虎耳草属山羊臭组的界定和系统发育_核糖体DNAITS序列证据
会议主要日程和议程.doc
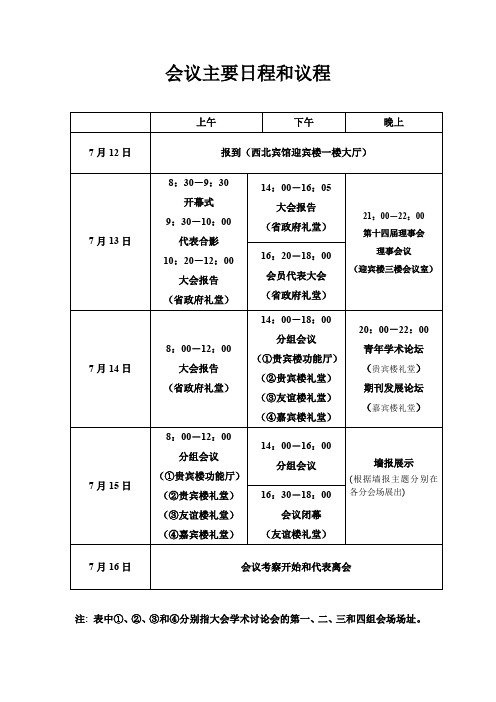
会议主要日程和议程注: 表中①、②、③和④分别指大会学术讨论会的第一、二、三和四组会场场址。
会议详细日程7月13日(星期日)上午会场:省政府礼堂(一)大会开幕式时间:8:30-9:30 主持人:武维华(二)大会特邀报告时间: 10:20-12:00 主持人: 武维华7月13日(星期日)下午会场:省政府礼堂(一)大会特邀报告(二)会员代表大会7月13日晚上(星期日)会场:迎宾楼三楼会议室时间:21:00-22:00第十四届理事会理事会议7月14日上午(星期一)会场:省政府礼堂大会特邀报告7月14日下午(星期一)分组会议1.系统与进化植物学(第一分会场)会场:贵宾楼功能厅2.植物生态与环境生物学(第二分会场)会场:贵宾楼礼堂3.植物发育与分子生物学(第三分会场)会场:友谊楼礼堂4.植物资源的可持续利用(第四分会场)会场:嘉宾楼礼堂7月14日晚上(星期一)20:00-22:00青年学术论坛会场:贵宾楼礼堂召集人:瞿礼嘉期刊发展论坛会场:嘉宾楼礼堂召集人:崔金鈡7月15日上午(星期二)分组会议1.系统与进化植物学(第一分会场)会场:贵宾楼功能厅2.植物生态与环境生物学(第二分会场)会场:贵宾楼礼堂3.植物发育与分子生物学(第三分会场)会场:友谊楼礼堂4.植物资源的可持续利用(第四分会场)会场: 嘉宾楼礼堂7月15日(星期二)下午(一)分组会议时间:14:00—16:00 1.系统与进化植物学(第一分会场)会场:贵宾楼功能厅2.植物生态与环境生物学(第二分会场)会场:贵宾楼礼堂3.植物发育与分子生物学(第三分会场)会场:友谊楼礼堂4.植物资源的可持续利用(第四分会场)会场:嘉宾楼礼堂(二)大会闭幕式时间:16:30-18:00 主持人:马克平会场:友谊楼礼堂7月16日(星期四)会后考察和代表离会。
【国家自然科学基金】_高山植物_基金支持热词逐年推荐_【万方软件创新助手】_20140731

科研热词 推荐指数 高山植物 3 生物量 3 高山红景天 2 高山林线 2 青藏高原 2 放牧 2 叶绿体dna 2 优势种 2 黄河源区 1 高山嵩草(kobresia pygmaea)草甸1 高寒山地 1 避难所 1 退化 1 连续扦插继代 1 运动行为 1 覆被变化 1 西藏八角莲 1 虎耳草属山羊臭组 1 藓类植物 1 草甸 1 花向日性 1 色季拉山 1 群落组成 1 群落多样性 1 红波罗花 1 繁育适应 1 繁殖特性 1 系统发育 1 空间动态 1 稳定碳同位素比值 1 种子萌发 1 种子大小 1 神经酰胺 1 碳、氮储量 1 界定 1 生理生态过程 1 生活型演变 1 生活型 1 生态生物学特性 1 生态特征 1 珍珠黄杨 1 物种组成 1 灌丛草甸 1 濒危植物 1 温度升高 1 温室气体通量 1 江河源区 1 水分利用效率 1 水分供应 1 气候变化 1 横断山区 1 横断山 1
107 108 109 110 111 112
soyasapogenol b rps16 rapd its序列 co2浓度升高 cca
1 1 1 1 1 1
107 108 109 110 111 112 113 114 115 116
传粉效率 交配系统 九龙山组 丽江 中侏罗统 世界遗产 twinspan cca cbmapk3基因 cbf/dreb1转录因子
科研热词 高山红景天 青藏高原 高山离子芥 融雪梯度 生物多样性 温度日较差 海拔梯度 新疆 土壤含水量 古生态环境 光合作用 五台山 义县组 高山雪场 高山草甸 高山苔原 高山绣线菊 高山流石滩 高山植物根 高山杓兰 高山、亚高山草甸 高寒草甸 马先蒿属 非生物胁迫 青海云杉 降维对应分析 长白山 通气组织 逆境适应性 退化草地 解剖 表达谱 表土花粉 菌根结构 菌丝结 自然遗产地 自交亲和性 继代次数 结构鉴定 红膜处理 红波罗花 红河流域 红景天苷产量 红景天甙 竹种资源 空间格局 种组 祁连圆柏 石羊河上游 短壳缝目 生长季长度 生长
【国家自然科学基金】_内转录间隔区(its)_基金支持热词逐年推荐_【万方软件创新助手】_20140801

推荐指数 3 3 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32
科研热词 序列分析 内转录间隔区(its) 种系发育关系 物种鉴定 内转录间隔区 its序列 黄鳝 香蕉枯萎病菌 铁皮石斛 鉴定 车前草白粉菌 车前草 蛇钩口线虫 蛇蛔虫 膜荚黄芪 胃瘤线虫 羌活 系统发育分析 稳定性 白粉菌 白粉寄生孢 生物学特性 单孢分离 分子系统学鉴定 凤仙花 准确性 亲缘关系 rdna-its序列 rdna pcr its/its2 its
推荐指数 3 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45
9种柴胡属植物的核糖体ITS序列及其在药材鉴定中的应用

9种柴胡属植物的核糖体ITS序列及其在药材鉴定中的应用谢晖;晁志;霍克克;吴炳义;潘胜利【期刊名称】《南方医科大学学报》【年(卷),期】2006(26)10【摘要】目的研究9种柴胡属药用植物rDNAITS序列的差异和规律,寻找可准确鉴定柴胡类药材的分子性状.方法 PCR扩增ITS序列,测序,DNAssist vetsion 2.2软件进行序列比对,并计算同源性.结果柴胡属植物ITS序列与外类群ITS序列比对同源性均小于75%,序列间两两比对同源性均大于88%,同种植物则大于99%.结论ITS序列作为柴胡类药材鉴定依据具有一定实用性和可靠性.【总页数】4页(P1460-1463)【作者】谢晖;晁志;霍克克;吴炳义;潘胜利【作者单位】复旦大学药学院生药教研室,上海,200321;南方医科大学中药学院,广东,广州,510515;复旦大学生命科学研究院,上海,200433;南方医科大学南方医院临床实验研究中心,广东,广州,510515;复旦大学药学院生药教研室,上海,200321【正文语种】中文【中图分类】R284【相关文献】1.胡椒属药材核糖体DNA的ITS序列分析及其分子鉴定 [J], 蔡诚诚;杨志业;谢晖;潘胜利2.基于2个核糖体DNA序列的国产白酒草属、小舌菊属和歧伞菊属(菊科紫菀族)分子系统学研究 [J], 钟彩霞;黎维平;杨秀林;唐明;廖威;陈三茂;3.基于2个核糖体DNA序列的国产白酒草属、小舌菊属和歧伞菊属(菊科紫菀族)分子系统学研究 [J], 钟彩霞;黎维平;杨秀林;唐明;廖威;陈三茂4.基于核糖体DNA的ITS序列和叶绿体trnT-trnL及trnL-trnF基因间区的菊花起源与中国菊属植物分子系统学研究(英文) [J], 赵惠恩;汪小全;陈俊愉;洪德元5.线粒体nad 1内含子2序列在石斛属植物分子鉴定中的应用(英文) [J], 张婷;王峥涛;徐珞珊;周开亚因版权原因,仅展示原文概要,查看原文内容请购买。
【国家自然科学基金】_nrdna_基金支持热词逐年推荐_【万方软件创新助手】_20140729

推荐指数 4 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
科研热词 系统发育 its 重组 贝叶斯分析 西川红景天 葡萄座腔菌属 白地霉 生物降解 润楠属 毡球壳属 樟科 杂交起源 普氏腔菌属 披碱草属 序列分析 小麦族 大聚颈腔菌属 地霉属 变叶海棠 双色孢腔菌属 分类鉴定 分离筛选 分化 trns-trng序列 sth基因组 rpl20-rps12序列 its序列
推荐指数 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
2011年 科研热词 推荐指数 遗传多样性 1 遗传分化 1 资源冷杉 1 系统发育 1 种间杂种 1 石耳目 1 石生真菌 1 真藓科 1 洋葱 1 大葱 1 地衣 1 内生菌 1 丝瓜藓属:ptychostomum 1 nrdna its序列 1 mtdna 1 its 1 gapc基因内含子 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
科研热词 its 系统发育 nrdna 虎耳草属山羊臭组 蓼科 花被片形态 花粉形态 翼蓼属 盐桦 界定 测序 桦木属 果实解剖 杂草稻 大血藤 地带性分化 单叶血藤 pcr扩增条件 its序列
推荐指数 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14
【国家自然科学基金】_近缘属_基金支持热词逐年推荐_【万方软件创新助手】_20140803

2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39
科研热词 推荐指数 菊属 2 聚类 2 系统进化 2 甘蔗 2 预处理 1 遗传多样性 1 遗传分析 1 通用性 1 近缘种属植物 1 解剖结构 1 菊花 1 荧光原位杂交 1 耐盐性 1 目标起始密码子多态性(scot) 1 盐害指数 1 特征 1 滇小檗 1 杂种鉴定 1 新组合 1 新名称 1 斑茅 1 数量性状 1 拟单性木兰 1 小檗属 1 定性多态性状 1 多倍体起源 1 含笑属 1 叶绿体转化 1 叶绿体基因组 1 叶 1 变性 1 十大功劳属 1 光萼荷属 1 体系优化 1 亲缘关系 1 ssr标记 1 rna编辑 1 leafy基因 1 dus测试 1
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
2011年 科研热词 近缘属 柑橘属 新纪录 dna条形码 襀翅目 编码序列 珠尾叉襀属 河南 新种 寄蝇科 噪寄蝇属 叉襀总科 中国 rbcl matk 推荐指数 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1
2014年 序号 1 2 3 4 5
2014年 科研热词 染色体数目 染色体形态 广义石山苣苔属 广义报春苣苔属 分类处理 推荐指数 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43
舶来赏析NewPhytologist:小麦-山羊草物种的染色体重建

舶来赏析NewPhytologist:小麦-山羊草物种的染色体重建近期,俄罗斯科学院瓦维洛夫遗传研究所Dr. Ekaterina D. Badaeva和瑞士波恩大学的Dr. Christian Parisod在New Phytologist 发表了综述论文:Chromosome restructuring among hybridizing wild wheats.小麦-山羊草复合群(Aegilops L. and Triticum L.)包含一大批自然分布于地中海和中亚地区的一年生二倍体和多倍体物种。
比较分析发现,圆锥小麦(Triticum turgidum, BBAA)和普通小麦(Triticum aestivum, BBAADD)也是在这些地区,从其祖先物种Ae. speltoides (genome SS=BB)、T.urartu (AA)和Ae. tauschii (DD)天然杂交和多倍体化演化而来。
由于这些野生物种的染色体基数为x=7,所以,在分子细胞遗传学上,将其称为小麦-山羊草复合群。
该复合群中物种中,其基因组超过80%由转座子序列组成(图1)。
这些物种的染色体是如何从其野生二倍体物种杂交、演化、基因组重建而来,其遗传机制值得深入探讨。
图1. 小麦-山羊草复合群染色体的序列组成A不同转座子序列的分布, B染色体臂的典型序列组成1、二倍体物种的网状进化小麦A, B, D基因组的祖先大约在4-5百万年前开始发生分化(图2)。
最近研究认为,Ae. mutica可能比Ae. speltoides更接近小麦B 染色体组的原始供体。
D染色体组的供体约在3百万年前加入了现代山羊草属多倍体物种的形成。
目前D组二倍体代表物种为Ae. tauschii,S基因组代表性物种为Ae. bicornis、Ae. longissima、Ae. searsii、Ae. sharonensis。
山羊草CUMN基因组的二倍体物种包括Ae. caudata (CC), Ae. umbellulata(UU), Ae. comosa(MM) 和Ae. uniaristata(NN)。
【国家自然科学基金】_dna分类_基金支持热词逐年推荐_【万方软件创新助手】_20140802
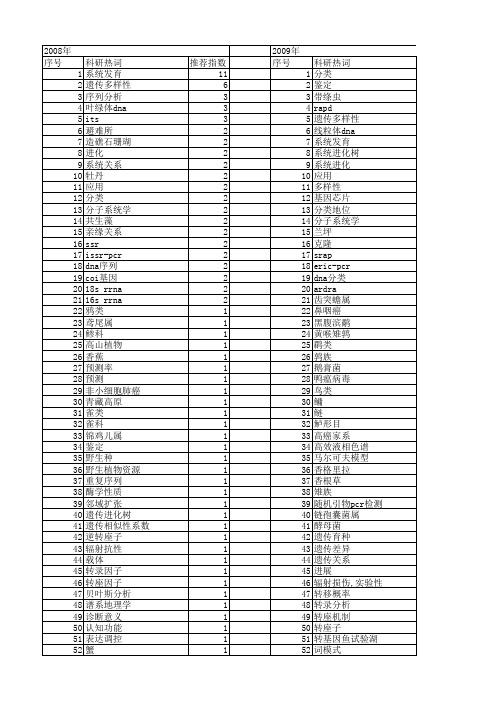
科研热词 分类 鉴定 带绦虫 rapd 遗传多样性 线粒体dna 系统发育 系统进化树 系统进化 应用 多样性 基因芯片 分类地位 分子系统学 兰坪 克隆 srap eric-pcr dna分类 ardra 齿突蟾属 鼻咽癌 黑腹滨鹬 黄喉雉鹑 鹛类 鹑族 鹅膏菌 鸭瘟病毒 鸟类 鳙 鲢 鲈形目 高癌家系 高效液相色谱 马尔可夫模型 香格里拉 香根草 雉族 随机引物pcr检测 链孢囊菌属 酵母菌 遗传育种 遗传差异 遗传关系 进展 辐射损伤,实验性 转移概率 转录分析 转座机制 转座子 转基因鱼试验湖 词模式
角蟾科 表达谱 表达序列标签 表型性状 补充描述 血雉 蛋白质结构 蛋白编码基因 蚂蚁 虫株 蔷薇科 蔷薇属 菌种资源 菌物 荧光定量pcr 茶树 自动显微图像处理 脂肪酸 肿瘤诊断 肠含物 肌阵挛-站立不能性癫痫 聚类分析 联合固氮菌 群体遗传关系 细菌多样性 细菌 细胞色素b基因 细粒棘球绦虫 线粒体细胞色素酶c亚基i基因 线粒体细胞色素b基因 纳米材料 红斑狼疮,系统性 红喉雉鹑 系统发育分析 空肠 稻属 种 盾蚧亚科 相互作用蛋白 皮脂蠕形螨 白灵菇 白地霉 白令海 瘤胃细菌 电活性小分子 电压门控性钠通道α 1亚基基因 电化学dna生物传感器 甲基萘醌 生物降解 生物多样性 甘薯 玉蜀黍属 玉米 猪
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
科研热词 系统发育 遗传多样性 序列分析 叶绿体dna its 避难所 造礁石珊瑚 进化 系统关系 牡丹 应用 分类 分子系统学 共生藻 亲缘关系 ssr issr-pcr dna序列 coi基因 18s rrna 16s rrna 鸦类 鸢尾属 鲹科 高山植物 香蕉 预测率 预测 非小细胞肺癌 青藏高原 雀类 雀科 锦鸡儿属 鉴定 野生种 野生植物资源 重复序列 酶学性质 邻域扩张 遗传进化树 遗传相似性系数 逆转座子 辐射抗性 载体 转录因子 转座因子 贝叶斯分析 谱系地理学 诊断意义 认知功能 表达调控 蟹
[doc]睡莲科的系统发育:核糖体DNAITS区序列证据
![[doc]睡莲科的系统发育:核糖体DNAITS区序列证据](https://img.taocdn.com/s3/m/9b8680ca32d4b14e852458fb770bf78a65293a77.png)
睡莲科的系统发育:核糖体DNAITS区序列证据植物分类43(1):22—30(2005)ActaPhytotaxonomicaSinica睡莲科的系统发育:核糖体DNAITS区序列证据刘艳玲徐立铭倪学明赵家荣(中国科学院武汉植物园武汉430074) PhylogenyoftheNymphaeaceaeinferredfromITSsequencesLIUYan?LingXULi-MingNIXue?MingZHA0Jia?R0ng (WuhanBotanicalGarden,theChineseAcademyofSciences,Wuhan43007 4,China) AbstractTheNymphaeaceaeareoneofthemostimportanttaxaforunderstand ingthe originandevolutionofprimitiveangiospermsduetoitsbasalpositioninthecla dogramsofthe angiospermsrecentlygivenbasedbothonseveralgenesequencesandonmorp hologicalcharacters,butphylogeneticrelationshipswithinthefamilyhavenotasyetbee nresolved.ThesequencesofnrDNAITSregionofllspeciesrepresentingsevengeneraoftheN ymphaeaceaeandoneoutgroup,Ceratophyllumdemersum,wereusedtoreconstructthephy logenyofthefamilyusingPAUP4.0b4A.Threemostparsimonioustrees(Length=l125,CI =O.7618and尺,-0.7214)wereobtained.Intheconsensustree:(1)Nelumbowasbasaltotheot hergenerawithabootstrapvalueoflo0%andoccupiedanisolatedposition.SOitcouldbe separated fromtheNymphaeaceaeandplacedinitsownfamily,Nelumbonaceae,andits ownorder,Nelumbonales;(2)Nupharwasmonophyletic,nestedinthebasalpositionofc ladeIIandwas stronglysupportedtobethesistergroupoftheothergenera(excludingNelumb o)oftheNymphaeaceae,suggestingthatitstraditionalinclusionintheNymphaeacea eshouldbemaintained;(3)CabombaandBraseniaweresistergroupandformedasubclad ewithabootstrapvalueof99%,indicatingtheircloseaffinity;(4)Nymphaeawasstronglysupported tObethesistergroupofasubcladecomprisingEuryaleandVictoriawithaboot strapvalueof94%.indicatingthattheyshouldbeplacedintheNymphaeaceae. KeywordsNymphaeaceae.ITS,phylogeny.