不同温度下太平洋牡蛎三倍体和二倍体生物能量学比较
大规模诱导生产太平洋牡蛎(Crassostrea gigas(Thunberg))3倍体的方法比较

大规模诱导生产太平洋牡蛎(Crassostrea
gigas(Thunberg))3倍体的方法比较
李赟;于瑞海;王昭萍;王如才;郑小东
【期刊名称】《高技术通讯》
【年(卷),期】2000(010)011
【摘要】利用6-DMAP和冷休克处理太平洋牡蛎受精卵大量生产牡蛎3倍体苗种用于养殖生产,结果表明,与冷休克相比,6-DMAP处理对胚胎发育存在较大的影响,幼虫孵化率有所下降,但3倍体率则基本稳定在60%以上 ,比冷休克高30%,综合分析认为,6-DMAP可作为比较理想的诱导药剂用于大量培育3倍体苗种的养殖.【总页数】3页(P1-3)
【作者】李赟;于瑞海;王昭萍;王如才;郑小东
【作者单位】青岛海洋大学水产学院,青岛,266003;青岛海洋大学水产学院,青岛,266003;青岛海洋大学水产学院,青岛,266003;青岛海洋大学水产学院,青
岛,266003;青岛海洋大学水产学院,青岛,266003
【正文语种】中文
【中图分类】S9
【相关文献】
1.低渗诱导太平洋牡蛎三倍体以及与其他诱导方法的比较 [J], 孔静;王昭萍;于瑞海;刘剑;张跃环
2.人工诱导长牡蛎Crassostrea gigas(Thunberg)三倍体发生的初探 [J], 林位琅
3.四倍体太平洋牡蛎(Crassostrea gigas Thunberg)自群繁殖的研究 [J], 施坤涛;王昭萍;于瑞海
4.两种方法诱导太平洋牡蛎三倍体在生产上的应用效果 [J], 于瑞海;王昭萍;王如才;田传远;郑小东
5.二倍体和三倍体太平洋牡蛎(Crassostrea gigas)繁殖前后生化成分及氨基酸组成的比较研究 [J], 孔令锋;王昭萍;于瑞海;王如才
因版权原因,仅展示原文概要,查看原文内容请购买。
二倍体与三倍体太平洋牡蛎(Crassostrea gigas)的细胞学比较研究

多倍 体 的快 速 生 长 现 象 几 乎 在 所 有 研 究 的 贝类 中都 得 以证 实 。如 侏 儒 蛤 Muii aeai l altrl n s
Sa 1 y[]
、
太 平 洋 牡 蛎 C a ssragg s 、 浦 珠 母 r sot i a E 合 e ]
、
Pica ama tn i( )5、 洲 牡 蛎C a— n td re s D. [ 美 i ] rs
假 说 来 解 释 多倍 体 贝 类 快 速 生 长 现 象 , 此 从 细 胞 学 的 角 度 研 究 多倍 体 快 速 生 长 的 机 理 是 很 因
必要的 。
本 研 究 从 细 胞 学 的 角 度 比较 了 二倍 体 和 三 倍 体 太 平 洋 牡 蛎 的 差 异 , 探 讨 多倍 体 的 快 速 为
维普资讯
第3 2卷 第 4期
青 岛 海 洋 大 学 学 报
J OU RNA L OF OCEAN N I ERSI U V TY OF I GDAO Q N
3 4 55 2( ): 1~ 5 6 5
20 0 2年 7月
Jl u y,2 02 0
唇 瓣 结缔组 织 中血 淋 巴核 径 、 肠 上皮 细 胞高 度 的大 小 。结 果显 示 , 倍 体鳃 上 皮 细胞 核 长轴 、 直 三 外 套 膜 与 唇 瓣 结 缔 组 织 中 血 淋 巴 核 径 显 著 大 于 二 倍 体 ( < O 0 ) 而 二 倍 体 和 三 倍 体 直 肠 上 皮 细 胞 P .1, 高 度 没 有 明 显 差 别 。本 研 究 结 果 支 持 了 多 倍 体 的 巨 态 性 假 说 , 揭 示 了 倍 性 对 细 胞 核 大 小 的 影 响 。 并 关键 词 太 平 洋 牡 蛎 ; 倍 体 ; 倍 体 ; 胞 学 二 三 细
葡萄牙牡蛎二倍体与太平洋牡蛎三倍体杂交生产异源四倍体的方法[
(19)中华人民共和国国家知识产权局(12)发明专利申请(10)申请公布号 (43)申请公布日 (21)申请号 202010673186.2(22)申请日 2020.07.14(71)申请人 厦门世倍海洋科技有限公司地址 361000 福建省厦门市思明区海岸后街54号205室(72)发明人 朱李贤 (74)专利代理机构 北京市炜衡律师事务所11375代理人 许育辉(51)Int.Cl.A01K 61/54(2017.01)A01K 61/17(2017.01)(54)发明名称葡萄牙牡蛎二倍体与太平洋牡蛎三倍体杂交生产异源四倍体的方法(57)摘要本发明公开了一种葡萄牙牡蛎二倍体与太平洋牡蛎三倍体杂交生产牡蛎异源四倍体的方法。
本发明以太平洋牡蛎三倍体作为母本,以全重为目标优选葡萄牙牡蛎二倍体作为父本,通过药物诱导杂交生产出牡蛎异源四倍体。
本方法获得的牡蛎异源四倍体具有广温的特性,其与普通葡萄牙牡蛎或太平洋牡蛎二倍体杂交生产的牡蛎三倍体适合我国南方和北方沿海养殖,推广应用前景广阔。
权利要求书1页 说明书4页 附图1页CN 112535128 A 2021.03.23C N 112535128A1.葡萄牙牡蛎二倍体与太平洋牡蛎三倍体杂交生产异源四倍体的方法,其特征包括以下几个步骤:(a)挑选太平洋牡蛎三倍体作为繁殖母本,挑选普通葡萄牙牡蛎二倍体作为繁殖父本,种贝分别放入种贝培育池进行促熟;(b)太平洋牡蛎种贝促熟至性腺成熟后,取得卵子,并于下一步开始前熟化;解剖促熟后葡萄牙牡蛎二倍体,获得精子,并于下一步开始前激活;(c)将熟化的卵子与激活的精子混合于水中,所述精子与卵子数量比例为5~10:1,控制受精卵密度保持在≤1万个/毫升;(d)观察受精卵发育情况,当第一极体出现50%~75%时,迅速加入细胞松弛素B(0.5~1.0mg/L)或6-二甲基氨基嘌呤(15~30mg/L)处理20~30min;(e)受精卵在药物处理结束后立即清洗并过滤药物,设置孵化密度,放置于黑桶中孵化;当发育至D型幼虫后,筛选幼虫,设置培育密度并放入黑桶培育;待幼虫出现眼点后,投放附着基质。
三种方法诱导熊本牡蛎三倍体的比较研究

doi: 10.7541/2021.2019.263三种方法诱导熊本牡蛎三倍体的比较研究李玲蔚 李海昆 于瑞海 马培振 张 哲 王永旺(中国海洋大学海水养殖教育部重点实验室, 青岛 266003)摘要: 为获得高效的熊本牡蛎的三倍体诱导方法, 分别比较了6-DMAP 、高盐和低盐3种诱导方法在不同的诱导浓度(盐度)、起始诱导时间和持续诱导时间下的卵裂率、孵化率和三倍体率, 同时比较了3种方法获得的幼虫的生长、存活和三倍体率变化情况。
结果表明, 在6-DMAP 诱导组中, 三倍体率和诱导效率分别可达37.97%—58.01%和34.30%—42.50%, 培育期间幼虫的平均存活率27.19%, 生长率13.03 μm/d, 三倍体率降低了24.94%; 在低盐诱导组中, 三倍体率和诱导效率分别可达7.32%—42.25%和2.17%—31.41%, 培育期间幼虫的平均存活率33.92%, 生长率12.71 μm/d, 三倍体率降低了20.64%; 在高盐诱导组中, 三倍体率和诱导效率分别可达7.47%—63.03%和6.58%—49.41%, 培育期间幼虫的平均存活率31.66%, 生长率13.08 μm/d, 三倍体率降低了17.64%。
综合来看, 高盐诱导是诱导三倍体熊本牡蛎的最优方法, 其诱导条件的最佳组合为盐度55, 起始诱导时间受精后15min, 持续诱导时间20min 。
研究为熊本牡蛎的三倍体诱导提供了技术支持, 对熊本牡蛎的多倍体育种具有重要的理论指导意义。
关键词: 熊本牡蛎; 6-DMAP; 高盐; 低盐; 三倍体率中图分类号: S966.9 文献标识码: A 文章编号: 1000-3207(2021)02-0352-08熊本牡蛎(Crassostrea sikamea )属于巨牡蛎属(Crassostrea ), 在中国、日本和韩国的沿海均有分布[1—3], 其独特的口味深受消费者的喜爱,被美国引进并成功地开展了生产养殖[4—6]。
单体三倍体太平洋牡蛎苗种生产技术

度 为6~ 10个 /毫 开 ,幼 虫 培 育 水温 为23℃。 幼虫 培 育 受 精 卵适 宜 诱导 密 度 ,经 我 们 多年经 验 认 为 200个 /
按 常 规 方法 培 育 每 两 犬测 量 幼 虫壳 长 和 密 度
毫 升 右 比较 适 宜 。再 就 是6一DMAP诱 导 的浓 度 和 处理
选 用 150 ̄300目的 废 旧筛 绢 、经 打 磨表 面 粗糙 的 塑 料
2.单体牡蛎生产方法和附着基的探讨 目前单
薄 膜和 鲍鱼 育 苗用 的白 色波纹 板 3种 , 以贝壳 采 茁器 做 体牡 蛎 生产 方 法主 要 有 两种 : 一 是 肾上 腺 素和 去甲 肾
5.采 苗 及 单体 牡 蛎 生产 在 幼 虫 出 现 眼 点 后 利 时 机及 处 理 时问 . 只要 保证 述 条件 ,牡蛎 的三 倍体
用 先 附着 后 脱基 的 方法 获 得 单体 三 倍体 牛t蛎 采 茼器 诱导 率 都 很 高 ,一 般在 80%以上 ,彘 高 达 95%。
别 雌雄 ,并 检 查精 卵 的成 熟 状况 挑 出 发育 好 的个 件
3.脱 基 幼 虫 附 茁 后 10天 左右 牛长 至 壳 长 I.0~
备 用 ,雌 雄 严格 分 开放 置 ,避 免 精 子污 染 。 将成 熟 的 1.5毫米,此时抖动筛绢和靼料薄膜可使小稚贝脱基 .
卵 巢逐 个 剥 离并 破 碎 ,先 用80目、 150目、250目的 筛 剥 离的 成活 率 为89.66%; 附在 波 纹板 上 的蛎 茁 在此 规
1.三倍体牡蛎诱导率的探讨 I 1前 贝类多倍体
水 中 孵化 、培 育 。
诱导 是近 几年研 究的热 点之 一.通 过 多年的 研究 发现 ,
不同发育期的太平洋牡蛎在不同干露状态下的成活率研究

