转录后加工
简述rna转录后加工过程

简述rna转录后加工过程摘要:1.RNA转录后加工过程的概述2.RNA转录后加工的主要步骤a.剪接b.剪切c.RNA编辑d.RNA降解3.各步骤的功能和意义4.实例分析5.RNA转录后加工在生物体中的作用6.研究RNA转录后加工的意义和前景正文:在我们生物体内,基因通过转录过程将DNA信息转化为RNA,但这只是RNA生命历程中的第一步。
接下来,RNA要经历一系列复杂的加工过程,才能最终发挥其生物学功能。
这个过程被称为RNA转录后加工。
RNA转录后加工的主要步骤包括剪接、剪切、RNA编辑和RNA降解。
剪接是指将RNA前体分子中的内含子去除,并将外显子连接成成熟的RNA分子。
这一过程通过特定的酶家族,如剪接酶,来实现。
剪切是指在RNA分子的3"端添加poly(A)尾巴,这是几乎所有真核生物RNA的共同特征。
RNA编辑则是指在RNA分子上发生碱基改变,这一过程依赖于特定的编辑酶和相应的底物。
最后,RNA降解是指RNA分子在细胞内的分解过程,这对于调控RNA水平和维持细胞内稳态至关重要。
这些加工过程对于RNA最终的生物学功能具有重要意义。
以剪接为例,它能消除RNA前体中无功能的RNA片段,使成熟的RNA更具特异性和高效性。
同时,RNA编辑能够改变RNA的序列,从而影响其翻译效率和稳定性。
在生物体中,RNA转录后加工涉及多种生物过程,如基因表达调控、病毒复制和免疫反应等。
对RNA转录后加工的研究,有助于我们深入了解生命过程中的基因表达调控机制,为治疗疾病和开发新型药物提供理论依据。
随着生物科学技术的不断发展,对RNA转录后加工的研究将越来越深入。
分子生物学 第三章 (下)转录后加工

5’m7pppNmNUUUU 寡聚 U 区 ds 单一序列 ds RNA RNA
AAAAA„„3’ Poly (A)
图 13-
hnRNA 的结构模型
hnRNA的物理结构是核 糖核蛋白颗粒 (hnRNP),颗粒中蛋白 质包围着hnRNA。体外 研究表明hnRNA的形状 是一个球体和一个与之相 连的纤维状结构。
第三章 (下) 转录后加工
转录后的加工(posttranscriptional modification) (1)减少部分片段:如切除5′端前导序列, 3′端拖尾序列和中部的内含子; (2)增加部分片段:5′加帽,3′加poly(A), 归巢和通过编辑加入一些碱基; (3)修饰:对某些碱基进行甲基化等。
真核tRNA的基因和原核不同: (1)真核的前体分子tRNA是单顺反子,但成 簇排列,基因间有间隔区; (2)真核tRNA基因一般都比原核tRNA基因多 得多,如酵母约有400个tRNA基因; (3) 5′端单磷酸核苷酸,表明已被加工过; (4) tRNA的前体分子中含有内含子。
真核tRNA内含子切除的特点:
真核细胞中rRNA的加工途径
(1) 切除5′端的前导序列; (2) 从41S的中间产物中先切下18S的片段。 Hela细胞的切点在18S和5.8S之间的ITS (内部转录间隔序列); L细胞的切点在 18S序列和ITS的交界处,称为先成熟。 (3) 部分退火,形成发夹结构; (4) 最后修正。
剪切位点
第一节tRNA和rRNA的加工
一. 原核的 tRNA和 rRNA的加工 参与蛋白质合成的 tRNA 和 rRNA 都不是最初 的转录产物 (1) 它们的5′端都是单磷酸,而原始的转 录 产物5′应是三磷酸; (2) 分子比初始转录物小; (3) tRNA含有特殊的碱基,这些碱基只有通 过一些化学修饰才可以得到。
转录后加工名词解释

转录后加工名词解释
转录后加工是指在基因组中进行转录的过程后,对转录产物(RNA分子)进行进一步的修饰和加工的过程。
转录是指在DNA模板上合成RNA分子的过程,而转录后加工则是在RNA分子合成完成后对其进行一系列的修饰和处理。
转录后加工的目的是为了产生成熟的RNA分子,使其能够发挥特定的功能。
在转录后加工过程中,RNA分子经历剪接、修饰和运输等多个步骤,以形成成熟的RNA分子。
剪接是转录后加工中最重要的步骤之一。
在剪接过程中,RNA 分子的内含子(非编码区域)会被剪除,而外显子(编码区域)则会被保留下来。
这样一来,通过剪接,一个基因可以产生多个不同的成熟RNA分子,从而扩大了基因的功能和多样性。
除了剪接,转录后加工还包括其他的修饰过程。
例如,RNA分子可能会经历5'端帽子的添加和3'端的聚腺苷酸尾巴的加入,这些修饰可以保护RNA分子免受降解,并有助于其在细胞内的稳定性和转运过程中的识别。
此外,转录后加工还可以包括RNA编辑、互补RNA合成和核糖体扫描等过程。
RNA编辑是指在转录后,RNA分子中的碱基序列可以发生改变,从而导致RNA分子的信息内容发生变化。
互补RNA合成是指利用RNA分子作为模板合成互补的DNA分子。
核糖体扫描是指RNA分子被核糖体识别并翻译成蛋白质的过程。
总的来说,转录后加工是一系列对转录产物进行修饰和加工的过程,通过这些过程,RNA分子可以获得特定的功能和稳定性,从而发挥其在细胞中的重要作用。
第8章 转录后加工

4、拼接(splicing)
Ø 大多数的真核生物基因是断裂基因;
Ø 其中编码序列称为外显子(exon),外显子之间的 介入序列称为内含子(intron);
Ø 少数真核生物基因(如组蛋白、干扰素)是连续的;
Ø 高等真核生物的基因中多数内含子比外显子长得多, 而低等真核生物(如酵母)的基因中内含子比较短而 且少见;
Ø 有些生物的rRNA前体含有内含子,需要拼接;
p.205
Ø 哺乳动物的18S, 28S, 5.8S rRNA gene 组成一个 转录单位,由RNA pol I 转录产生45S的前体分子;
Ø 5S rRNA gene 与不转录区域组成转录单位, 由RNA pol III转录;
small nucleolar RNA(snoRNA)
Ø 高度精确; Ø 依赖于多种顺式作用元件和反式作用因子; Ø 共转录事件;
顺式元件1
Ø 内含子具有一致的保守序列,即5’拼接点为 GU,3’拼接点为AG,称为BreathnathChambon规则,也称GU/GT-AG规则。
顺式元件2
为什么只有mRNA被加帽?
