部有个闸门,当离子通道接收到特定的讯号,离子通道蛋白质结构便会发生改变,因此造成闸门的开关。
麦金农对于钾离子通道的结构与作用机制的研究,是生物化学、生物物理等领域的一大突破,也为神经疾病、肌肉与心脏疾病的新药物开发,指引了新的方向。
生物膜离子通道

研究方法
离子通道结构和功能的研究需综合应用各种技术,包括:电压和电流钳位技术、单通道电流记录技术、通道 蛋白分离、纯化等生化技术、人工膜离子通道重建技术、通道药物学、基因重组技术及一些物理和化学技术。
电压钳位技术
一般而言,膜对某种离子通透性的变化是膜电位和时间的函数。通过玻璃微电极与细胞膜之间形成紧密封接, 利用电子学技术施加一跨膜电压并把膜电位固定于某一数值,可以测定该膜电位条件下离子电流随时间变化的动 态过程。利用药物或改变细胞内外的溶液成分,使其他离子通道失效,即可测定被研究的某种离子通道的功能性 参量,分析离子电流的稳态和动力学与膜电位、离子浓度等之间的关系,可推断该种通道的电导、活化和失活速 率、离子选择性等,并能测量和分析通道的门控电流的特性。
通道简介
生物膜离子通道活体细胞不停地进行新陈代谢活动,就必须不断地与周围环境进行物质交换,而细胞膜上的 离子通道就是这种物质交换的重要途径。人们已经知道,大多数对生命具有重要意义的物质都是水溶性的,如各 种离子,糖类等,它们需要进入细胞,而生命活动中产生的水溶性废物也要离开细胞,它们出入的通道就是细胞 膜上的离子通道。
钠通道
从电鳗电板分离的钠通道蛋白质分子量是208321,是由1820个氨基酸组成的多肽序列,可分为4个相似的区 段,每个区段中分别有较集中的正电性和负电性的氨基酸序列节段。多种钠通道构象模型的共同特征是:由多个 α螺旋跨膜排列组成通道,通道内侧应富含极性的氨基酸残基侧链,每个通道的控制部分由离子选择性滤器、活 化闸门和失活闸门3部分组成,其实体是氨基酸侧链的极性基团。膜电位变化时,电场诱导极性基团运动,使通道 局部构象发生变化,导致通道的开放、失活或关闭,并产生门控电流。
最新研究
离子通道和转运蛋白非各自为战
细胞膜离子通道的结构与功能

细胞膜离子通道的结构与功能细胞膜离子通道是一种膜蛋白,它通过细胞膜中的离子通道使离子在细胞膜上跨膜。
细胞膜离子通道可以根据其通透性和区分度被分为很多不同类型,包括钾离子通道、钠离子通道和氯离子通道等。
钾离子通道通过调节细胞的内外钾离子浓度和电位水平来控制许多生理生化过程。
钠离子通道可以影响神经肌肉细胞的动作电位,这对于神经传导非常重要。
氯离子通道在电动势调制中也起着重要的作用。
细胞膜离子通道的特殊性质大大增强了细胞对于不同的磁场、电场和其他刺激源的响应能力,并且对于人类健康和疾病的发生和进展也具有很大的影响。
一、细胞膜离子通道的基本结构细胞膜离子通道由一些单独的蛋白亚单位组成,它们通过与细胞膜中的脂质相互作用来形成细胞膜上的特异性孔道。
细胞膜离子通道主要由四个或五个轴突结构域组成。
每个结构域包含六个横式跨膜α螺旋,其中第四个α螺旋称为“色胺酸残基”,这个位置可以灵活转换造成离子通道的开放和关闭。
另外,细胞膜离子通道通常和一些调节因子和离子结合来形成复杂的膜蛋白,这些因子为它们对离子的通透性和特异性选择起到至关重要的作用。
细胞膜内面的大部分离子通道都是可打开的,而这种开启和关闭可以通过刺激信号、分子绑定、电化学影响、机械力等多种因素调节。
有些离子通道可以随着时间的推移发生“失活”或“不激活”,这导致它们对离子数量和流通率的影响发生改变。
二、细胞膜离子通道的功能细胞膜离子通道在许多细胞重要的生理生化过程中发挥了作用。
例如,在肌肉细胞中,钙离子和钾离子的通道通过细胞膜的电位差联合来调节细胞肌肉收缩。