研究简报不同发育期的太平洋牡蛎在不同干露状态下的成活率研究Ξ于瑞海,王昭萍,孔令锋,李 琪,郑小东(中国海洋大学,山东青岛,266003)摘 要: 用太平洋牡蛎的D 形幼虫、壳顶中期幼虫、眼点幼虫、稚贝、幼贝及成体为材料,在8~10℃和20~22℃干、湿环境下干露不同时间对所取样品的成活率进行了比较研究。
实验说明太平洋牡蛎其成体、D 形幼虫、壳顶中期幼虫、眼点幼虫、稚贝及幼贝在低温湿润的条件下干露时间长,成活率高,而在高温干燥的条件下干露相同的时间成活率明显降低。
在温度、湿度相同的条件下,其耐干能力为成贝>眼点幼虫>壳顶中期幼虫>幼贝>稚贝>D 形幼虫。
本实验为太平洋牡蛎幼虫的异地采苗,稚贝和幼贝的长途运输,成贝低温或干露保存提供了一定的科学依据。
关键词: 太平洋牡蛎;干露;成活率中图法分类号: S966.1 文献标识码: A 文章编号: 167225174(2006)042617204 太平洋牡蛎(Crassost rea gigas )因生长速度快,出肉率高,营养丰富,具有较高的经济价值,是我国1种重要的养殖贝类。
目前牡蛎随着国内外市场需求的大量增加,促进了我国牡蛎养殖业和加工出口业的发展,因此,加工和养殖单位每年都要购买大量的成贝和幼苗,但由于对干露时间缺乏充分了解,常出现大量死亡现象,造成较大的经济损失;另外由于幼虫培育期较长,附着基运输不便等,一些育苗单位往往需要进行异地采苗,也需要对幼虫的干露时间有所掌握[1]。
而以往对牡蛎各生长阶段不同干露时间下的成活率报道不多,且不够系统和详细,本文探讨了不同温度下、不同湿度下干露不同时间对太平洋牡蛎成贝、幼贝、稚贝及其幼虫成活率的影响规律,为科研提供理论依据,并对育苗、养殖生产和加工单位具有一定指导作用。
1 材料与方法1.1材料1.1.1太平洋牡蛎 太平洋牡蛎成贝、D 形幼虫、壳顶中期幼虫、眼点期幼虫均取自威海市初村文登水产养殖综合育苗基地。
三倍体太平洋牡蛎生产性育苗与养成初报

三倍体太平洋牡蛎生产性育苗与养成初报
王昭萍;李贇;王如才;郑小东;于瑞海;田传远;高强
【期刊名称】《海洋湖沼通报》
【年(卷),期】2000()3
【摘要】本文总结了1998~1999年三倍体太平洋牡蛎育苗与养成情况。
98年在荣成三4863中试基地利用6-DMAP、冷休克及CB抑制受精卵第二极体释放的方法诱导太平洋牡扳三倍体,其中以6-DMAP效果较好,诱导三倍体率为60.9%-90.5%。
共培育三倍体群牡蛎苗种4.13亿,海上养殖2000亩。
三倍体群牡蛎的生长明显快于二倍体对照群体,体重增加15.65%~41.89%。
1999年4~8月收获,三倍体群牡蛎比二倍体平均亩增产20%。
【总页数】6页(P34-39)
【关键词】太平洋牡砺;三倍体;育苗;养成
【作者】王昭萍;李贇;王如才;郑小东;于瑞海;田传远;高强
【作者单位】青岛海洋大学
【正文语种】中文
【中图分类】S968.3
【相关文献】
1.太平洋牡蛎三倍体育苗技术初步分析 [J], 刘景斗;刘溪运
2.太平洋牡蛎三倍体育苗技术初步研究 [J], 王在文
3.太平洋牡蛎三倍体苗种生产性培育试验 [J], 马庆涛;廖烈金;杨晓;李春晓;陈伟洲;郭森记;康叙钧
4.关于太平洋牡蛎的养殖技术之一太平洋牡蛎三倍体育苗技术措施 [J], 秦安庆;王世晔;邵淑芳;葛民兴;关茂贵
5.单体全三倍体太平洋牡蛎育苗技术的研究 [J], 于瑞海;王昭萍;施坤涛;孔令锋;牟宗宝;马化亮
因版权原因,仅展示原文概要,查看原文内容请购买。
温度和盐度对长牡蛎“海大1号”二倍体和三倍体幼虫生长与存活的影响

