DNA 聚合酶δ 结合蛋白38 是microRNA-291a-5p 的一
microRNA-361-5p在人类恶性肿瘤中的研究进展

2021,25(2):117-121.实用临床医药杂志Journal of Clinical Medicine in Practice-117-microRNA-361-5p在人类恶性肿瘤中的研究进展齐媛,郭宝良(哈尔滨医科大学附属第二医院乳腺外科,黑龙江哈尔滨,150000)摘要:研究表明,microRNA(miRNA)在恶性肿瘤的发生、发展中起重要作用。
miRNA通过各种机制参与细胞基因转录调控,其中与下游靶基因mRNA的特异性结合并使其降解为经典的调控机制。
近年来,miR-367-5p在恶性肿瘤中的表达失调得到了验证,通过调节与肿瘤生长、转移、上皮-间质转化(EMT)等方面相关的靶基因,进一步参与恶性肿瘤的增殖、凋亡、转移以及耐药性等相关生物学过程,并为恶性肿瘤的诊断及预后预测提供重要依据。
作者对miR-367-3p在不同肿瘤中的作用及相关机制进行综述,并展望其应用前景。
关键词:微小核糖核酸;恶性肿瘤;基因调控;靶基因中图分类号:R730.2;R329.2文献标志码:A文章编号:1672-2353(2027)02-177-35D0I:10.7619/jcmp.20201614Research progress of microRNA-311-5pin human malignant tumorsQI Yuan,GUO Baoliang(Department of'Breast Surgery,the Second^filiated Hospital of'Harbin MedicalUniversity,Harbin,Hedongjiaag,173000)Abstroch:Studids have showa thai microRNA(miRNA)plays ca inponaai aid in thd occao-naca ani deyelopmeat of tumors.MiRNA pdnicipdtds R thd resulatiou of callulao gess traa-scnptiou throorU diRenat mectanisms,amoo-which tha speciRc binCina ant dearaVatioo of dowa-stream tar-el geac mRNA is tha most classic reaulato—mechanism.Ia receai years,tha dysreaulatioo of miR-261-3p expnssioa in malinnaat tumors has baa yaified.By reaulatina tarad relatea to-tUat associatea with tumon growth,eaitUelial-meseachymai transitioo(EMT)and otUcn aspects,miR-361-3p furthen iavolve in tha relevaai bRlooical processas of rmainridat tumom,iacUrRa proliRratioo, dpoptosis,metastasis,v V drug resistaaca,v V R proviavs aa iRportaai basis foe tha diaaaosis ana prooaosis of mpin—vt tumoia.This卩1^>reviewea tha roles anC relatea mechanisms of miR-371-3p in diRereat tumors,and R s aaplicatiou prospect is prospectea.Key worCs:microRNA;malinnaat tumoo;uiv reaulatiou;tarael uiv1背景microRNA(miRNA)是一种非编码RNA,在进化中高度保守,人类基因2%的miRNA可通过调控网络影响机体近43基因的表达⑴。
大肠杆菌DNA聚合酶1的作用与应用资料

5’-3’外切核酸酶活性
从5'→3'方向水解DNA生长链前方的DNA链
,主要产生5'-脱氧核苷酸。这种酶活性只对
DNA上配对部分(双链)磷酸二酯键有切割活
力作用,方向是5'→3'。
3'
Mg2+
5'
5'
3'
DNA聚合酶I
3’-5’外切核酸酶活性
由3’端水解DNA链,反应底物是带3'-OH的双 链DNA或单链DNA,其活性从3'-OH端降解 ds-DNA,可被5' 3'聚合活性封闭,也可被 带5'磷酸的dNMP抑制
3' 5'外切酶活性
5'
3'
5' 3'聚合酶活性
切口平移
首先在镁离子存在下用少量DNaseI处理双链 DNA模板,产生少量切口。在切口处利用大 肠杆菌DNA聚合酶I的5’-3’外切核酸酶活 性是切口沿5’-3’方向移动,同时5’3’DNA聚合酶活性又不断合成DNA。
Mg2+ DNaseI
5'
3'
大肠杆菌DNA聚合酶I
切口平移法标记DNA
在Dnase的作用的基础上产生连内切口, 应用此酶的5’→3’聚合酶活性和5’→3’外切 酶活性的同步联合反应,使切口沿DNA链 从5’-端向3’-端移动。若用聚合反应的原料 dNTP带有放射性核素α-32P,就能使生成 的双链带有标记物。根据此原理可做DNA 杂交探针。
合成cDNA的第二链
大肠杆菌DNA聚合酶 I的活性
1 5’-3’DNA聚合酶活 性
2 5’-3’外切核酸酶活 性
中国药科大学生物化学精品课程习题核酸的代谢和蛋白质合成

中国药科大学生物化学精品课程习题核酸的代谢和蛋白质合成第一节核苷酸的代谢一、填空题1(人类对嘌呤代谢的终产物是。
2(痛风是因为体内产生过多造成的,使用作为黄嘌呤氧化酶的自杀性底物可以治疗痛风。
3(核苷酸的合成包括和两条途径。
4(脱氧核苷酸是由还原而来。
5(从IMP合成GMP需要消耗,而从IMP合成AMP需要消耗作为能源物质。
6(不能使用5-溴尿嘧啶核苷酸代替5-溴尿嘧啶治疗癌症是因为。
7(细菌嘧啶核苷酸从头合成途径中的第一个酶是。
该酶可被终产物抑制。
二、是非题1(黄嘌呤氧化酶既可以使用黄嘌呤又可以使用次黄嘌呤作为底物。
2(嘌呤核苷酸的从头合成是先闭环,再在形成N糖苷键。
3(IMP是嘌呤核苷酸从头合成途径中的中间产物。
4(真核细胞内参与嘧啶核苷酸从头合成的酶都位于细胞质。
5(嘧啶合成所需要的氨甲酰磷酸合成酶与尿素循环所需要的氨甲酰磷酸合成酶是同一个酶。
三、选择题(下列各题均有五个备选答案,其中只有一个正确答案) 1(嘌呤环1号位N原子来源于( )(A)Gln的酰胺N (B)Gln的α氨基N (C)Asn的酰胺N (D)Asp的α氨基N (E)Gly的α氨基N2(dTMP的直接前体是( )(A)Dcmp (B)dAMP (C)dUMP (D)dGMP (E)dIMP 3.人类嘧啶核苷酸从头合成的哪一步反应是限速反应,( )(A)氨甲酰磷酸的形成 (B)氨甲酰天冬氨酸的形成(C)乳清酸的形成 (D)UMP的形成(E)CMP的形成4(下面哪一种物质的生物合成不需要PRPP,( )+ (A)啶核苷酸 (B)嘌呤核苷酸 (C)His (D)NAD(P)(E)FAD 5(下列哪对物质是合成嘌呤环和嘧啶环都是必需的,( )(A)Gln/Asp (B)Gln/Gly (C)Gln/Pro (D)Asp/Arg (E)Gly/Asp四、问答题1(你如何解释以下现象:细菌调节嘧啶核苷酸合成的酶是天冬氨酸-氨甲酰转移酶,而人类调节嘧啶核苷酸合成的酶主要是氨甲酰磷酸合成酶。
分子生物学智慧树知到课后章节答案2023年下山东农业大学

分子生物学智慧树知到课后章节答案2023年下山东农业大学山东农业大学第一章测试1.格里菲斯转型实验得出了什么结论()答案:DNA是生命的遗传物质,蛋白质不是遗传物质2.现代遗传工程之父Paul Berg建立了什么技术()答案:重组DNA技术3.下列哪种技术可以用于测定DNA的序列()答案:双脱氧终止法4.RNA干扰是指由单链RNA诱发的基因沉默现象,其机制是通过阻碍特定基因的翻译或转录来抑制基因表达。
()答案:错第二章测试1.比较基因组学是基于基因组图谱和测序基础上,对已知的基因和基因组结构进行比较,来了解基因的功能、表达机理和物种进化的学科。
()答案:对2.以下哪项是原核生物基因组的结构特点()答案:操纵子结构3.细菌基因组是()答案:环状双链DNA4.下列关于基因组表述错误的是()答案:真核细胞基因组中大部分序列均编码蛋白质产物5.原核生物的结构基因多为单顺反子,真核生物的结构基因多为多顺反子。
( )答案:错6.病毒基因组可以由DNA组成,也可以由RNA组成。
( )答案:对第三章测试1.在原核生物复制子中以下哪种酶除去 RNA 引发体并加入脱氧核糖核苷酸?()答案:DNA 聚合酶 I2.使 DNA 超螺旋结构松驰的酶是()。
答案:拓扑异构酶3.从一个复制起点可分出几个复制叉?()答案:24.所谓半保留复制就是以 DNA 亲本链作为合成新子链 DNA 的模板,这样产生的新的双链 DNA 分子由一条旧链和一条新链组成。
( )答案:对5.DNA 的5′→3′合成意味着当在裸露3′→OH 的基团中添加 dNTP 时,除去无机焦磷酸 DNA链就会伸长。
( )答案:对第四章测试1.对RNA聚合酶的叙述不正确的是()。
答案:全酶不包括ρ因子2.原核生物RNA聚合酶识别启动子位于()。
答案:转录起始位点上游3.增强子与启动子的不同在于()。
答案:增强子与转录启动无直接关系4.启动子总是位于转录起始位点的上游。
应用于PCR技术的DNA聚合酶