摘要以金鱼藻Ceratoph),llumdemersum为外类群,使用PAUP4.0b4A 软件对睡莲科Nymphaeaceae植物7属11个代表种的ITS区序列进行了系统发育分析.采用最大简约法分析获得了39最简约树,步长为1125,一致性指数(c,)和维持性指数D值分别为0.7618和0.7214.利用3个最简约树获取严格一致树.结果表明:(1)莲属Nelumbo位于系统树的基部,自展支持率为100%,可从睡莲科中独立出来成立莲科Nelumbonaceae和莲目Nelumbonales;(2)萍蓬草属Nuphar是一单系类群,位于分支II的基部,并和睡莲科其他属(不包括莲属)植物聚在一起构成姐妹群,故萍蓬草属仍应置于睡莲科中;(3)水盾草属Cabomba和莼菜属Brasenia聚成一小支并构成姐妹群,自展支持率为99%,说明这两属之间亲缘关系较近;(4)睡莲属和芡实属Euryale,王莲属Victoria聚成一小支并构成姐妹群,自展支持率为94%,说明三者亲缘关系较近,仍应置于睡莲科中.关键词睡莲科;ITS;系统发育睡莲科Nymphaeaceae有9属约100种,广布于温带与热带地区(唐先华等,2003).随着新归化属植物——水盾草CabombacarolinianaA.Gray在浙江杭州,宁波,鄞县等地的发现2004.05.17收稿,2004.09.20收修改稿.通讯作者(Authorforcorrespondence.E—mail:**********************).1期刘艳玲等:睡莲科的系统发育:核糖体DNAITS区序列证据—(丁炳扬,2000),睡莲科在中国的分布或栽培已有7属:睡莲属NymphaeaL.,莲属NelumboAdans.,萍蓬草属NupharJ.E.Smith,芡实属EuryaleSalisb.exDC.,王莲属VictoriaLind1.,水盾草属CabombaAub1.,莼菜属BraseniaSchreb..另外不见于我国的两属为OndineaHartog(Hartog,1970;Helen,1977)和BarclayaWal1.(Bentham&Hooker,1862).形态和分子证据表明睡莲科是被子植物谱系的最早分支之--(Barkmaneta1.,2000;Soltiseta1..2000),也是原始双子叶植物中的一个重要类群,隶属于毛茛目,又兼有单子叶植物的若干性状,对探讨被子植物系统演化及单,双子叶植物起源等问题具有重大意义,因而倍受关注.国内外学者已从形态学(Osbom&Schneider,1988;Moseleyeta1.,1993;倪学明等,1994;Leseta1.,1997),解剖学(陈维培,张四美,1992;Moseleyeta1.,1993;Schneidereta1.,1995;Schneider&Carlquist,1996a,b;Hayeseta1.,2000;胡光万等,2003),孢粉学(Walker,1974,1976;张玉龙,1984;Osbometa1.,1991),染色体数及核型(韦平和等,1993,1994),酯酶同功酶(陆佩洪,陈维培,1993;陆佩洪等,1994)等方面做了大量的工作,但并未取得一致意见.有的学者主张作为一科处理,即睡莲科(Bentham&Hooker,1862;Caspary,1891);或两科处理,即莲科Nelumbonaceae或Cabombaceae和睡莲科(Hutchinson,1959;Les,1991);或三科处理,即Cabombaceae,睡莲科和莲科(Takhtajan,1980;Cronquist,1988;Williamson,1993a,b;Les,1999);还有的主张增加一萍蓬草科Nupharaceae(Takhtajan,1997)作为四科处理等不同观点.在基于大量分子资料而建立的APG(TheAngiospermPhylogenyGroup)系统(汤彦承等,1999)中,把莲科置于山龙眼目Proteales下,取消了睡莲目Nymphaeales,并把Cabombaceae合并到睡莲科中.而吴征镒等(2oo3)则支持将莲科升为莲目Nelumbonales隶属于毛茛纲,而睡莲目仍置于木兰纲,并调整为4个科:莼菜科Hydropeltidaceae,竹节水松科Cabombaceae,睡莲科和芡科Euryalaceae.有关睡莲科科内的分子系统发育及属间谱系分支研究较少.Les等(1991)以单基因rbcL 序列构建了睡莲科植物的分子系统发育树,支持将莲属独立出来成立莲科,隶属于莲目.利用rbcL,matK和18SrDNA序列,并结合重要的形态学性状重建的睡莲科植物(不包括莲属)系统发育树,又支持将水盾草属和莼菜属独立出来成立Cabombaceae(Leseta1.,1999),仍置于睡莲目.唐先华等(2003)仅获得了睡莲科4属植物的ITS序列,初步探讨]~ITS序列在睡莲科植物系统发育分析中的应用.核糖体DNA中的内转录间隔区(ITS)序列由于可以提供较丰富的信息位点,已被广泛用于研究被子植物属内种间,近缘属间乃至科内的系统发育和分类问题(Baldwineta1.,1995;刘忠等,2000;田欣,李德铢,2002).本文通过对睡莲科7属11种植物及外类群金鱼藻Ceratoph~,llumdemersumL.的nrDNAITS序列分析,构建了睡莲科植物的ITS系统发育树.并在此基础上结合一些重要的形态性状,探讨了科内的谱系发生关系,同时也试图为睡莲科植物的分类学位置研究提供新的证据.1材料和方法1.1材料实验材料包括睡莲科植物7属11种及外类群金鱼藻,其来源及基因库样品序号见表1.表1材料来源Table1o啦jnofmaterials植物分类43卷WBGCAS,WuhanBotanicalGarden,theChineseAcademyofSciences. 1.2总DNA提取,PCR扩增和DNA测序取上述植物的新鲜叶片约0.1g,采用改良的CTAB法(Doyle&Doyle,1987)提取总DNA.聚合酶链式反应(PCR)在MJ”PTC一225”型PCR仪上进行,用White等(1990)描述的一对通用引物ITS4和ITS5进行ITS区的扩增(ITS4序列:5’TCCTCCGCTTA TFGA TA TGC3’,ITS5序列:5’GGAAGTAAAAGTCGTAACAAGG3’).PCR反应程序为:94℃预变性4min,94℃变性lmin,50℃退火lmin,72℃延伸2min,30个循环:扩增产物经回收纯化后克隆~I|pMD18一T载体上并在含有X—Gal,IPTG和Amp的L一琼脂平板培养基上培养.挑选出阳性克隆送上海生工生物工程技术有限公司用ABI377型自动测序仪进行序列测定,测序引物为M13+/M13一.为了保证测序的准确性,对ITS序列正,反链进行测定并加以校准.1.3序列分析和系统树的构建本文测定的序列经ClustalX(Thompsoneta1.,1997)软件排序,个别位点做了必要的人工校正,以金鱼藻为外类群,用PAUP4.0b4A(Swofford,1998)软件进行系统发育分析.空位(gap)被编码为缺失(missing).最大简约性分析采用如下选项完成,即:树二组重新连接1期刘艳玲等:睡莲科的系统发育:核籍体DNAITS区序列证据25 (TBR),启发式搜索(Heuristicsearch),多重性选择(MUL TREESoption),ACCTRAN优化和100次随机附加的重复.利用自h~]0T(bootstral,,1Ooo次重复)(Fe1senstein,1985)检验各分支的置信度.2结果和分析如表2所示:本文测得的11种睡莲科植物的ITS1序列长度范围为257—274bp,GC百分含量为40.83%一63.12%,ITS2为244—274bp,GC—分含量为44.12%一63.26%,5.8SrDNA为156bp(水盾草插入两个碱基,5.8SrDNA为158b)),整个ITS区的长度范围为660—704bp.当空位作缺失处理时,ITS区全序列排序后的长度为73O位点,其中有114个变异位点,68个为系统发育的信息位点,分别占15.62%和9.32%.奇约性分析得到3个最简约树(MPTs),其步长为1125,一致性指数(CD和维持性指数(种分别为0.7618和O.7214. 表211种睡莲科植物的ITS长度和G+C含量Table2ThesizeandG+CcontentofITSofthe11speciesoftteNrrS1ITS2利用3个最简约树获取的严格一致树如图1尹亍示.睡莲科7属11个物种分别聚成两支,且分别获得100%的自展支持率.分支I为莲属的2个种,位于系统发育树的基部(自展支持率为100%).睡莲科的其他6属聚成一支,即分支Ⅱ,这一支又包括2个亚支,其中萍蓬草属的2个代表种以100%的自展支持率聚在一起,并位于相对基部的位置;另外一个亚支又分成2个小支.其中睡莲属,王莲属,芡实属和莼菜属水盾草属分别聚成一小支并构成姐妹群,且获得了较高的白展支持率(94%和99%),但这./J,支聚在一起的支持强度却不高(61%).3讨论3.1莲属的分类等级及其系统位置莲属由于具有离生心皮,无导管木质部,双珠被,厚珠心的倒生胚珠,八核蓼型胚囊,叶片状雄蕊,层状胎座等原始特征而被置于睡蹇科(韦平和等,1994).近年来,许多学者根据莲属独有的植物化学,形态学和细胞学特征?Takhtajan,1969;韦平和等,1994;倪学明植物分类43卷a厂—丽6l_一6694_一6899_——69l0086l00738049l0053454785443 CladeI\79l9l0l1NymphaeacoeruleaNymphaeacapensisEuryaleroxVictoriacruzianaCabombapiauhyensisCabombacarolinianaBraseniaschreberINupharbornetiiNupharsinensisNelumboluteaNelumboI’lUC’ireraCeratophyllumdemersum图1基于rrs序列分析得到的3个最简约树的严格一致树(步长为1125,一致性指数(c,)o.7618,维持性指数(肋为0.7214).分支下面的数值代表支长变化,分支上面的数值代表自展支持率.蟹?1?ThestrictconseⅡsust]’eeofthreeequallymostparsimonious仃ecsbasedOnITSsequenceswithallcharactersequallyweighted(I.~ngth=l125,CI=O.7618,.7214).Numbersbelowbranch esarebasesubstitutions;thoseabovebranchesarebootstrapvalues(%)with1000repficates.等,1994),支持莲属从睡莲科中独立出来成立莲科和莲目:莲属含苄基异喹啉类生物碱,果实具呼吸孔,种子单粒,无胚乳,两个子叶基部愈合,莲属叶片具排水器和厚壁异细胞,上下两表皮有气孔,属单唇型发育(Guptaeta1.,1968);莲属花粉为三沟花粉,外壁具明显的柱状层结构;睡莲科中的分类群是由古多倍体起源的,而只有莲属的染色体基数x=8,可推断为古二倍体(韦平和等,1993),同时它的染色体形态也显着不同于睡莲科其他类群,集中表现在莲属具有一对大的近端着丝点染色体,第2号染色体的一条长臂上存在明显的次缢痕以及可能出现的B染色体.基于cL基因(Leseta1.,1991)和本文ITS序列的分析表明:莲属是一单系类群,位于分支图的最基部(自展支持率为100%),应为睡莲科的原始类群(图1).鉴于莲属独有的形态,细胞学特征及来自cL,ITS序列的分子证据,作者支持将它从睡莲科中独立出来成立莲科,隶属于莲目(Leseta1.,1991;Li,1955).3.2萍蓬草属的分类等级及其系统位置萍蓬草属广布北半球的温带地区.形态解剖学的观察结果(Ito,1987;Moseleyeta1.,1993;Philbrick&Les,1996)和基于cL基因的序列分析(Leseta1.,1991)都支持萍蓬草属的传统分类地位,即隶属于睡莲科(Caspary,1891;Cronquist,1988).但最近的研究表明:萍蓬草属具有一些与睡莲科其他类群明显不同的特征(施国新等,1998;胡光万等,2003;Moseley,1965;Padgetteta1.,1999),有分离出来独立成科的可能性.这些特征主要包括:(1)该属是睡莲科中惟一没有热带种类的一个属;(2)果实不沉入水中成熟;(3)子房上位;(4)远轴花瓣上分布有蜜腺;(5)花粉无沟或槽,饰孔多刺;(6)花梗中仅具一个星状排列的维管束;(7)叶柄基本组织中无气道;(8)根顶端原始细胞具有分层特征,属封闭型.陆佩洪等(1994)通过对睡莲科叶片中酯酶同功酶的测定也发现:萍蓬草属具有3’4条独有的酶谱,显示出与其他属不同的特征.1期刘艳玲等:睡莲科的系统发育:核糖体DNAITS区序列证据萍蓬草属与睡莲科其他类群之间有着明显的差异,但它们在具根状茎,叶二型,花两性,辐射对称,单生花梗顶端,萼片,花瓣均为3至多数,雄蕊6至多数,纵裂,倒生胚珠1至多数等方面是基本一致的(倪学明等,1994).Les等(1999)基于rbcL,marK和18SrDNA的基因序列分析,并结合一些重要的形态性状,探讨了睡莲目植物的系统发生关系.其中萍蓬草属隶属于睡莲科(不包括莲属,莼菜属和水盾草属),位于睡莲科植物系统发育树的基部,并和Barclaya属保持较近的亲缘关系.本文的ITS序列分析结果也显示:萍蓬草属是一单系类群,位于分支Ⅱ(图1,CladeⅡ)的基部,并和睡莲科其他属(不包括莲属)植物以100%的自展支持率聚在一起构成姐妹群,故我们认为萍蓬草属应置于睡莲科中,保持其传统的分类地位(Caspary,1891;Cronquist,1988).3.3水盾草属和莼菜属的分类等级及其系统位置水盾草属主要分布在新大陆的热带和温带地区,莼菜属集中分布在北纬4O.以南的亚热带地区(于丹,1991),但两者在形态,花部,营养器官的解剖特征及孢粉学等方面有一些睡莲科其他属植物不具有的共征(胡光万等,2003;Ito,1986;Osbometa1.,1991):(1)除具有根状茎外,还具有沉水茎;(2)二者均为背生胎座;(3)花托维管束由带状束顺着花瓣,雄蕊形成分支的花托维管束组成(倪学明等,1994);(4)单沟花粉;(5)花丝短而背腹扁平;(6)叶柄中只有一个双韧维管束,位于叶柄的中央;(7)叶柄和叶肉中无星状石细胞和草酸钙结晶.基于这些共征,许多学者都赞同将水盾草属和莼菜属共同置于一新科——Cabombaceae中(Hutchinson,1959;Osborneta1.,1991;Schneider&Carlquist,1996;Le seta1.,1999).并根据其具下垂倒生,两层珠被,厚珠心的胚珠和维管束中有原生木质部空腔等事实,将Cabombaceae仍置于睡莲目(Takhtaj8.I1,1987;Cronquist.1988).ITS序列的分析结果表明(图1):水盾草属和莼菜属聚成一小支,并以99%的自展支持率构成姐妹群,说明这两属之间亲缘关系较近,聚在一起是十分自然的,这和来自rbcL,mark和18SrDNA基因序列的分子证据(Leseta1.,1999)是相吻合的.同时水盾草属,莼菜属又和王莲属,芡实属,睡莲属以61%的自展支持率聚在一起构成姐妹群,说明它们之间存在一定的亲缘关系,故我们支持将水盾草属,莼菜属置于睡莲科中.3.4睡莲科其他属的分类等级及其系统位置芡实属和王莲属植物叶片硕大,叶柄,花梗及果实有刺,两者不仅在外部形态上极为相似,而且在其他方面还有许多共同特征:二者都有雄蕊瓣化现象,柱头辐射排列,下位子房与花托结合在一起,环槽花粉等(Swindells,1983;倪学明等,1994).对过氧化物酶酶谱的类平均聚类分析也显示王莲属与芡实属有较近的亲缘关系(陆佩洪,陈维培,1994).来自rbcL,marK,18SrDNA(Leseta1.,1991,1999)及本文的ITS序列分析都表明:两者亲缘关系较近,并以较高的自展支持率聚在一起构成姐妹群.陈维培等(1992)通过对睡莲科植物叶片的比较解剖发现:睡莲属在气孔器,腺毛,吸水器,星状石细胞,草酸钙结晶及维管束结构和排列方式,染色体形态上和芡实属,王莲属更为接近(韦平和等,1994).ITS序列分析也表明:睡莲属和王莲属,芡实属聚成--’b支,并得到了较高的自展支持率(94%)(图1).基于形态,比较解剖学等方面的共征以及ITS序列的分子证据,我们认为芡实属,王莲属和睡莲属三者亲缘关系较近,且染色体核型均为2A类型(韦平和等,1994),仍应置于睡莲科中.28植物分类43卷睡莲科植物不同属之间虽存在着较大的差异,但在盾叶的形式和发育进程,维管束的结构和排列方式,只有管胞而无导管,双层珠被,子叶2片,上胚轴较短而下胚轴模糊等方面是基本一致的,这说明它们之间并非仅是由于水生环境而被归在一起的,其间确实存在一定的亲缘关系.致谢感谢浙江大学丁炳扬教授,姜维梅教授在水盾草的采集,华中农业大学重点实验室石磊博士,陈银华硕士在实验过程及武汉大学郭友好教授,王建波教授,陈进明博士在序列分析中给予的大力支持与帮助.参考文献BaldwinBG,SandersonMJ,PorterJM,WojciechowskiMF,CampbellCS,DonoghueMJ.1995,TheITSregionofnuclearribosomalDNA:avaluablesourceofevidenceonangiosper mphylogeny.AnnalsoftheMissouriBotanicalGarden82:247—277.BarkmanTJ,CheneryG,McNealJR,Lyons—WeilerJ,EllisensWJ,MooreG, WolfeAD.2000. Independentandcombinedanalysesofsequencesfromallthreegenomiccom partmentsconvergeontherootoffloweringplantphylogeny.Proceedingsoftl1eNationalAcademyofS ciencesUSA97:13166—13171.BenthalnG,HookerJD.1862.GeneraPlantarum.London:Reeve&Co. 1:45—48.CasparyR.1891.Nymphaeaceae.In:EnglerA,PrantlKAEeds.DieNatnrlich enPflanzenfamilien.Leipzig:V erlagvonWilhelmEngelmann.2:1—10.ChenW-P(陈维培),ZhangS-M(张四美)parativeleafanatomyofNymphaeaceae(s.1.).Acta PhytotaxonomicaSinica(植物分类)30:415—422.CronquistA.1988.Theevolutionandclassificationoffloweringplants.2nde d.NewY ork:TheNewY orkBotanicalGarden.DingB—Y仃炳扬).2000.CabombaAublet.