第54卷 第3期 2024年3月中国海洋大学学报P E R I O D I C A L O F O C E A N U N I V E R S I T Y O F C H I N A54(3):031~039M a r .,2024温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长与存活的影响❋张海宁1,徐成勋1,李 琪1,2❋❋(1.海水养殖教育部重点实验室(中国海洋大学),山东青岛266003;2.青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室,山东青岛266237)摘 要: 为确定长牡蛎(C r a s s o s t r e a g i ga s ) 海大1号 苗种规模化繁育的最适条件,本研究设置4个温度梯度(20㊁25㊁30和35ħ)和5个盐度梯度(20㊁25㊁30㊁35和40),分析了不同温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长和存活的影响㊂研究表明,当温度25ħ和盐度30时,长牡蛎 海大1号 二倍体和三倍体幼虫的受精率㊁孵化率和变态率最高㊂不同温度处理下,三倍体幼虫的生长速度和累计存活率始终高于二倍体幼虫㊂随着温度升高幼虫生长速度增加,但存活率下降,在25ħ时三倍体幼虫存活率最高(52.00%ʃ6.56%),温度为30ħ时,三倍体幼虫壳高增长最快(14.82μm ㊃d -1)㊂不同盐度下,三倍体幼虫的生长速度和累计存活率显著高于二倍体幼虫㊂幼虫的存活率随着盐度升高呈先升高再下降的趋势,三倍体幼虫累计存活率在盐度30时最高(52.00%ʃ6.56%);幼虫的生长速度随着盐度升高呈下降的趋势,三倍体幼虫壳高在盐度20时增长最快(18.22μm ㊃d -1)㊂研究结果表明,长牡蛎 海大1号 三倍体幼虫比二倍体幼虫表现出更强的温度㊁盐度的耐受性;长牡蛎 海大1号 三倍体苗种培育的适宜盐度为20~30,适宜温度为20~25ħ㊂本文可为长牡蛎 海大1号 三倍体苗种的规模化繁育提供参考㊂关键词: 长牡蛎;三倍体幼虫;温度;盐度;生长;存活;海大1号中图法分类号: S 968.3 文献标志码: A 文章编号: 1672-5174(2024)03-031-09D O I : 10.16441/j.c n k i .h d x b .20220514引用格式: 张海宁,徐成勋,李琪.温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长与存活的影响[J ].中国海洋大学学报(自然科学版),2024,54(3):31-39.Z h a n g H a i n i n g ,X u C h e n g x u n ,L i Q i .E f f e c t s o f t e m p e r a t u r e a n d s a l i n i t y o n t h e g r o w t h a n d s u r v i v a l o f d i p l o i d a n d t r i pl o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 [J ].P e r i o d i c a l o f O c e a n U n i v e r s i t y of C h i n a ,2024,54(3):31-39. ❋ 基金项目:现代农业产业技术体系项目(C A R S -49);山东省农业良种工程项目(2021L Z G C 027)资助S u p p o r t e d b y t h e A g r i c u l t u r e R e s e a r c h S y s t e m o f C h i n a (C A R S -49);t h e A g r i c u l t u r a l S e e d I m p r o v e m e n t P r o j e c t o f S h a n d o n g Pr o v i n c e (2021L Z G C 027)收稿日期:2022-12-30;修订日期:2023-02-04作者简介:张海宁(1996 ),男,硕士生,主要从事贝类遗传育种研究㊂E -m a i l :1501403997@q q.c o m ❋❋ 通信作者:李 琪(1966 ),男,博士,教授,博士生导师,主要研究方向为海洋贝类遗传育种及繁殖生物学研究㊂E -m a i l :qi l i 66@o u c .e d u .c n 长牡蛎(C r a s s o s t r e a g i ga s )作为全球性重要的经济贝类,具有生长速度快㊁抗逆性强㊁肉质鲜美㊁营养丰富等特点㊂长牡蛎 海大1号 是以生长速度和壳型为选育目标,通过群体选育培育出的新品种[1]㊂然而,长牡蛎 海大1号 与其他传统养殖的二倍体牡蛎一样,由于夏季繁殖期配子排放导致软体部消瘦,无法供应市场㊂与二倍体牡蛎相比,三倍体牡蛎育性相对较差,在繁殖季节也能保持良好的肉质,弥补夏季二倍体牡蛎市场短缺,可以保障全年供应[2]㊂因此,开展长牡蛎海大1号 三倍体苗种的规模化繁育与产业化应用对推动牡蛎养殖产业的发展具有重要意义㊂幼虫培育是牡蛎苗种生产的重要环节,了解幼虫对养殖环境的偏好对于最大限度地提高育苗场的产量至关重要,而环境条件在无脊椎动物幼虫发育中起着至关重要的作用㊂通常情况下,在探究培育幼虫适宜条件时,除食物的质量和数量,需要考虑温度和盐度的适应性[3-4]㊂温度是影响贝类幼虫生长和存活的重要环境因素,高温会导致海洋贝类幼虫大量死亡,而低温则会导致大部分海洋贝类幼虫生长缓慢[5-7]㊂盐度影响幼虫的生理反应,双壳贝类的渗透压调节功能受海水盐度的影响,盐度变化超过其自身调节能力是导致双壳贝类死亡的重要原因之一[8-9]㊂有关温度㊁盐度对贝类生长发育的影响已有大量报道,阐明温度和盐度对长牡蛎(C .g i g a s )[6]㊁岩牡蛎(C r a s s o s t r e a n i p-p o n a )[10]㊁栉孔扇贝(C h l a m y s fa r r e r i )[9]㊁菲律宾蛤仔(R u d i t a p e s p h i l i p p i n a r u m )[11]㊁钝缀锦蛤(T a pe s d o r s a t u s )[12]幼虫及稚贝生长发育的影响,可为优化贝类幼虫培育条件提供基础数据㊂研究幼虫培育的最佳中 国 海 洋 大 学 学 报2024年环境条件有助于提高幼虫的生长和存活,缩短幼虫培育周期的同时降低生产成本㊂本文研究了温度与盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长和存活的影响,旨在为长牡蛎 海大1号 三倍体苗种规模化扩繁提供理论基础和实践经验㊂1 材料与方法1.1实验材料实验在烟台莱州利滔育苗场进行,所用亲贝为1龄长牡蛎 海大1号 二倍体和四倍体,均取自山东荣成桑沟湾海区㊂挑选壳色性状优良㊁壳型规则㊁外表无损伤的个体作为亲贝,利用流式细胞仪(B e c k m a n C y t -o F L E X )检测亲贝倍性㊂1.2实验设计实验设计4个温度梯度:20㊁25㊁30和35ħ,海水盐度设置为30;设置5个盐度梯度:20㊁25㊁30㊁35和40,水温设置为25ħ㊂水温通过冷水机或加热器控制,使用水温计测定水温㊂低盐海水通过向过滤海水中加入曝气24h 的自来水配制,高盐海水通过向过滤海水中加入海水晶配制,使用盐度计(A T A G O )测量盐度㊂采用解剖法获得长牡蛎 海大1号 二倍体精卵和四倍体的精子㊂取长牡蛎 海大1号 二倍体卵子,260目筛绢过滤后,在砂滤海水中熟化1h ㊂二倍体和四倍体牡蛎的精子于授精前10m i n 挤出备用,授精时将卵液平均分为9份,并分别加入长牡蛎 海大1号 二倍体和四倍体的精子㊂每个实验组设置3个重复㊂1.3日常管理授精后放置于60L 聚乙烯桶中孵化㊂实验过程中每天换水1次,换水量为总体积的1/3,随着幼虫的生长,逐渐增加换水量至1/2㊂饵料以等鞭金藻(I s o c h -r ys i s g a l b a n )为主,后期辅助投喂亚心形扁藻(M a r i n e m i c r o a l g a )和小球藻(C h l o r e l l a v u l ga r i s ),每天投喂3次,投饵量根据幼虫的摄食和水中残饵情况确定㊂各实验组投饵㊁换水㊁充气等操作一致㊂1.4指标测定幼虫孵化后每4天取样一次,每组随机取30个幼虫测量㊂采用五点取样法取样,即从每组内的5个不同位置取样,每个位置取样20m L ,共取样100m L 幼虫,用卢戈氏液固定,在显微镜下测量其壳高㊂同时测定幼虫密度,每次测定后及时对密度进行均等校正,使各组保持一致,以消减密度效应的影响㊂根据初始密度㊁每次调整密度和测定的密度推算幼虫存活率㊂受精率为受精后4h 卵裂数占总卵数的百分比,孵化率为受精24h D 形幼虫占总受精卵数量的百分比,存活率为存活的幼虫数占初始实验幼虫总数的百分比,变态率为变态的幼虫数占附着基投放时实验幼虫总数的百分比㊂1.5数据分析数据统计分析采用S P S S 24.0软件处理,显著性水平设为P <0.05㊂2 结果2.1受精与孵化2.1.1不同温度对长牡蛎 海大1号 二倍体和三倍体受精率与孵化率的影响 25ħ实验组和30ħ实验组的长牡蛎 海大1号 二倍体与三倍体的受精率与孵化率均显著高于20ħ实验组和35ħ实验组(P <0.05),且温度为25和30ħ时二倍体组受精率显著高于三倍体组(P <0.05)(见图1)㊂温度为25ħ时,长牡蛎 海大1号 二倍体组与三倍体组的受精率最高,分别为92.67%ʃ2.52%和85.33%ʃ2.52%;温度为(同一指标具有相同字母表示组间差异不显著(P >0.05),具有不同字母表示组间存在显著差异(P <0.05)㊂S a m e i n d i c a t o r w i t h t h e s a m e l e t t e r m e a n st h a t t h e d i f f e r e n c e b e t w e e n g r o u p s i s n o t s i g n i f i c a n t (P >0.05),a n d w i t h d i f f e r e n t l e t t e r s m e a n s t h a t t h e r e i s a s i g n i f i c a n t d i f f e r e n c e b e t w e e n g r o u ps (P <0.05).)图1 不同温度下长牡蛎 海大1号 二倍体和三倍体的受精率(a )与孵化率(b)F i g .1 F e r t i l i z a t i o n r a t e (a )a n d h a t c h i n g r a t e (b )o f d i p l o i d a n d t r i p l o i d P a c i f i c o y s t e r H a i d a N o .1 e x p o s e d t o d i f f e r e n t t e m pe r a t u r e 233期张海宁,等:温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长与存活的影响35ħ时受精率最低,分别为51.67%ʃ2.52%和48.33%ʃ4.16%,且显著低于其他温度组(P<0.05)㊂在相同温度条件下,二倍体组孵化率高于三倍体组但差异不显著(P>0.05),温度为25ħ时,长牡蛎 海大1号 二倍体组与三倍体组的孵化率最高,分别为81.67%ʃ4.55%和76.41%ʃ5.78%;温度为35ħ时孵化率最低,分别为28.80%ʃ4.10%和25.27%ʃ3.91%,且显著低于其他温度组(P<0.05)㊂2.1.2不同盐度对长牡蛎 海大1号 二倍体和三倍体受精率与孵化率的影响盐度为25㊁30和35时,长牡蛎 海大1号 二倍体组受精率显著高于盐度为20和40时(P<0.05);盐度为25㊁30㊁35和40时,长牡蛎 海大1号 三倍体组受精率显著高于盐度为20时(P<0.05),盐度为30和35时二倍体组受精率显著高于三倍体组(P<0.05)(见图2)㊂长牡蛎 海大1号 二倍体组与三倍体组在盐度为30时的受精率最高,分别为92.67%ʃ2.52%和85.33%ʃ2.52%;盐度为20时受精率最低,分别为58.00%ʃ3.00%和59.00%ʃ2.00%,且显著低于其他盐度(P<0.05)㊂盐度为25㊁30和35时,长牡蛎 海大1号 二倍体组和三倍体组孵化率显著高于盐度为20和40时(P< 0.05)㊂盐度为20和35时二倍体组孵化率显著高于三倍体组(P<0.05)㊂盐度为30时 海大1号 二倍体组和三倍体组的孵化率最高,分别为81.67%ʃ4.54%和73.08%ʃ2.07%;盐度为20时 海大1号 二倍体组和三倍体组孵化率最低,分别为55.46%ʃ8.12%和46.27%ʃ3.16%,且显著低于其他盐度组(P<0.05)㊂(同一指标具有相同字母表示组间差异不显著(P>0.05),具有不同字母表示组间存在显著差异(P<0.05)㊂S a m e i n d i c a t o r w i t h t h e s a m e l e t t e r m e a n s t h a t t h e d i f f e r e n c e b e t w e e n g r o u p s i s n o t s i g n i f i c a n t(P>0.05),a n d w i t h d i f f e r e n t l e t t e r s m e a n s t h a t t h e r e i s a s i g n i f i c a n t d i f f e r e n c e b e t w e e n g r o u p s(P< 0.05).)图2不同盐度下长牡蛎 海大1号 二倍体和三倍体的受精率(a)与孵化率(b)F i g.2F e r t i l i z a t i o n r a t e(a)a n d h a t c h i n g r a t e(b)o f d i p l o i d a n d t r i p l o i d P a c i f i c o y s t e r H a i d a N o.1 e x p o s e d t o d i f f e r e n t s a l i n i t y2.2温度和盐度对幼虫生长的影响2.2.1不同温度对长牡蛎 海大1号 二倍体和三倍体幼虫生长的影响各温度水平下,长牡蛎 海大1号 三倍体幼虫始终保持生长优势,其壳高显著高于二倍体幼虫(P<0.05),且三倍体和二倍体幼虫生长最快和最慢的温度分别为30和20ħ(见图3)㊂当温度为35ħ时,第9天二倍体幼虫全部死亡,第13天三倍体幼虫全部死亡㊂第17天,温度为30ħ时,二倍体与三倍体幼虫的壳高达到最大值,分别为(302.17ʃ17.01)μm和(319.67ʃ14.26)μm;而温度为20ħ时,二倍体与三倍体幼虫的壳高最小,分别为(245.51ʃ17.23)μm和(254.05ʃ16.05)μm㊂2.2.2不同盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长的影响各盐度水平下,长牡蛎 海大1号 三倍体幼虫始终保持生长优势,三倍体幼虫的壳高显著高于二倍体幼虫(P<0.05),生长最快和最慢的盐度分别为20和40,而盐度25与30两组之间无明显差异(P>0.05)(见图4)㊂第17天,二倍体和三倍体幼虫的壳高均在盐度20时达到最大值,分别为(271.66ʃ20.43)μm和(320.49ʃ19.83)μm;在盐度40时出现最小值,分别为(213.46ʃ18.26)μm和(223.51ʃ27.78)μm㊂2.3温度和盐度对幼虫存活率的影响2.3.1不同温度对长牡蛎 海大1号 二倍体和三倍体幼虫存活率的影响各温度水平下,长牡蛎 海大1号 三倍体幼虫始终保持存活优势,三倍体幼虫的存活率显著高于二倍体幼虫(P<0.05)(见图5)㊂当温度为35ħ时,第9天二倍体幼虫全部死亡,第13天三倍33中 国 海 洋 大 学 学 报2024年(表示差异显著(P <0.05)㊂d e n o t e s s i gn i f i c a n t d i f f e r e n c e (P <0.05).)图3 不同温度下长牡蛎海大1号 二倍体和三倍体的幼虫壳高F i g .3 S h e l l h e i g h t o f d i p l o i d a n d t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t t e m pe r a t u r es (表示差异显著(P <0.05)㊂d e n o t e s s i gn i f i c a n t d i f f e r e n c e (P <0.05).)图4 不同盐度下长牡蛎海大1号 二倍体和三倍体的幼虫壳高F i g .4 S h e l l h e i g h t o f d i p l o i d a n d t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t s a l i n i t y433期张海宁,等:温度和盐度对长牡蛎 海大1号二倍体和三倍体幼虫生长与存活的影响(表示差异显著(P <0.05)㊂d e n o t e s s i gn i f i c a n t d i f f e r e n c e (P <0.05).)图5 不同温度下长牡蛎海大1号 二倍体和三倍体幼虫的存活率F i g .5 S u r v i v a l r a t e o f d i p l o i d a n d t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t t e m pe r a t u r e s 体幼虫全部死亡㊂第17天,二倍体在25ħ时存活率最高为32.67%ʃ4.04%,与在20ħ时无显著差异(P >0.05);三倍体幼虫在25ħ时存活率最高为52.00%ʃ6.56%,与在20ħ时无显著差异(P >0.05)㊂2.3.2不同盐度对长牡蛎海大1号 二倍体和三倍体幼虫存活率的影响 长牡蛎 海大1号 二倍体幼虫存活率在盐度为25时最高,其次为盐度20,盐度40时存活率最低;三倍体幼虫存活率在盐度为30时最高,其次为盐度25,同样在盐度40时存活率最低;盐度为25时,长牡蛎 海大1号 三倍体幼虫存活率高于二倍体幼虫,但差异不显著(P >0.05);盐度为30㊁35和40时长牡蛎 海大1号 三倍体幼虫存活率显著高于二倍体幼虫(P <0.05)(见图6)㊂2.4温度和盐度对幼虫变态率的影响2.4.1不同温度对长牡蛎海大1号 二倍体和三倍体幼虫变态率的影响 长牡蛎 海大1号 二倍体和三倍体在不同实验温度下的变态率如图7所示㊂长牡蛎海大1号 二倍体与三倍体幼虫变态率在25ħ时最高,在30ħ时最低,不同温度之间具有显著性差异(P <0.05)㊂在相同温度下,二倍体幼虫与三倍体幼虫的变态率无显著性差异(P >0.05)㊂2.4.2不同盐度对长牡蛎海大1号 二倍体和三倍体幼虫变态率的影响 长牡蛎 海大1号 二倍体和三倍体幼虫在不同实验盐度下的变态率如图8所示㊂长牡蛎 海大1号 二倍体幼虫和三倍体幼虫变态率在盐度为30时最高,在盐度为40时最低㊂二倍体和三倍体幼虫的变态率在盐度为20㊁25㊁30和40时无显著性差异(P >0.05),在盐度为35时,三倍体幼虫的变态率显著高于二倍体幼虫(P <0.05)㊂3 讨论3.1温度和盐度对长牡蛎海大1号 二倍体和三倍体受精率与孵化率的影响在本研究中长牡蛎海大1号 二倍体和三倍体的受精率与孵化率随着温度和盐度的升高,呈先升高后降低的趋势,这与H a n 等[13]在长牡蛎二倍体中的研究结果一致㊂在低温条件下,受精率与孵化率较低可能与温度影响受精卵内酶的活性有关,低温对其正常分裂分化产生了不利影响[14]㊂水温过高,会导致水中的53中 国 海 洋 大 学 学 报2024年(表示差异显著(P <0.05)㊂d e n o t e s s i gn i f i c a n t d i f f e r e n c e (P <0.05).)图6 不同盐度下长牡蛎海大1号 二倍体和三倍体幼虫的存活率F i g .6 S u r v i v a l r a t e o f d i p l o i d a n d t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t s a l i n i ty(同一指标具有相同字母表示组间差异不显著(P >0.05),具有不同字母表示组间存在显著差异(P <0.05)㊂S a m e i n d i c a t o r w i t h t h e s a m e l e t t e rm e a n s t h a t t h e d i f f e r e n c e b e t w e e n g r o u p s i s n o t s i gn i f i c a n t (P >0.05),a n d w i t h d i f f e r e n t l e t t e r s m e a n s t h a t t h e r e i s a s i gn i f i c a n t d i f f e r e n c e b e -t w e e n g r o u ps (P <0.05).)图7 不同温度下长牡蛎海大1号 二倍体和三倍体幼虫的变态率F i g .7 M e t a m o r p h o s i s r a t e o f d i p l o i d a n d t r i pl o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t t e m pe r a t u r e s 致病微生物滋生,增加耗氧,这可能是影响受精卵孵化的原因之一[15]㊂温度为25~30ħ,盐度为30~35时,海大1号 二倍体组受精率显著高于三倍体组,表明三倍体组受精卵可能存在一定的 受精卵不育 现象[16],Z h a n g 等[17]研究葡萄牙牡蛎三倍体受精卵与二倍体受精卵的卵裂率时也报道了类似结果㊂在本研究(同一指标具有相同字母表示组间差异不显著(P >0.05),具有不同字母表示组间存在显著差异(P <0.05)㊂S a m e i n d i c a t o r w i t h t h e s a m e l e t t e rm e a n s t h a t t h e d i f f e r e n c e b e t w e e n g r o u p s i s n o t s i gn i f i c a n t (P >0.05),a n d w i t h d i f f e r e n t l e t t e r s m e a n s t h a t t h e r e i s a s i gn i f i c a n t d i f f e r e n c e b e -t w e e n g r o u ps (P <0.05).)图8 不同盐度下长牡蛎海大1号 二倍体和三倍体幼虫的变态率F i g .8 M e t a m o r p h o s i s r a t e o f d i p l o i d a n d t r i pl o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o .1 u n d e r d i f f e r e n t s a l i n i t y中,长牡蛎 海大1号 二倍体在受精与孵化方面的表现优于三倍体㊂在受精和孵化阶段,控制水温在25~30ħ之间,盐度在25~35之间能减少两者之间的差异,提高三倍体的受精率与孵化率㊂3.2温度和盐度对长牡蛎海大1号 二倍体和三倍体幼虫生长与存活的影响贝类苗种培育成功的关键在很大程度上取决于其633期张海宁,等:温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长与存活的影响生命早期阶段的表现,如胚胎和幼虫阶段㊂随着温度升高, 海大1号 二倍体与三倍体幼虫的生长加快㊂这可能由于水温升高使贝类的新陈代谢与摄食消化等能力提高,从而使得幼虫具有更强的运动能力,生长更迅速[18]㊂值得注意的是,幼虫密度没有作为幼虫生长速率条件实验的一个因素进行检查,幼虫在高温下生长最快可能是由温度和幼虫密度的综合作用导致的㊂在温度对幼虫存活影响方面,三倍体幼虫的存活率显著高于二倍体幼虫,这表明三倍体幼虫的温度耐受性高于二倍体幼虫㊂高温条件下幼虫的高死亡率与之前报道的研究一致[13]㊂温度升高到一定程度,可能会导致水中致病微生物的滋生以及贝类血细胞和酶活性降低,从而对贝类的生长发育以及存活产生不利影响㊂在本研究中,在幼虫发育的整个阶段相对于长牡蛎二倍体幼虫 海大1号 三倍体幼虫表现出明显的生长优势,这可能与基因杂合度增加使得其生长性能得到改良有关[19];长牡蛎 海大1号 三倍体与二倍体相比,染色体组增加可能带来了剂量效应,即三倍体细胞内可能出现更多与生长相关的基因,使其生长速度加快㊂这在香港牡蛎(C r a s s o s t r e a h o n g k o n g e n s i s)[20]㊁美洲牡蛎(C r a s s o s t r e a v i r g i n i c a)[21-22]及扇贝(A r g o p e c t e n v e n t r i c o s u s)[23]等多种双壳贝类中已经证实㊂在本研究中,三倍体幼虫表现出快速生长特性,其有助于在苗种培育阶段缩短育苗周期从而降低养殖成本㊂盐度是影响贝类幼虫环境适应能力的另一个重要的非生物因素,对幼虫的生长和存活具有较大影响㊂迄今为止,已经进行了许多关于盐度对软体动物生产性能影响的研究[24-25]㊂在同等环境下,不同物种有不同的适宜盐度范围㊂在我们的研究中,盐度对生长和存活的影响是显著的,长牡蛎 海大1号 二倍体与三倍体幼虫存活率随着盐度的升高呈先升高再下降的趋势;幼虫的生长速度随着盐度的升高呈下降趋势㊂在以前对长牡蛎 海大1号 二倍体幼虫的研究中也发现了类似结果[26]㊂大量研究表明,盐度变化会对海洋贝类渗透压调节产生影响,进而影响个体的生长发育[27-29]㊂在本研究中高盐条件下生长速度降低,这可能是由于幼虫需要消耗大量的能量来进行渗透压调节,进而导致生长速度减缓㊂在高盐条件下,长牡蛎 海大1号 三倍体幼虫相对于二倍体幼虫表现出明显的存活优势,这可能是由于多倍体的染色体增加,从而进化出了新的功能,使潜在的生态位扩张或对环境变化反应的可塑性增强[30-31]㊂长牡蛎 海大1号 三倍体幼虫表现出得对温度和盐度的高耐受力,这可能与染色体组增加所引起的环境适应性增强有关㊂3.3温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫变态率的影响在本研究中,长牡蛎 海大1号 三倍体幼虫相对于二倍体幼虫具有生长快㊁变态时间短的特点㊂在温度实验中,长牡蛎 海大1号 三倍体幼虫与二倍体幼虫的变态率没有表现出明显的差异;而在盐度实验中,在高盐条件下长牡蛎 海大1号 三倍体幼虫相对于二倍体幼虫表现出显著的变态优势㊂总体来看,长牡蛎 海大1号 三倍体幼虫与二倍体幼虫的变态率差异不显著,这说明三倍体幼虫可以正常附着,变态形成稚贝㊂这与在长牡蛎㊁美洲牡蛎㊁香港牡蛎中的研究结果一致[22,32-36]㊂在本研究中,长牡蛎 海大1号 二倍体与三倍体幼虫的变态率受温度与盐度的影响较大,在生产过程中控制温度在25ħ左右,盐度在25~35之间能够有效提高幼虫的变态率㊂4结语为了探究长牡蛎 海大1号 三倍体幼虫对主要环境因子的适应能力,比较研究了温度和盐度对二倍体和三倍体幼虫生长和存活的影响㊂研究显示,长牡蛎 海大1号 三倍体幼虫培育的适宜盐度为20~30,适宜温度为20~25ħ㊂相对于长牡蛎二倍体 海大1号 三倍体在幼虫阶段具有生长快㊁变态时间短的特点,对温度和盐度具有更高的耐受能力㊂长牡蛎 海大1号 三倍体在幼虫阶段表现出的优势有助于提高在牡蛎苗种生产中的效率㊂参考文献:[1]李玲蔚,张哲,马培振,等. 海大1号 长牡蛎规模化人工育苗技术的研究[J].海洋湖沼通报,2017(4):139-144.L i L W,Z h a n g Z,M a P Z,e t a l.S t u d i e s o n t h e t e c h n i q u e s o f l a r g e-s c a l e a r t i f i c i a l r e p r o d u c t i o n o f t h e P a c i f i c O y s t e r H a i d a N o.1 [J].T r a n s a c t i o n s o f O c e a n o l o g y a n d L i m n o l o g y,2017(4):139-144.[2]王昭萍,王如才,于瑞海,等.多倍体贝类的生物学特性[J].青岛海洋大学学报,1998(3):60-65.W a n g Z P,W a n g R C,Y u R H,e t a l.B i o l o g i c a l c h a r a c t e r i s t i c s o f p o l y p l o i d s h e l l f i s h[J].J o u r n a l o f O c e a n U n i v e r s i t y o f Q i n g d a o, 1998(3):60-65.[3] K i n n e O.T h e e f f e c t s o f t e m p e r a t u r e a n d s a l i n i t y o n m a r i n e a n db r ac k i s h w a t e r a n i m a l s:2.S a l i n i t y a nd te m p e r a t u r e-s a l i n i t y r e l a-t i o n s[J].O c e a n o g r a p h y M a r i n e B i o l o g y A n n u a l R e v i e w,1964,2: 281-339.[4]J o y c e A,H o l t h u i s T D,C h a r r i e r G,e t a l.E x p e r i m e n t a l e f f e c t s o ft e m p e r a t u r e a n d p h o t o p e r i o d o n s y n c h r o n y o f g a m e t o g e n e s i s a n d s e x r a t i o i n t h e E u r o p e a n o y s t e r O s t r e a e d u l i s(L i n n a e u s)[J].J o u r n a l o f S h e l l f i s h R e s e a r c h,2013,32(2):447-458. [5]刘德经,张克存.西施舌幼虫及稚贝致死温度初步研究[J].动物学杂志,2001,36(1):29-31.L i u D J,Z h a n g K C.P r e l i m i n a r y s t u d y o n d e a t h t e m p e r a t u r e s o f s h e l l y o u n g a n d l a r v a e o f C o e l l o m a c t r a a n t i q u a t a[J].C h i n e s e J o u r n a l o f Z o o l o g y,2001,36(1):29-31.[6] L i Y G,X u C X,L i Q.E f f e c t s o f s a l i n i t y a n d t e m p e r a t u r e o n73中国海洋大学学报2024年g r o w t h a n d s u r v i v a l o f d i p l o i d a n d t e t r a p l o i d l a r v a e o f t h e P a c i f i co y s t e r,C r a s s o s t r e a g i g a s[J].A q u a c u l t u r e,2022,550:737809.[7]沈伟良,尤仲杰,施祥元.温度与盐度对毛蚶受精卵孵化及幼虫生长的影响[J].海洋科学,2009,33(10):5-8.S h e n W L,Y o u Z J,S h i X Y.T h e i n f l u e n c e o f t e m p e r a t u r e a n d s a l i n i t y o n z y g o t e s h a t c h i n g r a t e a n d l a r v a l g r o w t h o f S c a p h a r c a s u b c r e n a t a[J].M a r i n e S c i e n c e s,2009,33(10):5-8. [8]叶乐,赵旺,王雨,等.盐度与p H对长肋日月贝幼虫存活及生长的影响[J].南方农业学报,2015,46(9):1689-1703.Y e L,Z h a o W,W a n g Y,e t a l.E f f e c t s o f s a l i n i t y a n d p H o n s u r-v i v a l a n d g r o w t h o f A m u s i u m p l e u r o n e c t e s l a r v a e[J].J o u r n a l o f S o u t h e r n A g r i c u l t u r e,2015,46(9):1689-1703.[9]梁玉波,张福绥.温度㊁盐度对栉孔扇贝(C h l a m y s f a r r e r i)胚胎和幼虫的影响[J].海洋与湖沼,2008,39(4):334-340.L i a n g Y B,Z h a n g F S.E f f e c t s o f t e m p e r a t u r e a n d s a l i n i t y o n d e-v e l o p m e n t o f e m b r y o s a n d l a r v a e o f s c a l l o p C h l a m y s f a r r e r i[J].O c e a n o l o g i a a n d L i m n o l o g i a S i n i c a,2008,39(4):334-340. [10] W a n g T,L i Q.E f f e c t s o f s a l i n i t y a n d t e m p e r a t u r e o n g r o w t h a n ds u r v i v a l o f j u v e n i l e i w a g a k i o y s t e r C r a s s o s t r e a n i p p o n a[J].J o u r-n a l o f O c e a n U n i v e r s i t y o f C h i n a,2018,17(4):234-239. [11]杨东敏,张艳丽,丁鉴锋,等.高温㊁低盐对菲律宾蛤仔免疫能力的影响[J].大连海洋大学学报,2017,32(3):302-309.Y a n g D M,Z h a n g Y L,D i n g J F,e t a l.S y n e r g i s t i c e f f e c t s o fh i g h t e m p e r a t u r e a n d l o w s a l i n i t y o n i m m u n i t y o f M a n i l a c l a mR u d i t a p e s p h i l i p p i n a r u m[J].J o u r n a l o f D a l i a n O c e a n U n i v e r s i-t y,2017,32(3):302-309.[12]张柯馨,曹楚畑,刘志刚,等.钝缀锦蛤(T a p e s d o r s a t u s)稚贝的温度和盐度耐受性研究[J].海洋学报,2022,44(4):57-64.Z h a n g K X,C a o C T,L i u Z Q,e t a l.S t u d y o n t e m p e r a t u r e a n d s a l i n i t y t o l e r a n c e o f T a p e s d o r s a t u s j u v e n i l e s[J].H a i y a n g X u e-b a o,2022,44(4):57-64.[13] H a n Z Q,L i Q.D i f f e r e n t r e s p o n s e s b e t w e e n o r a n g e v a r i a n t a n dc u l t u r ed p o p u l a t i o n o f t he P a c if i c o y s t e r C r a s s o s t r e ag i g a s a t e a r l yl i f e s t a g e t o t e m p e r a t u r e-s a l i n i t y c o m b i n a t i o n s[J].A q u a c u l t u r e R e s e a r c h,2018,49(6):2233-2239.[14] G a g n a i r e B,F r o u i n H,M o r e a u K,e t a l.E f f e c t s o f t e m p e r a t u r ea n d s a l i n i t y o n h a e m o c y t e a c t i v i t i e s o f t h e P a c i f i c o y s t e r,C r a s-s o s t r e a g i g a s(T h u n b e r g)[J].F i s h&S h e l l f i s h I m m u n o l o g y, 2006,20(4):536-547.[15] G r u f f y d d L D,B e a u m o n t A R.A m e t h o d f o r r e a r i n g P e c t e nm a x i m u s l a r v a e i n t h e l a b o r a t o r y[J].M a r i n e B i o l o g y,1972,15: 350-355.[16]M a t t J L,A l l e n S K.H e t e r o p l o i d m o s a i c t e t r a p l o i d s o f C r a s-s o s t r e a v i r g i n i c a p r o d u c e n o r m a l t r i p l o i d l a r v a e a n d j u v e n i l e s a s r e v e a l e d b y f l o w c y t o m e t r y[J].A q u a c u l t u r e,2014,432(2):336-345.[17]Z h a n g Y H,Q i n Y P,Y u Z N.C o m p a r a t i v e s t u d y o f t e t r a p l o i d-b a s e d r ec i p r o c a l t r i p l o id P o r t u g ue s e o y s t e r s,C r a s s o s t r e a a n g u l a-t a,f r o m s e e d t o m a r k e t s i z e[J].A q u a c u l t u r e,2022,547: 737523.[18] R i c o-V i l l a B,P o u v r e a u S,R o b e r t R.I n f l u e n c e o f f o o d d e n s i t ya n d t e m p e r a t u r e o n i n g e s t i o n,g r o w t h a n d s e t t l e m e n t o f P a c i f i co y s t e r l a r v a e,C r a s s o s t r e a g i g a s[J].A q u a c u l t u r e,2009,287: 395-401.[19] W a n g Z P,G u o X M,A l l e n S K,e t a l.H e t e r o z y g o s i t y a n d b o d ys i z e i n t r i p l o i d P a c i f i c o y s t e r s,C r a s s o s t r e a g i g a s T h u n b e r g,p r o-d u ce df r o m m e i o s i sⅡi n h i b i t i o n a n d t e t r a p l o i d s[J].A q u a c u l t u r e,2002,204(3-4):337-348.[20]秦艳平,张跃环,莫日馆,等.香港牡蛎二倍体及三倍体幼虫生物学特征比较[J].海洋科学,2020,44(3):123-128.Q i n Y P,Z h a n g Y H,M o R G,e t a l.C o m p a r i s o n o f b i o l o g i c a lc h a r a c t e r i s t i c s b e t w e e nd i p l o i d a n d t r i p l o i d C r a s s o s t re a h o n g k o n-g e n s i s l a r v a e[J].M a r i n e S c i e n c e,2020,44(3):123-128.[21] D e g r e m o n t L,G a r c i a C,F r a n k L,e t a l.T r i p l o i d o y s t e r s i n t h eC h e s a p e a k e B a y:C o m p a r i s o n o f d i p l o i d a n d t r i p l o i d C r a s s o s t r e av i r g i n i c a[J].J o u r n a l o f S h e l l f i s h R e s e a r c h,2012,31(1):21-31.[22] C a l l a m B R,A l l e n S K,F r a n k-L a w a l e A.G e n e t i c a n d e n v i r o n-m e n t a l i n f l u e n c e o n t r i p l o i d C r a s s o s t r e a v i r g i n i c a g r o w n i n C h e s-a p e a k e B a y:G r o w t h[J].A q u a c u l t u r e,2016,452:97-106.[23]C e s a r A,R a m r e z a J,S t a n d i s h K,e t a l.T r i p l o i d c a t a r i n a s c a l l o p(A r g o p e c t e n v e n t r i c o s u s S o w e r b y I I,1842):G r o w t h,g a m e t o-g e n e s i s,a n d s u p p r e s s i o n o f f u n c t i o n a l h e r m a p h r o d i t i s m[J].A q-u a c u l t u r e,2000,186(1-2):13-32.[24]J i C,C a o L,L i F.T o x i c o l o g i c a l e v a l u a t i o n o f t w o p e d i g r e e s o fc l a m R ud i t a pe s p h i l i p p i n a r u m a s b i o i n d i c a t o r s of h e a v y m e t a lc o n t a m i n a n t s u s i n g m e t a b o l o m i c s[J].E n v i r o n T o x i c o l P h a r m a-c o l,2015,39:545-554.[25] K i m W,H u h H,H u h S H,e t a l.E f f e c t s o f s a l i n i t y o n e n d o g e-n o u s r h y t h m o f t h e M a n i l a c l a m,R u d i t a p e s p h i l i p p i n a r u m(B i-v a l v i a:V e n e r i d a e)[J].M a r i n e B i o l o g y,2001,138:157-162.[26]孟乾,李琪,张景晓.水温和盐度对长牡蛎 海大1号 早期生长发育的影响[J].海洋科学,2017,41(12):32-37.M e n g Q,L i Q,Z h a n g J X.E f f e c t o f t e m p e r a t u r e a n d s a l i n i t y o ne m b r y o n i c d e v e l o p m e n t,g r o w t h,s u r v i v a l,a n d s e t t i n g of t h e P a-c i f i c o y s t e r H a id a N o.1 l a r v a e[J].M a r i ne S c i e n c e,2017,41(12):32-37.[27] H i s E,B e i r a s R,S e a m a n M N L.T h e a s s e s s m e n t o f m a r i n e p o l-l u t i o n-b i o a s s a y s w i t h b i v a l v e e m b r y o s a n d l a r v a e[J].A d v a n c e s i nM a r i n e B i o l o g y,1999(37):1-178.[28] O'C o n n o r W A,L a w l e r N F.S a l i n i t y a n d t e m p e r a t u r e t o l e r a n c eo f e m b r y o s a n d j u v e n i l e s o f t h e p e a r l o y s t e r,P i n c t a d a i m b r i c a t aRÖd i n g[J].A q u a c u l t u r e,2004,229(1):493-506. [29]宁军号,庞云龙,宋坚,等.偏顶蛤胚胎和幼虫发育及温度对其浮游幼虫生长和发育的影响[J].中国水产科学,2015,22(2): 196-203.N i n g J H,P a n g Y L,S o n g J,e t a l.D e v e l o p m e n t o f e m b r y o a n d l a r v a o f M o d i o l u s m o d i o l u s a n d e f f e c t s o f t e m p e r a t u r e o n g r o w t ha n d d e v e l o p m e n t o f t h e p e l a g i c l a r v a[J].J o u r n a l o f F i s h e r y S c i-e n c e s of C h i n a,2015,22(2):196-203.[30] A d a m s K L,W e n d e l J F.P o l y p l o i d y a n d g e n o m e e v o l u t i o n i np l a n t s[J].C u r r e n t O p i n i o n i n P l a n t B i o l o g y,2005,8(2):135-141.[31]M o o r e R C,P u r u g g a n a n M D.T h e e v o l u t i o n a r y d y n a m i c s o fp l a n t d u p l i c a t e g e n e s[J].C u r r e n t O p i n i o n i n P l a n t B i o l o g y, 2005,8(2):122-128.[32] Q i n Y P,Z h a n g Y H,M o R G,e t a l.I n f l u e n c e o f p l o i d y a n d e n-v i r o n m e n t o n g r o w-o u t t r a i t s o f d i p l o i d a n d t r i p l o i d H o n g K o n go y s t e r s C r a s s o s t r e a h o n g k o n g e n s i s i n s o u t h e r n C h i n a[J].A q u a-c u l t u r e,2019,507:108-118.[33]I b a r r a A M,A s c e n c i o m i c h e l R,R a mír e z J L,e t a l.P e r f o r m a n c e833期张海宁,等:温度和盐度对长牡蛎 海大1号 二倍体和三倍体幼虫生长与存活的影响o f d i p l o i d a n d t r i p l o i d C r a s s o s t r e a g i g a s(T h u n b e r g,1793)g r o w ni n t r o p i c a l v e r s u s t e m p e r a t e n a t u r a l e n v i r o n m e n t a l c o n d i t i o n s[J].J o u r n a l o f S h e l l f i s h R e s e a r c h,2017,36(1):119-139.[34] A l l e n S K,D o w n i n g S L.P e r f o r m a n c e o f t r i p l o i d P a c i f i c o y s t e r s,C r a s s o s t r e a g i g a s(T h u n b e r g).I.S u r v i v a l,g r o w t h,g l y c o g e nc o n t e n t,a nd se x u a l m a t u r a t i o n i n y e a r l i n g s[J].J o u r n a l of E x-p e r i m e n t a l M a r i n e B i o l o g y a n d E c o l o g y,1986,102(2-3):197-208.[35] N e l l J A.F a r m i n g t r i p l o i d o y s t e r s[J].A q u a c u l t u r e,2002,210(1-4):69-88.[36] N e l l J A,P e r k i n s B.S t u d i e s o n t r i p l o i d o y s t e r s i n A u s t r a l i a:F a r m i n g p o t e n t i a l o f a l l-t r i p l o i d P a c i f i c o y s t e r s,C r a s s o s t r e ag i g a s(T h u n b e r g),i n P o r t S t e p h e n s,N e w S o u t h W a l e s,A u s-t r a l i a[J].A q u a c u l t u r e R e s e a r c h,2005,36(6):530-536.E f f e c t s o f T e m p e r a t u r e a n d S a l i n i t y o n t h e G r o w t h a n d S u r v i v a l o f D i p l o i d a n dT r i p l o i d L a r v a e o f P a c i f i c O y s t e r H a i d a N o.1Z h a n g H a i n i n g1,X u C h e n g x u n1,L i Q i1,2(1.K e y L a b o r a t o r y o f M a r i c u l t u r e(O c e a n U n i v e r s i t y o f C h i n a),M i n i s t r y o f E d u c a t i o n,Q i n g d a o266003,C h i n a;2.L a b o-r a t o r y f o r M a r i n e F i s h e r i e s S c i e n c e a n d F o o d P r o d u c t i o n P r o c e s s e s,P i l o t N a t i o n a l L a b o r a t o r y f o r M a r i n e S c i e n c e a n d T e c h-n o l o g y(Q i n g d a o),Q i n g d a o266237,C h i n a)A b s t r a c t:I n o r d e r t o p r o m o t e t h e d e v e l o p m e n t o f t h e t r i p l o i d i n d u s t r y o f a n e x c e l l e n t v a r i e t y o f P a c i f i c o y s t e r C r a s s o s t r e a g i g a s H a i d a N o.1 ,a n d o p t i m i z e i t s r e a r i n g c o n d i t i o n s a n d i m p r o v e t h e s u r v i v a l a n d g r o w t h r a t e,t h e e f f e c t s o f d i f f e r e n t t e m p e r a t u r e s a n d s a l i n i t i e s o n t h e g r o w t h a n d s u r v i v a l o f d i p l o i d a n d t r i p l o i d l a r v a e w e r e i n v e s t i g a t e d.F o u r t e m p e r a t u r e s(20,25,30a n d35ħ)a n d f i v e s a l i n i t i e s(20, 25,30,35a n d40)w e r e s e t u p f o r t h e e x p e r i m e n t s.T h e h i g h e s t f e r t i l i z a t i o n,h a t c h i n g a n d m e t a m o r p h o s i s r a t e s o f t h e d i p l o i d a n d t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o.1 w e r e f o u n d w h e n t h e t e m p e r a t u r e w a s25ħa n d s a l i n i t y w a s30.A t d i f f e r e n t s a l i n i t i e s,t h e g r o w t h a n d c u m u l a t i v e s u r v i v a l r a t e s o f t r i p l o i d l a r v a e w e r e s i g n i f i c a n t l y h i g h e r t h a n t h o s e o f d i p l o i d l a r v a e.T h e g r o w t h a n dc u m u l a t i v e s u r v i v a l r a t e s o f t r i p l o id l a r v ae w e r e c o n s i s t e n t l y h i g h e r t h a n t h o s e of d i p l o i d l a r v a e a td i f fe r e n t t e m p e r a t u r e s.T h e s u r v i v a l r a t e of t r i p l o i d l a r v a e d e c r e a s e d w i t h t e m p e r a t u r e,r e a c h i ng th e hi g h e s t a t25ħ(52.00%ʃ6.56%).T h e f a s t e s t g r o w t h i n s h e l l h e i g h t o f t r i p l o i d l a r v a e w a s o b s e r v e d a t30ħ(14.82μm㊃d-1).T h e s u r v i v a l r a t e o f t r i p l o i d l a r v a e i n c r e a s f i r s t a n d t h e n d e c r e a s e d w i t hi n c r e a s i n g s a l i n i t y,a n d t h e c u m u l a t i v e s u r v i v a l r a t e o f t r i p l o i d l a r v a e w a s h i g h e s t a t30(52.00%ʃ6.56%).T h e g r o w t h r a t e o f l a r v a e d e c r e a s e d w i t h i n c r e a s i n g s a l i n i t y,a n d t h e s h e l l h e i g h t o f t r i p l o i d l a r v a e g r e w f a s t e s t a t20(18.22μm㊃d-1).I n c o n c l u s i o n,t h e t r i p l o i d l a r v a e o f P a c i f i c o y s t e r H a i d a N o.1 s h o w e d s t r o n g e r t e m p e r a t u r e a n d s a l i n i t y t o l e r a n c e s t h a n t h e d i p l o i d l a r v a e.T h e o p t i m u m s a l i n i t y f o r t h e c u l t i v a t i o n o f t r i p l o i d P a c i f i c o y s t e r H a i d a N o.1 r a n g e d f r o m20t o30w i t h t h e m o s t s u i t a b l e v a r i e d b e t w e e n20a n d25ħ.O u r f i n d i n g s p r o v i d e d i m p o r t a n t i n f o r m a t i o n f o r t h e l a r g e-s c a l e b r e e d i n g o f t h e t r i p l o i d s e e d s o f P a c i f i c o y s t e r H a i d a N o.1 .K e y w o r d s:P a c i f i c o y s t e r;t r i p l o i d l a r v a;t e m p e r a t u r e;s a l i n i t y;g r o w t h;s u r v i v a l;H a i d a N o.1责任编辑朱宝象93。
三倍体牡蛎培育原理

