分子生物学:第十章真核生物基因表达调控
分子生物学李玉龙

2020 生物技术专业-分子生物学第一章:绪论1.基因表达的实质是遗传信息的转录和翻译。
2.基因的表达调控主要发生在转录水平和翻译水平上。
3.分子生物学发展过程概括为三个阶段:人类对DNA和遗传信息传递的认识阶段、重组DNA技术的建立和发展阶段、重组DNA技术的应用和分子生物学的迅猛发展阶段。
第二章:核酸的结构与功能1、判断、填空与选择考点:1.DNA 是主要的遗传物质。
2.核苷酸之间通过3’,5’磷酸二酯键连接形成核酸。
3.核苷酸是核酸的基本结构单位。
4.染色体分为常染色质和异染色质两类。
5.染色质分为组成型异染色质和兼性异染色质两类。
6.核酸是多核苷酸,核苷酸由含氮碱基、戊糖、磷酸构成。
核苷酸可以分解为核苷和磷酸,核苷可以分解为含氮碱基和戊糖。
7.稳定双螺旋结构的因素:碱基对之间形成的氢键、碱基堆积力、正负电荷的作用。
8.提出双螺旋模型有三个证据:X射线衍射法、DNA碱基等比例规律、DNA分子密度9.B-DNA是大多数DNA在细胞中的构象。
10.B-型螺旋就是Watson和Crick双螺旋√11.DNA每旋转一周,大约10个碱基对。
√12.染色质由最基本的结构单元核小体组成。
13.所有mRNA的3’端都有poly(A)结构。
×组蛋白mRNA的3’端无poly(A)结构14.检测DNA变性最简单的定性和定量方法是紫外吸收光谱变化。
15.Tm主要和DNA均一性、G-C碱基对含量、介质中离子强度有关。
16.DNA复性的两个必要条件是离子强度和较高的温度。
17.测定复性程度的3种方法:①减色效应②抗S1核酸酶水解DNA的量③羟基磷灰石柱层析。
18.分子杂交的类型与区分:①鉴定 DNA: Southern 印迹法;②鉴定 RNA: Northern 印迹法;③鉴定蛋白质:Western 印迹法。
第三章:基因与基因组的结构与功能1、判断、填空与选择考点:1.基因是遗传的基本单位,突变单位以及控制性状的功能单位。
分子生物学教案

分子生物学教案(整理版)第一章:分子生物学概述1.1 分子生物学的定义和发展历程介绍分子生物学的定义和研究范围回顾分子生物学的发展历程,如DNA双螺旋结构的发现等1.2 分子生物学的重要性和应用领域强调分子生物学在生物科学和生物技术领域的重要性介绍分子生物学在医学、农业、环境保护等领域的应用实例第二章:DNA与基因2.1 DNA的结构和功能详细描述DNA的双螺旋结构和特点解释DNA在遗传信息和基因表达中的作用2.2 基因的概念和分类介绍基因的定义和基本特征区分编码基因和非编码基因,以及原核生物和真核生物基因的区别第三章:基因表达与调控3.1 转录和翻译的过程详细解释DNA转录为mRNA的过程,包括启动子、转录因子等介绍mRNA翻译为蛋白质的过程,包括核糖体、tRNA等的作用3.2 基因表达调控机制介绍原核生物和真核生物中基因表达调控的机制和差异讨论转录因子、启动子、增强子等在基因表达调控中的作用第四章:分子克隆与基因工程4.1 分子克隆的基本原理和技术解释分子克隆的概念和基本步骤介绍PCR扩增、DNA连接、转化等分子克隆相关技术4.2 基因工程的应用和伦理问题讨论基因工程在生物制药、基因治疗等领域的应用探讨基因工程在伦理、安全、生态等方面的争议和问题第五章:蛋白质结构与功能5.1 蛋白质的结构层次介绍蛋白质的一、二、三级结构及其决定因素解释蛋白质结构与功能之间的关系5.2 蛋白质功能的多样性讨论蛋白质在生物体内承担的各种功能,如酶、结构蛋白、信号转导等介绍蛋白质工程在药物设计和疾病治疗中的应用第六章:酶学与酶工程6.1 酶的概述和特性介绍酶的定义、命名和分类解释酶的催化机制和酶活性的影响因素6.2 酶工程的应用和发展讨论酶在工业、医药、生物检测等领域的应用探讨定向进化、重组酶等技术在酶工程中的应用和发展第七章:RNA与非编码RNA7.1 RNA的结构和功能介绍RNA的种类、结构和功能解释mRNA、tRNA、rRNA等在蛋白质合成中的作用7.2 非编码RNA的研究进展讨论非编码RNA(如miRNA、siRNA、lncRNA等)的发现和功能探讨非编码RNA在疾病诊断、治疗和调控中的潜在应用第八章:蛋白质相互作用与信号转导8.1 蛋白质相互作用的基本概念介绍蛋白质相互作用的特点和机制解释生物信息学方法在蛋白质相互作用研究中的应用8.2 信号转导通路及其调控介绍细胞内主要的信号转导通路(如MAPK、Wnt、Notch等)讨论信号转导通路在细胞生长、分化、死亡等过程中的作用和调控机制第九章:基因组学与遗传变异9.1 基因组学的基本概念和技术介绍基因组学的研究内容、方法和进展解释基因组测序、基因组编辑等技术的原理和应用9.2 遗传变异与疾病讨论遗传变异在疾病发生中的作用和机制探讨遗传变异的检测、预测和疾病风险评估方法第十章:分子生物学实验技术10.1 分子生物学实验基本技术介绍PCR、电泳、免疫印迹等分子生物学实验技术解释实验操作步骤、条件和注意事项10.2 分子生物学实验设计与应用讨论分子生物学实验设计的原则和方法探讨实验结果的解读、数据分析和实验应用重点和难点解析一、分子生物学的定义和发展历程解析:了解分子生物学的概念和其发展历程对于理解后续内容至关重要。
真核生物基因表达调控

酸性激活域 (D/E-rich) 谷氨酰胺(Q)富含域 脯氨酸(P)富含域
蛋白质-蛋白质结合域 (dimerization, co-factors)
1) TF最常见的DNA binding domain
Zinc Finger
bZIP
Homeodomain
bHLH
(1) 锌指(zinc finger)
2. The pri5’ capping 3’ formation / polyA splicing
3. Mature transcripts are transported to the cytoplasm for translation
Chromatin
epigenetic control
Protein degradation RNA silencing
一般而言的基因表达调控范畴
二、基因表达的时间性及空间性
(一)时间特异性
按功能需要,某一特定基因的表达严格按 特定的时间顺序发生,称之为基因表达的时间 特异性(temporal specificity)。
Cys-X2-4-Cys-X3-Phe-X5-Leu-X2-His-X3-His C-terminal: α-helix binding DNA
常结合GC box
(2) 碱性亮氨酸拉链 bZIP
(3) 碱性螺旋-环-螺旋bHLH
bHLH蛋白(basic Helix-Loop-Helix)
2) TF常见的trans-activation domain
– usually expressed at high level – the level of their gene expression may vary
原核、真核生物基因及表达调控