(Cabombaceae),anewlynaturalizedgenusofChin a.ActaPhytotaxonomicaSinica(植物分类)38:198—2o0.DoyrleJJ.DoyleJL.1987.ArapidDNAisolationproceduref0rsmallquantifi esoffreshleaftissue.PhytochemicalBulletinl9:l1—15.FelsensteinJ.1985.Confidencelimitsonphylogenies:anapproachusingtheb ootstrap.Evolution39:783—791.GuptaSC,蹦iwalGS,AhuiaR.1968.ThestomataofNelumbonucirera:formation,distributi onanddegeneration.AmericanJoumalofBotany55:295—301.HartogCD.1970.Ondinea,anewgenusofNymphaeaceae.Blumea18:413—417.HayesV.SchneiderEL.CarlquistS.2000.RoralDevelopmentofNelumbonu cifera(Nelumbonaceae).InternationalJournalofPlantSciencel61:183—191.HelenIA.1977.AquaticPIantsofAustralia,Carlton:MelbourneUniversityP ress.139.HuG—w(胡光万),LiuK-M(刘克明),LeiL—G(雷立公)parativesmdyonleafanatomyofthree generainNymphaeaceae.LifeScienceResearch(生命科学研究)7:243—248.HutchinsonJ.1959.TheFamiliesofRoweringPlants.Oxford:Claredon.403—407.ItoM.1986.StudiesinthefloralmorphologyandanatomyofNymphaeales.I Ⅱ.RoralanatomyofBraseniaschreberiGme1.andCabombacarolinianaA.Gray.TheBotanicalMagazine (Tokyo)99:169—184.ItoM.1987.PhylogeneticsystematicsoftheNymphaeales.TheBotanicalMa gazine(Tokvo11o0:17—35.LesDH.GarvinDK.WimpeeCF.1991.Molecularevolufionaryhistoryofanc ientaquaticangiosperms. ProceedingsoftheNationalAcademyofSciencesUSA88:10119—10123. LesDH,SchneiderEL,PadgeaDL.1997.Waterlilyrelationshipsrevisited:le ssonslearnedfromanatomy.morphologyandmolecules.TheWaterGardenJoumal13:2l一28. LesDH,SchneiderEL,PadgeaDJ.1999.Phylogeny,classificationandfloral evolutionofwaterlilies(Nymphaeales):Asynthesisofnon—molecular,rbcLmatKand185rDNAda ta.SystematicBotany24:28—46.1期刘艳玲等:睡莲科的系统发育:核糖体DNAITS区序列证据LiH.L.1955.ClassificationandphylogenyofNymphaeaceaeandalliedfami Hes.AmericanMidlandNaturalist54:33—41.LiuZ(刘忠),WangX.Q(汪小全),ChenZ-D(陈之端),LinQ(林祁)’LuA-M(路安民).2000.111e phylogenyofSchisandraceaeinferredfromsequenceanalysisofthertrDNAI TSregion.ActaBotanicaSinica(植物)42:758—761.LuP-H(陆佩洪),ChenW.P(陈维培).1993.StudyontheperoxidaseisozymesofNymphaeaceaewith analysisofpolarordination.JournalofNanjingNormalUniversityfNaturalS cience)(南京师范大学(自然科学版))16(2):52—56.LuP.H(陆佩洪),ChenW.P(陈维培),XuH.X(许红星),LiuM(刘淼).1994.Studyontheesterase isozymesofNymphaeaceaewithnumericaltaxonomy.JournalofNanjingNo rmalUniversity(NaturalScience)(南京师范大学(自然科学版))l7(3):92—97. MoseleyMF.1965.MorphologicalstudiesofNymphaeaceae.III.Theflorala natomyofNuphar.Phytomorphology15:54—84.MoseleyMF,SchneiderEL,WilliamsonPS.1993.Morphologicalstudiesoft heNymphaeaceaesensulatoXIX.Phylogeneticinterpretationsfromselectedfloralvasculaturecharacter sintheNymphaeaceaesensulato.AquaticBotany44:325—342.Nix-M(倪学明),Y uB(於炳),ZhouY.J(周远捷),ZhaoJ-R(赵家荣).1994.Studiesonthephylogenetic relationshipsamongtheNymphaeaceae.JournalofWuhanBotanicalResear ch(武汉植物学研究)12:3ll一320.OsbornJ.SchneiderEL.1988.MorphologicalstudiesoftheNymphaeaceae XVI.ThefloralbiologyofBraseniaschreberi.AnnalsoftheMissouriBotanicalGarden75:778~794. OsbornJM,TaylorTN,SchneiderEL.1991.Pollenmorphologyandultrasllu ctureoftheCabombaceae:correlationswithpollinationbiology.AmericanJoumalofBotany78:1367一l378.PadgettDJ,LesDH,CrowGE.1999,PhylogeneticrelationshipsinNuphar(N ymphaeaceae):Evidencefrommorphology,chloroplastDNA,andnuclearribosomalDNA.American JournalofBotany86:l316—1324.PhilbrickCT,LesDH.1996.Evolutionofaquaticangiospermreproductivesy stems.BioScience46:813—826.SchneiderEL,CarlquistS.1996a.V esselsinNelumbo(Nelumbonaceae).A mericanJournalofBotany83:l101一ll06.SchneiderEL.CarlquistS.1996b.V esselsinBrasenia(Cabombaceae):newp erspecfivesonvesselorigininprimaryxylemofangiosperms.AmericanJournalofBotany83:l236一l240. SchneiderEL,CarlquistS,BeamerK,KohnA.1995.V esselsinNymphaeacea e:Nuphar’Nymphaea,andOndinea.InternationalJournalofPIantSciences156:857—862.ShiG?x(施国新),XieK?B(解凯彬),ChangF.C(常福辰),DingX.Y(丁小余).1998.Morphologicaland anatomicalobservationsofvegetativeorgaJlofNupharpumilum(Whimrn. DC.JournalofPlantResourcesandEnvironment(植物资源与环境)7:43-48.SoltisDE,SoltisPS,ChaseMW,MortME,AlbachDC,ZanisM,SavolainenV, HaimWH,HootSB,FayMF,AxtellM,SwensenSM,PrinceLM,NixonKC,FarrisJS.2000.Angiospe rmphylogenyinferredfroml8SrDNA,fL,andatpBsequences.BotanicalJournaloftheLirmeanSoci ety133:38l-461.SwindellsP.1983.Waterlilies.Portland,Oregon:TimberPress.20—21. SwoffordDL.1998.PAUP木4.O:Phylogeneficanalysisusingparsimonyf 木andothermethods).Betaversion4.0b4A.Sunderland:SinauerAssociates.tajanAL.1969.FloweringPlants:OriginandDispersa1.Edinburgh:Oliver& amp;Boyd.207.TakhtajanAL.1980.Outlineoftheclassificationoffloweringplants(Magnol iophyta).BotanicalReview46:225—359.Ta~tajanAL.1987.SystemaMagnoliophytorum.Leninopli:Nauka. TakhtajanAL.1997.DiversityandClassificationofFloweringPlants.NewY ork:ColumbiaUniversityPress.TangX?H(唐先华),ZhangX?Y(张晓艳),SlliS.H(施苏华),ZhongY(钟扬),Laix.L(赖旭龙).2003.A preliminaryphylogeneticanalysisofITSnrDNAsequencesinwater.1ilies.E arthScience---JournalofChinaUniversityofGeosciences(地球科学一一中国地质大学)28:97~101.TangY-C(汤彦承),LuA?M(路安民),ChenZ.D(陈之端).1999.Abriefintroductiontoanewordinalclassificationofthefloweringplants.ActaPhytotaxonomicaSinica(植物分类)37:608—621.植物分类43卷ThompsonJD,GbsonTJ,PlewinakF,JeanmouginF,HigginsDG.1997.Thecl ustal-Xwindowsinterface: flexiblestrategiesformultiplesequencealignmentaidedbyqualityanalysist ools.NucleicAcidsResearch25:4876—4882.TianX(田欣),LiD-z(李德铢).2002.AppficationofDNAsequencesinplantphylogeneticstudy.Acta BotanicaYunnanica(云南植物研究)24:l70一l84.WalkerJW.1974.ApertureevolutioninthepoHenofprimitiveangiosperms. AmericanJournalofBotany61:lll2一ll36.parativepoHenmorphology。
会议主要日程和议程
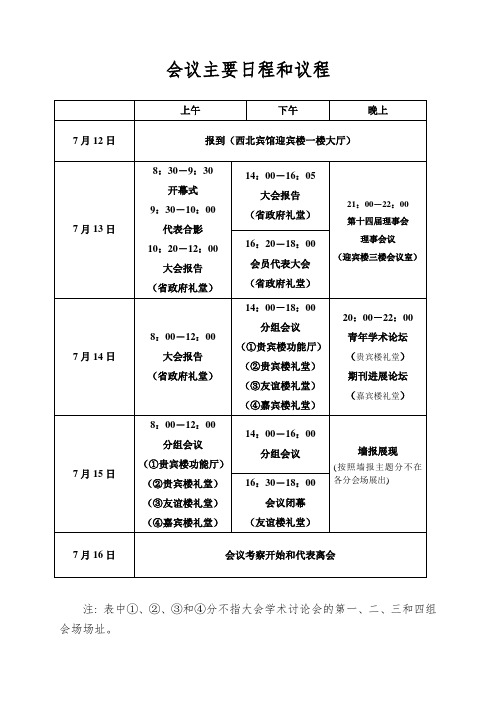
会议主要日程和议程
注: 表中①、②、③和④分不指大会学术讨论会的第一、二、三和四组会场场址。
会议详细日程
7月13日(星期日)上午会场:省政府礼堂
(一)大会开幕式
(二)大会特邀报告
时刻: 10:20-12:00 主持人: 武维华
7月13日(星期日)下午会场:省政府礼堂
(一)大会特邀报告
(二)会员代表大会
7月13日晚上(星期日)会场:迎宾楼三楼会议室
时刻:21:00-22:00 第十四届理事会理事会议7月14日上午(星期一)会场:省政府礼堂
大会特邀报告
7月14日下午(星期一)分组会议
1.系统与进化植物学(第一分会场)会场: 贵宾楼功能厅
7月14日晚上(星期一)20:00-22:00
7月15日上午(星期二)分组会议
7月15日(星期二)下午
(一)分组会议时刻:14:00—16:00
2.植物生态与环境生物学(第二分会场)会场: 贵宾楼礼堂
(二)大会闭幕式时刻:16:30-18:00
7月16日(星期四)会后考察和代表离会。
核糖体DNAITS区应用于虫草属无性型鉴定的初步研究[1]
![核糖体DNAITS区应用于虫草属无性型鉴定的初步研究[1]](https://img.taocdn.com/s3/m/64afb01759eef8c75fbfb3dd.png)
收稿日期:2002209230作者简介:凌建亚(1975-),男,2000级博士生,从事霉菌生物学研究.3通讯作者 文章编号:167129352(2003)0320117206核糖体DNA ITS 区应用于虫草属无性型鉴定的初步研究凌建亚 彭俊峰 张晗星 张长铠3(山东大学 微生物技术国家重点实验室,山东 济南 250100)摘要:虫草无性型鉴定是困扰学术界的一个难题.本文检索了互联网上报道的虫草及可能的无性型序列,应用相关软件构建进化树,研究了它们的同源性.从总体上看,核糖体DNA ITS 区序列研究与传统经典分类学相互印证进行虫草无性型鉴定是完全可行和必要的.关键词:虫草;无性型;ITS 区;同源性中国分类号:Q939.96 文献标识码:APreliminary study of identification to anam orph of Cordycepsby using its sequences of nrDNALI NGJian 2ya ,PE NGJun 2feng ,ZH ANG Han 2xing &ZH ANG Chang 2kai3(S tate K ey Laboratory of M icrobial T echnology ,Shandong University ,Jinan 250100,Shandong ,China )Abstract :The con figuration of anam orph of C ordyceps has the academic and practical importance.Phylogenetic Relationship am ong the genus Cordyceps and its probable anam orph were examined by using ITS region sequences of nrDNA.The data consisted of 25sequences for 11taxa were aligned by Clustal X ,a free multiple sequence alignment s oftware ,analyzed by neighbor 2joining (N J )tree ,and valued by Bootstraping method.The hom ology results indicated that the sequence difference within ITS region could provide an additional characteristic for identification to anam orph of the genus Cordyceps .K ey w ords :Cordycep ;anam orph ;ITS region ;hom ology 虫草属是一类虫生真菌,依据1996年Alex opou 2los 的分类,属于子囊菌纲(Ascomycetes ),麦角菌目(Clavicipitales ),麦角菌科(Clavicipitaceae ).迄今发现约有350余种[1],仅清水大典1994年编著的《原色冬虫夏草图鉴》就已记载了316种,我国现有记载70余种[2].由于该属大多数种采集困难,寄主范围广泛,世代交替的生活史有无性型和有性型阶段,加之传统真菌分类学圄于技术手段的限制,往往造成对虫草结构发育和生理片段资料的不同理解,使得虫草属种的鉴定和分类非常困难,尤其是无性型的鉴定一直是困扰学术界的难题.以冬虫夏草为例,争论十余年,先后报道的相关无性型菌株多达10属16种.随着分子生物学的迅速发展,找寻传统形态、生理、生化等经典分类指标之外的基因序列鉴定模式成为解决困难的重要途径之一.核糖体RNA (rRNA )是细胞中最古老的分子之一,具有结构、功能和进化的同源性.编码其rRNA 前体的基因即rDNA ,是基因组DNA 中中等重复并有转录活性的基因家族.位于18s -28s rDNA 之间的内转录间隔区(Internal Transcribed Sequence ,ITS 第38卷 第3期 V ol.38 N o.3 山 东 大 学 学 报 (理 学 版)JOURNA L OF SH ANDONG UNI VERSITY 2003年8月 Aug.2003 区),它包括进化上高度保守的5.8s rDNA和包含有rDNA前体加工信息的两个间隔区ITS21,ITS22.