三倍体牡蛎培育原理
三倍体牡蛎培育是一种养殖技术,通过使牡蛎细胞的染色体数目变为三倍体,旨在提高牡蛎的生长速度、抗逆性以及产量。
以下是三倍体牡蛎培育的基本原理:
1. 化学诱导:三倍体牡蛎的培育通常通过化学诱导实现。
在培育过程中,将二倍体牡蛎的卵子暴露在化学诱导剂(例如胱氨酸)中。
这些化学诱导剂能够干扰卵子的有丝分裂过程,导致多余的染色体在卵子中复制,形成三倍体。
2. 杂交:另一种常见的方法是通过二倍体雌性牡蛎的卵子与三倍体雄性牡蛎的精子进行杂交。
这种方法也能产生三倍体牡蛎,因为杂交导致了染色体的加倍。
3. 染色体加倍:通过这些方法,培育出的牡蛎细胞染色体数目由正常的二倍体变成了三倍体。
由于有额外的染色体,这些牡蛎通常具有更强的生长能力和更高的抗逆性。
培育三倍体牡蛎的目标是增加牡蛎的生产性能,提高其抗逆性,同时也可以改良其他性状,比如提高肉质品质。
这种技术的成功应用可以对养殖业产生积极影响,提高牡蛎的经济效益。
1/ 1。
不同卵密度对太平洋牡蛎三倍体诱导效果影响的研究