合肥学院Hefei University基因工程题目: 应用于PCR技术的DNA聚合酶系别:专业班级:学号:姓名:指导教师: 2015年 12 月 26日应用于PCR技术的DNA聚合酶摘要DNA聚合酶作为聚合酶链式反应(PCR)的重要因素之一,在PCR过程中起着至关重要的作用,在某种意义上甚至可以说,PCR技术就是耐热DNA聚合酶的技术。
故综合研究聚合酶并改善其酶学性能,已成为分子生物学工作者关注的热点之一。
本文就PCR技术产生至今DNA聚合酶的研究历史及现状进行简要综述。
关键词PCR;DNA聚合酶;酶学特性1 DNA聚合酶的分类从大肠杆菌DNA聚合酶工被分离和鉴定至今的50年时间里,已有100多个DNA聚合酶相关基因被克隆和测序,它们分别来自于嗜热极端细菌和古细菌。
多种天然的和重组的耐热DNA聚合酶也被鉴定和纯化,这些酶大部分是单体酶,在溶液中的分子量一般为80一115KD。
人们根据核普酸序列将大肠杆菌酶DNA聚合酶划分为四类,即DNA聚合酶工((Family A ) , DNA聚合酶II (Family B ) , DNA聚合酶III (Family C)和其它的(Family X)。
DNA聚合酶工主要参与DNA损伤的修复,同时在半保留复制中起辅助作用。
DNA聚合酶II堤DNA复制的主要酶[4]。
与大肠杆菌DNA聚合酶工同源的为A群,该群除酵母线粒体DNA聚合工外,都来自于原核生物,对双脱氧核普酸抑制剂敏感,因此可用于DNA序列分析。
包括E. col 栖热菌属(Themus)的DNA聚合酶I、芽胞菌属(Bacillus)的DNA聚合酶I、噬菌体T5、以及升DNA聚合酶等。
与大肠杆菌DNA聚合酶II同源的为B群,对双脱氧核普酸不敏感甚至可以接受诸如双脱氧一UTP、生物素一dUTP〕或荧光标记的dUTP〕等修饰过的核普酸,因此这类DNA聚合酶可用于聚合酶链式反应制备或标记核普酸。
与大肠杆菌。
NA聚合酶II洞源的为C群,C群可分为两个亚群,即以蛋白质引导的(protein-prime)和以RNA引导的(RNA-prime) DNA聚合酶。
基因工程试题库讲解

《基因工程》试题库(一)一、选择题(单选或多选)(每题2分,共计20分)1.下面哪一种特性不是密码所具有的? ( )(a)偏爱性(b)简并性(c)重叠性(d)连续性2. 黏性末端连接法,不仅操作方便,而且( )(a)产生新切点(b)易于回收外源片段(c)载体不易环化(d)影响外源基因的表达3. 用菌落杂交法筛选重组体时,( )(a)需要外源基因的表达(b)不需要外源基因的表达(c)要根据克隆基因同探针的同源性(d)上述说法都正确4. DNA在染色体的常染色质区压缩多少倍?( )(a)6倍(b)10倍(c)40倍(d)240倍(c)1000倍10000倍5. 对于一个特定的起点,引发体的组成包括:( )(a)在起始位点与DnaG引发酶相互作用的一个寡聚酶(b)一个防止DNA降解的单链结合蛋白(c)DnaB解旋酶和附加的DnaC,DnaT,PriA等蛋白(d)DnaB,单链结合蛋白,DnaC,DnaT,PriA蛋白和DnaG引发酶(e)DnaB解旋酶,DnaG引发酶和DNA聚合酶Ⅲ6. 下面哪些是在反转录病毒中发现的基因?( )(a)gag(b)pol(c)env(d)OnC7. 下列哪些基因以典型的串联形式存在于真核生物基因组?( )(a)球蛋白基因(b)组蛋白基因(c)rRNA基因(d)肌动蛋白基因8. 以下关于抗体类型转变的叙述哪些是正确的?( )(a)每种重链具有不同的功能(b)类型转变的次序按染色体上重链排列顺序进行(c)一旦一种类型转换发生,其他的转换将不再进行(d)也可通过可变剪接改变重链9. 下列哪些转录因子含有TBP?( )(a)TFⅡB (b)TFⅢA (c)SLl (d)TFⅡD (e)TFⅢB (f)UBFl 10.剪接小体的组装( )(a)按有序的途径一步步完成(b)涉及snRNP与水溶性蛋白(即不是任何snRNP组分的蛋白)(c)不需要ATP(d)伴随着多次snRNP的重组合(e)以上都正确二、判断题(每题1分,共计10分)1.一个带有反向重复序列的双链DNA经变性后,复性时其单链可形成发夹环。
MicroRNA简介

基因。
它们就像散落于天幕的星星一样,分散在人类的基因组中。
而在这些基因间存在的大片大片的DNA 片段是不能编码蛋白质的,即“非编码序列”。
由于功能不清,加州理工学院的大野·乾于1972年提出用“垃圾基因”的概念来形容它们。
30亿个碱基对组成的DNA序列中,只有1%—2%编码蛋白。
,其中的的信息是看似无用的“垃圾”。
能制造出无转译能力RNA的DNA序列。
此类DNA在真核生物的基因组中占有大多数。
远远超过编码基因,有很长的一段时间科学家们没有认为这些非编码的成蛋白质,但却在生命的舞台上扮演着不同的角色。
迄今为止,细胞中的rRNA、tRNA、snRNA、asRNA、snoRNA、miRNA、piRNA都是非编码“垃圾”DNA合成的。
它们参与到基因活化、基因沉默、基因印记、剂量补偿、蛋白合成与功能调节、代谢调控等众多生物学过程中。
科学家们已经发现:“垃圾”DNA的功能之一就是调节基因的活动,如同一道指令一样,控制着基因。
一些控制基因开和关的特殊蛋白(转录因子)能特异识别基因附近的非编码“垃圾”DNA,通过与它们相互作用参与基因的抑制与激活。
科学家还发现,大多数基因的开启和关闭是由附近的“垃圾”DNA控制的。
它们就像是基因的“分子”开关,调节基因的活动。
拥有更大基因组的哺乳动物中,虽然特殊的有功能的“垃圾”DNA的分布要比在酵母中分散,但却在编码蛋白序列的上下游区域内呈簇分布。
这部分的研究由于与疾病的相关性,因此收到科学家的瞩目。
这么少的基因耗费了如此大的基因组是不是太过于浪费啦?那么,在学术界和新闻媒体中广为流传的“基因的墓场”是真的吗?经过漫长岁月的进化,许多“垃圾DNA”序列被顽固地保留下来,这是不是在暗示它们有着不可或缺的功能?“垃圾”DNA对生物意味什么?海量“垃圾”的存在引发了科学家们浓厚的兴趣并重拾对“垃圾”序列的关注,也掀起了一场声势浩大的从垃圾中寻宝的浪潮。
“合”指的是合理。
亿万年的进化使不合理的基因基本上都被淘汰了,所以存在的基因其功能必定是合理的,至少具有合理性的一面。
taq dna聚合酶名词解释

taq dna聚合酶名词解释
Taq 聚合酶是从嗜热细菌水生栖热菌中提取的热稳定(热稳定)DNA 聚合酶。
它的主要功能是聚合酶链式反应(PCR) 技术,它使扩增特定DNA 序列的重复步骤自动化。
聚合酶链式反应可以使DNA 分子增加多达十亿倍。
这会产生大量特定基因,并在多种应用中下游使用。
Taq DNA 聚合酶已包含在称为家族A 的DNA 聚合酶家族中。
PCR 使用来自(仅)家族A 和家族B DNA 聚合酶的DNA 聚合酶。
家族DNA 聚合酶包括Tth 和Tma DNA 聚合酶以及Taq,并具有5'-3' 外切核酸酶活性,但通常缺乏3'-5'。
在没有3'-5' 核酸外切酶活性的情况下,A 族聚合酶在掺入碱基对时容易出错。
西南大学[1194]《生活中的DNA科学》
![西南大学[1194]《生活中的DNA科学》](https://img.taocdn.com/s3/m/b7004a4d453610661ed9f483.png)
1.A.V
2.B.X
44、原核RNA聚合酶可以4中NTP为底物催化RNA的合成。
1.从DNA到DNA
2.从DNA到RNA
3.从RNA到蛋白质
4.从蛋白质到RNA
20、识别10碱基序列的限制酶,大约每隔()进行一次切割。
1.410
2.104
3.210
4.102
21、下列事件中,不属于表观遗传调控的是()
1.DNA甲基化
2.组蛋白乙酰化
3.mRNA加尾
4.RNA干扰
22、重组DNA勺基本构建过程是将()
4.PrPC对蛋白酶K敏感
16、其中两对基因连锁,交换值为0,—对独立遗传()种
1.一种
2.二种
3.三种
4.四种
17、下列关于DNA连接酶的叙述中,正确的是()
1.DNA连接酶连接的是两条链碱基对之间的氢键
2.DNA连接酶连接的是黏性末端两条链主链上的磷酸和脱氧核糖
3.DNA连接酶连接的是黏性末端两条链主链上的磷酸和核糖
4.以上都是
11、下列哪种碱基不属于DNA/RNA勺碱基()
1.腺嘌呤
2.鸟嘌呤
3.次黄嘌呤
4.胸腺嘧啶
12、下列哪项不是法医DNA分析技术的衍生技术()
1.RT-PCR
2.SSP-PCR
3.PCR-SSOP
4.MVR -PCR
13、下列哪项不属于现在主要开发研究的微型化DNA分析仪器()
1.微芯片毛细管电泳装置
1、下面哪种酶是在重组DNA技术中不常用到的酶()
1.限制性核酸内切酶
2.DNA聚合酶
3.DNA连接酶
4.DNA解链酶才
2、长期接触X射线的人群,后代遗传病发病率明显升高,主要原因是该人群生 殖细胞发生()
病理学技术相关专业知识分类模拟题5