在rDNA多基因家族成员中,ITS区是多拷贝,以串连重复方式出现在一个或多个染色体基因位点上的,同时ITS区又有通过不等交换和基因转变等方式经历快速一致进化的特性,使得重复单位在基因组内保持了高度的一致性;而且ITS区两侧有高度保守序列存在,white等[3]于1990年设计了一整套真核生物通用扩增引物,所有这些特性有利于PCR扩增、测序和排序,而所得到的序列潜在信息变异可以有效地解决系统发育关系.近年来ITS区已成为真菌分子生物学研究中的热点.我们试图从G enbank数据库中现有的虫草相应ITS区序列着手,对其进行排序,构建了进化树,比较虫草及报道无性型相似性,探讨了其同源性.通过核糖体DNA ITS区序列研究阐明了分子生物学手段验证虫草属无性型的可能性.1 材料与方法1.1 实验材料截止2002年7月,G enbank上共有C ordyceps的序列552条,检索得到部分虫草及其报道的可能无性型ITS区数据共11种,25条(见表1).表1 所选虫草的序列号及其ITS区长度T able1 Sequence number of the selected C ordyceps sp and the length of ITS region虫草序列号ITS区长度(bp)Mariannaea pruinosa A J309337494C.pruinosa A J309338495C.pruinosa AB044635501C.hawkesii A J309341570 Paecilomyces hawkesii A J309342570C.tricentri A J309364392C.tricentri AB027376434 Paecilomyces sp.G97013A J309362469C.gunnii A J309340567 Paecilomyces gunnii A J309343569C.brittlebankoides A J309332537 Mearhizium anisopliae A J309333537C.takaomontana AB044637496 Paecilomyces tenuipes AB027380498C.emeiensis A J309347501 G liocladium sp G97013A J309346483C.sobolifera A J309351518Beauveria sobolifera A J309325518C.brongniartii A J309348483C.brongniartii AB027381480Beauveria brongniartii A J309349483itaris AF153266481Paecilomyces militaris A J309329481C.sinensis A J309356488Hir sutella sinebsus A J3093574901.2 方法利用基于渐近比对思想设计的Clustal X软件包将所选中的序列进行排序,序列间插入适当的空位以达到最大的同源性;使用Phylip96分析软件包中DNAdist程序建立距离矩阵;应用邻近-连接法(Neighbour2Joining method,N J)聚类;利用Clustal X 软件构建进化树比较其序列同源性;Bootstraping法进行检验,重复1000次;应用ClustalX附带N J P LOT win95软件显示进化树.2 结果与讨论2.1 典型ITS区序列的选择有关虫草的研究多年来主要集中于冬虫夏草(Cordyceps sinensis)和蛹虫草(Cordyceps militaris),相关的ITS序列报道分别多达18条和14条.为简化计算,同时更重要的是考虑到取样代表性的问题,我们事先用N J法对其序列进行了比较.通过相似性分析选取了其中的典型序列参与构建进化树;同样的处理分别应用于其它有较多条相关序列报道的古尼虫草(C.gunnii)、茧草(C.pruinosa)、蝉花虫草(C. sobolifera)、布氏虫草(C.brongniartii). 如图1所示,冬虫夏草所有ITS报道序列中, A J243777,AF291749与其它序列矩阵距离差异值非常大.即使考虑到扩增片段以及测序长度等因素的影响,也远远超过了一般意义上种及种间的ITS区序列的差异值(3%),其为冬虫夏草序列报道的可信度尚待进一步验证.因此我们选取A J309357与A J309356为冬虫夏草及无性型的代表序列.目前大多数所报道的蛹虫草ITS区序列高度一致,仅有微小差别;仅AB070373有较大差异(见图2),其为蛹虫草的ITS区报道可信性亦值得怀疑.我们选择了有代表性的AF153266与A J309329为蛹虫草及可能无性型的典型序列.同样,我们分析了其他有较多条ITS区序列报道的虫草(结果待报),适当选取了相应序列进行后续工作. 118 山 东 大 学 学 报 (理 学 版)第38卷 图1 冬虫夏草及其可能无性型ITS 区序列的树状图Fig.1 Ev olutionary tree of related ITS region sequences ofC ordyceps sinensis and its probable anam orph图2 蛹虫草及其可能无性型ITS 区序列的树状图Fig.2 Ev olutionary tree of related ITS region sequences ofCordyceps militaris and its probable anam orph2.2 虫草及相应报道无性型的距离矩阵(见表2)表2 虫草及其可能无性型的矩阵距离T able 2 Matrices length of C ordyceps and the probableanam orph12345678Hir sutella sinensis A J3093560.00000.00000.10960.22280.22220.33320.33220.3226C .sinensis A J3093570.00000.09700.20850.20790.32540.32540.3164C .emeiensis A J3093470.00000.23820.23750.35100.35100.3348Beauveriasobolifera A J3093250.00000.00190.43870.43620.4093C .sobolifera A J3093510.00000.44210.43960.4125Mearhiziumanisopliae A J3093330.00000.00000.2025C .brittlebankoidesA J3093320.00000.2029C .gunnii A J3093430.0000Paecilomyces hawkesiiA J309342C .hawkesii A J309341Paecilomyces gunnii A J309340Paecilomyces militarisA J309329C .militaris AF153266C .brongniartii A J309348Beauveria brongniartiiA J309349C .brongniartii AB027381Mariannaea pruinosaA J309337C .pruinosa A J309338Paecilomyces tenuipes AB027380C .takaomontana AB044637C .pruinosa AB044635G liocladium sp .G 97013A J309346C .tricentri A J309364C .tricentri AB027376Paecilomyces sp .G 97013A J309362910111213141516Hir sutella sinensis A J3093560.32550.33320.32460.29980.29110.33200.33200.3259C .sinensis A J3093570.31940.32710.31840.28900.28900.32230.32230.3161C .emeiensis A J3093470.33770.34870.34020.30950.29190.34130.34130.3322Beauveria soboliferaA J3093250.40890.42050.41210.36450.34630.39090.39090.3745C .sobolifera A J3093510.41210.42380.41530.36340.34530.38970.38970.3734Mearhiziumanisopliae A J3093330.20490.20800.20010.29680.28200.30340.30340.2914C .brittlebankoides A J3093320.20530.20850.20050.29680.28200.30340.30340.2914 第3期凌建亚,等:核糖体DNA ITS 区应用于虫草属无性型鉴定的初步研究119 910111213141516C.gunnii A J3093430.00530.00890.02530.22950.21580.25730.25730.2344 Paecilomyces hawkesii A J3093420.00000.01060.02710.23210.21840.26000.26000.2371C.hawkesii A J3093410.00000.03090.23540.22170.26340.26340.2403 Paecilomyces gunnii A J3093400.00000.22470.21100.25250.25250.2297 Paecilomyces militaris A J3093290.00000.00000.08990.08990.0883itaris AF1532660.00000.09200.09200.0897C.brongniartii A J3093480.00000.00000.0147 Beauveria brongniartii A J3093490.00000.0147C.brongniartii AB0273810.0000 Mariannaea pruinosa A J309337C.pruinosa A J309338Paecilomyces tenuipes AB027380C.takaomontana AB044637C.pruinosa AB044635G liocladium sp.G97013A J309346C.tricentri A J309364C.tricentri AB027376Paecilomyces sp.G97013A J309362171819202122232425 Hir sutella sinensis A J3093560.28980.28470.32990.33650.29960.33130.53580.57450.6328C.sinensis A J3093570.27580.27080.31680.32670.29300.32900.53580.57450.6154C.emeiensis A J3093470.31720.31190.32740.35070.32640.33980.48080.53940.5889 Beauveria sobolifera A J3093250.36720.36460.38500.40410.36090.39550.48280.52400.6697C.sobolifera A J3093510.36610.36350.38390.40290.36340.39430.48720.52810.6672 Mearhiziumanisopliae A J3093330.30930.30080.35620.36030.30250.34970.56620.57900.5658C.brittlebankoides A J3093320.30930.30080.35620.36030.30250.34970.56620.57900.5658C.gunnii A J3093430.29060.28280.30960.30100.25640.30040.51720.55050.5162 Paecilomyces hawkesii A J3093420.29340.28560.31240.30380.25900.30330.51080.54580.5200C.hawkesii A J3093410.29700.28910.31610.30740.26230.30390.52370.54580.5242 Paecilomyces gunnii A J3093400.29770.28710.32060.31100.26700.30490.52680.55980.5033 Paecilomyces militaris A J3093290.10620.09820.12890.12140.12000.32140.52010.53280.5006itaris AF1532660.10600.09790.12850.12350.11920.31060.52010.53280.4907C.brongniartii A J3093480.10070.09540.11640.12160.10990.29410.50460.53890.5049 Beauveria brongniartii A J3093490.10070.09540.11640.12160.10990.29410.50460.53890.5049C.brongniartii AB0273810.09670.09140.11410.11680.10520.27470.49470.52950.4891 Mariannaea pruinosa A J3093370.00000.00610.12690.11700.10780.30990.50810.54440.5131C.pruinosa A J3093380.00000.12130.10920.10250.30450.51260.52980.5089 Paecilomyces tenuipes AB0273800.00000.02460.11200.34030.53030.55180.5370C.takaom ontana AB0446370.00000.11220.33420.53500.55620.5280C.pruinosa AB0446350.00000.30910.51620.54280.5329 G liocladium sp.G97013A J3093460.00000.43760.45050.4562C.tricentri A J3093640.00000.05190.3832C.tricentri AB0273760.00000.4339 Paecilomyces sp.G97013A J3093620.00002.3 虫草及其相应报道无性型的树状图如图3所示,序列号间连线长度代表序列两两比较中产生的相似性得分所换算成的分化距离,亦即表1中所示矩阵值;线上数字代表Bootstraping法评估进化树的评估值.由图中可知:冬虫夏草A J309357与中华被毛孢(Hir sutella sinensis) A J309356;蛹虫草AF153266与蛹草拟青霉(Paecilo2 myces militaris)A J309329;布氏虫草AB027381、 120 山 东 大 学 学 报 (理 学 版)第38卷 A J309348与布氏白僵菌(Beauveria brongniartii )A J309349;高雄虫草(Cordyceps .takaomontana )AB044637与细脚拟青霉(Paecilomyces tenuipes )AB027380;蝉花虫草A J309351与Beauveria sobolifera A J309325;Cordyceps brittlebankoides A J309322与Mear 2hizium anisopliae A J309333Bootstraping 法评估重复1000次均聚在一起,且两两矩阵距离小于0.03(0.0246),亦即序列之间差异小于3%,甚至有的序列相似性高达100%,显示了二者间ITS 区序列的高度同源性.这是确证以上虫草无性型的有力证据,与经典分类的结果完全吻合.图3 虫草及其相应可能的无性型ITS 区序列的树状图Fig.3 Ev olutionary tree of ITS region sequences of Cordyceps sp and its probable anam orph 茧草A J309337与粉被马利娅霉(Paecilomyces te 2nuipes )A J309338的ITS 序列差异仅有0.61%,但与另一报道茧草AB044635相差甚大,分别为10.52%和10.78%.一般认为,生物样品序列间rDNA 相似值若低于90%,通常作为不同种或不同属处理[4],但具体到真菌种间差异亦有一般大于14%的报道[5].鉴于茧草及其可能无性型报道的ITS 区序列仅此3条,我们尚无法予以定论,该虫草无性型的确证有待进一步工作的验证.吹沫虫草(Cordyceps tricentri )曾有无性型A J309362的报道,但经我们验证,不同报道来源的该虫草序列AB027376与A J309364之间仅相差5.19%,而分别与该假定无性型相差38.32%和43.39%,提示A J309362可能并不是相应虫草的正确无性型.所报道的Cordyceps emeiensis 虫草仅有两个相关序列A J309347、A J309346,其矩阵距离差异值达到33.98%.二者就序列长度而言仅相差18bp (见表1),却有如此高的序列差异值,显示了这两个序列相似性之低,我们有理由相信A J309346不是该虫草的正确无性型.古尼虫草分布于贵州,最早作为国内新纪录种报道[6];亚香棒虫草(Cordyceps hawkesii )分布于湖北、湖南、广西,传统分类对二者子囊、子囊孢子、次生子囊孢子、分生孢子等特征描述有差别[7].古尼虫草及Paecilomyces gunnii 分别与亚香棒虫草、Paecilo 2myces hawkesii 四条序列两两间差异最大仅有3.09%,如此密切的亲缘关系,我们认为可能的解释是:1)古尼虫草与亚香棒虫草为近源种,可能rDNA 基因序列特别是ITS 区序列有着高度的同源性;2)古尼虫草寄主是蝙蝠蛾科Hepiaidae 幼虫;亚香棒虫草寄主是鳞翅目幼虫[7],亦有报道为蝙蝠蛾科新属新种-湖南棒蝠蛾[8~9].