关键 词 :6DMA 、 太平 洋牡 蛎 ,受精 卵密度 、 三倍 体 率 。 - P
中图分类号 :¥4 . 1 944
文献标识码 :A
—
翮 1 上 ・
— 青 j 一ຫໍສະໝຸດ 牡蛎是世界主要的养殖贝类 ,营养丰富 .俗称“ 中牛奶” 海 深受 国内外市场 的欢迎 ,但在 繁 殖季 节 , 由于 眭腺 发 育 ,消 耗 了大量 肝 糖 元 ,牡 蛎 停 止 生 长 甚 至 消 瘦 ,大 大 影 响 了牡 蛎 的 风味 ,而且在繁殖季节还会出现大量死亡的现象 ,并 且随着 目前养殖环境 的恶化 ,养殖品种 自身种质 的老化 ,疾病所造成 的危害越来越大 ,而三倍体具有育性差 、个体大 、生长快 、肉 质好 、抗病能力强等优点 ,显示 了牡蛎三倍体在养殖上 的广阔前景I ” 。 自18 年美 国人获得美洲牡蛎多倍体 以来 ,美国 、 日本等 国的许多学者利用 温度休克和 91 绌胞松驰素 B诱 导法 ,对太平洋牡蛎 、欧洲牡蛎及其它 贝类进行 多倍体研 究 ,取得 了不少 研究成果 ,有 的已经应用于生产 【 2 1 。在我 国,18 年正式进行 了牡蛎三倍体育苗 的研究 。当 98
维普资讯
海 洋 湖 沼 通 报
2 0 矩 02 Tr n a t n f e o o y a d L m n l g a s c i so a l g i o o y o Oc n n N 1 o
文 章 编号 : 10 --4 2( 0 2)0 —O 5 —O 3- 8 20 0 6 2 07 5
洋牡 蛎 的 受精 卵进 行 三 倍 体 诱 导 ,研 究 受精 卵的 密度 对 诱 导 效 果 的 影 响 , 以及 诱 导
后 受精 卵的孵化率和畸形率 。实验结果表 明:"受精 卵的密度为 4 万个/ 3 - ml时三倍
太平洋牡蛎的繁殖习性如何