原核、真核生物基因及表达调控引言现代生物学中“基因”一词甚为流行,细胞学、遗传学、生物化学等,以及各种生物学课本中,都涉及到“基因”一词。
甚至象典型的宏观生物学科——生态学,也把一片森林称为一个“基因库”[1]。
现代生物学已经完全证明,DNA 分子是由称为核普酸的有机分子线性聚合而成。
基因就是核普酸按一定顺序排列而成的DNA分子片段,它携带着遗传信息。
基因表达(gene expression)是指细胞在生命过程中,把储存在DNA顺序中遗传信息经过转录和翻译,转变成具有生物活性的蛋白质分子。
其实质就是遗传信息的转录和翻译。
在个体生长发育过程中,生物遗传信息的表达按一定的时序发生变化(时序调节),并随着内外环境的变化而不断加以修正(环境调控)[2]。
原核生物和真核生物的基因及表达过程有着差异。
随着世界分子生物学研究不断深入,基因表达技术有了很大的提高。
迄今为止,人们已经研究开发出多种原核和真核表达系统用以生产重组蛋白[3]。
一.原核、真核生物基因结构原核生物基因分为编码区与非编码区,所谓的编码区就是能转录为相应的信使RNA,进而指导蛋白质的合成,非编码区位于编码区的上游及下游。
[4]在调控遗传信息表达的核苷酸序列中最重要的是位于编码区上游的RNA聚合酶结合位点。
RNA聚合酶是催化DNA转录为RNA,能识别调控序列中的结合位点,并与其结合。
真核生物基因结构见图1:图1 真核生物基因结构二.原核、真核生物基因结构的区别最主要的在于真核基因是不连续的,而原核基因是连续的。
所谓真核基因的不连续,即一个基因的编码序列也叫外显子,被一个或多个非编码序列,又叫内含子所间隔。
[5]这些内含子和外显子同属一个转录单位,转录形成前体。
经过转录的加工,即切去内含子,重新连按外显子,从而得到成熟。
而绝大多数的原核基因是连续的,没有内含子的间隔,转录产生成熟。
不仅如此,而且凡在代谢途径上功能有关的多个基因可能紧密相联,与它们的调控基因一起组成一个操纵子,转录到一条链。
第十章原核生物基因表达的调控

表 16-4 E.coliσ 因子识别不同保守序列的启动子 基因 分子量 70KD 32KD 24KD 54KD 28KD 功能 普遍 热休克 热休克 氮饥饿 产生鞭毛 -35 序列 TTGACA CCCTTGAA ? CTGGNA CTAAA 间隔(bp) 16~18 13~15 ? 6 15 -10 序列 TATAAT CCCGATNT ? TTGCA GCCGATAA
基本概念
1.操纵子(operon)
很多功能上相关的结构基因在染色体上串连排列,由 一个共同的控制区来操纵这些基因的转录。包含这些结构 基因和控制区的整个核苷酸序列就称为操纵子(operon)。
一个完整的操纵子主要包括启动子、操纵基因、结构 基因和终止子。
2. 阻遏物和激活物(reperssor and activator)
2. 基因表达的极性效应
•在正常情况下原核基因表达时,其转录出来的mRNA随 即进行翻译,这时整个mRNA都覆盖着核糖体, ρ因子 无法接近mRNA,而RNA聚合酶早已越过前面的基因的 依赖ρ因子的终止子,所以转录实际上并不停止,而是继 续转录后续基因。如果在某一基因的依赖于ρ的终止子之 前发生无义突变,核糖体便从无义密码子上解离下来,翻 译停止,于是ρ就可以自由进入RNA并移动,直到赶上停 留在终止子上的RNA聚合酶,结果使RNA聚合酶释放, 不能再转录下游基因。
第十章 原核生物基因 表达的调控
生物的遗传信息是以基因的形式储藏在细 胞内的DNA(或RNA)分子中的。随着个体 的发育,DNA有序地将遗传信息,通过转 录和翻译的过程转变成蛋白质,执行各种 生理生化功能,完成生命的全过程。从 DNA到蛋白质的过程,叫做基因表达 (gene expression),对这个过程的调节 就称为基因表达调控(gene regulation或 gene control)。
基因表达调控

• CAP是同二聚体 含有DNA结合区和 cAMP结合位点 (需生成cAMP-CAP复合物才有活性)
DNA
CAP结合位点
P
OZ YA
cAMP-CAP复合物
CCAACCPPAAPP CACPAP 无葡萄糖,cAMP浓度高时
CAP
有葡萄糖,cAMP浓度低时
2、真核生物的基因调节蛋白
反式作用因子(trans-acting factor) 通过与另一基因的顺式作用元件相
互作用,调节其表达的蛋白质。
这种调节作用称为反式作用。
DNA
a
mRNA
蛋白质A
A
反式调节
Ab
顺式作用因子(cis-acting factor) 通过与自身基因的顺式作用元件相互作用,
调节自身基因表达的蛋白质
(三)协调表达
(一)基本表达
❖ 管家基因(housekeeping gene): 在一个生物体的几乎所有细胞中持续表达 的基因。 无论表达水平高低,管家基因较少受环境因素影 响,而是在个体各个生长阶段的大多数或几乎全 部组织中持续表达,或变化很小。
❖ 分子遗传学家将这类基因表达被视为基本或组成性 基因表达(constitutive gene expression)。
二、基因转录激活调节基本要素
• 特异DNA序列(如:启动子) • 调节蛋白(如:转录因子) • DNA-蛋白质、蛋白质-蛋白质相互作用 • RNA聚合酶 基因表达调控本质:
通过 DNA-蛋白质相互作用来实现
(一)特异DNA序列
1、 原核生物的操纵子
操纵子(operon):通常由2个以上的编码序
列与启动序列、操纵序列以及其它调节序列
解释基因表达的调控机制。