Ø 只有RNA聚合酶 II 合成的转录产物(mRNA、 部分snRNA)才有帽子结构;
Ø 因为加帽酶只能与RNA聚合酶 II 的CTD结构 域结合;而CTD是RNA聚合酶II 特有的。
Ø 加帽酶与CTD的磷酸化形式(延伸型)结合。 Ø 转录产物一旦从RNA聚合酶II中显露出来,就
可以与加帽酶接触。
2、3’端加尾
Ø 真核生物的大多数mRNA及其前体在3’端有约 250 nt 的连续的AMP。 Ø poly(A) 由poly(A) polymerase(PAP)添加; Ø mRNA进入细胞质后,其poly(A)可以被更新 : 不断地被RNase降解,再由细胞质中的PAP重新 合成。
rna转录后加工方式

rna转录后加工方式
RNA转录后加工(RNA post-transcriptional processing)是指在RNA分子合成之后,在细胞中对其进行修饰和修剪的过程。
这些加工方式可以使原始RNA分子成熟,并使其具有功能性。
以下是几种常见的RNA转录后加工方式:
剪接(Splicing):在真核生物中,基因的转录产物(前体mRNA)经过剪接过程,去除其中的内含子(intron),保留外显子(exon),从而形成成熟的mRNA分子。
剪接是通过剪接体(spliceosome)来完成的,其中包括snRNPs等辅助因子。
5'端修饰:RNA的5'端通常经过加上7-甲基鸟苷(7-methylguanosine)和三磷酸核苷酸链(PPP 链)的修饰,形成5'甲基鸟苷帽(5' cap)。
这个帽子在RNA稳定性、转运和翻译起重要作用。
3'端修饰:RNA的3'端通常经过加上聚腺苷酸(polyadenylation)的修饰。
这个poly(A)尾巴有助于RNA的稳定性、转运和翻译,并参与转录终止的过程。
RNA编辑:在一些生物体中,RNA的序列可以通过RNA编辑(RNA editing)进行改变。
这种编辑通常涉及碱基的替换、插入或删除,从而改变RNA的编码能力和功能。
RNA修饰:RNA分子可能会经历各种修饰,如甲基化、脱氨基、糖基化等。
这些修饰可以增强RNA的稳定性、调节翻译和识别,以及影响RNA的功能。
RNA转录后加工是一个复杂而精确的过程,它可以使原始的转录产物转化为功能性的RNA 分子。
这些加工方式对于基因表达调控和细胞功能起着重要的作用。
真核生物转录后的加工
1、核mRNA内含子剪接位点特征
内含子总是由GU开始,以AG结束,其规律称为GU-AG法则 (GU-AG rule) 或Chambon法则。
5´端剪接位点(供位)相邻的保守序列:5´-AG↓GUPuAGU-3´
分枝点保守序列:Py80NPy87Pu75APy95,其中A为百分之百保 守,且具有2′-OH。 3´端剪接位点(纳位)相邻的保守序列:5´-(Py)nNCAG-3´ mRNA前体正确剪接所必需的
剪接体解体与套索降解同步
4、具体的剪接机制
U1通过与 5´剪接点互补而结合
U2AF与 3´剪接点内含子结合
U2 识别并结合分支点 A ,并在 SF1 和 BBP 帮 助 下 使 内 含 子 的 5´端和 3´端带到一起
U4/U5/U6复合物与U1/U2结合
续上页
U1脱离
U4脱离,U6与U2间发生第一 次转酯反应,套索结构形成
第二次转酯反应,U2/U5/U6 与套索结构结合
成熟的mRNA释放
四、其他的内含子剪接方式
1、内含子的分类
根据基因的类型和剪接的方式,通常把内含子分为 四类。
I类:线粒体、叶绿体及低等真核生物细胞核的rRNA基因; II类:线粒体、叶绿体的mRNA基因; III类:大多数真核生物核mRNA的基因; tRNA内含子: tRNA基因。
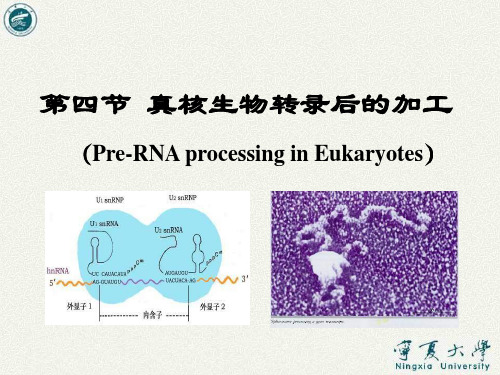
第四节 真核生物转录后的加工
(Pre-RNA processing in Eukaryotes)
多数转录的初始产物无生物活性,在生物体内 进行加工处理后才具有生物活性。
转录后加工( post-transcriptional processing):
是指将各种前体RNA(Pre-RNA)分子加工 转变成有功能的、成熟的各种 RNA (mRNA , rRNA或tRNA等) 的过程。
生物化学转录后加工

真核细胞tRNA加工过程
真核生物RNA的转录后加工
❖真核生物rRNA前体的加工 ❖真核生物tRNA前体的转录后加工 ❖真核生物mRNA前体的加工