太平洋牡蛎的繁殖习性如何太平洋牡蛎的性细胞在繁殖季节是分批成熟和排放的,因此繁殖期相当长。
它的性腺成熟、繁殖与水温、比重、潮汐关系较为密切。
太平洋牡蛎的性腺在水温16℃时开始形成,20℃逐渐增多,22℃-23℃时性腺成熟并开始排精产卵。
太平洋牡蛎初产时海水比重为1.016左右,未达到这个比重范围也开始大批量繁殖。
另外,太平洋牡蛎在海区大批量的繁殖活也开始大批量繁殖。
另外,太平洋牡蛎在海区大批量的繁殖活动多在大潮汐前1-2天或雨后,这主要是由于大潮期间水温、比重变化较大,对其排放精卵起着刺激诱导作用。
确定太平洋牡蛎的繁殖期,除了观测海况变化外,还得仔细地进行亲贝性腺成熟度的观测。
性腺成熟度一般采用测定生殖腺指数的办法,即生殖腺重量与整个软体部分重量的百分比(生殖腺指数=生殖腺重量/软体重量X100%)。
生殖腺指数的增减,可以反映出太平洋牡蛎的繁殖期。
当生殖腺指数的百分比逐渐增加,至45%左右时,则说明太平洋牡蛎的性腺已成熟。
太平洋牡蛎的繁殖属卵生型,即以胚胎发生直到匍匐附着之前是在海水中过浮游生活。
繁殖时,亲贝把精卵排放于海水中,营体外受精。
受精卵在水温23℃、比重1.019-1.022的条件下,经4小时左右发育成球形,且周身密生短小纤毛开始转动,称囊胚期。
受精后Il小时30分左右,发育成担轮幼虫。
担轮幼虫开始分泌贝壳,待到两片贝壳披盖整个软体,口前纤毛带增生细胞,并相对前移呈盘状,其上丛生纤毛,称面盘幼虫,此时大约在受精后18小时出现。
随着幼虫发育,壳顶隆起,称壳顶幼虫,当壳长在280微米左右时,出现眼点后面盘开始萎缩。
这时在口与肛门之间的棒状足逐渐发达,遇到适宜的附着物便在物体上爬行,并分泌足丝附着,幼体附着l-2天,足萎缩消失,进入固着生活。
太平洋牡蛎细菌菌相分析及-20℃冻藏过程中菌相变化