解释基因表达的调控机制。
> 原题:解释基因表达的调控机制基因表达调控是指在细胞中控制基因转录和翻译的过程。
通过调控基因表达,细胞可以根据内外环境的需求来合成所需的蛋白质。
基因表达调控涉及多个环节和分子机制。
一、转录调控1. 转录因子:转录因子是一类可以与DNA结合的蛋白质,它们能够促进或抑制特定基因的转录。
转录因子的结合位点通常位于基因的启动子区域,它们可以通过调控转录复合物的形成来影响RNA聚合酶的结合和启动转录的过程。
2. 染色质修饰:染色质修饰是指对DNA及其相关的蛋白质进行化学修饰,从而改变染色质结构和可访问性。
例如,DNA甲基化可以抑制某些基因的转录,而组蛋白乙酰化则可以促进基因的转录。
二、转录后调控1. RNA剪接:RNA剪接是一种将RNA前体分子中的内含子去除,将外显子连结起来的过程。
通过不同的剪接方式,可以产生不同的mRNA亚型,从而影响蛋白质的翻译。
2. mRNA降解:mRNA降解是指将mRNA分解为较小的碎片,从而停止蛋白质的合成。
通过调控mRNA的稳定性,可以控制基因的表达水平。
三、翻译调控1. 转运调控:通过调控mRNA的转运过程,可以控制mRNA的定位和稳定性。
这种调控方式可以影响基因的表达水平。
2. 蛋白质修饰:蛋白质修饰是指在翻译后对蛋白质进行化学修饰的过程。
蛋白质修饰可以影响蛋白质的功能、稳定性和亚细胞定位。
综上所述,基因表达调控涉及转录调控、转录后调控和翻译调控等多个层面和分子机制。
这些调控机制相互作用,共同影响基因的表达水平和细胞的功能。
对这些调控机制的深入研究,有助于我们更好地理解生物体的发育、生长和适应环境的能力。
《分子生物学》教案
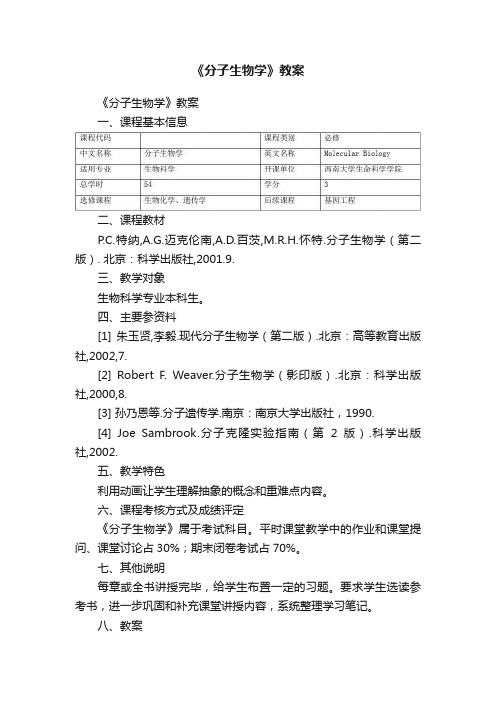
《分子生物学》教案《分子生物学》教案一、课程基本信息二、课程教材P.C.特纳,A.G.迈克伦南,A.D.百茨,M.R.H.怀特.分子生物学(第二版). 北京:科学出版社,2001.9.三、教学对象生物科学专业本科生。
四、主要参资料[1] 朱玉贤,李毅.现代分子生物学(第二版).北京:高等教育出版社,2002,7.[2] Robert F. Weaver.分子生物学(影印版).北京:科学出版社,2000,8.[3] 孙乃恩等.分子遗传学.南京:南京大学出版社,1990.[4] Joe Sambrook.分子克隆实验指南(第2版).科学出版社,2002.五、教学特色利用动画让学生理解抽象的概念和重难点内容。
六、课程考核方式及成绩评定《分子生物学》属于考试科目。
平时课堂教学中的作业和课堂提问、课堂讨论占30%;期末闭卷考试占70%。
七、其他说明每章或全书讲授完毕,给学生布置一定的习题。
要求学生选读参考书,进一步巩固和补充课堂讲授内容,系统整理学习笔记。
八、教案第一章绪论 Introduction(2学时)1、教学目标:掌握分子生物学的基本概念与研究内容;了解分子生物学发展简史和分子生物学的一些分支学科;了解分子生物学的发展趋势。
2、教学重点:分子生物学的产生及概念,分子生物学的研究内容3、教学难点:分子生物学的产生及概念,分子生物学的研究内容4、教学方法与手段:多媒体教学、自学与课堂讨论相结合5、教学过程第一节生命科学的回顾(20分钟)1、创世说与进化论;2、细胞学说;3、经典的生物化学和遗传学;4、DNA的发现。
第二节分子生物学的概念和研究内容(30分钟)1、什么是分子生物;2、分子生物学研究领域的三大原则;3、分子生物学研究领域。
第三节分子生物学发展简史(20分钟)1、孕育阶段;2、创立阶段;3、发展阶段。
第四节分子生物学实际应用的现状和展望。
(10分钟)第二章核酸的性质与结构(3学时)1、教学目标:掌握核酸的基本性质;掌握DNA的结构;掌握DNA分子变性、复性及分子杂交的原理。
分子生物学第十章

限制酶的发现
大肠杆菌的限制 — 修饰系统中,该系统由两种酶组成: 限制酶(分解和识别DNA的酶) 修饰酶(改变碱基结构,免遭限制酶分解)
甲基化修饰Am/Cm 两种酶作用于同一DNA的相同部位
(一)限制酶的命名
(1)以该菌株的属名的第一个字母及种名的头两个字母组 成三个斜体字母的略语表示 (2)同一种细菌若有不同菌株,则在属名种名后再加上株 名 (3)若一个菌株中有几种酶时,以罗马字加以区分 (4)同一属细菌种名头两个字母完全相同,其中一个菌的 酶可改用词头后的另一个字母来代替
• 一种质粒在一个细胞内存在的数目,称为质粒的拷贝数 • 质粒拷贝数由复制类型决定,并因此分为两类 1.严紧型质粒 复制与宿主染色体同步,拷贝数较少,1-3 个/细胞 2· 松弛型质粒 复制与宿主染色体不同步,可以单独复制,
拷贝数较多, 10-500个/细胞 • 在重组DNA技术中,为了提高目的DNA的扩增效率或目的基 因的表达效率,常采用松弛型质粒作为载体
个氨基酸
• ④多克隆位点,位于lacZ'编码区内 • ⑤大肠杆菌乳糖操纵子的调节基因lacI
pUC系列载体都是成对构建的。它们在结构上基本一致,只 是多克隆位点所含限制位点的排列顺序相反
2· 特点 和pBR322相比 • ①分子量更小,拷贝数更高,不用氯霉素处理就 可以在每个细胞内扩增500-700个拷贝 • ②所含的LacZ′适合于用α-互补和蓝白筛选技术
(二) pBR322载体
有两个抗药性基因,每 个抗性基因中均含有单克 隆位点,外源DNA插入后
使相应的抗性基因失活,
宿主细胞失去相应的抗药 性,不能在含相应抗生素 的培养基中生长,这一现 象称为插入失活。
质粒pBR322的抗性基因筛选示意图
分子生物学:真核基因表达调控习题与答案