Figure 6-21 Molecular Biology of the Cell (© Garland Science 2008)
原核生物rRNA前体的加工过程
初转录产物内部碱基互补配对, 折叠成一些茎环结构
continue
茎环结构有助于一些蛋白质结合形成核糖核 蛋白复合体(RNP, ribonucleoprotein)
(甲基化)
(M16)
(RNaseIII和RNaseE酶的剪切)
(M23) (M5)
原核生物RNA的转录后加工
分或全部poly(A),此时mRNA的寿命亦接近终点。
加尾反应所需的特定序列元件
特定序列元件:1)5‘-AAUAAA-3’’ 聚腺苷酸信号序列; 2)在其后11~20nt处紧随着一个“ 5‘-YA-3’ ”结构; 3)在其下游方向的GUGUGUG 。这些序列共同决定了聚 腺苷酸化位点(polyadenylation site)。
真核生物mRNA poly (A)长度并非固定不变 细胞核中的poly (A)长度平均为210±20 nt,细胞质poly (A )长度
190±20 nt。 输送到细胞质中的mRNA其poly (A)可由RNase切短,但又能经细胞质
poly(A)酶重新加长,保持有限的长度。 细胞质中mRNA的poly (A)长度总的趋势是逐渐变短,直到丢失大部
真核生物rRNA的特点
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、 28S rRNA和5S rRNA。其中前三者的基因组成一个转录单位, 由RNA pol Ⅰ合成,产生47S的前体,并很快转变成45S前 体;真核生物的5S rRNA由RNA pol Ⅲ合成121nt的转录产 物,几乎不需要加工。
5-转录、转录后加工
凝胶阻滞迁移实验
凝胶迁移或电泳迁移率实验(Electrophoretic Mobility Shift Assay,EMSA)是一种研究DNA结合蛋 白和其相关的DNA结合序列相互作用的技术,可用于 定性和定量分析。这一技术最初用于研究DNA结合蛋 白,目前已用于研究RNA结合蛋白和特定的RNA序列 的相互作用。 通常将纯化的蛋白和细胞粗提液和32P同位素标记 的DNA或RNA探针一同保温,在非变性的聚丙烯凝 胶电泳上,分离复合物和非结合的探针。DNA-复合 物或RNA-复合物比非结合的探针移动得慢。
~22bp
TBP
CTF/NF1
GC box
Octamer Octamer
GGGCGG
ATTTGCAT ATTTGCAT
~20bp
~20bp ~23bp
SP1
Oct-1 Oct-2
kB ATF
GGGACTTTCC GTGACGT
~10bp ~20bp
NF kB ATF
RNA聚合酶II的启动子的一致序列
控制着转录起始的频率 转录因子结合位点 增强子 决定了转录的起始点
帽子位点
GC框 CAAT框
GG(C/T)CAATCT
TATA框
TATAAAA
GGGCGG
-75
-25
A +1
(三)RNA聚合酶III的启动子 启动子位于转录区内。 在tRNA基因中启动子分为两部分:A区在 +8和+30之间,B区在+51和+72之间。两 段必须同时存在,否则不能起始。
Footprinting
硫酸二甲酯法
1. DNA与RNA聚合酶结合; 2. 在体系中加入硫酸二甲酯; 3. 碱基被甲基化,但与RNA聚合酶紧密结合的碱 基将不会被甲基化。 该方法可以发现与RNA聚合酶紧密结合的碱基。
分子生物学-转录后加工

小鼠免疫球蛋白μ重链基因可变剪接
14-14
可变剪接的多种模式
• 转录物可以按不同的模式进行可变剪接,产生具有多样性的转录本, 许多基因有2种以上的剪接方式,有的甚至达上千种。
• 几种常见的选择性剪接模式: • 1)不同启动子;2)忽略外显子;3)5’选择性剪接;4)3’选择性 剪接;5)保留内含子;6)多腺苷酸化
• 如果装配在snRNP的剪接因 子识别外显子,则称为外显子 界定(exon definition)
• 如果装配在snRNP的剪接因 子识别内含子,则称为内含子 界定(intron definition)
• 通过外显子-内含子边界(剪 接位点)突变,可以分析外显 子-内含子的界定类型
14-12
RNA Pol II的CTD参与外显子界定
内含子类型
特征