在神经元中,钠离子和钾离子的通道通过细胞膜内的电势差调节神经元的动作电位。
在胰岛细胞中,钙离子和钾离子的通道在胰岛素的分泌过程中发挥关键的作用。
除此之外,还有一些离子通道的活动还可以通过信号转导机制调节在细胞分化、增殖、凋亡、细胞周期、固态化和因子释放等方面。
可以说,细胞膜离子通道的开放和关闭对于机体的正常代谢和生物学等生理过程的控制有着极为重要的影响。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2010总复习一.基本定义和概念:欧姆定律:I=G×(E-E rev);膜电势=膜内-膜外;电流方向规定从胞内流向胞外为正;膜等效电路;离子选择性. E rev=RT/zF*ln(P A[A]o/P B[B]i)E rev=RT/F*ln((P K[K]o+P Na[Na]o+P Cl[Cl]i)/ (P K[K]i+P Na[Na]i+P Cl[Cl]o))动作电位产生原因:Na/Ca的内流, K的外流产生动作电位1.动作电位产生于Na通透性的增加:电击后,膜电位首先从V rest增加并超过0 mV达到最大值(叫去极化)之后开始下降至V rest或更低时叫复极化或超极化。
AP:All-or-none;有阈值(去极化10-15 mV);动作电位是一种脉冲式的电信号(常见于CNS中)。
见图。
有关动作电位的名词:去极化: 极性程度的减弱;复极化;超极化;动力学名词:激活activation:电导在一个短时间延时后急剧增加(去极化过程);失活:电导在一个短时延时后急剧增加后掉到低值(去极化过程中通道关闭);去激活deactivation:电导回到通道关闭的水平;尾电流:驱动力、通道开放;门电流/门电荷:位移电流,通道上电荷在电场力的策动下运动所致。
一般在10ms内消失,电流很小,可以通过阻断所有通道电流测得。
计算门电荷数/有效电荷:利用GV曲线的Slope得到门电荷或用方差法从电流中得到通道数或从门电流中得到总电荷数;O/C=exp(-(w-z g q e E)/k B T)O/(O+C)=1/(1+exp((w-z g q e E)/k B T))) 这里z g是等效电荷数。
,k为gv曲线中参数bQ10定义:会利用已知的Q10,计算速率常数随温度的变化;生物学中定义:Q10= k(T+10o C)/k(T),这里κ代表速率常数。
Eyring 速率定律:k(T) =k(0)exp(-∆G/RT). 用近似T=273>>10.任意温度间隔∆T的系数:Q∆T=(Q10)∆T/10.其它术语:Rundown:全细胞模式下,由于胞内的小分子能量物质可以通过电极流失而导致通道电流在很短时间内减小或消失,且不可恢复,可以在电极中加入ATP, Mg,cAMP等物质消除Rundown现象;Desensitization:加入Ach的几秒钟后,宏观终板电导减小或微观Ach电流减小。
这个过程叫配体通道去敏感。
Open Channel Block:当阻断剂在胞内有受体,而且仅当通道打开(去极化),才能阻断电流,也就是说当通道关闭时,TEA不能阻断电流,电流上升逐渐不变,所以阻断发生在静息电位时(关闭态)就不能进入孔内。
Use-dependent block:局部麻药的阻断效应,随着刺激频率的加快而增强。
Reope n:阻断剂NMS可以阻止Na通道失活和去激活,使通道在复极化时仍有开放特性,可以记录到hooker tail尾电流,且这个尾电流的值大于未阻断的水平,且时程延长。
依赖状态的阻断:QX,从胞外(以中快速率)在通道开放时阻断nAChR 通道,然后阻断通道关闭,延缓电流的去敏感。