第六章真核基因表达调控一、名词解释1.管家基因(housekeeping gene)2.可调节基因(regulated gene)3.顺式作用元件(cis-acting element)4.反式作用因子(trans-acting factors ) /转录因子(transcription factor, TF )5.基础转录因子(basal/general transcription factor)6.特异转录因子(special transcription factor)7.增强子9.锌指(zinc finger)结构10.亮氨酸拉链(leucine zipper)结构11.增强子12.基因表达13.RNAi14.miRNA15.SiRNA二、选择题1.关于基因表达调控的说法错误的是( )..A.转录起始是调控基因表达的关键B.环境因素影响管家基因的表达C.在发育分化和适应环境上有重要意义D.表现为基因表达的时间特异性和空间特异性E.真核生物的基因表达调控较原核生物复杂的多2.下列哪项属于可调节基因( )A.组蛋白编码基因B. 5s rRNA编码基因C.异柠檬酸脱氢酶编码基因D. 肌动蛋白编码基因E. 血红蛋白编码基因3.下列哪种染色质结构的变化不利于基因表达( ).A.组蛋白乙酰化B.核小体解聚C. CpG岛甲基化D.基因扩增E.染色质结构松散,对DNA酶I敏感4.下列哪项不属于真核生物基因的顺式作用元件( ).A.激素反应元件B.衰减子C.启动子D.沉默子E.增强子5.与RNA聚合酶相识别和结合的DNA片段是()A.增强子B.衰减子C.沉默子D.操纵子E.启动子6.下列哪项不参与调控真核细胞基因的特异性表达( ).A.反应元件B.特异转录因子C.增强子D.基础转录因子E.沉默子7.与原核生物相比较,真核生物的基因表达调控包括多个层次,下列哪项不是其调控复.杂性特有的分子基础( )A. 含有重复序列B. 断裂基因C. 转录与翻译分离D. 细胞内被膜性结构分隔形成多个区域E. 染色质结构8.能够与基础转录因子结合的是( )A.上游启动子元件B. TATA boxC.增强子D.反应元件E. Pribnow box9.有关基础转录因子的叙述,正确的是( )A.与非转录核心序列相结合B.决定基因表达的特异性C.其种类和数量在不同组织中差别很大D.辅助RNA聚合酶结合启动子E.在原核生物中的种类比真核生物少10.不属于特异转录因子的是( ).A. TF II DB. HSFC. APID.类固醇激素受体E. NF-K B11.关于特异转录因子的说法,正确的是( )A.调控管家基因的表达B.仅通过蛋白质一蛋白质相互作用进行调控C.仅通过DNA一蛋白质相互作用进行调控D.仅通过RNA一蛋白质相互作用进行调控E.起转录激活或者转录抑制作用12.锌指结构可能存在于下列哪种物质中( )A.阻遏蛋白B. RNA聚合酶C.转录因子D.端粒酶E.核酶13.下列哪种氨基酸在转录因子的转录激活结构域中含量丰富( )A. LysB. ArgC. AspD. HisE. Trp14.下列哪种因素不会影响真核细胞中mRNA的稳定性().A. 5'端帽子B. siRNAC. poly A尾D.去稳定元件E. miRNA15.小干扰RNA调节基因表达的机制是()A.封闭mRNA上的核蛋白体结合位点B.特异性降解靶mRNAC.形成局部双链,抑制靶mRNA的模板活性D. 使翻译出的蛋白质进入泛素化降解途径E.使翻译提早终止16.eIF-2对翻译起始具有重要的调控作用,下列哪项是它的活性形式()A.磷酸化B.脱乙酰化C.乙酰化D.脱磷酸化E. ADP-核糖基化17.不影响真核生物翻译起始的因素是( ).A. eIFB.帽子结合蛋白C. RNA编辑D. mRNA非翻译区的二级结构E. miRNA18.生物体在不同发育阶段,蛋白质的表达谱也相应变化,这主要取决于A.转录调控元件的差异B.翻译调控元件的差异C.基础转录因子的差异D. 特异转录因子的差异E. 翻译起始因子的差异19.下列哪项不是可调节基因的特点( ).A.组织特异性B.阶段特异性C.时间特异性D.空间特异性E.组成性表达20.下列哪一项是真核生物可调节基因的表达调控特有的机制( )A.基础转录因子B.衰减子C. RNA聚合酶D.增强子E.阻遏蛋白21.基础转录因子属于DNA结合蛋白,它们能够()A.结合转录核心元件B.结合增强子C.结合5’端非翻译区D. 结合3'端非翻译区E. 结合内含子22.特异转录因子不能够( ).A.结合RNA聚合酶B.结合基础转录因子C.结合其他特异转录因子D. 结合转录非核心元件E. 结合沉默子23.基因特异性表达的根本机制是( )A.顺式作用元件的种类不同B. RNA聚合酶活性的差异C.基础转录因子的质和量的差异D.特异转录因子的质和量的差异E.表达产物后加工过程的差异24.下列哪一类分子常具有亮氨酸拉链的结构特征( )A.生长因子B.酪氨酸蛋白激酶受体C. G蛋白D. 转录因子E. 丝/苏氨酸蛋白激酶25.管家基因的转录受哪些因素控制A. 基础转录因子B. 增强子C. 特异转录因子D. 启动子E. 反应元件26.真核生物的基因表达调控表现在A.转录水平B.翻译水平C.染色质水平D.转录后加工E.翻译后加工27.真核生物中,影响RNA聚合酶转录活性的因素包括A.启动子B.增强子C.基础转录因子D.衰减子E.特异转录因子28.真核生物基因表达的空间特异性的机制包括A.特异转录因子的种类不同B.同种特异转录因子的浓度不同C.特定组织的基因中存在组织特异性启动子D.特异转录因子的排列组合不同E.增强子等调控元件在不同组织的基因中分布不同29.转录因子的DNA结合结构域包含哪些结构类型A.螺旋一片层一螺旋B.锌指C.螺旋一转角一螺旋D.亮氨酸拉链E.螺旋一环一螺旋30.与siRNA相比较,miRNA的显著特点是A.单链B.在转录后水平发挥作用C.与靶mRNA碱基互补D.不降解靶mRNAE.个别碱基与靶mRNA序列不完全匹配31.转录因子的作用机制包括A. DNA—DNA相互作用B. DNA—RNA相互作用C. DNA一蛋白质相互作用D. RNA一蛋白质相互作用E.蛋白质一蛋白质相互作用32.真核生物的基因转录涉及哪些物质的相互作用A. operatorB. cis-acting elementC. polysomeD. trans-acting factorE. RNA polymerase33.在同一个体的不同组织中A.基因的表达谱不同B.基因组结构不同C.特异转录因子的种类不同D.存在的蛋白质的种类不同E.特异性启动子的种类不同34.真核生物独有的转录调控机制涉及A.启动子B.增强子C.转录因子D.组蛋白E. SD序列35.生物对环境的适应性表现在A.基因变异B.合成不同种类的mRNAC.合成不同种类的蛋白质D. 产物的反馈抑制E. 蛋白质活性的快速调节36.下列哪些情况对于真核生物的基因转录具有调控作用A.反式作用因子的磷酸化B.类固醇激素与胞内受体结合C.特定DNA序列的甲基化D.组蛋白的乙酰化E.蛋白质因子的羟基化37.管家基因的含义是A.在各组织细胞中都表达B.在特定的组织细胞中表达C. 在不同发育阶段都表达D. 在特定的发育阶段表达E.表达程度在不同时空条件下差异显著38.关于特异转录因子的描述,正确的是A.在所有组织细胞中组成性表达B.在不同组织细胞中存在的种类不同C. 在不同组织细胞中的浓度不同D. 调控管家基因的转录E.是真核生物基因表达特异性的根源所在三、简答题1.简述顺式作用元件与反式作用因子对基因表达调控的影响。
真核生物DNA水平上的基因表达调控