病理学技术相关专业知识分类模拟题5B型题以下提供若干组考题,每组考题共同在考题前列出A、B、C、D、E五个备选答案,请从中选择一个与考题关系最密切的答案。
A.核苷酸在核酸链上的排列顺序B.tRNA的三叶草结构C.DNA双螺旋结构D.DNA的超螺旋结构E.DNA的核小体结构1. 属于核酸一级结构的描述是答案:A2. 属于核糖核酸二级结构的描述是答案:B3. 属于真核生物染色质中DNA的三级结构的描述是答案:E核酸的一级结构指核苷酸链上核苷酸的排列顺序。
DNA的二级结构是双螺旋结构,在此基础上还可以形成更高级的超螺旋结构。
真核生物的DNA可在双螺旋基础上形成核小体(三级结构),再多次盘绕成染色体。
RNA单链内部也可以形成局部互补的螺旋区,如tRNA的二级结构(三叶草结构)及局部螺旋扭曲形成的三级结构(“倒L形”结构)。
A.双股DNA解链成单股DNAB.单股DNA恢复成双股DNAD.DNA分子超螺旋解开E.磷酸二酯键断裂4. DNA变性是指答案:A5. DNA复性是指答案:B6. DNA水解是指答案:EDNA变性是指在某些理化因素作用下维系DNA双螺旋的次级键断裂,DNA双螺旋分子解开成单链的过程。
DNA变性可以使其理化性质发生一系列改变,包括黏度下降和OD260紫外吸收增加(增色效应)等。
DNA复性指变性后解开的单链重新缔合形成双螺旋。
DNA复性后出现OD260紫外吸收降低(减色效应)。
A.tRNAB.mRNAC.hnRNAD.snRNAE.rRNA7. 组成核糖体的是答案:E8. 含有较多稀有碱基和核苷的是答案:A9. 蛋白质生物合成的模板是10. mRNA的前体是答案:C11. 带有遗传密码的是答案:B12. 参与hnRNA的剪接和转运的是答案:D动物细胞内主要含有mRNA、tRNA、rRNA三种核糖核酸。
其中mRNA含有遗传密码,作为蛋白质合成的模板;tRNA活化、转运氨基酸并识别mRNA上的密码,参与蛋白质的合成,其一级结构中含有较多的稀有碱基;rRNA与蛋白质组成核糖体作为蛋白质合成的场所。
几种不同DNA聚合酶比较

合理选择耐热DNA聚合酶是PCR成败与否的一个关键因素,如何选择最合适的耐热聚合酶,是进行PCR实验首先要考虑的问题。
目前市面上有许多耐热DNA聚合酶,虽然名称各不相同,但主要区别在于特异性、保真性、耐热性、扩增速率、扩增片段长度等几个指标。
Taq DNA聚合酶存在于水生嗜热菌(Thermus aquaticus)的嗜热DNA聚合酶,可在74℃复制DNA,在95℃仍具有酶活力。
该酶可在离体条件下,以DNA为模板,延伸引物,合成双链DNA。
这个酶只有5′→3′DNA聚合酶活性和5′→3′的外切酶活性,缺少3′→5′的外切酶活性,即具有5’-3’DNA聚合酶活性和对双链DNA特异的5’-3’外切核酸酶活性,无3’-5’外切酶活性。
使用该产品扩增得到的PCR产物的3’末端附有一个“A”碱基,可直接用于TA克隆。
由于不具有3'-5'外切酶活性,因而在合成中对某些单核苷酸错配没有校正功能。
TaqDNA聚合酶还具有非模板依赖性活性,可将PCR双链产物的每一条链3'加入单核苷酸尾,故可使PCR产物具有3'突出的单A核苷酸尾;另一方面,在仅有dTTP存在时,它可将平端的质粒的3'端加入单T核苷酸尾,产生3'端突出的单T核苷酸尾。
应用这一特性,可实现PCR产物的T-A克隆法。
适用于:普通PCR,TA克隆Super Taq DNA聚合酶Super Taq DNA聚合酶是由耐热的Taq DNA聚合酶和具有3’-5’外切酶活性的高保真DNA聚合酶按照最合适的比例混合而成,具有扩增效率高和错配率低的优良性能,使用该产品扩增得到的PCR产物的3’末端附有一个“A”碱基,可直接用于TA克隆。
适用于:要求效率高保真性好的PCR反应。
UlltraPF™DNA聚合酶UlltraPF™DNA Polymerase是一种耐热的超保真DNA聚合酶,具有5’-3’聚合酶活性和3’-5’外切酶活性,具有比Pfu DNA Polymerase更好的保真性能和更快的扩增效率。
部编版高中生物必修二第四章基因的表达带答案专项训练
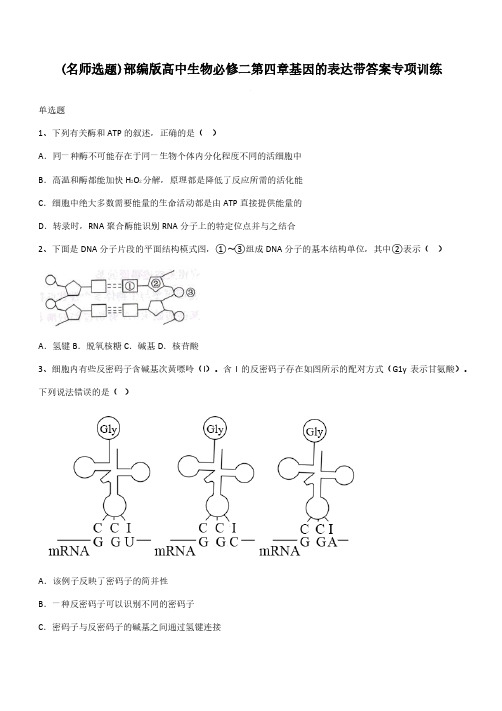
(名师选题)部编版高中生物必修二第四章基因的表达带答案专项训练单选题1、下列有关酶和ATP的叙述,正确的是()A.同一种酶不可能存在于同一生物个体内分化程度不同的活细胞中B.高温和酶都能加快H2O2分解,原理都是降低了反应所需的活化能C.细胞中绝大多数需要能量的生命活动都是由ATP直接提供能量的D.转录时,RNA聚合酶能识别RNA分子上的特定位点并与之结合2、下面是DNA分子片段的平面结构模式图,①〜③组成DNA分子的基本结构单位,其中②表示()A.氢键B.脱氧核糖C.碱基D.核苷酸3、细胞内有些反密码子含碱基次黄嘌呤(I)。
含I的反密码子存在如图所示的配对方式(G1y表示甘氨酸)。
下列说法错误的是()A.该例子反映了密码子的简并性B.一种反密码子可以识别不同的密码子C.密码子与反密码子的碱基之间通过氢键连接D.密码子与反密码子空间结构的不同造成其功能不同4、豌豆的圆粒和皱粒是由R、r基因控制的一对相对性状,当R基因插入一段800个碱基对的DNA片段时就成为r基因。
豌豆种子圆粒性状的产生机制如下图所示。
下列分析错误的是()A.R基因插入一段800个碱基对的DNA片段属于基因突变B.图示说明基因可通过控制酶的合成间接控制生物体的性状C.过程①需RNA聚合酶参与,能发生A-U、C-G、T-A配对D.参与过程②的mRNA上每三个相邻碱基都能决定一个氨基酸5、人体内苯丙氨酸的代谢途径如下图所示,下列叙述错误的是()A.缺乏酶⑤会导致人患白化病B.缺乏酶③会导致人患尿黑酸症C.据图可知基因能通过控制蛋白质的结构直接控制生物体的性状D.苯丙酮酸积累可阻碍脑的发育,需给此类患儿提供不含苯丙氨酸的特殊奶粉6、作为生物学的核心规律之一,中心法则反映了遗传信息的传递和表达规律。
在下图所示的中心法则中虚线表示少数生物遗传信息的流向。
下列有关遗传信息流向的说法错误的是()A.DNA、RNA都是遗传信息的载体,蛋白质是遗传信息的表达产物B.ATP可以为图中遗传信息的传递过程提供能量C.过程②和④所需模板不同,所需的原料、酶都相同D.染色体上基因的②③过程是在不同结构中进行的7、下列关于密码子的叙述,错误的是()A.一种氨基酸可能有多种与之相对应的密码子B.起始密码子是核糖体进行翻译的起点C.每种密码子都有与之对应的氨基酸D.信使RNA上的GCA在人细胞中和猪细胞中决定的是同一种氨基酸8、关于基因控制蛋白质合成的过程,下列叙述正确的是A.一个含 n 个碱基的 DNA 分子,转录的 mRNA 分子的碱基数是 n / 2 个B.细菌的一个基因转录时两条 DNA 链可同时作为模板,提高转录效率C.DNA 聚合酶和 RNA 聚合酶的结合位点分别在 DNA 和 RNA 上D.在细胞周期中,mRNA 的种类和含量均不断发生变化多选题9、当细胞中缺乏氨基酸时,负载tRNA(携带氨基酸的tRNA)会转化为空载tRNA(没有携带氨基酸的tRNA)参与基因表达的调控。
2021年安师大分子生物学实验课考核考试题库和答案