Adaptation:感觉通道对于一种恒定的刺激会产生减少冲动发放频率的适应性反应。
它的产生原因:电压依赖,胞内的mg离子阻断效应。
1、什麽是内向整流?它的产生原因?胞内的Mg2+等阻断;(C5)内向整流K通道(Inwared rectifier)心脏的非起博细胞。
去极化时关闭。
又叫Kir 通道。
功能为当膜电位比静息电位更负时Kir 通道会将膜电位拉回来。
而它的小外向电流使得膜电位维持在E K。
这种通道有多种,如K ir6 即K ATP通道在胞内ADP/ATP的比增高时,通道打开。
又叫ATP 阻断。
K ir3 在G蛋白被激活时打开。
在心脏中它们又叫K Ach(GIRK4)。
Kir 通道的GV曲线的上升斜率较缓慢。
所以其等效门电荷较Na通道为小。
内向整流来自胞内的多价离子,如Mg2+。
什麽是外向整流?复极化期突触前和突触后间隙的连接通道:像一个二极管。
电流是外向整流的。
信号传导的单方向性。
这也意味着突触前和突触后细胞的不对称性。
所以两边的通道不同。
延迟整流通道的含义?(C3)延迟整流钾通道。
(delayed rectified potassium channel)不同的钾通道对TEA有不同灵敏度;不同激活特性(快,慢,很慢);失活;Ca依赖;电压依赖;ATP敏感的钾通道。
电流存在一定的延时性,快(缩短动作电位),慢(帮助动作电位复极化)。
二.实验中设计指令电压的方案设计得到以下结果:激活(GV)曲线(有三种方法:稳态、峰值或尾电流法);电导定义:g Na=I Na/(E-E Na),g K=I K/(E-E K)E Na和E K电动势。
描述函数为:1/(1+exp((V-V0.5)/b))或为a/(1+exp((V-V0.5)/b))或a/(1+exp((V-V0.5)/b))+c这里,a, b(>0), c和V0.5为待定常数。
G 与几率的关系:I=nip o=GE; 当n, i, E为常数时,G与P o成正比。
尾电流与IV和GV如果有较大的尾电流的话,用尾电流测GV要好于用峰值和稳态值。
稳态失活曲线;g Na是由两个动力学过程组成.激活和失活一旦通道失活,通道的再打开必须要经过去激活过程。
(超极化)描述函数为:1/(1+exp((V-V0.5)/b))或为a/(1+exp((V-V0.5)/b))或a/(1+exp((V-V0.5)/b))+c 这里,a, b(<0), c和V0.5为待定常数。
失活恢复曲线;前一个是条件脉冲,后一个是检验脉冲。
描述函数是:1-exp(-t/τ);a*(1-exp(-t/τ)); a*(1-exp(-t/τ))+b这里,a, b和τ为待定常数。
平均电流迹线;一组电流迹线IV曲线的尾电流法;膜电容简易测量方法;膜平均电容:C M=1 F/cm2膜电容:C=Q/E;dE/dt=I c/C 这里C= 0A/d。
dE/dt= I c/C=-E/RCE=E0exp(-t/RC)= E0exp(-t/ ) Fig.1.2阻断剂的对通道阻断的IC50浓度实验:多种浓度测量法和单一浓度测量法。
1.单一毒素浓度测定K d值法求阻断剂的K D:每隔一个固定时间比如10秒,膜电压从保持电压跳到检验电压以得到电流的蜂值,然后加药测量电流峰值随时间的改变至一相对稳态,然后用没有毒素的溶液灌洗至基本完全恢复。
2.药和毒素和受体作用:受体:含有结合位点的分子。