研究基因调控主要应回答3个问题:
① 什么是诱发基因转录的信号? ② 基因调控主要是在哪一步(模板DNA的转录、mRNA
的成熟或蛋白质合成)实现的? ③ 不同水平基因调控的分子机制是什么?
第一节:真核生物的基因结构与转录活性
概述
目录
第一节:真核生物的基因结构与转录活性
第二节:真核基因的转录
第三节:反式作用因子
第四节:真核基因转录调控的主要模式
第五节:其他水平上的基因调控
概述
真核生物和原核生物由于基本生活方式不同所决定基因表达调控上 的巨大差别。
原核生物的调控系统就是要在一个特定的环境中为细胞创造高速生 长的条件,或使细胞在受到损伤时,尽快得到修复,所以,原核生 物基因表达的开关经常是通过控制转录的起始来调节的。
酶,另一种是从头合成型甲基转移酶,前者主要在甲基化母链(模板链) 指导下使处于半甲基化的DNA双链分子上与甲基胞嘧啶相对应的胞嘧啶甲 基化。该酶催化特异性极强,对半甲基化的DNA有较高的亲和力,使新生 的半甲基化DNA迅速甲基化,从而保证DNA复制及细胞分裂后甲基化模式 不变。后者催化未甲基化的CpG成为mCpG,它不需要母链指导,但速度 很慢。
1.“开放”型活性染色质(active chromatin) 结构对转录的影响
真核基因的活跃转录是在常染色质上进行的。转录发生之 前,染色质常常会在特定的区域被解旋松弛,形成自由DNA。 这种变化可能包括核小体结构的消除或改变,DNA本身局部结 构的变化等,这些变化可导致结构基因暴露,促进转录因子与 启动区DNA的结合,诱发基因转录。
一、基因家族 二、真核基因的断裂结构 三、真核生物DNA水平上的基因表达调控 四、DNA 甲基化与基因活性的调控
生物化学(本科)第十章-基因表达调控及其相关细胞信号转导通路-随堂练习与参考答案

生物化学(本科)第十章基因表达调控及其相关细胞信号转导通路
随堂练习与参考答案
第一节概述第二节原核基因的转录调控第三节真核基因的转录调控第四节相关细胞信号转导通路
1. (单选题)基因表达调控的基本控制点是( )
A. 基因结构活化
B. 转录起始
C. 转录后加工
D. 蛋白质翻译及翻译后加工
E. mRNA从细胞核转运到细胞浆
参考答案:B
2. (单选题)分解代谢物基因激活蛋白(CAP)对乳糖操纵子表达的影响是( )
A. 正性调控
B. 负性调控
C. 正性调控、负性调控都可能
D. 无调控作用
E. 可有可无
参考答案:A
3. (单选题)阻遏蛋白识别操纵子中的( )
A.启动基因
B.结构基因
C.操纵基因
D.内含于
E.外显子
参考答案:C
4. (单选题)目前认为基因表达调控的主要环节是( )
A. 翻译后加工
B. 转录起始
C. 翻译起始
D. 转录后加工
E. 基因活化
参考答案:B。
分子生物学名词解释

分子生物学名词解释第十章DNA的生物合成1、半保留复制:复制时,母链双链DNA解开成两股单链,各自作为模板指导子代合成新的互补链。
子代细胞的DNA双链,其中一股链从亲代完整的接受过来,另一股单链则完全重新合成。
由于碱基互补,两个子细胞的DNA双链,都和亲代母链DNA碱基序列一致。
这种复制方式称为半保留复制。
2、半不连续复制:领头链连续复制,随从链不连续复制,这就是复制的半不连续性。
3、双向复制:复制时,DNA从起始点向两个方向解链,形成两个延伸方向相反的复制叉,称为双向复制。
4、冈崎片段:DNA复制时,随从链形成的不连续片段。
5、复制子:是独立完成复制的功能单位,从一个DNA 复制起点起始的DNA复制区域称为复制子。
6、引发体:复制起始时,原核生物由解链酶、DnaC、DnaG、结合到DNA复制起始区域形成的复合结构,叫引发体。
7、领头链:DNA复制时,顺着解链方向生成的子链,复制是连续进行的,叫做领头链。
8、随从链:DNA复制时,不能顺解链方向连续复制,复制方向与解链方向相反的子链叫做随从链。
9、端粒:指真核生物染色体线性DNA分子末端的结构,由末端DNA序列和蛋白质构成。
10、框移突变:指由于核苷酸的插入或缺失突变引起的三联体密码的阅读方式改变,造成蛋白质氨基酸排列顺序发生改变,其后果是翻译出的蛋白质可能完全不同。
11、引物:是由引物酶催化合成的短链RNA分子。
12、逆转录:以RNA为模板在逆转录酶的作用下合成双链DNA的过程。
第十一章RNA的生物合成1、转录:生物体以DNA为模板合成RNA的过程称为转录。
2、结构基因:基因组中,能转录出RNA的DNA区段。
3、不对称转录:在双链DNA分子上,一股链用作模板,另一股链不转录;模板链并非永远在同一条DNA单链上。
4、TATA盒:基因的转录起始点上游多具有典型的TATA序列,通常认为是启动子的核心序列。
5、Pribnow盒:原核生物中,在起始密码子上游有一个由5-6个核苷酸组成的共有序列,以其发现者的名字命名为Pribnow盒,这个框的中央位于起点上游10bp处,所以又称—10序列,是转录的解旋功能部位,一般较保守。
真核基因表达调控