剪接体内含子(spliceosomal introns)
细胞核,mRNA,由剪接体催化切除
tRNA内含子(tRNA introns)
细胞核或古菌tRNA基因,由蛋白催化切除
自切割I类内含子(self-splicing group I introns) 细胞器,由RNA催化切除
自切割II类内含子(self-splicing group II introns)细胞器,由RNA催化切除
• 如果在6个位点发生2个不同事件,将会产生26 = 64种结果
14-15
可变剪接示例:果蝇性别决定
• 果蝇的性别决定涉及sxl(sex lethal,性别致死)、 tra (transformer, 转换)和dsx(doublesex, 双性别)三个基因前体mRNA的可变剪接
• 这3个基因的剪接存在级联反应:sxl基因雌性特异性剪接可产生活性 蛋白,进一步增强sxl基因的雌性特异性剪接,同时引发tra基因的雌性 特异性剪接,再进一步引发dsx的雌性特异性剪接
《现代分子生物学》第五章 3 真核生物的转录后加工

各种参与剪接的成分形成一个剪接体系, 称为剪接体(spliceosome)。该体系由 几种snRNP和大量的其他的蛋白质分子 组成,这些蛋白质分子称为剪接因子, 估计有40多种。 剪接点和分支点序列由剪接体识别, snRNA和蛋白质都参与了识别,特别是 snRNA之间以及与mRNA间的碱基配对 起重要作用。
RNA印迹法(RNA blotting)可用于分析核 RNA剪接过程中的中间体,从而确定内含 子的去除顺序。 实验中可以发现含有不同内含子的中间产 物,因此内含子的去除似乎没有特定的顺 序,但也发现有一定的规律,说明剪接有 特定的途径。
剪接反应并不是按内含子在RNA前体上 的顺序进行的。 RNA的构象影响剪接点的选择。在去除 特定的内含子后,RNA的构象发生一定 的变化,再选择新的剪接点。
剪接体Splicesome: snRNA:U1, U2, U4, U5, U6 snRNP:U+蛋白 质
剪接体按一定的顺序组装,已分离到一些 组装的中间体。只有组装完整的剪接体才 有功能。在剪接反应中,剪接体还会释放 和添加某些成分。 转酯反应只是磷酸酯键的直接转移,没有 水解反应的出现,因此不需要外部的能量, 也不需要供能物质如ATP或GTP的参与。 剪接体中起催化转酯反应的成分尚未弄清 楚,也不知道是蛋白质还是RNA在起作用。
一、核基因mRNA的剪接 一、核基因mRNA的剪接
mRNA的基因在低等真核生物中只有少数含 有内含子,为不连续基因。随着进化程度的 增高,不连续基因的数目不断增加。 细胞核中有一类RNA,十分不稳定,平均长 度比mRNA要长,序列的复杂程度非常高, 称为核内不均一RNA( heterogeneous nuclear RNA,hnRNA )。
RNA processing 1
第三十七章 转录后加工

锥体虫COXIII mRNA前体在编 辑过程中插入大量的尿苷酸
gRNA介导的编辑过程
小肠上皮细胞内发生的Apo B蛋白的Pre-mRNA的编辑
编辑的意义
利什曼原虫细胞色素b mRNA通过编辑创造终止密码子
rRNA前体的后加工
*
*
细菌-剪切、修剪和修饰 真核生物 1)剪切、修剪和修饰 (需要snoRNA) 2)剪接 (某些真核生物,如四膜虫)
细菌rRNA前体的后加工
真核细胞核rRNA前体的后加工
由snoRNA 指导的rRNA前体的修饰
snoRNA指导的2′-O甲基化核糖和假尿苷形成
四膜虫rRNA前体的自我剪接
tRNA前体的后加工
*
*
细菌 1) 剪切和修剪 2) 修饰 3) 添加CCA (如何需要) 真核生物 1) 剪切和修剪 2) 修饰 3) 添加CCA 4) 剪接(某些真核生物)
加尾反应由两步组成:
1. 剪切——在3′-UTR一个特定序列上游10-30 核 苷酸序列的位置切开 2. 添加腺苷酸(100-200个)产生多聚A尾巴
加尾信号
真核细胞核mRNA前体的加尾反应
为什么必须有尾巴?
保护mRNA免受3′-外切核酸酶的消化,提高 mRNA的稳定性。 PABP能够与帽子相互作用增强mRNA的可翻 译性,提高其翻译的效率; 影响最后一个内含子的剪接; 某些本来缺乏终止密码子的mRNA通过加尾 反应创造终止密码子。在UG后加尾可产生 UGA,在UA后加尾产生UAA; 通过可变加尾调节基因的表达。
第三十七章 转录后加工
提纲
一.