有可能是同一虫草无性型菌着生于不同寄主,对不同地理环境适应的结果表现为虫草表观特征的不同;3)同一虫草(或无性型)不同生长发育阶段的表观特征细微差别造成了同物异名现象.3 总结早期的ITS -1和ITS -2系统发育分析比较了潜在信息位点占全序列位点的百分率,发现ITS 两 第3期凌建亚,等:核糖体DNA ITS 区应用于虫草属无性型鉴定的初步研究121 个间隔序列之间变异的分布和根据趋异值的估计是近似的.B.G.Baldwin等A[10]认为,ITS-1和ITS-2两套资料进化上是相似的,具有互补性,是两套同质的(hom ogeneous)性状序列,因此根据两套资料得到的系统树要比根据其中任何一个间隔序列资料得到系统树分辨率高,内部支持强度大.我们采信了以上的观点,同时考察高保守的5.8srDNA序列,将整个ITS区序列在虫草属无性型鉴定中应用.结果表明,测定ITS区序列进行虫草无性型鉴定,并与传统分类相互印证是完全可行和必要的,ITS序列测定法在分类学研究中具有潜在价值.但是,鉴于目前虫草属的研究现状,前期工作必须严谨、扎实,传统分类是一切工作的基础.对于分子生物学数据不能简单的予以采信,不宜贸然否定传统的鉴定分类,毕竟,宏观的形态进化与微观的分子进化并不一定同步.同时,ITS区序列可以被用来开展虫草无性型相关半知菌属的工作,解决诸如蛹虫草无性型先后被归于头孢霉属、轮枝霉属和拟青霉属,至今尚有争论的问题,为经典分类奠定坚实的基础.核糖体DNA作为分子系统学的热点受到分类学家的广泛关注.分子系统学着眼于从分子水平为地球各级生物分类鉴定提供客观依据,是分子生物学与分类学相互渗透的结果.核糖体ITS区适用于种及种间的分类阶元.但是我们应该注意到,毕竟ITS区序列较短,用于分类系统学研究的性状数量是有限的.开展虫草菌资源普查工作,广泛的收集虫草品种,继续应用分子生物学手段进行虫草菌的鉴定工作,特别是长序列18s、28s核糖体基因的测定,而且必要的条件下可以选取高保守蛋白基因序列进行研究,在大量积累分子数据的基础上构建虫草属完整的系统进化树.这是虫草属无性型鉴定进一步工作的方向.参考文献:[1]肖宏亮,高孔荣,谭盈科.冬虫夏草及其菌丝体研究进展[J].中国食用菌,1997,16(2):3~5.[2]李增智,黄勃,李春如,等.确证冬虫夏草无性型的分子生物学证据I.中国被毛孢与冬虫夏草之间的关系[J].菌物系统,2000,19(1):60~64.[3]White T J,Bruns T,T ee S,et al.Applification and direct se2quencing of fungal ribos omal RNA genes for phylogenetics[M].In:M A Innis,DH G elfand,JJ Sninsky,et al(eds).PCR Protocols:A G uide to Methods and Applification.San Dieg o.C A:Academic Press,Inc.1990,315~322.[4]葛颂,Schaal B A,洪德元.用核糖体DNA的ITS序列探讨裂叶沙参的系统位置[J].植物分类学报,1997,35(5):385~395.[5]余仲东,张星耀,曹支敏.真菌核糖体基因间隔区研究概况[J].西北林学院学报,2000,15(2):107~112. [6]梁宗琦.一种国内未见报道的虫草菌-古尼虫草[J].真菌学报,1983,2(4):258~259.[7]卯晓岚.中国大型真菌[M].郑州:河南科学技术出版社,2000.[8]吴德龙,王卫芳,薛芳森,等.亚香棒虫草的研究[J].江西农业大学学报,1997,19(2):31~36.[9]朱弘复,王林瑶.冬虫夏草与蝙蝠蛾[J].动物学集刊,1985,(3):30.[10]B.G.Baldwin,Michael J,Sanders on J.et al.,The ITS re2gion of nuclear ribos omal DNA:A valuable s ource of evidence on angiosperm phylogeny[J].Ann Miss ouri Bot G ard.1995, 82:247~277.(编辑:于善清) 122 山 东 大 学 学 报 (理 学 版)第38卷 。
【国家自然科学基金】_球茎_基金支持热词逐年推荐_【万方软件创新助手】_20140801
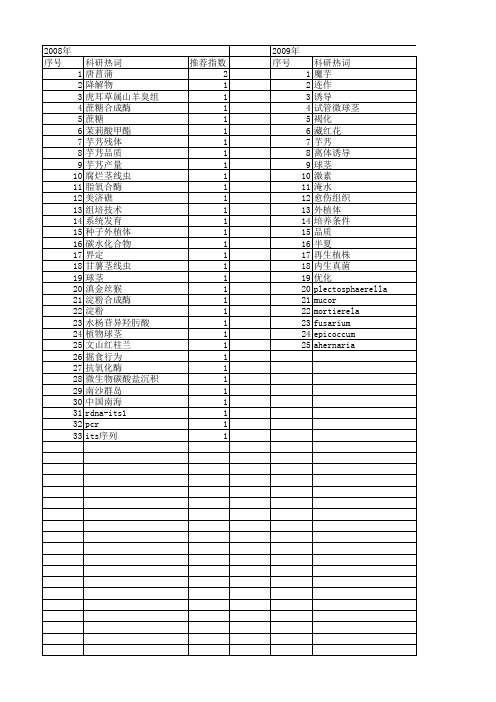
推荐指数 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
科研热词 青天葵 遗传变异 谢君魔芋 萝卜硫素 组织培养 系统进化分析 类原球茎 福青1号青花菜 生物多样性 狗蔷薇 烟台海域 毛状不定根 显微观察 改良catb法 抗污染 嗜盐菌 品比 发育过程1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7
2011年 科研热词 香蕉 锯齿状腺瘤 绒毛管状腺瘤 纤维绒毛锯齿状腺瘤 氮磷钾 异型增生 分布 推荐指数 1 1 1 1 1 1 1
2012年 序号 1 2 3 4 5 6 7 8 9 10 11
科研热词 香蕉 绿色荧光蛋白标记 生理小种 尖孢镰刀菌古巴专化型 定殖 侵染 预测模型 贮藏期 荸荠腐烂病 花部特征 繁育系统 紫背天葵 盐胁迫 生理特性 环境因素 濒危原因 杂交指数 总生物碱 半夏 一氧化氮(no)
推荐指数 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33
科研热词 唐菖蒲 降解物 虎耳草属山羊臭组 蔗糖合成酶 蔗糖 茉莉酸甲酯 芋艿残体 芋艿品质 芋艿产量 腐烂茎线虫 脂氧合酶 美济礁 组培技术 系统发育 种子外植体 碳水化合物 界定 甘薯茎线虫 球茎 滇金丝猴 淀粉合成酶 淀粉 水杨苷异羟肟酸 植物球茎 文山红柱兰 掘食行为 抗氧化酶 微生物碳酸盐沉积 南沙群岛 中国南海 rdna-its1 pcr its序列
赖草属物种的its序列变异及其系统推断

赖草属物种的its序列变异及其系统推断赖草属(Poaceae)是禾本科的一个大属,包括了众多具有重要经济价值和生态意义的物种。
ITS序列(Internal Transcribed Spacer)是植物核糖体DNA(rDNA)中的一个关键区域,具有种间和种内变异。
研究赖草属物种的ITS序列变异对于了解其系统发育关系和分类具有重要意义。
目前已经有很多研究对赖草属物种的ITS序列进行了分析。
这些研究主要集中在以下几个方面:1. 序列变异分析:通过对不同物种的ITS序列进行PCR扩增和测序,发现赖草属物种间的ITS序列存在一定的变异。
这些变异可用于鉴别不同物种,并为物种的分类和系统发育提供依据。
2. 分子系统学分析:基于ITS序列的分子系统学研究揭示了赖草属物种的进化关系。
例如,研究发现赖草属可分为若干个分支,而这些分支之间的亲缘关系与形态学、生态学等特征密切相关。
3. 遗传多样性分析:通过对ITS序列变异的研究,可以了解赖草属物种的遗传多样性。
这对于种质资源保护和遗传改良具有重要意义。
4. 物种鉴定:ITS序列变异可用于物种鉴定,尤其在赖草属这类形态特征相近的物种中,ITS序列分析具有较高的准确性。
然而,ITS序列研究也存在一些局限性。
例如,由于序列长度较短,ITS序列变异有时不足以明确划分物种。
此外,不同物种间的序列变异程度也可能存在差异,导致部分物种难以区分。
因此,在研究赖草属物种的ITS序列变异时,需结合其他分子标记和形态学等证据,以更准确地揭示其系统发育关系。
总之,赖草属物种的ITS序列变异及其系统推断研究对于理解该属物种的进化关系、分类和种质资源保护具有重要意义。
然而,还需进一步探讨其他分子标记和多种数据来源的整合,以提高研究准确性。
【国家自然科学基金】_高山地区_基金支持热词逐年推荐_【万方软件创新助手】_20140801
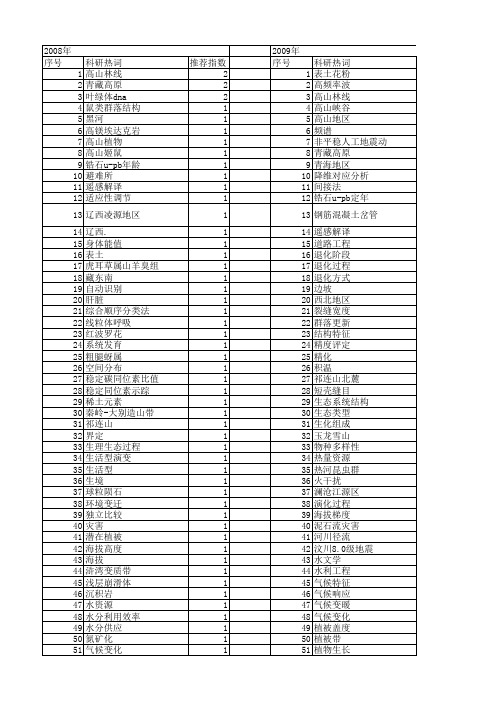
科研热词 表土花粉 高频率波 高山林线 高山峡谷 高山地区 频谱 非平稳人工地震动 青藏高原 青海地区 降维对应分析 间接法 锆石u-pb定年 钢筋混凝土岔管 遥感解译 道路工程 退化阶段 退化过程 退化方式 边坡 西北地区 裂缝宽度 群落更新 结构特征 精度评定 精化 积温 祁连山北麓 短壳缝目 生态系统结构 生态类型 生化组成 玉龙雪山 物种多样性 热量资源 热河昆虫群 火干扰 澜沧江源区 演化过程 海拔梯度 泥石流灾害 河川径流 汶川8.0级地震 水文学 水利工程 气候特征 气候响应 气候变暖 气候变化 植被盖度 植被带 植物生长 植物区系
推荐指数 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
科研热词 高山林线 青藏高原 叶绿体dna 鼠类群落结构 黑河 高镁埃达克岩 高山植物 高山姬鼠 锆石u-pb年龄 避难所 遥感解译 适应性调节 辽西凌源地区 辽西. 身体能值 表土 虎耳草属山羊臭组 藏东南 自动识别 肝脏 综合顺序分类法 线粒体呼吸 红波罗花 系统发育 粗腿蚜属 空间分布 稳定碳同位素比值 稳定同位素示踪 稀土元素 秦岭-大别造山带 祁连山 界定 生理生态过程 生活型演变 生活型 生境 球粒陨石 环境变迁 独立比较 灾害 潜在植被 海拔高度 海拔 浒湾变质带 浅层崩滑体 沉积岩 水资源 水分利用效率 水分供应 氮矿化 气候变化 武夷山自然保护区
《2024年羊草类受体蛋白激酶基因鉴定与功能研究》范文

《羊草类受体蛋白激酶基因鉴定与功能研究》篇一一、引言羊草作为一种重要的牧草资源,在农业生态系统中扮演着重要的角色。
近年来,随着分子生物学技术的快速发展,基因鉴定与功能研究逐渐成为生物学领域的研究热点。
本文以羊草类受体蛋白激酶基因为研究对象,旨在通过对该基因的鉴定及功能研究,为羊草的遗传育种、生态利用和改良提供理论基础。
二、材料与方法1. 材料本研究所用材料为羊草的基因组DNA及cDNA。
2. 方法(1)基因克隆与序列分析:利用PCR技术,从羊草基因组DNA中克隆出受体蛋白激酶基因,并进行序列分析。
(2)生物信息学分析:利用生物信息学软件,对克隆得到的基因序列进行注释、预测其编码的蛋白质结构及功能。
(3)转基因技术:构建过表达和敲低表达载体,利用转基因技术将其导入羊草细胞或组织中,研究其对羊草生长发育及抗逆性的影响。
(4)生理生化分析:通过对转基因羊草的生理生化指标进行测定,如酶活性、蛋白质含量等,探究受体蛋白激酶基因的功能。
三、结果与分析1. 基因克隆与序列分析通过PCR技术成功克隆出羊草类受体蛋白激酶基因,序列分析表明该基因具有典型的激酶结构域,属于受体蛋白激酶家族成员。
2. 生物信息学分析利用生物信息学软件对克隆得到的基因序列进行注释,预测其编码的蛋白质具有激酶活性,参与信号传导等生物学过程。
此外,通过跨膜结构域预测,发现该蛋白具有多个跨膜结构域,可能为膜蛋白。
3. 转基因技术及生理生化分析(1)过表达实验:将过表达载体导入羊草细胞或组织中,发现过表达该基因的羊草表现出更强的抗逆性及生长优势。
(2)敲低表达实验:通过RNA干扰技术敲低该基因的表达,发现羊草的生长速度及抗逆性明显降低。
(3)生理生化分析:测定转基因羊草的酶活性、蛋白质含量等生理生化指标,发现过表达该基因的羊草具有更高的酶活性和蛋白质含量,有利于提高其生物量和抗逆性。
四、讨论本研究鉴定了羊草类受体蛋白激酶基因,并对其功能进行了初步研究。
虎耳草属的资源分类及育种进展

㊀Guihaia㊀Jan.2024ꎬ44(1):193-206http://www.guihaia-journal.comDOI:10.11931/guihaia.gxzw202212037唐世梅ꎬ张霞ꎬ陈倩如ꎬ等ꎬ2024.虎耳草属的资源分类及育种进展[J].广西植物ꎬ44(1):193-206.TANGSMꎬZHANGXꎬCHENQRꎬetal.ꎬ2024.AdvancesontaxonomyandbreedingofSaxifragaTourn.exL.[J].Guihaiaꎬ44(1):193-206.虎耳草属的资源分类及育种进展唐世梅1ꎬ2ꎬ3ꎬ4ꎬ张㊀霞2ꎬ4ꎬ陈倩如2ꎬ4ꎬ蔡文淇2ꎬ4ꎬ李炎林2ꎬ4ꎬ田代科1ꎬ3∗(1.中国科学院分子植物科学卓越创新中心辰山科研中心/上海辰山植物园ꎬ上海201602ꎻ2.湖南农业大学园艺学院ꎬ长沙410128ꎻ3.上海市资源植物功能基因组学重点实验室ꎬ上海201602ꎻ4.湖南省中亚热带优质花木繁育与利用工程技术研究中心ꎬ长沙410128)摘㊀要:虎耳草属(SaxifragaTourn.exL.)资源丰富且分布广泛ꎬ全球有440~500个种ꎬ其分类一直受到植物学家的关注ꎮ该属具有重要的观赏和药用价值ꎬ欧洲多国非常重视其观赏资源的开发利用ꎮ虎耳草属的品种培育距今已有150多年的历史ꎬ到2022年国际虎耳草协会(TheSaxifrageSociety)网站收录了1692个品种ꎬ但只有1个品种来自中国ꎮ可见ꎬ我国虽然是虎耳草属的多样性中心之一ꎬ但对其观赏资源的开发利用远远落后于欧美甚至日本ꎮ该文从虎耳草属的种质资源㊁分类概况㊁育种进展等方面进行综述ꎬ并简要介绍该属资源的利用现状ꎬ为今后我国虎耳草属的分类㊁育种及应用提供借鉴ꎮ结果表明:(1)虎耳草种质资源丰富ꎬ但属下系统演化关系仍存在诸多问题ꎬ有待整合形态和系统发育学手段开展系统而深入的研究ꎮ(2)该属品种主要通过杂交育种和变异筛选的方式育成ꎬ英国㊁捷克共和国㊁德国㊁荷兰为培育品种最多的国家ꎮ(3)我国对该属的品种选育工作起步晚ꎬ育成品种少且育种方式单一ꎮ关键词:虎耳草属ꎬ栽培品种ꎬ种质资源ꎬ育种ꎬ研究进展中图分类号:Q945ꎻQ949㊀㊀文献标识码:A㊀㊀文章编号:1000 ̄3142(2024)01 ̄0193 ̄14AdvancesontaxonomyandbreedingofSaxifragaTourn.exL.TANGShimei1ꎬ2ꎬ3ꎬ4ꎬZHANGXia2ꎬ4ꎬCHENQianru2ꎬ4ꎬCAIWenqi2ꎬ4ꎬLIYanlin2ꎬ4ꎬTIANDaike1ꎬ3∗(1.ChenshanScienceResearchCenterofCASCenterforExcellenceinMolecularPlantSciencesꎬShanghaiChenshanBotanicalGardenꎬShanghai201602ꎬChinaꎻ2.CollegeofHorticultureꎬHunanAgriculturalUniversityꎬChangsha410128ꎬChinaꎻ3.ShanghaiKeyLaboratoryofPlantFunctionalGenomicsandResourcesꎬShanghai201602ꎬChinaꎻ4.HunanKeyProvincialMid ̄SubtropicalHighQualityFlowerandTreeBreedingandUtilizationEngineeringTechnologyResearchCenterꎬChangsha410128ꎬChina)收稿日期:2023-05-14基金项目:上海市绿化和市容管理局科研专项(G182412)ꎮ第一作者:唐世梅(1998-)ꎬ硕士ꎬ研究方向为国产虎耳草属的多样性及观赏价值评价ꎬ(E ̄mail)1648943359@qq.comꎮ∗通信作者:田代科ꎬ博士ꎬ研究员ꎬ博士生导师ꎬ研究方向为植物分类及多样性评价㊁植物迁地保育㊁观赏植物种质创新及利用等ꎬ(E ̄mail)dktian@cemps.ac.cnꎮAbstract:SaxifragaTourn.exL.isrichinspeciesdiversitywithabout440-500speciesandbroaddistribution.Thetaxonomicalissuesofthisgenushavebeencontinuouslyconcernedbythebotanists.Saxifragahasimportantornamentalandmedicinalvalues.SomeofEuropeancountrieshavemadegreateffortsontheexploitationandutilizationofitsornamentalresources.ChinaisoneofthediversitycentersofSaxifragaꎬbutithasbeenlaggedfarbehindEuropeꎬUSAandevenJapaninexploitationandutilizationonornamentalvalueofthisgenus.TheworldbreedingworkinSaxifragastarted150yearsago.Bytheendof2022ꎬthe1692namesofcultivarshadbeenon ̄linelistedbyTheSaxifrageSocietyꎬbutonlyonecamefromChina.InthispaperꎬthegermplasmresourcesofSaxifragaꎬandprogressonitsclassificationandbreedingarereviewedꎬandalsotheutilizationofitsornamentalresourcesisbrieflyintroducedꎬwhichwillprovideanimportantreferencefortaxonomicalresearchꎬbreedingandapplicationofthisgenusinChina.Theresultsareasfollows: (1)Saxifragaisrichingermplasmsꎬbutmanyscientificissuesremainintheevolutionaryrelationshipsofitssubordinatesystemsꎬwhichrequireasystematicandin ̄depthinvestigationbyintegratingmorphologicalandmolecularbiologymethods.(2)Thecultivarsofthisgenusaremainlyraisedthroughcross ̄breedingandmutantselectionꎬandthemajorityofcultivarsarefromtheUKꎬCzechRepublicꎬGermanyꎬandtheNetherlands.