子遗传学的奠基石。
Gregor Mendel (1822-1884).
The Father of Genetics
在孟德尔遗传学基础上, Morgan 又提出了 基因学说。 1910年,Morgan和他的助手们发现了第一只 白眼雄果蝇,称为突变型。正常情况下,果蝇
都是红眼的,称为野生型。Morgan将白眼雄果
Actually, they had. That morning, Watson and
Crick
had
figured
out
the
structure
of
deoxyribonucleic acid, DNA. And that structure —
a "double helix" that can "unzip" to make copies
控制遗传信息流动的基本机制——RNA干扰 方面的杰出贡献而获得诺贝尔生理医学奖。
1928 年,英国科学家 Griffith 等人发现,具有
光滑外表的S型肺炎链球菌能使小鼠发病,具有 粗糙外表的R型细菌没有致病力。荚膜多糖能保
护细菌免受动物白细胞的攻击。
美国著名的微生物学家 Avery首先用实验证明 基因就是DNA分子。他将光滑型致病菌(S型) 烧煮杀灭活性以后再侵染小鼠,发现这些死细 菌自然丧失了致病能力。
• 1993 年,美国科学家 Roberts 和 Sharp 因发
现 断 裂 基 因 ( introns ) 而 获 得 Nobel 奖 ; Mullis 由 于 发 明 PCR 方 法 而 与 加 拿 大 学 者 Smith ( 第 一 个 设 计 基 因 定 点 突 变 ) 共 享 Nobel化学奖。
分子生物学:原核基因表达调控模式

添加葡萄糖后,细菌所需要的能量便可从葡萄糖得到 满足,葡萄糖是最方便的能源,细菌无需开动一些不 常用的基因去利用这些稀有的糖类。
葡萄糖的存在会抑制细菌的腺苷酸环化酶活性,减少
环腺苷酸(cAMP)的合成,与它相结合的蛋白质,
即 环 腺 苷 酸 受 体 蛋 白 CRP 又 称 分 解 代 谢 物 激 活 蛋 白 CAP,因找不到配体而不能形成复合物。
负控诱导 阻遏蛋白不与效应物(诱导物)结合时,结 构基因不转录;与之结合则转录。
负控阻遏 阻遏蛋白与效应物结合时,结构基因不转录。 阻遏蛋白作用的部位是操纵区。
在正转录调控系统中,调节基因的产物是激活蛋 白(activator)。
正控诱导系统 效应物分子(诱导物)的存在使激活蛋白 处于活性状态;
葡萄糖 cAMP Lac操纵子被抑制
DNA
+ + + + 转录
CAP P O Z Y A
CAP CAP CAP CAP 无葡萄糖,cAMP浓度高时
CAP
有葡萄糖,cAMP浓度低时
协调调节
负性调节与正性调节协调合作
阻遏蛋白封闭转录时,CAP不发挥作用 如没有CAP加强转录,即使阻遏蛋白从P上解聚仍无转录活性
23
• 乳糖操纵子的控制模型,其主要内容如下:
① Z、Y、A基因的产物由同一条多顺反子的mRNA分子所编码。 ② 这个mRNA分子的启动子紧接着O区,而位于I与O之间的启动子区(P), 不能单独起动合成β-半乳糖苷酶和透过酶的生理过程。 ③ 操纵基因是DNA上的一小段序列(仅为26bp),是阻遏物的结合位点。 ④当阻遏物与操纵基因结合时,lac mRNA的转录起始受到抑制。 ⑤诱导物通过与阻遏物结合,改变它的三维构象,使之不能与操纵基因结 合,从而激发lac mRNA的合成。当有诱导物存在时,操纵基因区没有被阻 遏物占据,所以启动子能够顺利起始mRNA的合成。
简述真核生物基因表达调控过程

简述真核生物基因表达调控过程真核生物基因表达调控过程是指在真核生物细胞中,如何通过一系列的调控机制,将基因中的遗传信息转化为蛋白质,以实现细胞功能的正常发挥。
基因表达调控过程可以分为转录调控和转录后调控两个阶段。
在转录调控阶段,首先是在细胞核中进行转录。
细胞核中的DNA被RNA聚合酶酶识别并解链,形成单链mRNA。
但并不是所有基因都会被转录,细胞会根据需要选择性地进行转录。
这是通过转录因子的作用来实现的。
转录因子是一类能够与DNA特定序列结合的蛋白质,它们能够促进或抑制转录的进行。
转录因子的结合位点位于启动子区域,当转录因子结合到启动子区域时,会引发一系列的反应,包括启动RNA聚合酶的活性和引导其结合到合适位置上,从而促使转录的进行。
转录因子的表达受到多种因素的调控,如细胞内的信号分子、细胞周期等。
转录后调控是指在mRNA合成后,通过一系列的调控机制来决定其在细胞中的命运。
mRNA在合成后需要经过剪接、修饰和运输等过程。
剪接是指将mRNA中的内含子去除,将外显子进行连接的过程。
通过剪接的不同方式,可以生成不同的mRNA亚型,从而在翻译过程中产生不同的蛋白质。
修饰是指在mRNA上加上帽子和尾巴等化学修饰,这些修饰可以保护mRNA不被降解,并帮助mRNA与翻译机器结合。
运输是指mRNA离开细胞核,进入到细胞质中,进一步参与翻译过程。
这个过程受到RNA结合蛋白的调控。
在翻译过程中,mRNA被核糖体识别并翻译成蛋白质。
这个过程也受到多种调控机制的影响。
一方面,mRNA上的启动子序列会影响翻译的起始位置,从而决定蛋白质的翻译起始位点。
另一方面,mRNA的稳定性也会影响翻译的效率和蛋白质的表达水平。
mRNA 的稳定性受到RNA结合蛋白和非编码RNA的调控。
总的来说,真核生物基因表达调控过程是一个复杂而精细的调控网络。
通过转录调控和转录后调控的相互作用,细胞可以根据内外环境的需要,在不同的时空位置上产生不同类型的蛋白质,以实现细胞功能的正常发挥。
分子生物学-真核生物基因表达调控