分子生物学实验课考核考试题库1.PCR原理是什么?2.PCR普通体系应加入哪些试剂?3.PCR防止其他DNA污染, 应注意哪些问题?4.影响PCR产物产量因素重要有哪些?5.引物设计应注意哪些问题?6.简述克隆某一原核生物基因片段普通操作环节?7、什么是质粒?质粒基本性质有哪些?8、质粒抽提实验中溶液I、II、III各有什么作用?9、碱裂解法抽提质粒DNA基本原理是什么?10、在碱裂解法提取质粒DNA操作中应注意哪些问题?11.在碱裂解法提取质粒DNA时, 酚/氯仿作用是什么?12.碱裂解法提取质粒DNA分子普通有几种构象?电泳时移动速率有何差别?13.提取植物基因组DNA基本原理是什么?在操作过程中应当注意什么?14、在植物基因组提取过程中, DNA 降解也许因素?15.在植物基因组提取过程中, 提高DNA 产量办法?16.在使用苯酚进行DNA提取时应当注意什么?17、在提取植物基因组DNA过程中, 使用苯酚与氯仿作用是什么?18、在植物基因组提取过程中, 该如何检测和保证基因组DNA质量?19、什么是限制性内切酶?20、常用Ⅱ型限制性内切酶切割双链DNA后会产生末端或末端。
21.下列关于限制性内切酶描述, 错误是: ()A. 一种限制性内切酶只能辨认一种特定脱氧核苷酸序列B. 限制性内切酶活性受温度影响C. 限制性内切酶能辨认和切割RNAD. 限制性内切酶可以从原核中提取22、既有一长度为l 000 bpDNA分子, 用限制性核酸内切酶EcoR I酶切后得到DNA分子仍是l 000 bp, 用Kpn I单独酶切得到400 bp和600 bp 2种长度DNA 分子.用EcoR I, Kpn l同步酶切后得到200 bp和600 bp 2种长度DNA分子。
请画出该DNA也许酶切图谱。
23.一种限制性内切酶辨认序列和切割位点是5ˊ一G↓GATCC一3ˊ, 在质粒上有该酶一种切点, 请画出质粒被该限制性内切酶切割后所形成黏性末端。
DNA聚合酶的定义

DNA聚合酶的定义发布时间: 2010-6-3 浏览次数: 775 次DNA聚合酶的定义DNA聚合酶(DNA polymerase)是细胞复制DNA 的重要作用酶。
DNA聚合酶 , 以DNA为复制模板,从将DNA由5'端点开始复制到3'端的酶。
DNA聚合酶的主要活性是催化DNA的合成(在具备模板、引物、dNTP 等的情况下)及其相辅的活性。
真核细胞有5种DNA聚合酶,分别为DNA聚合酶α(定位于胞核,参与复制引发具5'-3'外切酶活性),β(定位于核内,参与修复,具5'-3'外切酶活性),γ(定位于线粒体,参与线粒体复制具5'-3'和3'-5'外切活性),δ(定位核,参与复制,具有3'-5'和5'-3'外切活性),ε(定位于核,参与损伤修复,具有3'-5'和5'-3'外切活性)。
原核细胞有3种DNA聚合酶,都与DNA链的延长有关。
DNA聚合酶I是单链多肽,可催化单链或双链DNA 的延长;DNA聚合酶II则与低分子脱氧核苷酸链的延长有关;DNA聚合酶III在细胞中存在的数目不多,是促进DNA链延长的主要酶。
DNA聚合酶的发现在50年代的中期,A. Kornberg和他的同事们就想到DNA的复制必然是一种酶的催化作用,于是决心分离出这种酶并研究其结构和作用机制。
为了达到这个目的,他们分离的蛋白,然后加到体外合成系统中即同位素标记的dNTP、Mg2+及模板DNA,经过大量的工作,于1956年终于发现了DNA聚合酶Ⅰ(DNA polymerase Ⅰ,DNA pol Ⅰ)原来称为Kornberg酶。
以后又相续发现了DNA pol Ⅱ和DNA pol Ⅲ。
开始人们以为DNA pol I是细菌中DNA复制主要的酶类,后来发现DNA pol Ⅰ的突变株照样可以复制,才清楚它并不是主角。
microRNA-199a-5p对口腔鳞状细胞癌细胞上皮间质转化及侵袭转移的影响机制

第45卷第18期2222年18月贵州医科大学学报Vol.45No.18 JOURNAL OF GUIZHOU MEDICAL UNIVERSITY2222.18micaRNAN99aNy对口腔鳞状细胞癌细胞上皮间质转化及侵袭转移的影响机制*卢静8,李杰4,张莹8,吴栋材4(8-北京市朝阳区中医医院口腔科,北京190222;2.北京大学第一医院口腔科,北京-0234)[摘要]目的:探究MicaRNAT99gAp通过靶向激酶p((KKp)-核因子k B(NF n B)信号通路影响口腔鳞状细胞癌细胞上皮间质转化及侵袭转移的机制。
方法:将离体培养的SCCN5、CALN7、TCANn3、SCCN OSCC和293口腔鳞状细胞癌细胞分为空白对照组(n=24) )NC模拟组(n=24)和miRT99gAp模拟物组(n=24)°逆转录定量聚合酶链反应(RT-fPCR)分析miRN99gNp和IKKp的转录水平,CCKN法检测细胞增殖水平,流式细胞仪检测细胞凋亡和周期分布,蛋白质印迹法检测抗IKKB、总p65、核磷酸化(cA p65、NF-rB(EB)-2抑制剂、y-I k B t、组蛋白H3、6-肌动蛋白及a-微管蛋白的表达水平,荧光素酶测定试剂盒定量检测NF-rB活性,进行Tran-swell迁移和侵袭分析肿瘤细胞的侵袭和迁移能力。
结果:在OSCC细胞中miR-199g-5h表达下调(P<0.01);与无淋巴结转移的细胞比较,有淋巴结转移的OSCC细胞中miRN99gNp表达降低(P<0.05);OSCC细胞SCCN5、CALN7、TCAAn3和SCCT中miRN99a-5p下调(P<0.05);RTPCR分析显示,与TCA-8113和SCCT细胞的NC 对照组比较,用mCT99gAp模拟物转染后,mCN99gAp的表达上调;CCK8分析表明,与模拟NC转染的细胞比较,mCN99gTp模拟物转染抑制了细胞活力,在TCA-8113和SCCT细胞中观察到的凋亡诱导;流式细胞仪分析的结果表明,在miRN99gTp过表达后,IKK[3表达下调,但在TCA-8113和SCCT细胞中敲低miRN99gTp后,IKK|3的表达上调;miRT99aAp模拟物抑制了pEB-c的水平,miRN99a-5p模拟物在蛋白质水平上均抑制了ph5表达,p65的过表达抑制了ZEB1、TWIST1、E-钙黏蛋白和波形蛋白的表达,减轻了miRN99gTp对OSCC细胞的迁移和侵袭的抑制。
sPE_患者血清miR-199a-5p、miR-142-3p表达变化及与产妇血管内皮损伤、围产儿结局

sPE患者血清miR-199a-5p、miR-142-3p表达变化及与产妇血管内皮损伤、围产儿结局的关系张丽萍,李霞,谢艳淮安市妇幼保健院产科,江苏淮安223002摘要:目的 观察重度子痫前期(sPE)患者血清微小核糖核酸-199a-5p(miR-199a-5p)、微小核糖核酸-142-3p (miR-142-3p)表达变化,并分析其与产妇血管内皮损伤、围产儿结局的关系,为围产儿结局的改善提供参考。
方法 选取sPE患者200例(观察组)和产检健康孕妇200例(对照组),采用RT-PCR法测算两组血清miR-199a-5p、miR-142-3p,酶联免疫吸附试验检测两组血清可溶性细胞内皮因子(sEng)、可溶性血管内皮生长因子受体-1(sFlt-1)、内皮素-1(ET-1)、血管内皮生长因子(VEGF),Pearson相关分析法分析观察组血清miR-199a-5p、miR-142-3p 与血管内皮功能相关指标的相关性。
根据围产儿结局将观察组分为结局不良组(29例)和结局良好组(171例),采用单因素分析法及多因素Logistic回归分析法分析患者血清miR-199a-5p、miR-142-3p表达与围产儿结局的关系。
结果 与对照组比较,观察组血清miR-199a-5p表达及VEGF水平降低,血清miR-142-3p表达及sEng、sFlt-1、ET-1水平升高(P均<0.05)。
观察组血清miR-199a-5p表达与sEng、sFlt-1、ET-1水平呈负相关,与VEGF水平呈正相关(P 均<0.05);血清miR-142-3p表达与sEng、sFlt-1、ET-1水平呈正相关,与VEGF水平呈负相关(P均<0.05)。
结局不良组和结局良好组年龄、早发型sPE比例、并发水肿比例、并发子痫比例、并发胎盘早剥比例、并发产后出血比例、血清miR-142-3p表达变化、血清miR-199a-5p表达变化比较,P均<0.05;多因素Logistic回归分析显示,早发型sPE、胎盘早剥、miR-142-3p高表达是sPE围产儿结局不良的危险因素(P均<0.05),miR-199a-5p高表达是sPE围产儿结局不良的保护因素(P<0.05)。
MiR-21-5p_通过下调电压门控钾离子通道Kv1.1_表达减轻大鼠三叉神经痛