太平洋牡蛎细菌菌相分析及-20℃冻藏过程中菌相变化
太平洋牡蛎细菌菌相分析及-20℃冻藏过程中菌相变化
以胶州湾太平洋牡蛎(Crassostrea gigas)为研究对象,分析了生鲜牡蛎的细菌菌相及在-20℃冻藏过程中的菌相变化情况.研究发现,生鲜牡蛎的初始菌相比较复杂,从生鲜牡蛎中分离到的85株细菌分别属于14个属(科),其中Pseudomonas和Vibrionaceae为优势菌,比例分别为22.4%和20.0%.冻藏过程中,各属(科)细菌的数量都呈下降趋势,Flavobacterium、Staphylococcus、Micrococcus、Lactococcus 和Lactobacillus由于其较强的耐冻性,成为牡蛎冻藏过程中的优势菌.
作者:曹荣薛长湖许加超李兆杰作者单位:中国海洋大学食品科学与工程学院,青岛,266003 刊名:食品与发酵工业 ISTIC PKU 英文刊名: FOOD AND FERMENTATION INDUSTRIES 年,卷(期):2006 32(3) 分类号: Q93 关键词:太平洋牡蛎细菌菌相 -20℃冻藏菌相变化。
太平洋牡蛎二倍体和三倍体的生长比较

太平洋牡蛎二倍体和三倍体的生长比较
曾志南;林琪
【期刊名称】《上海水产大学学报》
【年(卷),期】1999(008)002
【摘要】比较研究了在福建养殖的太平洋特蛎二倍体和三倍休的生长,结果表明,经17个月养殖的太平洋牡蛎三倍体壳长、壳高、体重和软体部重量分别比二倍体增加14.41%、7.66%、35.27%和73.25%。
在太平洋特蛎繁殖季节5 ̄8月,三倍体体重和软体部重量生长明显超过二倍体.还就温带及亚热带海区养殖的太平洋牡蛎二倍体和三倍体的生长进行讨论。
【总页数】5页(P119-123)
【作者】曾志南;林琪
【作者单位】福建省水产研究所厦门;福建省水产研究所厦门
【正文语种】中文
【中图分类】S968.321
【相关文献】
1.太平洋牡蛎杂交三倍体与诱导三倍体的生长比较 [J], 孔静;王昭萍;于瑞海;张跃环;刘剑;李晓瑜;李雅琳;郭希明
2.合浦珠母贝二倍体、三倍体和非整倍体群体的基因杂合度与生长比较 [J], 何毛贤;沈琪;林岳光;姜卫国;潘金培
3.栉孔扇贝三倍体与二倍体的生长比较 [J], 杨爱国;王清印;张岩;刘志鸿;刘萍
4.僧帽牡蛎三倍体和二倍体的生长比较 [J], 曾志南;林琪;吴建绍
5.合浦珠母贝三倍体和二倍体的生长比较 [J], 姜卫国;许国强;林岳光;李刚
因版权原因,仅展示原文概要,查看原文内容请购买。
虾夷扇贝二倍体与三倍体脂肪酸组成的比较

摘要 :采用毛细管色谱柱对虾夷扇贝二倍体与三倍体的脂肪酸组成进行了检测 。结果表明 : ①两者闭壳肌和 外套膜的脂肪酸组成相似 ,特征为多烯酸 ( PUFA) > 饱和脂肪酸 (SFA) > 单烯酸 (MUFA) , PUFA 以 n23 系列为 主 ,主要脂肪酸为 22∶6n23 (23. 9 %~28. 9 %) 、20∶5n23 (17. 6 %~20. 6 %) 、16∶0 (11. 0 %~19. 4 %) 、18∶0 (7. 3 %~ 8. 1 %) 、20∶4n26 (2. 8 %~5. 5 %) ; ②生殖腺的脂肪酸种类相似 , 但两者多数脂肪酸的含量差异显著 , 其中三倍 体的 SFA 和 MUFA 含量显著高于二倍体 ( P < 0. 01) ,而 PUFA 含量显著低于二倍体 ( P < 0. 01) 。 关键词 :虾夷扇贝 ; 二倍体 ; 三倍体 ; 脂肪酸 中图分类号 :S963. 161 文献标识码 : A
1. 1 材料
虾夷扇贝二倍体和三倍体样品 (表 1) 于 2000 年 4 月取自大连长海县獐子岛。
1. 2 倍性检验
用 Partec PAS - Ⅲ型细胞流式仪检测倍性 。
1. 3 脂肪酸甲酯的制备
用生理盐水将贝体洗净 ,活体解剖 ,分别取闭壳肌 、外套膜 、生殖腺装于塑料袋中速冻于 - 28 ℃冰柜 中保存 。用玻璃匀浆器匀浆待测样品 ,然后准确称取 1g 样品 , 加入 2 %的 B HT 甲醇溶液 。按照改进的 Folch 法[8]萃取脂质 。将所得脂质用 KOH - 甲醇于 70 ℃下皂化 1h 后 , 用 B F3 催化法[9] 制取脂肪酸甲 酯 。最后转移浓缩到石油醚中供色谱分析用 。
三倍体太平洋牡蛎不同组织中挥发性风味成分分析

三倍体太平洋牡蛎不同组织中挥发性风味成分分析郑泽文;周子权;陈志;陈建楠;薛婷;林岗【期刊名称】《福建农业科技》【年(卷),期】2024(55)2【摘要】为探究太平洋牡蛎不同组织中挥发性风味成分的差异,揭示牡蛎总体风味的来源及形成机制。
采用SPME-GC-MS技术结合多元统计分析的方法,对太平洋牡蛎闭壳肌、肝胰腺、鳃和性腺的挥发性风味成分进行了检测。
结果表明:闭壳肌中检测到18种物质,肝胰腺中检测到11种物质,鳃中检测到8种物质,性腺中检测到6种物质。
闭壳肌、鳃和性腺中主要成分为醛类物质,肝胰腺中主要成分为醇类物质,4个组织中共检测出28种挥发性风味成分,其中大多数化合物为醛类、酮类、醇类等;采用聚类热图分析、关键风味成分筛选以及主成分分析比较4个不同组织的挥发性风味成分,结果显示太平洋牡蛎中各组织风味差异明显,影响太平洋牡蛎风味的主要物质为醛类和酮类,太平洋牡蛎不同组织中的各种风味物质相互协调,造就了太平洋牡蛎独特的海洋鲜香风味。
研究结果对指导太平洋牡蛎风味研究和产品加工利用有重要意义。
【总页数】9页(P1-9)【作者】郑泽文;周子权;陈志;陈建楠;薛婷;林岗【作者单位】福州海洋研究院海洋活性物质与制品技术研发中心;福建省淡水水产研究所【正文语种】中文【中图分类】TS254.4【相关文献】1.南疆多浪羊不同部位脂肪组织中挥发性风味成分分析2.SPME-GC-MS分析近江牡蛎酶解液挥发性风味成分3.工厂化栽培的不同杏鲍菇菌株非挥发性风味成分分析及风味评价4.利用概率加权回归和主成分分析评估低盐诱导太平洋牡蛎三倍体幼虫的效果5.不同犊牛肉加工产品中挥发性风味成分分析因版权原因,仅展示原文概要,查看原文内容请购买。
二倍体和三倍体牡蛎的外观鉴别

二倍体和三倍体牡蛎的外观鉴别
曾志南;林琪;吴建绍;陈木
【期刊名称】《海洋科学》
【年(卷),期】1999()1
【摘要】对二倍体、三倍体僧帽牡蛎和太平洋牡蛎外套膜和生殖腺外部形态进行
了周年观察。
结果表明,从外套膜和生殖腺外观特征上的差异可鉴别出牡蛎二倍
体和三倍体。
1997年8月至1998年6月,分析了两种牡蛎样品266个,其鉴别准确率达96.99%。
【总页数】3页(P51-53)
【关键词】僧帽牡蛎;太平洋牡蛎;二倍体;三倍体;牡蛎
【作者】曾志南;林琪;吴建绍;陈木
【作者单位】福建省水产研究所
【正文语种】中文
【中图分类】Q959.215.3
【相关文献】
1.温度对不同规格长牡蛎三倍体和二倍体代谢影响的比较 [J], 周一兵;黄璞祎;李晓艳;杨大佐;张国范
2.盐度对三倍体和二倍体太平洋牡蛎代谢影响的比较 [J], 李晓艳
3.饥饿对二倍体和三倍体长牡蛎呼吸和排泄的影响 [J], 王芳;王昭萍;董少帅;吕宝
强
4.香港牡蛎二倍体及三倍体幼虫生物学特征比较 [J], 秦艳平; 张跃环; 莫日馆; 李军; 肖述; 马海涛; 喻子牛
5.熊本牡蛎三倍体与二倍体性腺发育特征比较 [J], 武祥伟;张跃环;肖述;秦艳平;喻子牛
因版权原因,仅展示原文概要,查看原文内容请购买。
能量收支在水产养殖中的应用

能量收支在水产养殖中的应用冯硕恒;米海峰;张璐;潘化祥;王武刚;吴业阳;孙瑞健【摘要】The research on energy budget model in aquaculture can not only assess the position and role of the aqua-cultural organisms in the ecosystem,but also evaluate the effects of the aqua-cultural organisms on the environment.This article summarized the progress and research status of the energy budget model in aquaculture in respect of both aquatic ecosystem and animal individual.It explained the reasons for the growth difference of aqua-cultural organisms with differ-ent environmental factors,feed and breeding methods,in the mean time,it also provided strong supports for the highly effi-cient and sustainable development of the aquaculture industry.%通过对水产养殖中能量收支模型的研究,不仅可以对养殖生物体在生态系统中所占的位置和所起的作用进行评定,同时可以对养殖生物对环境产生的影响进行评估。
本文从水域生态系统和动物个体水平上综述水产养殖中能量收支的研究现状。
一方面解释了不同环境因子、饵料、养殖方式使养殖生物体生长产生差异的原因,同时还为水产养殖产业高效、可持续发展提供有力支撑。
开始试养3倍体牡蛎

开始试养3倍体牡蛎
孙国凤
【期刊名称】《生物技术通报》
【年(卷),期】1995(000)003
【摘要】广岛县最近宣布9月开始试养3倍体牡蛎。
操作染色体而生产的鱼介类符合水产厅规定的“三倍体鱼等的水产生物的利用要领”,6月才能接受认可并实施。
养殖业者进行养殖,试验场和县合作进行监控。
估计最早于95年10月上市。
一到夏季,由于2倍体牡蛎生殖器官的发育吸收营养,所以牡蛎瘦小没有商品价值。
但由于3倍体牡蛎生殖器官不发育,所以在没有二倍体商品时也可以上市。
特别是与广岛县竞争牡蛎产地的宫城县相比,水温高,牡蛎的上市时间晚。
期望3倍体牡蛎能弥补这些缺憾。
【总页数】1页(P33-33)
【作者】孙国凤
【作者单位】
【正文语种】中文
【中图分类】S968.3
【相关文献】
1.美国的单体牡蛎在威海试养成功 [J], 缪国荣
2.太平洋牡蛎的渤海金州湾试养结果的报告 [J], 刘庆余
3.北海湾近江牡蛎试养区采苗试验的初步探讨 [J], 梁广耀
4.日本试养引进的三倍体牡蛎 [J], 孙国凤
5.“橙色牡蛎”在山东威海荣成下海试养 [J], 齐鲁晚报
因版权原因,仅展示原文概要,查看原文内容请购买。
太平洋牡蛎多倍体研究