细菌的转录后加工
mRNA前体的后加工 rRNA前体的后加工 tRNA前体的后加工
第三十七章 转录后加工
2. CFI和CFII = “剪切因子”
3. PAP = “poly(A) 聚合酶”
加尾反应由两步组成:
1. 剪切——在3′-UTR一个特定序列上游10-30 核 苷酸序列的位置切开
2. 添加腺苷酸(100-200个)产生多聚A尾巴
加尾信号
真核细胞核mRNA前体的加尾反应
牛的PAP与Mg2+-ATP形成的复合物的三维结构
剪接信号
剪接通过2次转酯反应
在转酯反应中,1个磷酸二酯键被转移到另1个羟基上 没有水解,无能量的损失
参与剪接反应的5种snRNA
snRNA U1 U2 U4
U5 U6
互补性 内含子的5' -端
分支点
U6 snRNA
上游外显子 和下游外显子
U4 (和U2)
功能
识别和结合5'-剪接点
识别和结合分支点。在剪接体组装中,也与 U6 snRNA 配对。
酵母Pre-tRNA的剪接
古菌的转录后加工
古菌的转录后加工在某些方面分别与细菌和真核生物一样,在 某些方面仅类似于细菌或真核生物,还有少数是古菌特有的。
tRNA后加工包括:5′-端和3′-端的剪切和修剪 ;核苷酸的修饰 ; 添加CCA ;含有内含子的还包括剪接,但古菌的tRNA剪接与 真核生物一样,由一系列专门的蛋白质组成的酶按照一定的次 序催化完成的
真核细胞剪接体的装配示意图
次要剪接途径
* GU-AG规则适合绝大多数断裂基因,以此为剪接信号 的剪接途径被称为主要剪接途径。但某些断裂基因的 少数内含子的剪接信号并不遵守GU-AG规则,而是以 AT开头,AC结尾。除此以外,这一类内含子在5'-剪 接点和分支点上具有高度保守的序列,分别是 ATATCCTY和TCCTTRAY。含有与这两段保守序列互 补序列的U11和U12-snRNAs参与AT-AC内含子的剪接, 另外两种snRNAs即U4atac和U6atac分别代替U4和U6 参与这种剪接途径,只有U5被证明同时参与主要剪接 途径和次要剪接途径。
《现代分子生物学》第五章 3 真核生物的转录后加工
真核细胞中rRNA的加工途径 真核细胞中rRNA的加工途径
(1) 切除5′端的前导序列; (2) 从41S的中间产物中先切下18S的片段。 (3) 部分退火,形成发夹结构; (4) 最后修正。
真核细胞中rRNA的加工 真核细胞中rRNA的加工
目前还不清楚45S前体在剪切位点断裂后是否 就产生成熟的末端,还是要经进一步的加工。 整个加工过程需要蛋白质的参与,可能形成核 蛋白体的形式。 rRNA的加工过程还需要snoRNA (small nucleolar RNAs )的参与。 真核生物的5S rRNA是和tRNA转录在一起的, 经加工处理后成为成熟的5S rRNA。
真核细胞核mRNA的加帽反应 真核细胞核 的加帽反应
不同真核生物的mRNA可有不同的帽子结构, 同一种真核生物的mRNA也常有不同的帽子 结构。 帽子结构的作用: 1.为核糖体识别RNA提供信号 2.增加mRNA的稳定性 3.为mRNA向胞质的运输提供信号 4 与某些RNA病毒的正链RNA的合成有关。
各种参与剪接的成分形成一个剪接体系, 称为剪接体(spliceosome)。该体系由 几种snRNP和大量的其他的蛋白质分子 组成,这些蛋白质分子称为剪接因子, 估计有40多种。 剪接点和分支点序列由剪接体识别, snRNA和蛋白质都参与了识别,特别是 snRNA之间以及与mRNA间的碱基配对 起重要作用。
1. rRNA的转录后加工 rRNA的转录后加工
真核生物有4种rRNA,即5.8S rRNA、18S rRNA、28S rRNA和5S rRNA。其中前三者 的基因组成一个转录单位,产生47S的前 体,并很快转变成45S前体。 45S前体上有许多甲基化的位点,在转录 过程中或转录后被甲基化。甲基基团主要 是加在核糖上。甲基化是45S前体最终成 为成熟rRNA区域的标志。
分子生物学:转录后加工

tRNA的切割
酵母的内切酶是一种异源四 聚体蛋白质,
Sen34亚基和Sen2亚基分别 切割3`和5`剪接位点。
Sen54亚基通过“测量”成 熟tRNA结构中某个点的距离 来确定切割位点的位置。
尽管Sen15亚基的功能未知, 但其在酵母中是必需的。
反密码子环的第一个碱基和 3`剪接位点前的碱基之间的 AI碱基配对对3`剪接位点的 切割是必需的。
核糖2`位点甲基化作用需要C/D型snoRNA的 参与。
rRNA中假尿苷(pseudouridine,ψ)的合成需要 H/ACA型snoRNA。
[1] 马伟超, 李一婧, 王会宁. 脑特异性表达的snoRNA研究进展 [J]. 天水师范学院学报, 2010, 30 (5):29-32.