中南大学学报(医学版)J Cent South Univ (Med Sci)2024, 49(1) MiR -21-5p 通过下调电压门控钾离子通道Kv1.1表达减轻大鼠三叉神经痛周雪雯1, 2,郭刚文1,余珊子3,胡蓉1[1. 中南大学湘雅三医院疼痛科,长沙 410013;2. 浙江大学医学院附属第一医院麻醉科,杭州 310009;3. 湖南省人民医院(湖南师范大学附属第一医院)麻醉科,长沙 410002][摘要] 目的:三叉神经痛(trigeminal neuralgia ,TN)是一种临床上常见的神经病理性疼痛。
电压门控性钾通道(voltage-gated potassium channel ,Kv)已被证实参与TN 的发生、发展,但具体机制仍不明确。
微RNA(microRNA ,miR)可通过调节三叉神经节(trigeminal ganglion ,TG)上Kv 通道的表达及神经元兴奋性,参与神经病理性疼痛。
本研究旨在探索TN 模型中TG 上Kv1.1和miR-21-5p 的关系,评估miR-21-5p 是否对Kv1.1有调控作用,为TN 的治疗提供新的靶点。
方法:将48只SD 大鼠随机分为6组:1)假手术组(sham 组,n =12),大鼠仅在术侧切口缝合,不结扎神经;2)Sham+agomir NC 组(n =6),sham 大鼠通过脑立体定位注射方法于术侧TG 微量注射agomir NC ;3)Sham+miR-21-5p agomir 组(n =6),sham 大鼠通过脑立体定位注射方法于术侧TG 微量注射miR-21-5p agomir ;4)TN 组(n =12),采用铬肠线慢性缩窄性眶下远端神经损伤(chronic constriction injury of the distal infraorbital nerve ,dIoN-CCI)法构建TN 大鼠模型;5)TN+antagomir NC 组(n =6),TN 大鼠通过脑立体定位注射方法于术侧TG 微量注射antagomir NC ;6)TN+miR-21-5p antagomir 组(n =6),TN 大鼠通过脑立体定位注射方法于术侧TG 微量注射miR-21-5p antagomir 。
MicroRNA在昆虫-病毒互作中的作用研究进展