k1T+R←→TR (这里,假定只有一个结合位点)k-1d([TR])/dt=k1[T][R]-k-1[TR]=0 (平衡态)K d=k-1/k1=[T][R]/[TR]受体结合Toxin的比率(通道被阻断的比率):y=[TR]/([TR]+[R])=1/(1+K d/[T])自由受体率:1-y=1/(1+[T]/K d)另一种看法:k1[T]O←-----→Bk-1dP O/dt=-k1[T]P O+k-1(1-P O);P B=1-P O平衡时有-k1[T]P O+k-1(1-P O)=0P O=k-1/(k1[T]+k-1)=1/(1+[T]/K d); P B= k1[T]/(k1[T]+k-1)=1/(1+K d/[T])多次重复上述计算后,我们有:Hill 方程(n个结合位点):y=1/(1+(K d/[T])n) 协同性; n 是Hill 系数K d=IC50, EC50 Fig. 3.3Na通道阻断剂量曲线电压依赖的阻断的测量方法:在不同电压下阻断电流。
得到IC50(V) Exp(qV)其中Ca电流,Na电流,K电流,nAChR电流的Protocol各不相同,原因是它们的失活和反转电位各异。
比如, HV A和LV A不同的是LV A有失活,HV A却没有失活等等。
问:如何记录LV A电流?如何记录nAChR电流(静息时加Ach刺激)?电压分类:HV A(高电压激活,无失活或部分失活,慢失活).LV A (低电压激活,完全失活,快失活)确定离子通透性的GHK方程:E rev=RT/F*ln((P K[K]o+P Na[Na]o+P Cl[Cl]i)/(P K[K]I+P Na[Na]i+P Cl[Cl]o))。
这里P A/P B是通透比注意尾电流法中的瞬时电流值的选取的原则是避免电容电流的最近点,而非Peak值。
用非稳态方差法求通道总数N和单通道电流的要点时:各Trace是非平均Trace;消除漏电流;三.通道结构(第十三章和第十七章)1.nAChR,Na, K, Ca 以及阳离子通道的一级拓扑分子结构特点2.nAChR,Na, K, Ca 以及阳离子通道中的pore结构特点,阻断及离子选择性机制。
3.钾通道过滤器的氨基酸序列特征序列是什麽?TXXTXGYG;该序列的排列相当于水合离子结构。
这被叫作Signature Sequence。
四个TXGYG在前腔形成K离子过滤器。
至少有两个跨膜部分的P环就可形成孔道。
4.探测通道孔区结构的方法:即用Cysteine,Alanine等突变结合氧化剂MTS和其他阻断剂的方法SCAM (substituted-cystein accessibility method)方法:(如果必要的话,去除所有的原生型cysteine)1)逐一用半胱氨酸Cysteine替换到所希望的各残基,但要保持功能不变。
2)再用MTS检验是否有永久性阻断(-SS-引起永久型阻断)。
5.比较K V,K ATP和KCNQ等离子通道的辅助β亚基与其α亚基的结合方式异同和β亚基的功能。
四.阻断机制(第十六章和第十七章)1.阻断速率的单通道鉴别;阻断剂在通道结构研究中的应用?三种time scale2.Hill方程y=1/(1+(K d/[T])n); Hill系数n的含义?Hill 方程(n个结合位点):y=1/(1+(K d/[T])n);k1T+R TR (这里,假定只有一个结合位点)k-1d([TR])/dt=k1[T][R]-k-1[TR]=0 (平衡态)K d=k-1/k1=[T][R]/[TR]受体结合Toxin的比率(通道被阻断的比率):y=[TR]/([TR]+[R])=1/(1+K d/[T]) 自由受体率:1-y=1/(1+[T]/K d)3.质子阻断Na通道的机制;TEA和失活的竞争机制要点;QA阻断nAChR通道机制;表面电荷理论:通过降低pH来减少负表面电荷,从而影响单通道电导酸基理论:通过降低pH使孔内重要的酸基得以改变,从而影响单通道电导因为质子的结合点在电场中,它的结合和释放速率常数均是电压依赖的。