(3)ThebreedingofSaxifragastartedverylateinChinaꎬwithonlyfewcultivarsfromonebreedingmethod.Keywords:Saxifragaꎬcultivarꎬgermplasmresourceꎬbreedingꎬresearchprogress㊀㊀虎耳草属(SaxifragaTourn.exL.)是虎耳草科(Saxifragaceae)中最大的属ꎬ也是分类最为复杂㊁困难的属之一(潘锦堂ꎬ1992ꎻ张梦华ꎬ2018)ꎮ该属主要由耐寒性的多年生草本组成ꎬ稀一年或二年生草本ꎬ花瓣5ꎬ通常为辐射对称ꎬ稀两侧对称ꎬ雄蕊10ꎬ花丝棒状或钻形(潘锦堂ꎬ1992)ꎮ虎耳草属的很多种类为药材ꎬ在化学成分和药理活性方面ꎬ国内外对部分种类开展过比较深入的研究ꎮ其中ꎬ已知的药理活性包括抗菌抗病毒疗效㊁抗炎抗氧化功能以及抗突变㊁抗肿瘤㊁抗雌雄激素的作用ꎬ甚至有护肝㊁诱导成纤维细胞凋亡㊁降血糖㊁镇咳作用等(左国营等ꎬ2007ꎻ于淑玲ꎬ2009ꎻ李玉兰ꎬ2011)ꎮ挪威虎耳草(S.oppositifoliaL.)的叶子被用作茶ꎬ其叶和花可食用ꎬ因其富含维生素C而作为因纽特人饮食中必不可少的一部分(Nyvollꎬ2019)ꎮ此外ꎬ虎耳草属的高山类群具有其他大部分高山植物所不具有的优良特性ꎮ例如ꎬ该属很多种类寿命长㊁可全年观叶㊁多花㊁花色多样(白㊁黄㊁浅黄㊁橙㊁红㊁淡紫㊁紫和粉色)㊁抗虫性好以及栽培条件下往往能保持野生状态下的植物形态ꎬ而许多其他高山植物在花园里的表现通常不如在野外(Sellarsꎬ2019ꎻMalcolmꎬ2008)ꎮ鉴于虎耳草属的优良观赏特性ꎬ在全球育种者的长期努力下ꎬ培育出大量品种并在园林中应用ꎮ截至2022年底ꎬ负责虎耳草属栽培品种国际登录的权威机构TheSaxifrageSociety(国际虎耳草协会)网站(https://www.saxifrage.org/)已收录了1692个虎耳草属的品种名称ꎮ在国外ꎬ部分观赏价值高的虎耳草属种类和品种已被广泛应用于花境布置㊁岩石园点缀㊁盆栽观赏等ꎬ而国内对虎耳草属观赏价值的开发利用程度还远远不够ꎬ目前仅局限于虎耳草(S.stoloniferaCurt.)一种ꎮ研究方面主要关注虎耳草属的药用价值ꎬ忽视了其观赏价值(Takedaetal.ꎬ2020ꎻKawaharaetal.ꎬ2021)ꎮ近年来ꎬ虎耳草因其漂亮的株形㊁叶形㊁叶色和极强的栽培适应性等优点而在国内越来越多地被作为盆栽观赏㊁盆景点缀㊁岩石园和地被植物等进行应用(孔令亚等ꎬ2014ꎻ刘嘉等ꎬ2016)ꎮ但是ꎬ该属其他种类的观赏价值开发利用方面还是空白ꎬ至今还没有培育出一个杂交品种ꎬ与国外相比差距甚大ꎮ为了让科技工作者和大众尽快了解这种差距ꎬ促使我国园艺界对虎耳草属资源调查分类和其观赏价值开发利用的重视ꎬ本文对虎耳草属的全球资源及分类状况㊁虎耳草属的育种历史㊁进展和方法等进行分析总结ꎬ并对我国虎耳草属育种和观赏价值的开发利用提出一些有益思考和建议ꎮ1㊀虎耳草属资源及分类虎耳草属全球有440~500个种ꎬ约占虎耳草科的三分之二(Tkachetal.ꎬ2015)ꎮ虎耳草属以适应寒冷的多年生草本植物为主ꎬ广泛分布于北半球ꎮ该属起源于北美的落基山ꎬ向南扩散到南491广㊀西㊀植㊀物44卷美洲的安第斯山和火地岛ꎬ在北部经白令海峡扩散到欧亚大陆ꎮ在这些地区ꎬ其高度的生物多样性可与蒿属(ArtemisiaL.)㊁薹草属(CarexL.)㊁毛茛属(RanunculusL.)和早熟禾属(PoaL.)等相媲美(Ebersbachetal.ꎬ2016)ꎮ虎耳草属生于林缘㊁草原㊁冻土地带㊁岩坡石隙ꎬ其多样性中心包括中国 喜马拉雅山脉㊁高加索山脉㊁阿尔卑斯山脉以及北极的山区(Elvenetal.ꎬ2011)ꎮ虎耳草属在生长习性㊁形态㊁营养特性㊁生殖特性以及花粉粒和种子的微观形态上都表现出显著多样性ꎬ因此该属在研究高山植物系统演化㊁物种进化等领域常作为模式类群(Ebersbachetal.ꎬ2018)ꎮ在繁殖方式方面ꎬ虎耳草属除了主要通过种子繁殖外ꎬ有些种类还可通过匍匐茎㊁球茎㊁珠芽等方式繁殖ꎮ该属的授粉方式通常为昆虫传粉ꎬ少数自交授粉(Zhmylevꎬ2001)ꎮ我国虎耳草属南北均产ꎬ主要分布于西南㊁青海和甘肃等省的高山地区ꎻ其中云南分布最多ꎬ约100种ꎬ多生长在海拔3000m以上的高山岩石上及石缝间(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)ꎮ虎耳草属组间㊁组内ꎬ或种下分类ꎬ均有植物学者关注研究ꎬ使用的分类方式包括传统形态分类㊁分子系统学(核糖体ITS片段㊁叶绿体片段㊁叶绿体基因组㊁ddRAD ̄seq等)及数量分类学(Zhangetal.ꎬ2015ꎻFujiietal.ꎬ2020)方式等ꎮ林奈(1753)正式建立了虎耳草属ꎬ当时该属仅含31种但无属下分类ꎮHaworth(1803)首次对该属进行了属下等级分类ꎬ将其划分为6个组[Sect.NudicaulesHaw.㊁石荷叶组(Sect.IrregularesHaw.)㊁舌形组(Sect.LigulataeHaw.)㊁山羊臭组(Sect.CiliataeHaw.)㊁Sect.LobataeHaw.㊁Sect.StoloniferaeHaw.]ꎮ虎耳草属最早的分类系统由Engler和Irmscher(1916 1919)建立ꎬ后来由Gornall(1987)修订ꎬ包括15个组㊁19个亚组和34个系ꎬ当前被国际虎耳草协会使用ꎮ近年来ꎬ虎耳草属分子系统的研究获得迅速发展ꎬ相关研究表明该属独立为一个单系ꎬ并与莛虎耳草属(MicranthesHaw.)互为姊妹支系(Xiangetal.ꎬ2012ꎻTkachetal.ꎬ2015ꎻDengetal.ꎬ2015)ꎬTkach等(2015)利用254种虎耳草属植物在分子系统发育分析中确定了虎耳草属的主要分支ꎬ并与以往的3个分类处理进行了比较ꎬ确认了该属至少有13个组和9个亚组ꎮ对虎耳草属的组间关系研究表明ꎬ虎耳草组(Sect.Saxifraga)与山羊臭组互为姊妹单系(Vargasꎬ2000ꎻGaoetal.ꎬ2015)ꎬ舌形组与糙叶组[Sect.Trachyphyllum(Gaudin)W.D.J.Koch]是多系类群(Dechaineetal.ꎬ2013ꎻDechaineꎬ2014)ꎮConti等(1999)根据ITS和matK基因对分布在欧洲的舌形组进行了分析ꎬ结果显示ꎬ舌形组是一个多系类群ꎬ该组的一些类群与垫状组(Sect.PorphyrionTausch)聚成一支ꎻ张得均等(2008)根据分子系统地理研究成果表明山羊臭组分支内所涉及的5个亚组分为两大支ꎮZhang等(2020)利用ITS㊁trnL ̄F㊁PSBA ̄trnH和matK重组了石荷叶组的系统发育关系ꎬ结果支持东亚的石荷叶组与北美的Sect.HeterisiaRaf.各为一支ꎮ此外ꎬ近年来与该属近缘种间(Gaoetal.ꎬ2017ꎻMagotaetal.ꎬ2018ꎻMagotaetal.ꎬ2021)或种内(Abbott&Comesꎬ2003ꎻJørgensenetal.ꎬ2006ꎻWinkleretal.ꎬ2012ꎻMagotaetal.ꎬ2021)遗传分化相关的研究也有报道ꎬ其中涉及最多的物种包括广泛分布于欧洲(西班牙㊁意大利和希腊北部)㊁亚洲(喜马拉雅山)和北美(南至科罗拉多)的挪威虎耳草ꎬ该种有8~9个亚种(Malcolmꎬ2008)ꎬ以及广泛分布于日本的齿瓣虎耳草(S.fortuneiHook.f.)等ꎮ在我国ꎬ虎耳草一名始载于南宋医药学家王介著的«履巉岩本草»(张慧等ꎬ2021)ꎮ1992年潘景堂确定我国有虎耳草属植物203种ꎬ并将其分为8个组:双喙组[Sect.Birostres(Gornall)C.Y.Wu&J.T.Pan]㊁斑点组[Sect.Punctatae(Engl.)J.T.Pan]㊁石荷叶组㊁球茎组(Sect.MesogyneSternb.)㊁小花组[Sect.Micranthes(Haw.)D.Don]㊁山羊臭组㊁舌形组和点地梅形组(Sect.AretiariaSternb.)(潘锦堂ꎬ1992)ꎮFloraofChina(FOC)记载国产该属植物216种ꎬ重新分为7组:小花组㊁石荷叶组㊁糙叶组㊁山羊臭组㊁球茎组㊁垫状组㊁虎耳草组(Panetal.ꎬ2001)ꎬ同以往相比变化较大ꎮ在物种方面ꎬ国产虎耳草属已有245种发表(图1:A)ꎬ其中FOC中收录了216种ꎬ2001 2022年发表了29种(图1:B)ꎮ挪威虎耳草在1753年被命名ꎬ是国产虎耳草属最早发表的种类ꎬ这与林奈命名虎耳草属的时间相同(Linnaeusꎬ1753)ꎮ此后ꎬ1760 1820年仅有1个虎耳草属种类被命名ꎻ1910 1920年是命名发表的高峰期ꎬ共有46种被发表ꎬ与虎耳草属最早的分类系统建立时间相对应(Engler&Irmscherꎬ1958ꎻ5911期唐世梅等:虎耳草属的资源分类及育种进展数据来源于FloraofChina(2001)及其后发表文献㊁国际植物名称索引(IPNI)和世界植物(POWO)在线ꎻ2020s:2020 2022ꎮThedataarefromFloraofChina(2001)ꎬlaterpublicationsꎬInternationalPlantNamesIndex(IPNI)andPlantsoftheWorldOnline(POWO)ꎻ2020s:2020 2022.图1㊀虎耳草属国产种类发表历史及21世纪该属全球物种增加状况Fig.1㊀HistoryofChineseSaxifragadescribedwithtimeperiodandgloballyincreasedspeciesnumberinthe21stcentury691广㊀西㊀植㊀物44卷Englerꎬ1916 1919)ꎻ第二个发表高峰期为1990 2000年ꎬ共有24种被描述发表ꎮ从全球范围来看ꎬ根据国际植物名称索引(InternationalPlantNamesIndexꎬIPNI)(https://www.ipni.org/)和世界植物在线(PlantsoftheWorldOnlineꎬPOWO)(https://powo.science.kew.org/)的数据查询结果(图1:Bꎬ表1)ꎬ2001 2022年共有40个种类发表ꎬ其中最多的一年(2001年)发表15种ꎬ均来自中国ꎬ有7年(2002ꎬ2004ꎬ2005ꎬ2011ꎬ2013 2015)全球没有种类发表ꎮ在最近这22年期间ꎬ我国发表29种ꎬ其中2001年发表数量较集中ꎬ此后每年发表的种类很少ꎮ2㊀虎耳草属育种进展2.1育种历史、品种数量及其特点虎耳草属的育种历史已有150多年ꎮ通过对国际虎耳草协会的品种数据库进行检索和统计分析发现ꎬ共检索到1870 2020年有培育时间记载的虎耳草属品种1455个ꎬ其中ꎬ奥地利的Kerner和Anton于1870年培育的 Guthrieana 是最早有明确育种者姓名及品种培育年份记载的品种ꎮ按照每10年培育出的品种数量统计发现波动较大ꎬ如1870 1880年培育的虎耳草属品种最少ꎬ仅4个ꎻ随后品种培育总数逐渐增加ꎬ到1930 1940年和1940 1950年又分别减少到7和13个ꎻ而1990 2000年培育的品种最多ꎬ达377个(图2)ꎮ自1890年以来培育的虎耳草属品种数量占比66.19%ꎬ相比1890年以前ꎬ这是一个惊人的数据ꎬ说明了近40年以来该属品种培育的快速兴起ꎮ根据不同虎耳草属品种的来源对其进行划分ꎬ可分为3类ꎬ即来源于直接选育(920个)㊁杂交(454个)和品种群(296个)ꎬ各占比54.66%㊁27.75%和17.59%ꎮ1880 1980年育出的虎耳草属品种主要来源于杂交选育ꎻ1980 2000年育出的虎耳草属品种主要来源于原种的直接选育(434个)和杂交育种(190个)ꎬ各占比59.37%和25.99%ꎻ2000年之后以选择育种方式为主(图2)ꎮ按照品种所培育的国家分析ꎬ据不完全统计共有来自16个国家培育了1487个虎耳草属品种ꎮ其中ꎬ英国㊁捷克共和国㊁德国㊁荷兰和美国培育的品种数分别为609㊁390㊁197㊁120和78个ꎬ合计占总数的93.75%ꎻ其他国家培育的品种少ꎬ如意大利㊁日本㊁瑞士和爱尔兰分别培育了27㊁27㊁19㊁6个品种ꎻ加拿大㊁奥地利和法国仅分别培育了5㊁3㊁2个品种ꎬ中国㊁挪威㊁瑞典和比利时均只分别培育了1个品种(图3)ꎮ根据不同年份和国家对虎耳草属品种进行检索ꎬ收录品种中均有年份和国家记载的虎耳草属品种共计1380个ꎮ从育种历史上看ꎬ育种历史最为悠久的国家有英国㊁瑞士和奥地利ꎬ在1870 1880年就有虎耳草属的品种选育ꎻ其次ꎬ德国的虎耳草属品种记录首次出现在1890 1900年ꎻ随后ꎬ爱尔兰和法国在1910 1920年有虎耳草属品种育出ꎮ从不同国家所培育的虎耳草属品种数量比较ꎬ英国排名第一ꎬ共培育了556个(40.29%)ꎻ虽然捷克共和国在1930 1940年才出现虎耳草属品种记载ꎬ时间均晚于上述欧洲6国ꎬ但其培育的品种总量仅次于英国ꎬ为383个(27.75%)ꎻ排在第三位的为德国ꎬ共计168个(12.17%)ꎻ而瑞士和奥地利虽然在虎耳草属的育种方面起步早ꎬ但育出的品种总数分别仅有14和2个ꎮ英国和德国分别自1870年和1890年以来均有虎耳草属品种培育出ꎻ荷兰和美国培育虎耳草属品种起步较晚ꎬ分别于1990年和1960年才有品种出现ꎬ但品种总数分别居第4㊁第5位ꎮ特别是荷兰ꎬ仅1900 2020年30年间的品种数就达到全球第4位ꎬ可见该国的育种发展速度之快ꎮ反观我国ꎬ虎耳草属品种的首次登录时间为2014年(Tianꎬ2014)ꎬ并且为该属首个有正式名称的品种ꎮ因此ꎬ从育种历史和培育的品种数量上看ꎬ我国与发达国家相差甚远ꎮ2.2虎耳草属的育种方式2.2.1直接选育㊀直接选育是培育虎耳草属新品种最常用㊁最有效的方法ꎮ特别是自1990年以来ꎬ通过直接选育的方式将野生变种㊁变型或变异个体培育成符合观赏目标的园艺品种成为虎耳草属的主要育种方法ꎮ通过此方式培育的品种有912个ꎬ占总品种数的53.90%ꎬ分别来源于Sect.PorphyrionTausch(591个)㊁舌形组(189个)㊁石荷叶组(106个)㊁虎耳草组(24个)和Sect.GymnoperaD.Don(2个)5个组ꎮ国际虎耳草协会对部分直接选育的品种来源的记录并没有都具体到种ꎬ甚至部分只记录到组㊁亚组或系ꎬ推测其可能是由于原始育种记录信息不全的缘故ꎬ尤其是早期培育的品种ꎮ7911期唐世梅等:虎耳草属的资源分类及育种进展891广㊀西㊀植㊀物44卷表1㊀2001—2022年中国虎耳草属发表种类Table1㊀SpecieslistofSaxifragapublishedfrom2001to2022inChina序号Latinname发表时间PublishedtimeNo.中文名(文献)Chinesename(Reference)拉丁名1奔子栏虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.benzilanensisH.Chuang20012碧江虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.bijiangensisH.Chuang20013菖蒲桶虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.champutungensisH.Chuang20014大海虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.dahaiensisH.Chuang20015东川虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.dongchuanensisH.Chuang20016东旺虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.dongwanensisH.Chuang20017拟繁缕虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.habaensisC.Y.WuexH.Chuang20018横断山虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.hengduanensisH.Chuang20019景东虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.jingdongensisH.Chuang200110泸水虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.lushuiensisH.Chuang200111大心虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.megacordiaC.Y.WuexH.Chuang200112小叶虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.minutifoliosaC.Y.WuexH.Chuang200113细虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.pseudoparvulaH.Chuang200114山箐虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.valleculosaH.Chuang200115多痂虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.versicallosaC.Y.WuexH.Chuang200116斑玛虎耳草(Panetal.ꎬ2006)S.banmaensisJ.T.Pan200617丁青虎耳草(Panetal.ꎬ2006)S.dingqingensisJ.T.Pan200618大理虎耳草(KunmingInstituteofBotanyꎬChineseAcademyofSciencesꎬ2006)S.daliensisH.Chuang200619小中甸虎耳草(Panetal.ꎬ2007)S.xiaozhongdianensisJ.T.Pan200720泸定虎耳草(Panetal.ꎬ2007)S.ludingensisJ.T.Pan200721大桥虎耳草(Wangetal.ꎬ2008)S.daqiaoensisF.G.Wang&F.W.Xing200822克纲虎耳草(Zhangetal.ꎬ2017)S.kegangiiD.G.ZhangꎬY.Meng&M.H.Zhang201723罗霄虎耳草(Zhangetal.ꎬ2018)S.luoxiaoensisW.B.LiaoꎬL.Wang&X.J.Zhang201824绿瓣虎耳草(Gaoetal.ꎬ2018)S.viridipetalaZ.X.Zhang&Gornall201825大明山虎耳草(Zhaoetal.