在脊椎动物rRNA中的保守位置上有 100种以上的2`-O-甲基基团。
的mRNA前体的直接证据; ③ 两者5`端都有帽子结构,其他的RNA分子和前体都没
tRNA的连接
真核细胞中rRNA的加工途径
真核生物的18S,5.8S和28S rRNA基因是串联在一起 形成一个转录本,初始转录本为45S前体(高等真核 生物)或更小(低等真核生物如酵母中为35S)
5S RNA是由RNA聚合酶III从单独的基因转录的,这和 原核的rRNA基因不同。
真核生物转录后加工方法

真核生物转录后加工方法1 真核生物转录后加工方法真核生物转录后加工(Transcription Post-Processing)是指在真核生物转录前实施多种变异来改变或增强其转录本表达。
它可以改变转录本结构或增强蛋白质表达效率。
真核生物转录后加工方法是可以精确协调转录因子与转录本的相互作用,从而达到增强特定基因的表达的目的。
转录后加工是研究基因表达和调控过程的主要工具,也用于开发转基因技术来改变宿主品系,或分离提高表达特定编码蛋白质功能的基因。
1.1 真核生物转录后加工技术真核生物转录后加工技术包括各种类型的转录本变异,如遗传变异(突变、转录因子缺失、RNA聚合酶选择性识别)、外源RNA干扰、DNA整合、去氧核糖核酸(RNAi)、CRISPR/Cas流程等。
1.1.1 遗传变异遗传变异是真核生物转录后加工的一种常见的技术,它涉及到各种形式的变异突变,如突变、转录因子缺失、RNA聚合酶选择性识别等。
这些技术可用于改变转录本结构,如增加剪接位点来增强表达量,减少反式转录等。
1.1.2 外源RNA干扰外源RNA干扰技术用特定的外源RNA来干扰细胞内特定的mRNA,从而减少特定基因的表达。
它也可以被用来调节特定基因的表达,或作为一种突变诊断的技术。
1.1.3 DNA整合DNA整合是让因子和特定的DNA序列整合到宿主RNA聚合酶II靶位点以调控特定基因的表达。
目前,常见的DNA整合方式包括荧光融合和杂交方法。
1.1.4 RNAiRNAi是通过使用特定的21~23碱基接头的双链RNA裂解靶标mRNA来减少特定基因的表达,从而抑制特定基因的功能。
1.1.5 CRISPR/Cas技术CRISPR/Cas技术是通过使用特定的RNA-目标-核酸(sgRNA)和多聚蛋白或酶来精确结合特定的DNA片段以调节特定基因的表达。
1.2 真核生物转录后加工作用真核生物转录后加工技术广泛应用于各种科学研究领域,其作用主要有:(1)用于研究转录调控机制(2)用于纯化特定编码蛋白质的基因(3)用于改变宿主基因组,进而改变宿主品系(4)用于生产新型转基因植物,提取编码蛋白的特定基因(5)用于开发医学应用1.3 真核生物转录后加工技术的局限性尽管真核生物转录后加工技术在研究转录调控和改变宿主品系、开发转基因技术方面发挥了重要作用,但它也具有一定的局限性。
chapter 7转录后加工

A poly(A) tail is added to the 3’ end of the transcript in 2 steps… 1) cleavage: the RNA is cut 10-30 nucleotides downstream of a specific sequence in the 3’UTR 2) addition of A’s (100-200 are added) to generate a poly(A) tail
poly(A)的功能
增加mRNA的稳定性 将携带或缺少poly(A)的球蛋白mRNA 注入到蛙卵中,结果发现,在6小时后缺少poly(A)的球蛋白 mRNA不再进行翻译,而携带poly(A)的处理仍然正常合成球 蛋白。最简单的解释是,poly(A)有助于增加mRNA的稳定性 提高mRNA翻译效率 mRNA的翻译需要PAB I(poly(A) binding protein I,poly(A)结合蛋白I)的蛋白参与,它们与 poly(A)的结合可提高mRNA的翻译效率。如果在mRNA样品 中加入过量的poly(A),可急剧降低蛋白质合成的数量。原因 在于大量poly(A)与PAB I因子竟争性结合,使mRNA缺少必 需的PAB I,因而不能有效翻译。此外适当延长poly(A)核苷 酸数目可控制mRNA的翻译。活细胞中很少mRNA的poly(A) 长度少于30 nt,低于这一长度mRNA不能翻译。 poly(A)可影响mRNA前体最后一个内含子的剪切,缺少poly (A)使剪接效率降低5-10倍。
注意稀有碱基
tRNA转录后的加工与修饰
(1)RNaseP (由蛋白质和RNA组成的复 合体)可切除E.coli前体tRNA5’的前导序列 (41nt)。该酶不识别特殊的序列,而是 识别二级结构——发夹所组成的tRNA。
分子生物学:转录及转录后加工

α rpo A 40 2 装配亚基:与启动子上游元件和
活化因子结合.
β rpo B 155 1 催化中心:结合核苷酸底物,催
β' rpo C 160 1 化磷酸二酯键形成,与模板DNA
结合。
σ rpo D 32~92 1 识别亚基:可识别启动子,促进
转录的起始。
ω
9
1 促进RNA酶组装和稳定的作用
4.2 真核生物的RNA聚合酶
真核细胞核RNA聚合酶共同的性质
* 都是大的多亚基蛋白质(500-700 k) * 都有两个大的与大肠杆菌RNA聚合酶 、和′亚
基同源的亚基,因此活性中心是保守的 * 没有与大肠杆菌σ因子同源的亚基
真核生物的RNA聚合酶结构与功能的比较
名称