MicroRNA在昆虫-病毒互作中的作用研究进展吴萍【摘要】MicroRNA(miRNA)是一类小的非编码RNA,广泛参与生长发育、细胞凋亡、肿瘤、抗病毒等一系列重要的生命过程。
近年来的研究发现,miRNA在宿主与病毒的互作机制中发挥重要作用。
文中结合近几年在昆虫miRNA抗病毒免疫作用方面的研究成果,简要综述了miRNA的产生机制,与靶基因的结合特点,昆虫宿主编码的miRNA及病毒编码的miRNA在昆虫宿主-病毒互作中的作用。
%MicroRNA(miRNA)as small non-coding RNA widely participates in a series of important biological processes including development,cell apoptosis,tumor and anti-viral responses.Recently,studies suggest that micorRNAs play key roles in host-virus interaction.This article reviewed the mechanism of microRNAs biogene-sis,interaction between miRNAs and target genes,the roles of miRNA in host-virus interaction based on the re-cent achievements in insects.【期刊名称】《江苏科技大学学报(自然科学版)》【年(卷),期】2016(030)002【总页数】6页(P183-188)【关键词】昆虫;病毒;microRNA;互作【作者】吴萍【作者单位】江苏科技大学蚕业研究所,江苏镇江212018【正文语种】中文【中图分类】S881.2MicroRNA(miRNA)是一类由18~25个核苷酸组成的小的非编码RNA.miRNA 广泛存在于动物、植物和病毒等生物中,在物种进化中高度保守,并具有时空表达特异性,参与包括生长发育、细胞凋亡、肿瘤、抗病毒等一系列重要的生命过程[1-2].RNA干扰 (RNA interference) 作为一种先天的抵抗病毒感染的防御机制最早是在植物中发现的[3], 随后研究发现,RNA干扰在昆虫自身免疫体系中也扮演着重要角色.研究表明:宿主编码的miRNA在受到病毒感染后,其表达水平会发生显著变化[4].差异表达的miRNAs可能是由宿主免疫应答反应引起或由病毒感染诱导的调控因子导致.另一方面,病毒编码的miRNA也可调控自身与病毒复制相关基因或宿主免疫相关基因,从而进一步增加了宿主—病毒互作的复杂性.文中结合近几年昆虫miRNA在抗病毒免疫机制中的作用研究成果,就miRNA的产生,与靶基因的调控及在昆虫—病毒互作中的作用研究进展作一简要综述.1.1 标准途径通常,由宿主细胞和DNA病毒编码的miRNA由标准途径生成.即首先由RNA 聚合酶Ⅱ将细胞核内编码 miRNA 的基因转录成初级转录物 (primary miRNA, pri-miRNA),初级转录物除具备典型的mRNA特征外(含5’端帽子和3’端poly A 结构),还具有1个或多个茎环结构.初级转录物在位于细胞核中的Drosha酶和DGCR8/Pasha 蛋白的共同作用下被加工成约 70 nt 的不完全配对茎—环结构的miRNA前体(precursor miRNA, pre-miRNA).而后 pre-miRNA 在核输出蛋白-5(Exprotin-5)的帮助下,从细胞核转运到细胞质,被细胞质中的Dicer 酶及其辅助因子TRBP切割成3’端有两个碱基突出的 18~25 个核苷酸长度的双链 miRNA:miRNA*分子.成熟的miRNA与Ago1或Ago2等复合物形成RNA诱导沉默复合体(RNA induced silencing complex, RISC),通过与靶mRNA的特定序列互补或不完全互补结合,诱导靶mRNA剪切或者阻止其翻译而对靶基因表达进行调控[5-9].1.2 非标准途径近年来,研究发现, miRNA前体可不依赖于由Drosha酶和DGCR8蛋白组成的miRNA加工子(Microprocessor)而从其他非标准途径生成(图1).例如,有些miRNA前体的生成无需Drosha酶,可由剪接体剪切短的含有发卡结构的内含子(称为mirtrons)而来[10];有些miRNA前体可由tRNase Z 酶剪切tRNA而来,如miR-1983[11];还有些miRNAs可来源于小核仁RNA(snoRNA),这类RNA主要位于核仁上,参与特定位点的甲基化和其他RNA的假尿苷化[12].以上来源的miRNA虽然其前体的生成可不依赖Drosha酶,但成熟的miRNAs仍需Dicer酶在细胞质中的剪切.1.3 RNA病毒基因组来源的miRNARNA病毒曾被认为是不可能编码miRNA的,理由如下:1) 大多数RNA病毒的复制都在细胞质中进行,而细胞质中缺乏Drosha酶,因而初级转录物没法形成miRNA 前体.2) 病毒编码的miRNA会诱导RNA干扰应答从而降解RNA病毒自身的基因组.事实上,miRNA生成的非标准途径无需Drosha酶的剪切,这为RNA病毒编码miRNA提供了可能性.研究表明:当DNA病毒编码的miRNA前体插入RNA 病毒基因组中,可被成功地加工为成熟的miRNA.如来源于EB病毒(Epstein barr virus)的pre-miR-BART2R插入到TBEV (Flavivirus tick-borne encephclitis virus)基因组中,被加工成miR-BART2,而不影响TBEV病毒的复制[14].来源于宿主的miR-124可从重组的正链RNA病毒、辛德毕斯病毒(Sindbis virus)中产生[15].近年来,在反转录病毒,如人类免疫缺陷病毒(Human immunodeficiency virus, HIV)、牛白血病病毒(Bovine leukemia virus, BLV)、猴泡沫病毒(Simian foamy viruse,SFV)中发现了大量的miRNAs[16-18].在RNA病毒中,如西尼罗病毒(West Nile virus, WNV)、登革热病毒(Dengue virus, DENV)以及甲型肝炎病毒(Hepatitis A virus, HAV)也被证实可编码miRNA[19-21].一般认为,人类以及大多数动物的miRNA 通过与靶基因 mRNA 的3’非翻译区(3’UTR区)完全或不完全互补配对,诱导靶mRNA的降解或抑制靶基因蛋白的合成,从而参与基因的表达调控.种子序列(成熟miRNA 5’端的第2~8个核苷酸)与靶基因3’UTR区的结合通常要求完全互补.越来越多的研究表明,种子序列的完全配对并不是必需的,可以允许有至多1个碱基凸起或错配[22].种子序列也不仅仅局限于miRNA 5’区,miRNA 3’端或中间序列与靶基因的配对也能发挥对靶基因的调控作用[22-23].起初人们认为,在动物中,miRNA与靶基因的结合配对仅局限于靶基因mRNA的3’UTR区.现在很多证据表明这类特异性结合也可发生在靶基因mRNA的5’非翻译区或开放阅读框区域.例如,由巨细胞病毒(cytomegalovirus)编码的miR-US25-1可以和细胞周期调控相关基因(BRCC3, EID1, CD147等)的5’UTR配对结合并抑制其表达[24].Argonaute (AGO) HITS-CLIP以及 PAR-CLIP数据分析表明,将近一半的AGO 与miRNA的结合位点位于开放阅读框内[25-26].研究表明:miRNA对靶基因不仅具有负调控作用,即降解靶RNA或翻译抑制,miRNA对靶基因还有正调控作用,这可能是因为增加了靶基因mRNA的稳定性或与靶基因启动子序列的结合引发了翻译激活.例如,miR-128可以诱导与脑发育相关基因的表达[27].文献[28]肝脏中特异表达的miR-122 与丙型肝炎病毒(Hepatitis C virus,HCV) mRNA 5’UTR区给合从而引起病毒增殖,同时还增加了HCV基因组RNA的稳定性.有些miRNAs 可以和靶基因启动子的TATA框结合使得靶基因的表达水平上调[29].一个miRNA可作用于多个靶基因,一个靶基因又可同时受多个miRNA调控,这使得miRNA与靶基因之间的调控更为复杂.3.1 昆虫编码的miRNA病毒与宿主相互作用是病毒感染致病以及免疫的生物学基础,病毒感染宿主后通过多种信号传导途径引起应答基因,包括miRNAs表达水平的变化.近些年,很多研究学者利用日趋先进的高通量测序技术对宿主受病原菌入侵后,宿主miRNA的表达谱进行了广泛研究.宿主感染病毒后miRNA表达谱的变化至少归为两类:一是为病毒的增殖提供更适宜的细胞内环境;二是诱导宿主免疫应答反应从而抑制病毒增殖[30-31].例如,文献[4]采用Solexa高通量测序技术对家蚕质型多角体病毒 (Bombyx mori cytoplasmic polyhedrosis virus, BmCPV)感染中肠及对照中肠组织进行了小RNA测序分析,58个microRNAs在家蚕感染中肠组织和对照中肠组织中的表达有显著差异.差异表达的miRNAs的靶基因预测分析表明大多数靶基因的功能与应激和免疫相关.文献[32]预测在果蝇中,有超过70个miRNAs 参与果蝇免疫系统相关的Toll信号通路、Imd信号通路以及JNK和JAK/STAT信号通路的调控,推测由miRNA介导的RNA干扰途径可能是果蝇先天的广谱型免疫机制.冈比亚按蚊编码(Anopheles gambiae)的两个miRNA, aga-miR-2304以及aga-miR-2390经预测可能靶向作用于抑制素和酚氧化酶原基因[33].文献[34]的研究发现,烟草天蛾(Manduca sexta)在受到病原菌侵染后,其miRNA表达谱变化涉及到编码病原物模式识别、酚氧化酶原激活、抗菌肽合成以及保守免疫信号转导等多个免疫应答相关通路的基因的表达.MiR-8是经实验室验证的,和昆虫免疫应答相关的miRNA.它可负调控果蝇中编码Drosmycin 以及Diptericin等抗菌肽(AMP)基因的表达[35],使得AMP在非感染情况下维持较低水平,从而确保免疫反应维持内稳态.果蝇中若缺失miR-8,则其体内的AMP将显著升高.miR-8对AMP的负调控也在小菜蛾(Plutella xyloshella)的幼虫实验中得到了证实.该研究还发现,miR-8可正向调控Serpin-27的表达,Serpin-27可抑制Toll通路的激活.当感染病原菌时,由于miR-8表达下调从而抑制Serpin-27表达,进而激活Toll通路[36].家蚕编码的miR-278-3p 可通过调控IBP2基因的表达进而调控BmCPV的增殖[37].研究表明:宿主基因组编码的microRNA既可调控宿主的基因也可作用于病毒的基因,从而可调控病毒的增殖.文献[38]的研究发现谷实夜蛾(Helicoverpa zea)Hz-miR24在感染囊泡病毒(ascovirus)晚期可下调ascovirus 病毒基因组编码的DNA依赖的RNA聚合酶及其beta亚基的转录水平.家蚕编码的bmo-miR-8可靶作用于多个家蚕核型多角体病毒(BmNPV)基因,抑制bmo-miR-8的表达可导致家蚕脂肪体中BmNPV病毒滴度减少为原来的1/8[39].蚊子细胞C6/36感染登革热病毒后,miR-252表达水平上调3倍,抑制了miR-252的表达,登革热病毒基因组RNA水平上调1.5倍[40].文献[41]的研究发现Wolbachia(一种节肢动物生殖组织内的共生细菌)感染蚊子后,可诱导在蚊子中特异表达的aae-miR-2940的表达,aae-miR-2940又可通过诱导宿主金属蛋白酶(metalloprotease)基因的表达,显著提高宿主细胞中Wolbachia的感染浓度.研究发现,aae-miR-2940也可通过诱导金属蛋白酶基因而正向调控WNV的复制[42].此外,病毒还可通过破坏宿主miRNA来抑制宿主的先天免疫,实现帮助其自身繁殖的目的[43].3.2 昆虫病毒编码的miRNA病毒编码的miRNA可靶作用于病毒基因,从而可调控病毒基因组复制或抑制宿主抗病毒应答反应.第一个经鉴定的昆虫病毒编码的miRNA是来源于棉铃虫囊泡病毒编码的HvAV-miR-1.它可下调病毒编码的DNA聚合酶进而影响病毒基因组的复制[44].家蚕核型多角体病毒(BmNPV)编码的bmnpv-miR-3在BmNPV感染早期,可靶作用于病毒DNA结合蛋白p6.9基因以及其他晚期表达基因[45],这对BmNPV的复制非常有利.首先,可自主调控BmNPV增殖,避免过度复制;其次可抑制晚期表达基因在感染早期的表达;最后可调控宿主免疫应答反应[46].苜蓿银纹夜蛾杆状病毒AcMNPV编码的AcMNPV-miR-1可靶作用于ODV-E25[47].该基因是一个晚期表达基因,编码约25 kDa的包涵体类病毒(ODV)的结构蛋白,同时也可编码出芽病毒(BV)的膜[48-49].研究发现,AcMNPV-miR-1过表达可促进ODV的形成,虽对BV病毒的复制无影响但可削弱其传染性[50].抑制AcMNPV-miR-1的表达会显著降低包涵体的数量.棉铃虫杆状病毒(Helicoverpa zea nudivirus-1,HzNV-1)在潜伏期仅表达一个非蛋白编码基因,即持续感染相关基因(persistency-associated gene 1, pag1)[51].pag1编码的2个miRNAs可降解病毒早期基因hhi1的转录本,对病毒维持潜伏期发挥重要作用.若pag1基因缺失,HzNV-1病毒将不能进入潜伏期.另一方面,研究表明,昆虫病毒的miRNA也可通过调控宿主基因的表达从而建立病毒的感染并在宿主体内成功复制.这可能是通过调节对某一特定通路起决定性作用的靶基因而导致的[52].例如,家蚕核型多角体病毒(BmNPV)编码的BmNPV-miR-1可下调核输出蛋白Exportin-5的协同因子Ran的表达,从而阻断miRNA 前体运送至细胞质,进而影响感染BmNPV细胞中成熟miRNA的表达丰度[39].再如,由WNV病毒编码的 KUN-miR-1可以正向调控蚊子细胞中编码GATA结合蛋白4基因(GATA- 4)的表达.沉默GATA- 4基因的表达可降低WNV病毒RNA的复制水平,这表明KUN-miR-1诱导表达的GATA- 4在WNV 病毒复制中发挥着重要作用[19].miRNA在昆虫—病毒互作中扮演着重要角色,其调控机制是网络化的、多层次化的.宿主编码的miRNA通过调控自身相关基因及病毒基因,从而激活免疫应答反应,抵制病毒的侵染.反之,病毒编码的miRNA靶向作用于自身基因及宿主基因,一方面提高与病毒复制相关基因的表达,另一方面抑制宿主免疫相关基因的表达,从而达到在宿主体内顺利建立感染及繁殖的目的.迄今为止,miRNA在昆虫—病毒互作机制中的研究尚属冰山一角,广泛深入地研究这一互作机制,可为揭示昆虫免疫机制提供新的研究思路,对昆虫病毒病的鉴定与防治研究具有重要的科学理论意义和实际应用价值.【相关文献】[ 1 ]SHOMRON N, GOLAN D, HORNSTEIN E. An evolutionary perspective of animal microRNA s and their targets[J].Biomed Research International, 2009(1):83-99.[ 2 ]ZHANG G, HUSSAIN M, O'NEILL S L, et al. Wolbachia uses a host microRNA to regulate tra nscripts of a methyltransferase contributing to dengue virus inhibition in Aedes aegypti[J]. Proceedings of the National Academy of Sciences, 2013, 110(25): 10276-10281.[ 3 ] LINDBO J A, SILVA-ROSALES L, PROEBSTING W M, et al. Induction of a highly specific antiviral state in transg enic plants: implications for regulation of gene expression and virus resistance[J]. Plant Ce ll, 1993, 5(5): 1749-1759.