ꎬ2019)S.damingshanensisW.B.LiaoꎬW.Y.Zhao&J.H.Jin201926神农虎耳草(Zhangetal.ꎬ2019)S.shennongiiL.WangꎬW.B.Liao&J.J.Zhang201927绿花虎耳草(Zhangetal.ꎬ2021)S.viridifloraX.J.ZhangꎬT.DengꎬJ.T.Chen&H.Sun202128文县虎耳草(Zhangetal.ꎬ2022)S.sunhangianaT.DengꎬX.J.Zhang&J.T.Chen202229阳朔虎耳草(Chenetal.ꎬ2022)S.yangshuoensisHaiL.ChenꎬW.B.Xu&YanLiu2022数据来源于国际虎耳草协会(2022年前)ꎮ下同ꎮThedataarefromTheSaxifrageSociety(before2022).Thesamebelow.图2㊀虎耳草属历年培育出的品种数量Fig.2㊀NumberofSaxifragacultivarsraisedindifferentperiods图3㊀世界各国培育的虎耳草属品种数量比较Fig.3㊀NumbercomparisonofSaxifragacultivarsproducedfromdifferentcountries㊀㊀垫状组大约有110个种分布于欧洲㊁喜马拉雅山脉和中国西南部ꎬ大多是株高仅几厘米的垫状植物(Brochmann&Hapnesꎬ2001)ꎮ值得注意的是ꎬ表2中的S.burseriana和S.marginata都属于Ser.Kabschiaꎬ而Ser.Kabschia和Ser.Engleria都是Subsect.Porophyllum的下级分类单位ꎬ挪威虎耳草则来自于Subsect.Oppositifoliaeꎮ由此计算出表中对Subsect.Porophyllum直接选育的品种有527个ꎬ说明育种家热衷于通过对Subsect.Porophyllum直接选育来筛选新品种ꎬ也说明Subsect.Porophyllum的原种具有较高的观赏价值且变异丰富ꎻ挪威虎耳草也是来自于Sect.Porphyrion的下级分类单元对叶亚组中值得关注的类群ꎬ由此种筛选出28个品种ꎮ舌形组包括在欧洲和非洲北部的约10个种(Brochmann&Hapnesꎬ2001)ꎬ其中锥花虎耳草(S.paniculata)和匙叶虎耳草(S.cochlearis)的分布非常广泛ꎬ其他种的分布则呈现破碎化(Malcolmꎬ2008)ꎮ按照Gornall(1987)的分类ꎬ舌形组中大多数都有美丽的白花ꎬ但也有一些开红色或粉色花的优良品种ꎬ如 Foster sRed HareKnollBeauty Rosea (Sellarsꎬ2019)ꎮ通过对该组进9911期唐世梅等:虎耳草属的资源分类及育种进展表2㊀选择育种获得10个及以上品种的亲本来源Table2㊀Originofcultivarsmorethantenthroughselectionbreeding序号No.选择育种的来源Sourceofbreeding品种数Numberofcultivars1Subsect.Porophyllum2872Ser.Kabschia1683Sect.Ligulatae1014S.fortunei865S.paniculata536Ser.Engleria387S.oppositifolia288S.burseriana249S.cochlearis1210S.stolonifera1211S.marginata10㊀注:数据来源于国际虎耳草协会(2022年前)ꎮ下同ꎮ㊀Note:ThedataarefromTheSaxifrageSociety(before2022).Thesamebelow.行直接选育所产生的品种共166个ꎬ包括从锥花虎耳草和匙叶虎耳草分别直接选育的53和12个品种(表2)ꎮ锥花虎耳草为多年生植物ꎬ非常耐寒㊁不易染病ꎬ也不需要过多打理(Malcolmꎬ2008)ꎬ这可能是它能被广泛选育并应用于园林中的重要原因之一ꎮ虽然舌形组有约10个种ꎬ但对该组的资源开发利用与有近110个种的垫状组相比并不逊色ꎮ石荷叶组有10~20种ꎬ分布于亚洲东部ꎬ直立且株高(含花序)可达40cmꎬ有大的带叶柄的基生叶(Gornallꎬ1994)ꎮ据作者不完全统计ꎬ虎耳草属的品种中通过齿瓣虎耳草选择育种培育出的品种共计86个ꎮ1993 2015年由英国皇家园艺学会(RoyalHorticulturalSocietyꎬRHS)给51个虎耳草属品种颁发了花园优秀奖(AwardofGardenMeritꎬAGM)ꎬ其中70.59%的品种来自选择育种ꎬ有14个品种来自齿瓣虎耳草ꎬ其中13个品种在2015年获奖ꎮ不仅如此ꎬ由于日本的27个品种中有24个来自齿瓣虎耳草ꎬ其原因可能是齿瓣虎耳草的变种 大文字草(S.fortuneivar.incisolobataEngl.&Irmsch.)的野生资源分布广泛ꎬ在日本的北海道㊁本州㊁四国㊁九州等地均产ꎬ多样性高ꎬ自然变异大ꎬ叶㊁花形㊁色各异ꎬ花大而多ꎬ因此大文字草品种繁多ꎬ美不胜收(图4:A)ꎮ大文字草的花瓣不等长ꎬ上侧3瓣明显短于下侧2瓣ꎬ从而组成一个 大 字ꎬ由此得名(刘嘉等ꎬ2016)ꎮ同为石荷叶组的虎耳草不仅是药材还是一种野菜ꎬ如在中国有虎耳草炒鸡蛋ꎻ在日本则拥有更多千奇百怪的做法ꎬ如虎耳草叶天妇罗(一种油炸食物)㊁凉拌㊁煮汤等ꎬ人们对它味道的评价是具有一股青草香ꎬ颇为奇特ꎮ日本育种者培育的虎耳草品种七变化 ( NanaHenge )㊁ 御所车 ( Tricolor )和 极姬雪の下 ( GokuhimeYukinoxita )等很受国内外爱好者的欢迎(图4:BꎬCꎬF)ꎮ 七变化 在冬季的新叶为纯白色ꎬ随着生长叶片转为绿白相间ꎬ老叶会泛起红晕ꎬ甚至有全株雪白的情况ꎬ变化多端ꎬ十分新奇ꎻ 御所车 的叶片有不规则的白色或浅粉色镶边ꎬ叶中部绿色或绿中带粉ꎬ典雅华美ꎻ 极姬雪の下 植株矮小ꎬ叶片小而密集ꎬ最大叶仅如无名指指甲般大小ꎬ娇小玲珑ꎬ十分可爱ꎬ是做微型盆景的上佳材料(陶隽超等ꎬ2016)ꎮ除此之外ꎬTian(2014)从野外发现的虎耳草变异株筛选培育而成的新品种 黑魁 ( HeiKui )ꎬ叶片莲座状紧凑排列ꎬ植株健壮㊁株型优美ꎬ其叶莲座状排列ꎬ叶表面脉间规则分布大小不等的纺锤形黑眉状斑纹ꎬ观赏性高ꎬ极具园林应用潜力(图4:G)ꎬ在北京举行的 第二届园林植物新品种新技术交流会 被评为最具潜力新品种(张哲ꎬ2016)ꎮ 黑魁 虎耳草也是我国首个自主培育和首个国际登录的虎耳草品种(田代科ꎬ2015)ꎮ此外 雪纹 ( XueWen )和 天目恩赐 ( TianmuEnci )为上海辰山植物园2020年申报完成国际登录的品种(图4:DꎬE)ꎬ前者叶片表面沿着叶脉具有宽窄不等的白色条纹ꎬ该品种尽管在我国园林应用的历史悠久ꎬ但在完成国际登录前一直没有具体的品种名ꎬ同更为常见的纯绿色叶片个体一起在民间统称为虎耳草ꎻ后者为新选育出的新品种ꎬ叶片表面脉区白纹狭窄ꎬ脉间为深绿和黑条斑(但对光照较敏感)ꎬ密被粉红色长毛ꎬ生长旺盛ꎬ观赏价值高ꎬ尚未大量推广(田代科ꎬ2022)ꎮ虎耳草组分布最广(欧洲㊁非洲北部㊁亚洲㊁北美和南美洲)ꎬ该组由70~85个种组成ꎻSect.Gymnopera(4个种)都分布在欧洲的山区(Webb&Gornallꎬ1989)ꎮ虎耳草组的物种数虽然远多于舌形组和石荷叶组ꎬ并且在物种分布范围上占优势ꎬ但数据显示通过直接选育出的该组品种数却相对较少ꎮ实际上ꎬ全球育种者统一把该组部分观赏002广㊀西㊀植㊀物44卷A.大文字草系列品种ꎻB. 七变化 ꎻC. 御所车 ꎻD. 雪纹 ꎻE. 天目恩赐 ꎻF. 极姬雪の下 ꎻG. 黑魁 ꎮA.CultivarsofS.fortuneivar.incisolobataꎻB. NanaHenge ꎻC. Tricolor ꎻD. XueWen ꎻE. TianmuEnci ꎻF. GokuhimeYukinoxita ꎻG. HeiKui .图4㊀部分观赏虎耳草属品种举例Fig.4㊀SomeornamentalcultivarsofSaxifraga表3㊀超过10个杂交品种的杂交群亲本组合Table3㊀Grexwithcrossparentsinvolvedincultivarsmorethanten序号No.杂交群名Grexname亲本1Parent1亲本2Parent2亲本3Parent3亲本4Parent4品种数Numberofcultivars1S.ˑmegaseaefloraS.aretioidesS.lilacinaS.mediaS.burseriana672S.ˑpoluanglicaS.aretioidesS.lilacinaS.mediaS.poluniniana343S.ˑanglicaS.aretioidesS.lilacinaS.media 254S.ˑborisiiS.marginataS.ferdinandi ̄coburgi 245S.ˑboydiiS.aretioidesS.burseriana 216S.ˑelisabethaeS.burserianaS.sancta 217S.ˑirvingiiS.burserianaS.lilacina 158S.ˑpetraschiiS.burserianaS.tombeanensis 149S.ˑsalmonicaS.burserianaS.marginata 1110S.ˑconcinnaS.cinereaS.dinnikii 1011S.ˑhornibrookiiS.lilacinaS.stribrnyi 1012S.ˑlismorensisS.aretioidesS.lilacinaS.mediaS.georgei10㊀注: 表示无ꎮ㊀Note: indicatesno.价值较高的种或杂交品种归为了SaxifrageMossyGroup栽培群ꎮ2.2.2杂交育种㊀杂交育种是选育新品种的重要培育方式之一ꎮ在已登录的虎耳草属所有杂交品1021期唐世梅等:虎耳草属的资源分类及育种进展表4㊀超过10个杂交品种的相关原生种亲本Table4㊀Speciesproducingmorethantenhybridcultivars序号No.原生种Species杂交品种数Numberofhybridcultivars中国分布DistributioninChina发表年份Yearpublished1Saxifrageburseriana200无No17532S.aretioides194无No18013S.lilacina188无No19044S.media151无No17735S.marginata85无No18226S.ferdinandi ̄coburgi65无No19017S.poluniniana51无No19588S.sancta41无No18439S.stribrnyi38无No190210S.tombeanensis29无No186911S.sempervivum24无No190612S.cinerea22无No195813S.dinnikii13无No189214S.corymbosa11有Yes1843种中ꎬ垫状组的组内种间或品种间杂交产生的杂交品种共计456个(98.06%)ꎬ并且均来自Subsect.Porophyllum的3个亚系间或亚系内(Kabschia㊁Engleria㊁Tetrameridium)杂交ꎬ与Malcolm(2008)的专著记载相同ꎻ舌形组的组内种间或品种间的杂交品种共计仅9个(1.94%)ꎬ来自5个杂交组合ꎬ但未见组间杂交品种的记载ꎮ垫状组共登录有92个杂交组合ꎬ使用最多的是杂交组合是S.ˑmegaseaeflora(S.aretioides㊁S.burseriana㊁S.lilacina㊁S.media)ꎬ共培育出67个品种ꎻ其次是S.ˑpoluanglica(S.aretioides㊁S.lilacina㊁S.media㊁S.poluniniana)共培育出34个品种ꎻ居第3位的杂交组合为S.ˑanglica(S.aretioides㊁S.lilacina㊁S.media)ꎬ共育出25个品种(表3)ꎮ来自捷克共和国的RadvanHorn'y和KarelMirkoWebr发表的PorophyllumSaxifrages曾详细地介绍了该亚组3个亚系的种及杂交品种(Malcolmꎬ2008)ꎮ虎耳草属的原生种作为亲本参与杂交培育的品种数如表4所示ꎬ其中涉及S.burseriana(1753年发表)的品种最多ꎮ14个参与杂交的原生种中除了近优越虎耳草(S.corymbosa)外ꎬ其余13个原生种在我国均不产ꎬ说明国外主要利用当地的种质资源进行新品种培育ꎬ我国的虎耳草属种质资源很少被欧美用于育种ꎮ研究表明ꎬ垫状组的自然杂交种分布于欧洲和高加索地区ꎬ喜马拉雅地区也可能广泛存在该组的自然杂交种(Malcolmꎬ2008)ꎮ英国育种家Gornall(1994)在SomeaspectsofhybridisationinthegenusSaxifraga一文中详细介绍了虎耳草属人工杂交育种方法的步骤ꎮ用酒精给镊子消毒ꎬ并在母本的花药成熟前去雄ꎬ使用空茶袋㊁薄纱袋甚至纸袋来套袋以避免去雄的花朵被污染ꎻ当母本的柱头变得潮湿或膨胀时ꎬ用镊子或清洗干净的毛笔取父本的成熟花粉在母本柱头表面轻轻摩擦ꎬ每次授粉前都须给镊子消毒ꎻ授粉后将母本重新套袋ꎬ挂上标签并在笔记本上记录日期ꎬ以及父本母本的信息等ꎮ虎耳草属植物的花粉产生后ꎬ会在几天或几周后失活ꎬ若花粉被弄湿ꎬ花粉活力仅可保持几个小时ꎮ一般来说双核花粉的花粉活力比三核花粉持续时间更长ꎮ虎耳草属中除山羊臭组为三核花粉外ꎬ其他均为双核花粉ꎮ因此ꎬ除山羊臭组外ꎬ其他虎耳草属物种的花粉均适合低温条件下保存以后使用ꎮ可在花药裂开前收集花药保存在干燥的小瓶中放入冰箱冷藏ꎬ这样花粉活力可保持一个月ꎬ冷冻起来可以保持更长时间ꎬ达一年ꎬ但不同种间会存在差异ꎮ此外ꎬRichardGornall还介绍了虎耳草属人工杂交的实验设计及衡量杂交是否成功的标准等ꎮ2.3虎耳草属品种群«国际栽培植物命名法规»(第九版)规定:品种群是基于一定相似性的品种㊁植物个体或植物集合体的正式等级ꎬ品种群的品种可能来自直接选育ꎬ也可能来自杂交(向其柏和臧德奎ꎬ2016)ꎮ国际虎耳草协会数据库中共有来自68个品种群的296个品种ꎬ占总品种数的17.49%ꎮ品种数排在前三位的品种群分别为SaxifrageMossyGroup㊁SaxifrageBluesGroup和SaxifrageLondonPrideGroupꎬ分别有138(46.62%)㊁24(8.11%)和22个品种(7.43%)ꎬ其他品种群的品种总数均小于等于6个ꎮSaxifrageMossyGroup由虎耳草组的大部分原种和杂交种组成ꎬ该栽培群植株低矮ꎬ叶片细小紧凑ꎬ形似苔藓植物ꎬ因此通常被称为苔藓虎耳草ꎬ该类品种花色丰富ꎬ有花色纯白的 Fleece ꎬ白色花瓣上带明显浅绿脉络的 Schwefelblüte 等ꎻ花色深浅不一的红色系ꎬ如浅粉色的 KnaptonPink ㊁粉红的 Glassel sCrimson ㊁玫红的 Triumph ㊁深红的 WelshDragon 等ꎮ这类虎耳草形态比较接近于生长在高山的部分种类ꎬ多数202广㊀西㊀植㊀物44卷被应用于岩石园或盆栽ꎬ其花量大ꎬ花色鲜艳ꎬ少数当成地坛草花ꎮSaxifrageBluesGroup的叶莲座状着生ꎬ叶细小ꎬ紧凑排列在地面形成一个半球形ꎬ镶嵌着生粉色㊁橘红或玫红的花朵ꎬ花瓣圆形ꎬ其中 Satchmo 的花瓣还有浅波浪状缺刻ꎮSaxifrageLondonPrideGroup来自Sect.Gymnopera(除S.cuneifolia外)ꎬ具长叶柄ꎬ叶片深绿或浅绿ꎬ圆形或匙形ꎬ具缘齿ꎬ椭圆形的白色花瓣上点缀着粉红色和黄色的斑点ꎬ雄蕊和花药为粉色ꎮ2.4虎耳草属品种的杰出育种者欧洲的育种者对虎耳草属的育种贡献最大ꎬ其中德国的育种者Sündermann和Franz共同培育出115个品种ꎬ为育种者之最ꎬ所育品种数占德国的68.45%ꎬ包括本国的首个虎耳草品种ꎮ这些品种中ꎬ有88个来源于杂交育种ꎬ26个来自选择育种ꎬ1个来自栽培群ꎮ品种数第二多的育种者为捷克共和国的Lang和Karelꎬ两位在1990 2020年共同培育出100个品种ꎬ最近一次记载为2017年ꎬ该年培育出的品种有7个ꎮ这些品种中ꎬ通过栽培群选育的最多ꎬ有71个ꎬ来自杂交育种的有16个ꎬ源于选择育种的有13个ꎮ荷兰的Moerland和Bart是品种数量第三多的育种者ꎬ二者于2000 2020年培育出99个品种ꎬ占荷兰总数的82.50%ꎮ3㊀问题与展望3.1虎耳草属的分类作为研究高山植物系统演化㊁物种进化和多样性等领域的特征类群(邓家彬ꎬ2015)ꎬ近二十年来ꎬ虎耳草属的分子系统学研究发展迅速ꎬ但该属的属下系统演化关系仍存在诸多问题(胡晶晶ꎬ2020)ꎬ组的划分存在不确定性ꎬ并不能真实地反映虎耳草属属下等级的自然发生规律ꎬ有待整合形态和系统发育学手段开展系统而深入的研究ꎮ在资源调查和新物种描述方面ꎬ进入21世纪后ꎬ除了2001年发表15种最多外ꎬ此后每年发表的种类不超过4种ꎬ甚至这期间有7年无新种描述发表ꎬ说明21世纪全球虎耳草属的经典分类学研究几乎停滞不前ꎮ云南为我国虎耳草属种类分布最多的省份ꎬ2001年发表的15种均分布在该省ꎬ显然同该省具有丰富的生物多样性相关ꎮ21世纪我国发表虎耳草属种类29个ꎬ其中石荷叶组由原来7种增加到现在的15种ꎬ新增加的种类发现于广西㊁广东㊁湖南㊁江西和甘肃ꎬ因此推测该组还可能有新种待调查ꎮ此外ꎬ2001年发表的15种国产虎耳草属种类均来自从事虎耳草专属植物分类研究的同一学者ꎬ而此后发表的种为非虎耳草专属的不同学者参与的工作ꎬ说明国内现已缺少专门针对这一类群开展研究的分类学者ꎬ我国的虎耳草属家底尚未清楚ꎬ急需年轻分类学家针对该属开展系统的资源调查㊁分类及其修订工作ꎬ而面向全球开展虎耳草属的资源调查和分类研究可能面临更长期㊁更艰巨的挑战ꎮ3.2国际虎耳草属的育种趋势及展望根据国际虎耳草协会的数据库数据统计ꎬ在有年份记载的品种数中ꎬ仅在1990 2022年培育的虎耳草属品种数占比66.19%ꎬ并且在将花卉业作为支柱产业的荷兰自1995年培育第1个虎耳草属品种以来ꎬ陆续培育出大量优良虎耳草属品种ꎬ说明了近半个世纪虎耳草属育种的兴起ꎮ但是ꎬ该属的育种大都局限于欧美国家ꎬ亚洲除日本外还处于落后阶段ꎬ目前其他洲的育种还是空白ꎮ现有记载的虎耳草属品种主要通过杂交育种和变异筛选的方式育成ꎮ杂交育种主要利用Subsect.Porophyllum的3个系间或系内杂交ꎬ组间或属间杂交未见报道ꎬ组间或属间杂交可能成为未来的重要育种方向ꎮ当前ꎬ传统的育种方式还是主流ꎬ但将来一定会选择利用诱变㊁分子技术等进行辅助育种ꎬ而这些育种方法需要不断摸索ꎮ在育种材料选择上ꎬ国际交流的日趋频繁带动了种质资源的快速流通ꎬ为育种家们提供了亲本选择的便利ꎮ但是ꎬ在引进国外的优良原种和品种时ꎬ除了主要考虑其观赏价值以外ꎬ还应充分考虑到品种对本土气候和土壤条件的环境适应性ꎮ3.3中国虎耳草属的育种现状及展望相比国际上的虎耳草属育种历程长和品种数量多ꎬ虽然中国开展该属的育种工作还很少且才刚开始不久ꎬ但也说明育种潜力巨大ꎬ因为中国的气候和资源多样性均很高ꎮ中国虎耳草属的育种存在明显不足:(1)育种起步晚ꎬ2013年才完成该属品种的首次国际登录ꎬ中国也才有了第一个虎耳草品种名( 黑魁 )ꎻ(2)品种数量太少ꎬ截至2022年底仅有3个品种(其中1个已被英国虎耳草协会数据库收录)ꎻ(3)育种材料及方式单一ꎬ品种仅通过野外变异筛选培育而成ꎬ杂交育种还没有起步ꎮ育种3021期唐世梅等:虎耳草属的资源分类及育种进展。