RNA聚合酶I (A) RNA聚合酶II (B) RNA聚合酶 Ⅲ(C)
⑹ 都遵从碱基配对规律—— 但转录忠实性要低于DNA复制;
⑺ 转录与复制都受到严格的调控。
3.转录和复制的区别
复制
模板
两条链均复制
原料 酶
dNTP DNA聚合酶
产物
子代双链DNA
配对 高度进行性
A-T;G-C 中途不停止
转录 不对称转录
NTP RNA聚合酶 mRNA,tRNA,rRNA A-U;T-A;G-C 可一段一段复制
第七章 转录及转录后加工
主要内容:
第一节 转录的复制机理与体系 第二节 与转录起始和终止有关的DNA结构 第三节 原核生物和真核生物转录
第四节 转录后加工过程及其机制
1955年,Brachet分别用洋葱根尖和变形虫 进行实验。如果往洋葱根尖细胞和变形虫中加入 RNA酶分解细胞质中的RNA,细胞中的蛋白质合成 就会停止。而再加进从酵母菌中提取的RNA,则 又能够合成一定数量的蛋白质。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
酵母Pre-tRNA的剪接
古菌的转录后加工
古菌的转录后加工在某些方面分别与细菌和真核生物一样,在 某些方面仅类似于细菌或真核生物,还有少数是古菌特有的。 tRNA后加工包括:5′-端和3′-端的剪切和修剪 ;核苷酸的修饰 ; 添加CCA ;含有内含子的还包括剪接,但古菌的tRNA剪接与 真核生物一样,由一系列专门的蛋白质组成的酶按照一定的次 序催化完成的 rRNA后加工:其基因以多顺反子的形式存在,通过剪切和修剪 释放出三种rRNA。核苷酸的修饰与真核生物一样,需要各种特 殊的小RNA(相当于真核生物的snoRNA)与蛋白质形成的 RNP去锁定修饰目标。但对于含有内含子的rRNA的剪接与其 tRNA剪接机制相似,完全不同于真核生物 mRNA后加工:后加工类似于细菌,没有帽子结构,一般也没 有多聚A尾巴。对于少数具有多聚A尾巴的mRNA而言,其加尾 的机制和功能完全不同于真核生物,倒与含有多聚A尾巴的细 菌相似。含有内含子的mRNA的剪接机制既不同于真核生物, 也不同于细菌,而是类似于它的tRNA的剪接
SnRNAs
*一般由60nt~300nt组成,且富含U * 对于剪接十分重要
*ቤተ መጻሕፍቲ ባይዱ调节序列的特异性和剪接反应的精确性
* 折叠成特定的二级结构,这对催化十分重要
*snRNP中起催化作用的是RNA,不是蛋白质
*参与剪接的只有U1、U2、U4、U5和U6
剪接体的组装与剪接反应
*
*
*
*
*
分支点结合蛋白(BBP)识别并结合分支点,剪接因 子U2AF与富含嘧啶的序列结合; U2-snRNP取代BBP,并与分支点的一致序列配对。 分支点内的一个A无互补配对的U,突出在双螺旋外而 被激活,为剪接的第一次转酯反应提供了便利; U1-snRNP 与5'-剪接点的一致序列互补配对; U4/U6 -U5-snRNP三聚体进入剪接体。
剪接体的组装和去组装
分支点A2' –羟基的突出以及5' –剪接点和3'–剪接点的结构
真核细胞剪接体的装配示意图
次要剪接途径
* GU-AG规则适合绝大多数断裂基因,以此为剪接信号 的剪接途径被称为主要剪接途径。但某些断裂基因的 少数内含子的剪接信号并不遵守GU-AG规则,而是以 AT开头,AC结尾。除此以外,这一类内含子在5'-剪 接点和分支点上具有高度保守的序列,分别是 ATATCCTY和TCCTTRAY。含有与这两段保守序列 互补序列的U11和U12-snRNAs参与AT-AC内含子的剪 接,另外两种snRNAs即U4atac和U6atac分别代替U4和 U6参与这种剪接途径,只有U5被证明同时参与主要 剪接途径和次要剪接途径。
为什么只有mRNA才会加尾?
* 与加帽反应一样,只有mRNA才会加尾, 也是因为聚合酶II最大亚基上的CTD重复 序列被TFIIH磷酸化,但是磷酸化位点为 Ser2 。 Ser2 的 磷 酸 化 将 加 尾 因 子 招 募 到 mRNA 前体上进行加尾反应。
mRNA前体的剪接
剪接这种后加工方式是在发现基因断裂的现象后确定 的。1977年,由Phillip Sharp和Richard Roberts领导两个 实验小组几乎同时在腺病毒的晚期表达基因中发现蛋 白质基因断裂现象。 进一步研究表明,基因断裂是真核细胞及其病毒的基 因组中的普遍现象,在高等生物的基因组中,只有很 少的蛋白质基因是连续的(如组蛋白和干扰素),但 在低等的真核生物,断裂基因却不多见。 不同断裂基因含有的内含子数目不一定相同,同样内 含子大小也会有差别。一般说来,一个典型的真核生 物蛋白质的基因由10% 的外显子序列和90%的内含子 序列组成。
锥体虫COXIII mRNA前体在编 辑过程中插入大量的尿苷酸
gRNA介导的编辑过程
小肠上皮细胞内发生的Apo B蛋白的Pre-mRNA的编辑
编辑的意义
利什曼原虫细胞色素b mRNA通过编辑创造终止密码子
rRNA前体的后加工
* *
细菌-剪切、修剪和修饰 真核生物 1)剪切、修剪和修饰 (需要snoRNA) 2)剪接 (某些真核生物,如四膜虫)
细菌rRNA前体的后加工
真核细胞核rRNA前体的后加工
由snoRNA 指导的rRNA前体的修饰
snoRNA指导的2′-O甲基化核糖和假尿苷形成
四膜虫rRNA前体的自我剪接
tRNA前体的后加工
*
*
细菌 1) 剪切和修剪 2) 修饰 3) 添加CCA (如何需要) 真核生物 1) 剪切和修剪 2) 修饰 3) 添加CCA 4) 剪接(某些真核生物)
为什么只有mRNA被snRNPs剪接?