3 基因重排与交换
将一个基因从远离启动子的地方移到距它很
Hale Waihona Puke 近的位点从而启动转录,这种方式称为基因 重排。
通过基因重排调节基因活性的典型例子是免
疫球蛋白和T-细胞受体基因的表达。
V、C和J基因片段在胚胎细胞中相隔较远。编码产生免疫球蛋白的细胞发 育分化时,通过染色体内DNA重组把4个相隔较远的基因片段连接在一起, 从而产生了具有表达活性的免疫球蛋白基因。
发育早期:只有一个着丝点行使功能,
从头合成型甲基转移酶:催化未甲基化的CpG成 为mCpG
基因丢失
在细胞分化过程中,可以通过丢失掉某些基
因而去除这些基因的活性。某些原生动物、 线虫、昆虫和甲壳类动物在个体发育中,许 多体细胞常常丢失掉整条或部分的染色体, 只有将来分化产生生殖细胞的那些细胞一直 保留着整套的染色体。
一.
基因丢失: 在细胞分化过程中,某些原生动物、线虫 、昆虫等体细胞通过丢失某些基因而除去 这些基因的活性。 马蛔虫:只有一对染色体,染色体上有许 多着丝点。
假基因
是基因组中因突变而失活的基因,无蛋白质产
物。
一般是启动子出现问题。
8.2 DNA水平的基因表达调控
1染色质水平的调节:“开放”型活性染色质
(activechromatin)结构对转录的影响
2基因扩增
3基因重排与交换
4
DNA甲基化与基因活性的调控
1 染色质状态对基因表达的调控
能相关的基因,这些基因成套组合称为基因家族。 如:编码组蛋白、免疫球蛋白和血红蛋白的基因都 属于基因家族 同一家族中的成员有时紧密地排列在一起,成为 一个基因簇(gene cluster) 。
1、简单多基因家族
真核生物基因表达的调控

第10章真核生物基因表达的调控本章教学要求1.熟悉真核基因组的结构特点、真核生物在DNA水平、转录水平和翻译水平上基因表达调控的特点。
2.掌握以下概念:顺式作用元件、反式作用因子、启动子、增强子,熟悉沉默子、基本转录因子、特异转录因子。
3.了解转录因子的结构特点。
本章教学重点和难点1、真核生物在DNA水平和转录水平基因表达调控的特点。
2、转录因子的结构特点。
教学方法与手段讲授与交流互动相结合,采用多媒体教学。
授课内容10.1 真核生物基因表达调控的特点和种类一、真核生物基因表达调控的特点原核生物的调控系统就是要在一个特定的环境中为细胞创造高速生长的条件,或使细胞在受到损伤时,尽快得到修复,所以,原核生物基因表达的开关经常是通过控制转录的起始来调节的。
真核基因表达调控的最显著特征是能在特定时间和特定的细胞中激活特定的基因,从而实现"预定"的、有序的、不可逆转的分化、发育过程,并使生物的组织和器官在一定的环境条件范围内保持正常功能。
真核生物基因表达调控与原核的共同点:•基因表达都有转录水平和转录后的调控,且以转录水平调控为最重要;•在结构基因上游和下游、甚至内部存在多种调控成分,并依靠特异蛋白因子与这些调控成分的结合与否调控基因的转录。
真核生物基因表达调控与原核的不同点:1、真核基因表达调控的环节更多:转录与翻译间隔进行,具有多种原核生物没有的调控机制;个体发育复杂,具有调控基因特异性表达的机制。
2、真核生物活性染色体结构的变化对基因表达具有调控作用:DNA拓扑结构变化、DNA 碱基修饰变化、组蛋白变化;3、正性调节占主导,且一个真核基因通常有多个调控序列,需要有多个激活物。
二、真核生物基因表达调控的种类根据其性质可分为两大类:一是瞬时调控或称为可逆性调控,它相当于原核细胞对环境条件变化所做出的反应。
瞬时调控包括某种底物或激素水平的升降,及细胞周期不同阶段中酶活性和浓度的调节。
二是发育调控或称不可逆调控,是真核基因调控的精髓部分,它决定了真核细胞生长、分化、发育的全部进程。
真核生物基因表达调控