太平洋牡蛎多倍体研究
田传远;赵厚均
【期刊名称】《黄渤海海洋》
【年(卷),期】1999(017)002
【摘要】本文综述了太平洋牡蛎多倍体产生的途径、诱导方法、诱导结果、诱导
机制、倍性鉴别方法、繁殖、生长、生理指标、抗逆性和口味等。
提出:6-DMAP诱导牡蛎三倍体的技术前景广阔;四倍体与二倍体杂交的方法是产生三倍体的最好途径;四倍体是生物方法产生三倍体的中间材料,可以存活,应加大力度地研究和开发;三倍体的性腺能够发育,并可产生具有繁殖力的配子;在良好的环境里,四倍体的生长和抗逆性较二倍体优势;在繁殖季节,三
【总页数】9页(P75-83)
【作者】田传远;赵厚均
【作者单位】青岛海洋大学水产学院;山东省渔业技术推广站
【正文语种】中文
【中图分类】S968.321
【相关文献】
1.咖啡因+热刺激法诱导太平洋牡蛎多倍体研究初报 [J], 王菊生
2.关于太平洋牡蛎的养殖技术之二太平洋牡蛎工厂化育苗技术的研究 [J], 曾明江
3.苎麻多倍体诱导研究Ⅰ.苎麻多倍体诱导的基础研究 [J], 董延瑜;郑思乡;洪亚辉;任春梅;刘国民;邓明其;程尧楚
4.四川省桑属(Morus)植物多倍体的研究Ⅱ、四川省桑树多倍体的地理分布(西南
农业大学与四川省丝绸公司合作研究成果) [J], 郭明武;朱勇;向仲怀;蒋同庆;徐继汉5.苎麻多倍体诱导研究——Ⅰ. 苎麻多倍体诱导的基础研究 [J], 董延瑜;郑思乡;洪亚辉;任春梅;刘国民;邓明其;程尧楚
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Abstract:The energy budgets,assimilation and conversion efficiency of l_year-old triploid,(,'F(1,{SOMFe(I gigas
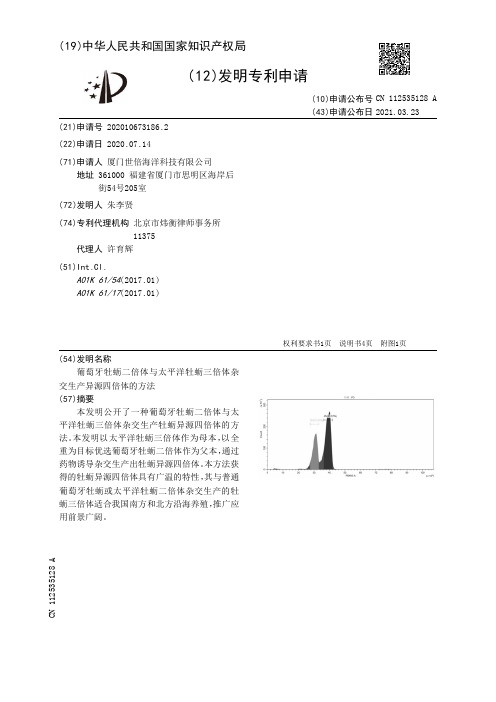
Ⅷcompared with those of sibling diploids.At 5—300c,32%of the energy ingested by diploid oysters WflS
关于三倍体贝类倍性效应研究,现在主要集中于三倍体(3n)与二倍体(2n)生活力和生长特征、性腺 发育、抗病性和软体部组织生化成分等方面的比较。例如,Tabarini(1984)报道了3n海湾扇贝(Argopecten irradituns)的生长和生殖特征;Stanley等(1984)研究了个体多态位点杂合度对美洲牡蛎(Crassostrea “rginica)生长的影响;Mason和Shumway(1988)比较了软壳蛤(Mya aremtria)3n和2n的能量分配特征; Allen和Dowing(1986.1990)、Bruce等(1992)、Akashige和Fushimi(1992)、曾志南等(1999)先后报道了太 甲洋牡蛎3n生长、十殖和绀织生化成分含量季节变化;Hand和Nell等(1993,1994,1998)对悉尼岩牡蛎 (Saccostrea COmID”Fc如如)3n和2n存活、生长、死亡及其养殖潜力进行了比较。关于太平洋牡蛎3n能量收 支特征的研究国内外尚未见报道。本文在实验室条件下比较了太平洋牡蛎3n和2n在不同温度下的生 长预算、能量转化效率和能量收支特征。以揭示三倍体能量利用对策和和能量转化效率的规律,为阐明 其能茸代谢机殚,客观地评估进入现行养殖系统3n群体的生产力特征及其变化趋势.并建立健康的养 殖牛态系和改进养殖工艺提供依据。
同化效率测定:将饵料和收集的粪便分别抽滤到玻璃纤维滤膜,使用前将其在500。c下焚烧6 h,自 称重。用0.5M甲酸铵洗去盐分,并在同样条件下做一空白对照后,65℃烘干48 h,称重,在马福炉(s] 一4—10)内焚烧(5000C,6 h)以测定灰分。根据灰重比例法测定牡蛎的同化效率(Ab)
Ab=1一(Fas}I/Fq。)/(Ea5ll/E。rg∞)
l∞ ** l" ∞1
lⅨl
lW 《m
m
w∞ %m 5∞ Bm
¨1h啪061,P《0 ##铀5 2n%§#&m¥*^2。a自{&%,Ⅺ&自镕№目KI∞g目目¥“*Ⅱ#¥
OOl,‰f∞4*9《o 01),Ⅲ№女#∞97%≈誊%fⅢ自(R。=∞1w.
23n㈣:黜l(4蜒0 =“㈣81p+M 自&l*镕日镕m#《&Ⅻ℃*#T,*i(53 P《0 01j,*#WK{*日∞i#%自.t日**自J目m№i自±*“t日£目镕*Ⅱ#镕ⅢⅨ%。 50±20∞)啦"Ⅱ々2n№i№i十*^《: 蛐m g*+3 66u 23啪+7*U
1材料和方法
1.1实验材料
实验于1999年3—6月进行。实验贝来源于大连市金州区红星养殖场1998年诱导群体的1龄贝。 实验牡蛎先在室内驯化10d,每日投喂小球藻(Cholovella spp.)四次,使培养水体小球藻浓度保持在每毫 升6×104细胞。
1.2实验方法
境培养箱内(MFD一233),每日分别升温或降温0—5。C,直至达到所需实验温度。在各温度下培养一届 后开始各项实验。
牡蛎311和2n的生态生长效率(K1)和组织生长效率(K2)分别如下式计算:KI=P/C x
100%.K2=P/A×100%
2结果和分析
2.1三倍体和_二倍体的生长预算及其它能量组分
牡蛎3n和2n在小同温度下的生长预算(kJ·个。1·d)等能量组分如表1。由表1可见,3n和2n的嘻 长预算均随温度升高而增加。
万方数据
水产学报
24卷
对表1进行方差分析的结果表明,温度和倍性均对牡蛎氨氮排泄率有极显著的影响(R。=40.621, P(0.001;R㈨d=79.48。P<O.001),3n的排泄能量显著高于2n,约为2n的2.06倍。 2.3牡蛎3n和2n的性腺指数
由表3可见,3n牡蛎性腺指数平均是5%,极值4.2%和7.5%;2n平均为25.6%,变幅在19.1%~
虽然r述方差分析未反映出倍性效应对牡蛎呼吸代谢的显著作用,但3n和2n仍表现出一定程度 的差异。由图l和图2可见,2n和3n在干重(70.75±3.86)mg时,生长预算(1(J·ind“-d)与温度在‘定 范同内是直线相关的,但超过20℃则开始降低,尤其在30℃下.2 n生长预算出现负值,这意味着牡蛎在 该体重和温度范围内个体能量需求不足,导致体内贮存能量损耗。在lO~30℃范围内,牡蛎2n和3n的 生长能与呼吸能之比(P/R)均随温度升高而降低,而呼吸能与同化能之比(R/A)则随温度而上升,它们 与温度(t,℃)的关系可分别如下式表达:
耗氧率和氨氮排泄率测定:呼吸实验在5.5L广口瓶中进行,采用Winkler法测定牡蛎的耗氧率。{{ 泄实验利用荼氏试剂法测定牡蛎排入水中的氨氮。
摄食率测定:摄食实验在不同温度下进行。实验时问为8ht饵料浓度为6×104细胞’mL一1 o每隔‘ h测定饵料浓度,并将其凋至实验所要求的浓度。饵料浓度测定应用流式细胞仪进行。
passed.3 65%was ammonia excretion,41 66%Was respired and lost at heat,and 22.81%was used for growth.Under same condition,among the energy fed by triploid,the feces accounted for 23%,the metabolism about 40 65%.ammonia excxeiion about 7 54%,and about 28.78%was usedfor growth In general.tfiploids spent 26%more energy for growth than diploids did,13%more for assimilation,106%more for ammonia exl.、retion and consumed 97%less energy for respiration By means of covedates test,the growth,assimilation, and ammonia excretion energy of oysters were affected significantly by polyploid and temperature.No significant difference was found between diploids and triploids in oxygen consumption.The net growth efficiencies of diploid andtriploid groups were 33 9%and 37.4%respectively,andthe gross growth efficiencies ofthemwere 22 8% and 28.8%.1rd'luence of polyploid and temperature on Kl of oysters Was significant,but the variation of K2 may be attributableto ambienttemperature.Theratio ofgonad dryweight and drytissue oftriploid groupWas 5%Off
505
average.and diploid individuals about 25.6%.
Key words:Crassostrea舒拶triploidy;energy budget;energy conversion efficiency;temperature
=倍体太平洋牡蛎(Crassostrea giga,-)于80年代中期在美围诱导、培育并且养殖成功。在我国,此项 研究近年也取得了很大的进展,叭类三倍体产业化的发展十分迅速,并形成了一定的生产规模。H前, 我国贝类养殖的困境主要是种质资源衰退、动物抗逆能力下降、养殖海区的自身污染和生态环境老化 等。造成这种情况的外在原因是海域养殖过度,适食饵料供应不足,海水污染加剧;内在原因则足贝类 种质资源衰退、动物能量收支失衡、牛态效率低下所致。困此,在我国海区养殖总体环境较差,菜些海区 的环境尚有恶化趋势的大背景下,希望通过三倍体来改良贝类的种质资源,并在一定程度上改善』J!类养 殖产业所面临的问题。
万方数据
㈣g/A 38川∞) 10惦∞+2∞aIⅣ=0 817 F;13
P/P.z2.31 950—6“50E(口=O 968,F_68 21,“0*)
'2am:2R1/8A㈣9+2770—6㈣(m=O%,F_61 5目m《Ⅳ=0 m,F;2l 51 P《0 06) 44.P<0 005)
自l*女目nfm,《*自¥&ⅢⅨ.2nK/A∞m**&#pt'R∞女*日《目kf 3n.
表1牡蛎3n和2n在不同温度和体重下的摄食能和生长预算(kJ/个·d)
The姻州energy Tab.1
and scope of growtlI of扯叫枷and diptoid oysters in differem ttmlperatures alld weights
注:2n f‘晕=(52 5±19 17)rIlg;3n干蘑=(60 17±26 27)ragn