* * *
RNA聚合酶的CTD是剪接必需的 剪接体内的剪接因子与CTD作用 这保证来自RNA聚合酶II的转录物处于 和剪接机构正确的相互作用的位置。
可变剪接和反式剪接
可变剪接
1. 2. 剪接是精确的,但是... 一种mRNA前体可以通过去除不同的内含子/ 外显子组合而剪接成两种以上的mRNA,从 而导致一个基因可以编码多种蛋白质 可变剪接受到调节 果蝇的性别决定与可变剪接有关 发生在两个mRNA分子之间的剪接 比较罕见 自然界只发现在锥体虫和秀丽线虫
骑行纳古菌tRNAGlu前体的反式剪接
大肠杆菌 tRNATyr的后加工
核糖核酸酶 P –一种真正的核酶
一种核酸内切酶–参与tRNA前体5′-端的后加 工 大肠杆菌核糖核酸酶 P: 14-k的多肽链 + 一 种 377核苷酸长的RNA ( M1 RNA) 在高浓度的Mg2+下,分离的M1 RNA能单独 和剪切大肠杆菌的tRNA前体。核糖核酸酶P 中的多肽链提高 M1 RNA的剪切速率,允 许它在生理Mg2+浓度下起作用。
RNA polII CTD的磷酸化与加尾和加帽反应的关系
3′-加尾
加尾内因:加尾信号 ATG 加尾外因:
终止密码子
mRNA
1. CPSF = ―剪切/多聚腺苷酸化特异性因子(3个 亚基)识别mRNA前体 3′-UTR上的 AAUAAA 一致序列,并与此结合 2. CFI和CFII = ―剪切因子” 3. PAP = ―poly(A) 聚合酶”
3. 4.
1. 2. 3.
反式剪接
果蝇的性别决定与可变剪接
反式剪接
mRNA前体的编辑
* 定义 任何发生在转录后编码区内的序列变化 * 主要类型 1) 尿苷酸的插入 2) C→U (哺乳动物的Apo B-48) 3) A→I (哺乳动物谷氨酸受体的一个亚基) * 机制 1) gRNA 2) 特殊的脱氨酶 * 意义 1) 提高遗传信息的容量 2) 创造终止密码子和起始密码子
加帽和甲基化
* * * * 帽子结构 加帽反应:共转录 为什么只有mRNA才会加帽? 功能
为什么要有帽子?
提高mRNA的稳定性 参与识别起始密码子的过程,提高mRNA 的可翻译性 有助于mRNA通过核孔从细胞核运输到细 胞质 提高剪接反应的效率。
为什么只有mRNA加帽?
* 之所以只有mRNA和某些snRNA才有帽子 结构,是因为它们都由聚合酶II催化,当 TFIIH磷酸化CTD重复序列中的Ser5以后, 它即可以将转录因子DSIF 招募到转录复合 物。DSIF随后将另一种转录因子NELF招 募进来,以阻滞转录。上述暂停允许加帽 酶进入,来修饰转录物的5'-端。第三种转 录因子P-TEFb是一种激酶,在帽子结构形 成不久也被招募到复合物,然后磷酸化 CTD的Ser2和NELF,NELF随之失活,聚 合酶II继续延伸。
R-环技术:真核基因内含子数目与结构分析
变性
与成熟的mRNA杂交; 在电镜下观察
DNA 模板链 成熟的mRNA
R环实验的结果及其对结果的解释
鸡卵清蛋白基因结构及其Pre-mRNA的后加工
mRNA前体的剪接机制
*
*
* * *
mRNA前体的剪接是高度精确的。其精确 性一方面取决于位于外显子和内含子交界 处的剪接信号(可以将其视为内因),另 外一方面取决于5种被称为snRNP的核糖核 酸蛋白质复合物(可以将其视为外因)。 剪接反应的“内因”——剪接信号 剪接反应的―外因‖——snRNP和剪接因子 剪接反应——两次转酯反应 剪接体的组装
细菌mRNA前体的后加工
在细菌,mRNA很少有后加工。但某些 噬菌体mRNA会发生最简单的剪切反应, 将一个多顺反子切割成单顺反子,也有 某些噬菌体的mRNA需要经过相对复杂 的剪接反应才能成熟(如T4噬菌体编码 的胸苷酸合酶)。
真核细胞mRNA前体的后加工
* 加工形式 1) 5′-端 = 加帽 2) 3′-端 = 加尾 3) 内部 = 剪接 4) 内部=甲基化 5) 编码区=编辑 * 后加工机制
第三十七章 转录后加工
提纲
一.
细菌的转录后加工
mRNA前体的后加工 rRNA前体的后加工 tRNA前体的后加工
1. 2. 3.
二.
真核生物的转录后加工
mRNA前体的后加工 rRNA前体的后加工 tRNA前体的后加工
1. 2. 3.
三.
古菌的转录后加工
转录后加工
基因转录的直接产物被称为初级转录物。初 级转录物一般是无功能的,它们在细胞内必 → 须经历一些结构和化学的变化即所谓的转录 后加工以后才会有功能。转录后加工可能是 RNA的功能所必需的,也可能提供基因表达 调控的一种手段。 RNA所能经历的后加工方式可达10种以上, 但后加工反应的本质要么是增减一些核苷酸 序列,要么是修饰某些特定的核苷酸。
剪接信号
剪接通过2次转酯反应
在转酯反应中,1个磷酸二酯键被转移到另1个羟基上 没有水解,无能量的损失
参与剪接反应的5种snRNA
snRNA U1 U2 U4 互补性 内含子的5' -端 分支点 U6 snRNA 功能 识别和结合5'-剪接点 识别和结合分支点。在剪接体组装中,也与 U6 snRNA 配对。 结合并失活U6。在剪接体组装中,U4-U6之间 的碱基配对被U2-U6之间的剪接配对所取代。 U6也取代U1 与5' -剪接点的相互作用。 与相邻的2个外显子结合,以防止它们离开剪 接体。 在剪接体中与U2结合。