真核生物基因表达调控第十章作业1. 简述真核生物基因表达调控的7个层次。
①染色体和染色质水平上的结构变化与基因活化②转录水平上的调控,包括基因的开与关,转录效率的高与低③RNA加工水平的调控,包括对出事转录产物的特异性剪接、修饰、编辑等。
④转录后加工产物在从细胞核向细胞质转运过程中所受到的调控⑤在翻译水平上的控制,即对哪一种mRNA结合核糖体进行翻译的选择以及蛋白质成量的控制⑥对蛋白质合成后选择性地被激活的控制,蛋白质和酶分子水平上的剪接等的控制⑦对mRNA选择性降解的调控2. 真核基因表达调控与原核生物相比有何异同?相同点:①与原核基因的调控一样,真核基因表达调控也有转录水平调控和转录后水平的调控,并且也以转录水平调控为最重要;②在真核结构基因的上游和下游(甚至内部)也存在着许多特异的调控成分,并依靠特异蛋白因子与这些调控成分的结合与否调控基因的转录。
不同点:①原核细胞的染色质是裸露的DNA,而真核细胞染色质则是由DNA与组蛋白紧密结合形成的核小体。
②在原核基因转录的调控中,既有激活物参与的正调控,也有阻遏物参与的负调控,二者同等重要。
③原核基因的转录和翻译通常是相互偶联的,即在转录尚未完成之前翻译便已开始。
④真核生物大都为多细胞生物,在个体发育过程中发生细胞分化后,不同细胞的功能不同,基因表达的情况也就不一样,某些基因仅特异地在某种细胞中表达,称为细胞特异性或组织特异性表达,因而具有调控这种特异性表达的机制。
3. DNA 甲基化对基因表达的调控机制。
甲基化抑制基因转录的机制:DNA甲基化会导致某些区域DNA 构象改变,包括甲基化后染色质对于核酸酶或限制性内切酶的敏感度下降,更容易与组蛋白H1相结合,DNaseⅠ超敏感位点丢失,使染色质高度螺旋化, 凝缩成团, 直接影响了转录因子与启动区DNA的结合效率的结合活性,不能启始基因转录。
DNA的甲基化不利于模板与RNA聚合酶的结合,降低了转录活性。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第三节 基因重排调节
沙门氏菌鞭毛蛋白由不同基因编码: H1基因表达产生H1型鞭毛蛋白细菌处于I相 H2基因表达产生H2型鞭毛蛋白细菌处于II相 I相和II相细菌在细胞分裂时,以1/1000概率产生另一相的后代,称为相 变。
H1基因和H2基因位于染色体上不同区域。H2基因与另一个编码H1阻遏 蛋白的基因rh1紧密连锁,两个基因协同表达。沙门氏杆菌的相取决于 H2-rh1是否表达。
RNA干扰(RNAi)的影响
• RNA干扰是外源或内源性双链RNA(dsRNA)触发同源mRNA 特异性降解,从而使相应基因沉默。(反义RNA是利用完全互 补的RNA与同源性mRNA杂交,封闭mRNA,阻断基因的表 达。)
• RNAi的重要特征包括Dicer切割产生小分子RNA和RNA诱导的 沉默结合(RNAinduced silencing complex, RISC)的形成。 RISC可以导致转录或转录后水平的基因沉默。
ppGpp的生成:
• 氨基酸缺乏时,刺激relA基因表达。relA编码严谨因子( stringent factor ,SF )。
• SF位于核糖体上,催化ATP转移焦磷酸给GTP或GDP,分别形成 pppGpp或ppGpp。
(p)ppGpp的作用:可能是严紧控制的关键
• 抑制rRNA和tRNA的转录 • 降低RNA聚合酶在转录合成mRNA时的延伸速度
• 低渗透压时,omp F合成增多, omp C合成受抑制。
• omp C基因转录时,omp C上游有一段DNA序列,以相反的方向同时 转录一个174核苷酸的RNA,这RNA能与omp F的前导序列中44个核 苷酸(包括SD序列)以及编码区域(包括起始密码子AUG)形成杂合 双链,从而抑制了omp F mRNA的翻译。
• RNA干扰机制
Dicer:将dsRNA 精确切割成siRNA(Small interfering RNA (siRNA)的酶. RISC:RNA 诱导的沉默复合物 (RNA - induced Silencing Complex ,RISC)
microRNA(miRNA)
• MicroRNA(miRNA)是近几年在真核生物中发现的一类内源性的具 有调控功能的码RNA,其大小长约20~25 nt,主要发挥基因转录后 水平调控作用。
• 仅仅依靠弱化作用,色氨酸的存在可以使转录水平抑制10倍,色氨 酸阻抑蛋白的作用可以抑制70倍,合在一起可抑制700倍。在His操 纵子中,弱化作用是唯一反馈调控机制。
第二节 严谨反应
当细菌发现它们处于很差的生长环境,缺乏足够的氨基酸来维持蛋白质 合成时,它们停止大部分代谢活动来保存能源,这种反应称为严紧反应 (或应急反应,stringent response) 严紧反应时,细胞产生两种非正常的核苷酸,在电泳时呈现两个特殊斑 点,称为魔点I和魔点II 两斑点分别为ppGpp(鸟苷四磷酸)和pppGpp (鸟苷五磷酸)
不同物种miRNA具有保守性
第十一章 真核 生物表达调控
真核生物染色质被包裹在细胞核内,基因的转录(核内)和翻译 (细胞质内)被核膜所隔开,核内RNA的合成与转运,细胞质中 RNA的剪接和加工等都属于真核生物基因调控的范围。
• 反义RNA的抑制机理:
• 细胞核中:反义RNA和正链RNA结合,形成双链,阻止正义 RNA的加工或转运。
• 细胞质中:反义RNA通常和mRNA5’端形成双链,抑制其翻译。
反义RNA的抑制机理:
• 反义RNA调节E.coli外膜蛋白变化
• 渗透压变化后E.coli外膜蛋白的变化
• 高渗透压时,omp C合成增多, omp F合成受抑制;
蛋白质合成的自体调控
• RF2合成的自体调控
…CUU U GAC 25 26
• RF2充分时,核糖体A位到达第25个密码子后面的UGA时,RF2促成 肽链合成的终止。(不完全肽链,无RF2活性)。
• RF2不足时,核糖体向前划过一个核苷酸,继续合成第26个氨基酸, 直到此基因的最后的终止密码子。
• 核糖体蛋白合成的自体调控
• 在色氨酸丰富时,核糖体迅速翻译直接到达前导肽末端。由于核糖 体封闭了序列2,3-4发夹得以形成终止子结构,转录可能被终止, 转录不能进行到结构基因区域,结构基因的转录被关闭。这一过程 称为弱化作用。
• 在色氨酸缺乏时,由于缺乏色氨酸氨酰-tRNA,核糖体在序列1内停 滞,封闭了序列1,序列2-3形成发夹结构,终止子发夹不能形成, 转录可以继续到trpE及其下游。
• 核糖体蛋白基因组成若干个操纵子。它们的表达与rRNA相协调是通 过调节蛋白实现的,这种调节蛋白都是核糖体蛋白本身,而且都是核 糖体上与rRNA结合的蛋白。
• mRNA与调节蛋白相结合的序列与rRNA与该蛋白结合的序列有很大 的同源性,二者都能与起调控作用的核糖体蛋白相结合,但结合能力 rRNA>mRNA。
• 当细胞中有游离rRNA时,调节蛋白优先和rRNA结合;当rRNA被饱 和后,多余的调节蛋白就与mRNA上的结合位点结合,这些结合位点 靠近或包含SD序列,阻碍mRNA的翻译。
反义RNA( antisense RNA )的调控
• 反义RNA也叫:干扰mRNA的互补RNA(mic-RNA, mRNAinterfering complementary RNA )。
原核生物转录 后调控
第一节色氨பைடு நூலகம்操纵子
结构基因:包括色氨酸合成途径中相关的5个结构基因trpE 、trpD、 trpC 、trpB、 trpA。
启动子:Ptrp ,控制所有5个结构基因的表达。
弱第化作一用(节att色enu氨atio酸n)操纵子
• 在大肠杆菌中转录和翻译是紧密偶联的。前导肽编码序列位于序列1 内,两个色氨酸密码子位于互补序列1内,终止密码子位于1和2之间。
H2-rh1是否表达受其上游一段995bp核苷酸DAN序列的排列方向所控制。 H2基因启动子位于该序列末端,995bp序列中有一个hin基因,其蛋白 产物可催化995序列的倒位。当启动子序列朝向H2基因时,H2-rh1表达; 倒位后,启动子背离H2基因,H2-rh1不表达,H1基因表达。
第四节 翻译水平调控