[ 4 ]WU P, HAN S, CHEN T, et al. Involvement of microRNAs in infection of silkworm with Bom byx mori cytoplasmic polyhedrosis virus (BmCPV) [J]. PLOS One, 2013, 8(7):e68209.[ 5 ]TRUJILLO R D, YUE S B, TANG Y, et al. The potential functions of primary microRNAs in tar get recognition and repression [J]. Embo Journal, 2010, 29(19): 3272-3285[ 6 ]LEE Y, AHN C, HAN J, et al. The nuclear RNase III Drosha initiates microRNA processing [J]. Nature, 2003, 425(6956): 415-419[ 7 ]DU T, ZAMORE PD. microPrimer: the biogenesis and function of microRNA [J]. Developme nt, 2005, 132(21): 4645-4652.[ 8 ]GHILDIYAL M, XU J, SEITZ H, et al. Sorting of Drosophila small silencing RNAs partitions m icroRNA* strands into the RNA interference pathway[J]. RNA, 2010, 16(1): 43-56.[ 9 ] YANG J S, SMIBERT P, WESTHOLM J O, et al. Intertwined pathways for Argonaute-mediated microRNA biogenesis in Drosophila[J]. Nucleic Acids Research, 2014, 42(3): 1987-2002.[10]OKAMURA K, HAGEN J W, DUAN H, et al. The mirtron pathway generates microRNA-class regulatory RNAs in Drosophila[J]. Cell, 2007, 130(1): 89-100.[11]BABIARZ J E, HSU R, MELTON C, et al. A role for noncanonical microRNAs in the mammali an brain revealed by phenotypic differences in Dgcr8 versus Dicer1 knockouts and small R NA sequencing[J]. RNA, 2011, 17(8): 1489-1501[12]TAFT R, GLAZOV E, LASSMANN T, et al. Small RNAs derived from snoRNAs[J]. RNA, 2009, 15(7): 1233-1240.[13]ASGARI S. Role of microRNAs in arbovirus/vector interactions[J]. Viruses, 2014, 6(9):3514-3534.[14]ROUHA H, THURNER C, MANDL C W. Functional microRNA generated from a cytoplasmic RNA virus[J]. Nucleic Acids Research, 2010, 38(22): 8328-8337.[15]SHAPIRO J S, VARBLE A, PHAM A M, et al. Noncanonical cytoplasmic processing of viral m icroRNAs[J]. RNA, 2010, 16(11): 2068-2074.[16] BENNASSER Y, LE S Y,Man L Y, et al. HIV-1 encoded candidate micro-RNAs and their cellular targets[J]. Retrovirology, 2004, 1(1):1-5.[17]ROSEWICK N, MOMONT M, DURKIN K, et al. Deep sequencing reveals abundant noncano nical retroviral microRNAs in B-cell leukemia/lymphoma[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(6): 2306-2311.[18]KINCAID RP, CHEN Y, COX J E, et al. Noncanonical microRNA (miRNA) biogenesis gives ris e to retroviral mimics of lymphoproliferative and immunosuppressive host miRNAs[J]. Mbi o, 2014,5(2):e00074-88.[19] HUSSAIN M, TORRES S, SCHNETTLER E, et al. West Nile virus encodes a microRNA-like small RNA in the 3′untranslated region which up-regulates GATA4 mRNA and facilitates virus replication in mosquito cells[J]. Nucleic Acids Research,2012, 40(5):2210-2223.[21] SHI J, DUAN Z, SUN J, et al. Identification and validation of a novel microRNA-like molecule derived from a cytoplasmic RNA virus antigenomeby bioinformatics and experimental approaches[J]. Virology Journal, 2014,11(1):1-14.[22]HELWAK A, KUDLA G, DUDNAKOVA T, et al. Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding[J]. Cell, 2013, 153(3): 654-665.[23]SHIN C, NAM J W, FARH K K, et al. Expanding the microRNA targeting code: Functional sit es with centered pairing[J]. Molecular Cell, 2010, 38(6): 789-802.[24] GREY F, TIRABASSI R, MEYERS H, et al. A viral microRNA down-regulates multiple cell cycle genes through mRNA 5′UTRs[J]. Plos Pathogens, 2010, 6(6): e 1000967.[25]FANG Z, RAJEWSKY N. The impact of miRNA target sites in coding sequences and in 3′UT Rs[J]. Plos One, 2011, 6(3): e18067.[25] CHI S W, ZANG J B, MELE A, et al. Argonaute HITS-CLIP decodes microRNA-mRNA interaction maps[J]. Nature, 2009, 460(7254): 479-486.[26] HAFNER M, LANDTHALER M, BURGER L, et al. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP[J]. Cell,2010, 141(1): 129-141.[27]BRUNO I G, KARAM R, HUANG L, et al. Identification of a microRNA that activates gene ex pression by repressing nonsense-mediated RNA decay[J].Molecular Cell, 2011, 42(4):500-510.[28] CONRAD K D, GIERING F, ERFURTH C, et al. MicroRNA-122 dependent binding of Ago2 protein to hepatitis C virus RNA is associated with enhan ced RNA stability and translation stimulation[J]. Plos One, 2013, 8(2): e56272.[29] ZHANG Y, FAN M, ZHANG X, et al. Cellular microRNAs up-regulate transcription via interaction with promoter TATA-box motifs[J]. RNA,2014, 20(12):1878-89.[30] CAMERON J E, FEWELL C, YIN Q, et al. Epstein-Barr virus growth/latency III program alters cellular microRNA expression[J]. Virology, 200 8, 382(2):257-266.[31]WANG F Z, WEBER F, CROCE C, et al. Human cytomegalovirus infection alters the expressi on of cellular microRNA species that affect its replication[J]. Journal of Virology, 2008, 82( 18): 9065-9074.[32]FULLAOND A, LEE S Y. Identification of putative miRNA involved in Drosophila melanogaster immune response[J]. Development Comparative Immunology, 2012, 36(2): 267-273.[33] THIRUGNANASAMBANTHAM K, HAIRUL-ISLAM V, SARAVANAN S, et al. Computational approach for identification of anopheles ga mbiae miRNA involved in modulation of host immune response[J]. Applied Biochemistry & Biotechnology, 2013, 170(2): 281-291.[34]ZHANG X, ZHENG Y, JAGADEESWARAN G, et al. Identification of conserved and novel mic roRNAs in Manduca sexta and their possible roles in the expression regulation of immunit y-related genes[J]. Insect Biochemistry & Molecular Biology,2014, 47(1): 12-22.[35] CHOI I K, HYUN S. Conserved microRNA miR-8 in fat body regulates innate immune homeostasis in Drosophila[J]. Developmental & Co mparative Immunology,2012, 37(1):50-54.[36] ETEBARI K, ASGARI S. Conserved microRNA miR-8 blocks activation of the Toll pathway by up-regulating Serpin 27 transcripts[J]. RNA Biology,2013,10(8):1356-1364.[37] WU P, QIN G, QIAN H, et al. Roles of miR-278-3p in IBP2 regulation and Bombyx mori cyto plasmic polyhedrosis virus replication[J]. Gen e, 2016,575(2Pt1):264-269.[38] HUSSAIN M, ASGARI S. Functional analysis of a cellular microRNA in insect host-ascovirus interaction[J]. Journal of Virology, 2010, 84(1):612-620.[39] SINGH C P, SINGH J, NAGARAJU J. A baculovirus-encoded microRNA(miRNA) suppresses its host miRNA biogenesis by regulating the Expo rtin-5 cofactor Ran[J]. Journal of Virology, 2012, 86(15): 7867-7879.[40] YAN H, ZHOU Y, LIU Y, et al. miR-252 of the Asian tiger mosquito Aedes albopictus regulates dengue virus replication by su ppressing the expression of the dengue virus envelope protein[J]. Journal of Medical Virol ogy,2014, 86(8): 1428-1436.[41]HUSSAIN M, FRENTIU F D, MOREIRA L A, et al. Wolbachia utilizes host microRNAs to mani pulate host gene expression and facilitate colonization of the dengue vector Aedes aegyp ti [J]. Proceedings of the National Academy of Sciences, 2011, 108(22): 9250-9255. [42]Slonchak A, Hussain M, Torres Morales S, et al. Expression of mosquito microRNA aae-miR-2940-5p is down-regulated in response to West Nile virus infection to restrict viral replication[J]. Journal of Virology, 2014, 88(15): 8457-8467.