广东紫珠核糖体DNA ITS序列分子鉴定

广东紫珠核糖体DNA ITS序列分子鉴定作者:李家敏,姜琼,汪剑鸣来源:《湖北农业科学》 2014年第10期李家敏,姜琼,汪剑鸣(宜春学院化学与生物工程学院,江西宜春336000)摘要:采用改良CTAB法提取广东紫珠(CallicarpakwangtungensisChun.)植物总DNA,以通用引物ITS4和ITS5PCR扩增核糖体DNAITS序列,并运用MEGA5.2软件分析ITS序列和构建系统发育树。
结果表明,广东紫珠的核糖体DNAITS序列长度为658bp,ITS1和ITS2的长度分别为229、267bp,G+C含量分别为67.39%、70.30%。
用NJ法根据ITS1与ITS2序列构建系统发育树,ITS片段在种间存在差异,可区分紫珠属中不同的种。
测序分析和系统发育树构建,能够对广东紫珠进行准确的分子鉴定,为紫珠属中药材的种类鉴定和种间的分类地位提供分子生物学理论依据。
关键词:广东紫珠(CallicarpakwangtungensisChun.);ITS序列;分子鉴定中图分类号:Q78文献标识码:A文章编号:0439-8114(2014)10-2435-05广东紫珠(CallicarpakwangtungensisChun.)为马鞭草科紫珠亚属多年生落叶植物,适合生长于海拔100~1000m的丘陵山地,主要分布于江西、湖南、福建、贵州等省,广东和广西等地也有少量分布。
广东紫珠的干燥茎、枝、叶均可入药,具有收敛止血、清热解毒等功效,收载于《中国药典》(2010年版),可用于治疗咯血、便血、肺胃出血、崩漏、肺热咳嗽、咽喉肿痛、烧伤、热毒疮、外伤出血等。
化学成分研究表明,广东紫珠富含熊果酸、槲皮素、没食子酸、水杨酸等化学成分[1];药理研究显示具有较强的抑菌作用和显著的止血效果[2],临床上广东紫珠主要用于治疗宫颈糜烂出血、阴道炎、宫颈炎等,是抗宫炎片、抗宫炎颗粒和抗宫炎胶囊等中成药的主要成分[3]。
耳草属岭南药材DNA分子鉴定研究

耳草属岭南药材DNA分子鉴定研究黄娟;李西文;徐文;丘小惠;徐江;黄志海【期刊名称】《世界科学技术-中医药现代化》【年(卷),期】2016(018)008【摘要】目的:本研究应用ITS2序列作为DNA条形码对9种广东耳草属药材进行鉴定研究.方法:收集广东耳草属药材样本,通过植物基因组DNA提取、PCR扩增以及产物的双向测序获得ITS2序列,所得序列采用CodonCode Aligner进行拼接.用MEGA 6.06进行比对,分析种内和种间遗传距离,并构建系统进化树.结果:9种耳草属植物65条序列长度范围为208-224 bp,共有92个碱基变异位点,种内遗传距离(0.002)明显小于种间遗传距离(0.202).NJ和ML聚类树结果基本一致,除耳草与金毛耳草关系较近难以区分外,其余7个物种种内样本分别聚在一支,表现出单系性.结论:ITS2序列基本能够准确的鉴定广东耳草属药材,可作为耳草属药材基原植物鉴定的候选条形码序列.【总页数】7页(P1401-1407)【作者】黄娟;李西文;徐文;丘小惠;徐江;黄志海【作者单位】广东省中医院第二临床医学院广州 510006;中国中医科学院中药研究所北京 100700;广东省中医院第二临床医学院广州 510006;广东省中医院第二临床医学院广州 510006;中国中医科学院中药研究所北京 100700;广东省中医院第二临床医学院广州 510006【正文语种】中文【中图分类】R282【相关文献】1.菊属植物DNA分子鉴定研究进展 [J], 胡志刚;夏叶;郑贝;胡志强;高欢欢;刘合刚2.耳草属植物化学成分及药理活性研究进展 [J], 杨元;姜艳艳;石任兵3.耳草属植物中环烯醚萜类化合物的研究进展 [J], 邵明国;罗永明;王文哲;杨美华4.耳草属植物的化学研究Ⅷ.黄毛耳草化学成分的分离和鉴定 [J], 彭江南;冯孝章;梁晓天5.耳草属植物化学成分的研究VI.牛白藤化学成分的研究 [J], 彭江南;冯孝章因版权原因,仅展示原文概要,查看原文内容请购买。
虎耳草属山羊臭组的界定和系统发育:核糖体DNA ITS序列证据

虎耳草属山羊臭组的界定和系统发育:核糖体DNA ITS序列证据张得钧;陈生云;高庆波;段义忠;陈世龙【期刊名称】《植物分类学报》【年(卷),期】2008(046)005【摘要】虎耳草属Saxifraga山羊臭组sect.Ciliatae是该属中最大的一个组,共有175种,主要分布在喜马拉雅地区,我国分布有166种,占总种数的95%;其中,112种为中国特有.约80%的种类分布在我国青藏高原和两南地区,是中国喜马拉雅植物成分的代表类群.山羊臭组内物种分化十分显著,分类处理也很困难,该组是否为单系类群,组下的系统发育关系也不清楚,均需进一步验证.本文测定了虎耳草属山羊臭组及其他组33种植物样品的核糖体DNA内转录间隔区ITS序列,并从GenBank调取虎耳草组sect.Saxifraga等组和近缘属唢呐草属Mitella共22种植物的该序列.ITS分析结果表明:(1)所研究的山羊臭组类群聚为单独一支,而且与垫状组sect.Porphyrion、虎耳草组、球茎组sect.Mesogyne和仪在欧洲分布的sect.Cymbalaria和sect.Cotylea等8个组聚成的另一分支构成姊妹群;(2)根据形态特征建立的山羊臭组的3个亚组即唐古拉亚组subsect.Hirculoideae、莲座状亚组subsect.Rosulares和具芽亚组subseet.Gemmiparae各自聚为一支,但是莲座状亚组这一支的支持率较低.同时,山羊臭组的鞭匐枝亚组subsect.Flagellares和subsect.Hemisphaericae的代表类群单独聚为一支.位于具芽亚组类群分支内部而不能成立;(3)唐古拉亚组和莲座状亚组又聚为一亚分支与具芽亚组构成姊妹群,而且具芽亚组最早从山羊臭组这一支中分化出来.我们的研究还发现山羊臭组内种间形态分化较大,而ITS碱基变异较小,这可能是山羊臭组类群在青藏高原及毗邻地区的高山环境下物种快速分化的结果.【总页数】9页(P667-675)【作者】张得钧;陈生云;高庆波;段义忠;陈世龙【作者单位】中国科学院高原生物适应与进化重点实验室,中国科学院西北高原生物研究所,西宁,810001;中国科学院研究生院,北京,100049;中国科学院高原生物适应与进化重点实验室,中国科学院西北高原生物研究所,西宁,810001;中国科学院研究生院,北京,100049;中国科学院高原生物适应与进化重点实验室,中国科学院西北高原生物研究所,西宁,810001;中国科学院研究生院,北京,100049;中国科学院高原生物适应与进化重点实验室,中国科学院西北高原生物研究所,西宁,810001;中国科学院研究生院,北京,100049;中国科学院高原生物适应与进化重点实验室,中国科学院西北高原生物研究所,西宁,810001【正文语种】中文【中图分类】Q94【相关文献】1.从核糖体DNA ITS区序列研究甘蔗属及其近缘属种的系统发育关系 [J], 陈辉;范源洪;向余颈攻;蔡青;张亚平2.基于叶绿体trnL-F,rbcL序列和核核糖体ITS序列探讨蓼属(蓼科)头状蓼组的系统发育 [J], 赵大鹏;王康满;侯元同3.金缕梅科银缕梅属与帕罗堤属的亲缘关系——核糖体DNA ITS序列证据 [J], 李建华;A.LinnBogle;AnitaS.Klein;潘开玉4.报春花属藏报春组、毛茛叶报春组的界定和系统发育关系 :核糖体DNAITS序列证据(英文) [J], 郝刚;胡启明;李南淑5.金粟兰属的系统发育:核糖体DNAITS区序列的证据(英文) [J], 孔宏智;陈之端因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
植物分类学报 46 (5): 667–675 (2008) doi: 10.3724/SP.J.1002.2008.07091 Journal of Systematics and Evolution (formerly Acta Phytotaxonomica Sinica) 虎耳草属山羊臭组的界定和系统发育:核糖体DNA ITS序列证据1,2张得钧1,2陈生云1,2高庆波1,2段义忠1陈世龙*1(中国科学院高原生物适应与进化重点实验室, 中国科学院西北高原生物研究所西宁 810001)2(中国科学院研究生院北京 100049)Circumscription and phylogeny of Saxifraga sect. Ciliatae:Evidence from nrDNA ITS sequences1,2De-Jun ZHANG 1,2Sheng-Yun CHEN 1, 2Qing-Bo GAO1,2Yi-Zhong DUAN 1Shi-Long CHEN *1(Key Laboratory of Adaptation and Evolution of Plateau Biota, Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810001, China)2(Graduate University of Chinese Academy of Sciences, Beijing 100049, China)Abstract The largest section of the genus Saxifraga (Saxifragaceae), sect. Ciliatae, consists of 175 morphologi-cally diverse species. This section is mainly distributed in the Qinghai-Tibetan Plateau and adjacent regions of southwest China and more than 80% of the total number of species are endemic to this region. It remains unknown whether this section is monophyletic and up to now no study has been conducted on the infra-sectional phylogeny. In this study, ITS sequences of the nuclear ribosomal DNA were firstly determined for 33 species mainly from this section and related sections. We further downloaded the corresponding sequences of the same DNA region for the other 22 species of Saxifraga and Mitella from GenBank. All sequences were together used to construct the phy-logenetic trees. The main implications of the phylogenetic analyses include: (1) sect. Ciliatae, as traditionally defined, constitutes as a monophyletic clade and its sister group is a well supported clade that includes species from 8 sections such as sect. Porphyrion, sect. Saxifraga and sect. Mesogyne; (2) three morphological subsections, i.e., subsect. Gemmiparae, subsect. Hirculoideae and subsect. Rosulares were tentatively recovered despite the relatively low statistic bootstrap support for the last one; however, subsect. Flagellares and subsect. Hemi-sphaericae were not recognized as separate entities, and nested within subsect. Gemmiparae; (3) subsect. Hircu-loideae and subsect. Rosulares clustered together as sister subclades while subsect. Gemmiparae diverged early. In addition, our results suggest that the paired variation of ITS sequences in sect. Ciliatae is relatively low between the sampled species in spite of their diverse morphology. It is suggested that such a scenario may mirror rapid speciation in this section that probably trigged by the uplifts of the Qinghai-Tibetan Plateau and the extensive selection pressure under the alpine environments.Key words circumscription, ITS, phylogeny, Saxifraga sect. Ciliatae.摘要虎耳草属Saxifraga山羊臭组sect. Ciliatae是该属中最大的一个组, 共有175种, 主要分布在喜马拉雅地区, 我国分布有166种, 占总种数的95%; 其中, 112种为中国特有。
约80%的种类分布在我国青藏高原和西南地区, 是中国喜马拉雅植物成分的代表类群。
山羊臭组内物种分化十分显著, 分类处理也很困难, 该组是否为单系类群, 组下的系统发育关系也不清楚, 均需进一步验证。
本文测定了虎耳草属山羊臭组及其他组33种植物样品的核糖体DNA内转录间隔区ITS序列, 并从GenBank调取虎耳草组sect. Saxifraga等组和近缘属唢呐草属Mitella共22种植物的该序列。
ITS分析结果表明: (1)所研究的山羊臭组类群聚为单独一支, 而且与垫状组sect. Porphyrion、虎耳草组、球茎组sect. Mesogyne和仅在欧洲分布的sect. Cymbalaria和sect. Cotylea 等8个组聚成的另一分支构成姊妹群; (2)根据形态特征建立的山羊臭组的3个亚组即唐古拉亚组subsect. Hirculoideae、莲座状亚组subsect. Rosulares和具芽亚组subsect. Gemmiparae各自聚为一支, 但是莲座状亚组这一支的支持率较低。
同时, 山羊臭组的鞭匐枝亚组subsect. Flagellares和subsect. Hemisphaericae的代表类群单独聚为一支, 位于具芽亚组类群分支内部而不能成立; (3)唐古拉亚组和莲座状亚组又聚为一亚分支与具芽亚组构成姊妹群, 而且具芽亚组最早从山羊臭组这一支中分化出来。
———————————2007-06-21收稿, 2007-09-19收修改稿。
* 通讯作者(Author for correspondence. E-mail: slchen@;Tel.: 86-971-6110067)。
植物分类学报 Journal of Systematics and Evolution 2008 46卷5期668我们的研究还发现山羊臭组内种间形态分化较大, 而ITS碱基变异较小, 这可能是山羊臭组类群在青藏高原及毗邻地区的高山环境下物种快速分化的结果。
关键词界定; ITS序列; 系统发育; 虎耳草属山羊臭组虎耳草属Saxifraga L.是虎耳草科Saxifragaceae 最大的属, 约有400种(Engler & Irmscher, 1916, 1919; Spongberg, 1972; Kohlein, 1984; Gornall, 1987; Webb & Gornall, 1989; 潘锦堂, 1992; Pan et al., 2001), 主要分布在北极和北温带, 该属的多样性中心在北美西部、东亚和喜马拉雅–青藏高原地区; 生于林缘、草原、冻土地带、岩坡石隙。
长期以来, 该属在形态学、细胞学、生境等方面的变异很大, 因此虎耳草属分类学研究一直存在争议。
自Linnaeus (1753)建立虎耳草属以来, 先后有很多分类学家对该属进行了分类学研究, 如Haworth (1812, 1821)虽摒弃了Linnaeus建立的广义虎耳草属分类系统, 将该属分成16个属,但Linnaeus的虎耳草属分类系统还是被后来的许多学者接受并进一步完善(Don, 1822; Tausch, 1823; Seringe, 1830; Engler, 1872; Engler & Irmscher, 1916, 1919; Gornall, 1987; Webb & Gornall, 1989; 潘锦堂, 1992; Pan et al., 2001)。
中国现有虎耳草属植物216种, 其中139种是中国特有种(Pan et al., 2001)。
一直以来, 对我国虎耳草属植物进行分类学及相关研究的人较少。
20世纪90年代以前, 少数分类学家曾对该属进行过研究(吴征镒, 1984; 吴征镒, 潘锦堂, 1990); 1976年以后, 我国植物分类学家潘锦堂从形态学、解剖学、区系地理学等方面对我国分布的虎耳草属植物进行了详细的研究, 在Engler和Irmscher (1916–1919)的虎耳草属分类系统基础上, 确定我国共有虎耳草属植物203种, 并将其分为8组7亚组31系和4亚系。