[43]TROBAUGH D, GARDNER C, SUN C, et al. RNA viruses can hijack vertebrate microRNAs to suppress innate immunity[J]. Nature, 2013, 506(7487): 245-248.[44] HUSSAIN M, TAFT R J, ASGARI S. An insect virus-encoded microRNA regulates viral replication[J]. Journal of Virology, 2008, 82(18): 9164-9170.[45] SINGH C P, SINGH J, NAGARAJU J. Bmnpv-miR-3 facilitates BmNPV infection 1 by modulating the 2 expression of viral P6.9 and other late genes in Bombyx mori[J]. Insect Biochemistry & Molecular Biology, 2014, 49: 59-69. [46] MORENO-HABEL D, BIGLANG-AWA I, DULCE A, et al. Inactivation of the budded virus of Autographa californica M nucle opolyhedrovirus by gloverin[J]. Journal of Invertebrate Pathology,2012, 110(1): 92-101. [47]ZHU M, WANG J, DENG R, et al. A microRNA encoded by Autographa californica nucleopo lyhedrovirus regulates expression of viral gene ODV-E25[J]. Journal of Virology, 2013, 87(23):13029-13034.[48] RUSSELL R, ROHRMANN G. A 25-kDa protein is associated with the envelopes of occluded baculovirus virions[J]. Virology, 1993, 195(2): 532-540.[49]WANG R, DENG F, HOU D, et al. Proteomics of the Autographa californica nucleopolyhedr ovirus budded virions[J]. Journal of Virology, 2010, 84(14): 7233-7242.[50]CHEN L, HU X, XIANG X, et al. Autographa californica multiple nucleopolyhedrovirus odv-e25 (Ac94) is required for budded virus infectivity and occlusion-derived virus formation[J]. Archives of Virology, 2012, 157(4):617-625.[51] WU Y L, WU C P, LIU C Y, et al. A non-coding RNA of insect HzNV-1 virus establishes latent viral infection through microRNA[J]. Scientific Reports, 2011, 1(1) : 60.[52] HUSSAIN M, ASGARI S. MicroRNAs as mediators of insect host-pathogen interactions and immunity[J]. Journal of Insect Physiology,2014, 70:151-158。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
生物工程学报Chin J Biotech 2010, August 25; 26(8): 1095−1101 Chinese Journal of Biotechnology ISSN 1000-3061 cjb@©2010 CJB, All rights reserved.DNA聚合酶δ结合蛋白38是microRNA-291a-5p的一个靶基因林德球,林辽华华南师范大学生命科学学院,广州 510631摘要: DNA聚合酶δ结合蛋白38 (DNA Polymerase delta-interacting protein 38,PDIP38) 是2003年新鉴定的一个基因,目前认为其可能在DNA修复、有丝分裂以及血管平滑肌细胞迁移中起重要作用。
根据本实验室前期在胚胎干细胞中对该基因的研究,认为microRNA可能在PDIP38的调控过程中发挥了重要作用。
为证实这种推论,运用生物信息学方法预测发现在胚胎干细胞中高表达的microRNA——microRNA-291a-5p (miR-291a-5p) 与PDIP38的开放阅读框 (ORF) 有一个配对非常理想的靶位点,通过构建该靶位点的报告基因载体以及ORF表达载体,分别进行荧光素酶报告基因分析以及细胞转染和Western blotting方法。
结果证明miR-291a-5p能够直接调节PDIP38的蛋白表达。
进一步运用real-time PCR和Western blotting分析证明了在胚胎干细胞中miR-291a-5p能够调节内源PDIP38的蛋白表达而对其mRNA表达无影响,这些都证明PDIP38确实是miR-291a-5p的一个靶基因。
关键词: microRNA,miR-291a-5p,PDIP38,胚胎干细胞DNA polymerase delta-interacting protein 38 is a target gene of microRNA-291a-5pDeqiu Lin, and Liaohua LinCollege of Life Science, South China Normal University, Guangzhou 510631, ChinaAbstract:DNA polymerase delta-interacting protein 38 (PDIP38) was identified in 2003 as a human DNA polymerase delta interacting protein which plays important roles in DNA repair, mitosis and vascular smooth muscle cells (VSMCs) migration. Our previous study showed that PDIP38 was expressed in mouse embryonic stem (ES) cells and upregulated in protein levels after differentiation from ES cells, while the expression in mRNA levels was not changed. We supposed that microRNA played key roles in the regulation of PDIP38 and the differentiation of ES cells. By bioinformatics assay, we predicted that PDIP38 was a potential target of microRNA-291a-5p (miR-291a-5p). Futhermore, we validated the possibility of miR-291a-5p to regulate the protein expression of PDIP38. Using luciferase reporter assay, realtime PCR and western blot methods, we firstly demonstrated that miR-291a-5p directly inhibited the expression of PDIP38. The present results shed a new light on the study of PDIP38 and miR-291a-5p in the differentiation of ES cells.Keywords:microRNA, miR-291a-5p, PDIP38, embryonic stem cellReceived:February 9, 2010; Accepted: May 20, 2010Supported by: National Natural Science Foundation of China (No. 30430490).Corresponding author: Deqiu Lin. Tel/Fax: +86-20-85214720; E-mail: lindq168@国家自然科学基金 (No. 30430490) 资助。
1096 ISSN1000-3061 CN11-1998/QChin J Biotech August 25, 2010 Vol.26 No.8MicroRNA (miRNA) 是一类约22 nt 的高度保守的非编码RNA 分子。
在动物中,miRNA 主要以不完全配对的方式结合到靶基因3′非翻译区 (3′UTR) 的多个互补位点,在转录后水平沉默其靶基因的表达。
在植物中,miRNA 主要以完全或几乎完全配对的方式结合到靶基因编码区的互补位点,引起mRNA 降解[1-2]。
miRNA 参与包括发育、分化、增殖、调亡和机体病变等多种生理过程,其异常表达会导致发育缺陷、癌症、肥胖、早老性痴呆等多种疾病的产生[3-6]。
和许多mRNA 类似,miRNA 的表达也具有时空特异性[3,7-8]。
近年来的研究发现,miRNA 在干细胞的自我更新、分化及其子代细胞的分化中发挥了重要的作用。
在胚胎干细胞,精原干细胞和各种体组织干细胞中,miRNA 也发挥了类似的功能[9-10]。
PDIP38是DNA 聚合酶δ结合蛋白38 (DNA Polymerase delta-interacting protein 38) 的缩写。
PDIP38是一个新鉴定的基因[11],因而目前的研究比较少。
PDIP38可以和增殖性细胞核抗原 (Proliferating cell nuclear antigen ,PCNA) 结合[11]。
PCNA 也可以和DNA 聚合酶δ结合。
在DNA 复制过程中,PCNA 募集DNA 聚合酶δ到DNA 上修复损伤的DNA ,以提高前导链的合成效率[12]。
此外,PDIP38也和线粒体内DNA 类核的一些组成蛋白结合,并且在整个有丝分裂的过程中,PDIP38都定位于有丝分裂的纺锤体[13-14]。
最新的研究则证明PDIP38在血管平滑肌细胞 (VMSCs) 迁移中发挥关键作用[15]。
在前期的研究中,我们在mRNA 和蛋白水平上分别检测了小鼠胚胎干细胞中PDIP38的表达,发现PDIP38在胚胎干细胞中有表达,同时在胚胎干细胞形成类胚体发生分化以后其蛋白表达水平显著增高,而mRNA 水平则未见有明显的变化。
根据对PDIP38的初步研究结果,我们认为很可能miRNA 在PDIP38的表达调节中发挥了重要的作用。
由此我们对PDIP38进行了miRNA 结合位点的生物信息学预测,发现在PDIP38的开放阅读框 (ORF) 末尾处有一个和microRNA-291a-5p(miR-291a-5p)配对非常理想的靶位点,而有研究者已经证实miR-291a 的基因簇成员在胚胎干细胞中高表达,并且在细胞周期调节中具有重要作用[16-17]。
根据以上的研究背景和生物信息学预测的结果,我们认为miR-291-5a 很有可能调节PDIP38蛋白的表达,进而影响胚胎干细胞的分化。
通过荧光素酶报告基因分析、real-time PCR 以及Western blotting 分析等方法我们证实了PDIP38确实是miR-291-5a 的一个靶基因。
1 材料与方法1.1 载体构建从小鼠的大脑cDNA 文库中扩增出PDIP38的961~1 860 bp 和1 101~1 860 bp 两个片段以及它的ORF 区域。
分别克隆到pMIR-REPORT 载体 (Ambion) 的萤光素酶基因后面的Xho I 位点和Sac I 位点之间以及C 端带有Flag 标签的pCDNA3.1载体的Xba I 和Sal I 位点之间。
具体如下,在50 μL 体系中加入5 μL 10× KOD Plus PCR buffer ,5 μL 2 mmol/L 的dNTPs ,2 μL 25 mmol/L MgCl 2,20 μmol/L 的正反向引物各1 μL ,cDNA 模板1 μL ,KOD Plus 1 μL (TOYOBO),用水补至50 μL 。
PCR 程序为:94℃变性2 min ;94 30 s ℃,60 30 ℃s ,72 ℃1 min ,30个循环;72 5 min ℃。
PCR 产物琼脂糖电泳后,用琼脂糖凝胶回收试剂盒 (上海生工) 回收目的条带。
对于靶位点的克隆,回收片段及pMIR-REPORT 用Xho I 和Hin d III (TaKaRa) 双酶切,37℃消化2 h ;对于ORF 的克隆,PCR 片段及载体用Xba I 和Sal I (TaKaRa) 双酶切。
酶切产物用PCR 回收试剂盒 (上海生工) 回收。
载体和片段的连接用T4 DNA 连接酶 (Fermentas) 在16℃进行。
连接产物转化到大肠杆菌DH5α感受态细胞,挑阳性克隆用PCR 进行鉴定,并送华大基因公司测序确认。
用高纯度质粒小抽试剂盒 (北京天根) 提取转染级的质粒。
扩增片段所用的引物见表1。
1.2 细胞培养及转染报告基因分析采用的细胞为293T 细胞,用含10%胎牛血清 (Invitrogen) 的高糖DMEM 培养基 (Invitrogen) 培养。
采用小鼠胚胎干细胞R1细胞系,进行Western blotting 检测内源基因表达ES 细胞培养在0.1%明胶 (Sigma) 铺盘的经丝裂霉素林德球等: DNA 聚合酶δ结合蛋白38是microRNA-291a-5p 的一个靶基因1097表1 文中所用的引物序列Table 1 The primer sequences in this studyPrimer name Primer sequence (5′−3′)target+forward GTCGAGCTCCCAGTATAGCAGCCATGT CTCTCtarget −forward GTCGAGCTCCACCCTCAGGCCTTCACT GGTAtarget+/target −reverse GACGAGCTCAGTCCAATTCTCTGGCTC CTGTORF forward GAGTCTAGAA TGGCTGGTTGTGTGGCCCG ORF reverse GAGGTCGACCTACCAGTGAAGGCCTGA GGGTPDIP38 forward GACAGTTGGTGTGTTTGAGGT PDIP38 reverse GGTAGTAAGTGTGAGTTTTGCC GAPDH forward TGA TGACA TCAAGAAGGTGGTGAAG GAPDH reverseTCCTTGGAGGCCA TGTGGGCCA T(上海生工) 处理后的单层小鼠胚胎成纤维 (Mouse embryonic fibroblasts ,MEF) 细胞上,培养液成分为含有10%胎牛血清 (Gibco)、2 mmol/L 谷氨酸 (Gibco)、0.1 mmol/L β-巯基乙醇(Gibco)、白血病抑制因子(Leukemia inhibitory factor ,LIF ,Chemicon 公司) 的DMEM 培养液,每天换液,隔天传代。