发育生物学 第8章 花
(推荐)植物发育生物学被子植物花器官发育的分子模型

5.边缘滑动模型
边 缘 滑动模型(shifting border model或sliding boundarymodel) (图)解释了B功能基因表达区域的可塑性, 在花发育进程中, B功能基因的表达区域扩展到外层而导 致花瓣状器官的分化, 使外轮器官与内层花瓣在形态上具 有一致性(如单子叶植物百合、郁金香; 基部核心双子叶 植物毛茛、耧斗菜等), 这种B 功能基因功能延伸到外轮 花器官的分子模型又称为修饰的ABC 模型(modified ABC model) , 但此种分子模型并不适用于所有的单子叶植物 类群。
就拟南芥而言, A+E 功能基因控制萼片发育; A+B+E 功 能基因控制花瓣发育; B+C+E功能基因控制雄蕊发育; C+E 功能基因控制雌蕊发育。
3.四分子模型
通过凝胶阻滞、酵母双杂交等分子生物学实验,人们发现 花的同源蛋白能通过聚合作用形成同源或者异源二聚体, 进而组装形成多聚复合体发挥作用。为了解释这些蛋白如 何通过相互作用来调控花器官的发育,Theissen 等结合MADS蛋白多聚体的研究,提出了“四因子”模 型(quartet model),认为花器官是由4 种同源异型蛋白复合体通过结合在目标基因启动子区域来 调节基因开闭,进而调控花器官的发育。
Wild-type:
A功能基因包括拟南芥的APETALA1 (AP1)和AP2 基因、金鱼草 的AP2-like基因LIPLESS1和LIPLESS2(LIP1、LIP2), 单独决
定第1轮萼片属性。
A-function mutant:
B功能基因包括拟南芥的APETALA3 (AP3)和PISTILLATA(PI) 基因、金鱼草的DEFICIENS (D E F) 和GLOBOSA (GLO)基因,
植物发育生物学8

雌蕊原基(PP)分化末期
• 从一般花的分化过程看,很可能最顶端的细胞停止
分裂进而发生PCD是花分化的前提。
• 许多植物在特殊情况下花的顶端可以生出一营养枝,
也可以再分化出一小的花枝或花。这些说明花芽和
花序芽最顶端的分生组织并没有在花器官的分化中
被完全消耗掉。
桂花花顶端又生出一叶(左)或一小花(右)
始。?
2.花序和花分生组织的发育
① 花序和花分生组织的结构
② 拟南芥花序分生组织发育的遗传控制 • 突变体:emf1-2和tfl • EMF基因通过促进营养发育,抑制花序的形成。EMF2蛋 白是一个复合物,核心蛋白包括 CLF 蛋白、 FIE 蛋白和 IRA1蛋白的MULTICOPY SUPPRESSOR组分。EMF1和 EMF2复合物共同作用,抑制开花同源异型基因AG。 • TFL1促进花序分枝;TFL2对控制开花时间的基因(FT)
草原龙胆的花芽分化进程
形态分化前期
萼片原基(CaP)分化期
花瓣原基(PeP)分化期
雄蕊原基(SP)分化期
雌蕊原基(PP)分化初期
雌蕊原基(PP)分化末期
草原龙胆的花芽分化进程
萼片原基(CaP)分化期
花瓣原基(PeP I)分化期
花瓣原基(PeP II)分化期
花瓣原基(PeP III)分化期
雄蕊原基(SP)分化期
2.幼龄向成年转变的遗传调控
① 突变体及其相关基因 • 幼龄延长突变体:玉米的 Tp1-3 、 Cg 、 d1 、 d3 、 d5 、 an1 、 vp8;拟南芥ga1-3、ga4-1、ga5-1。
• 早熟突变体及相关基因:玉米gl15、miRNA172;拟南芥sqn
、 hst、zip、serrate、SPL与miRNA156、TCL1与GL1。
花的形态及发育PPT课件
第11页/共40页
花的类型
双被花
单被花
无被花
两性花
单性花♀/♂ 单性花♀/♂
花萼和花冠合称花被
第12页/共40页
无性花
二、禾本科植物的花
颖片 外稃 内稃 雄蕊 雌蕊 浆片
第13页/共40页
三、花芽的分化
第14页/共40页
第九章 性细胞的形成与有性生 殖过程
9.1雄蕊的发育与结构
一、花丝 二、花药的发育和结构 三、花粉粒的发生、形成和发 育
• 多体雄蕊:一朵花中的雄蕊 的花丝连合成多束。
• 聚药雄蕊:花药合生,花丝 分离。
第7页/共40页
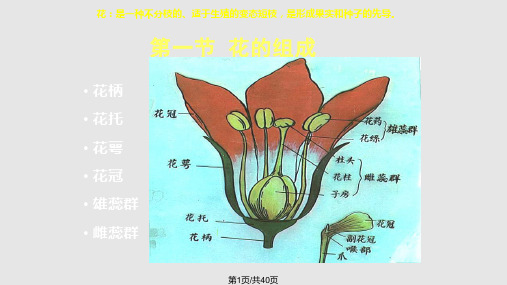
5、雌蕊群 ---雌蕊:柱头、花柱、子房
• 心皮:是组成雌
蕊的基本单位,是 具有生殖作用的变 态叶。
• 背缝线 • 腹缝线
第8页/共40页
A 雌蕊的类型
单雌蕊:一朵花
中只有一个由单 心皮构成的雌蕊。
大孢子
第24页/共40页
成熟胚囊
雌配子体
卵细胞
雌配子
9.2 雌蕊的发育和结构
一 雌蕊的发育
1 柱头 2 花柱 3 子房的结构
背束
腹 缝
线
背
缝
内外表皮
线
子房壁
薄壁细胞 维管束
子房室 胎座 胚珠
腹束
第25页/共40页
二 胚珠的发育和结构、类型
1 胚珠的发育
胎 座 细
平周 分裂
胚 珠 原
胞
基
前端细胞 后端细胞
四面体型
第20页/共40页
三、花粉粒的发生、形成和发育
2 花粉粒的发育与精子的形成 单核靠边期 二 细 胞 期
二 细 胞 花 粉 粒
《发育生物学》课程笔记

《发育生物学》课程笔记第一章:发育生物学的概述一、发育生物学的定义和研究范围1. 定义:发育生物学是生物学的一个分支,它专注于研究生物体从单个细胞(通常是受精卵)开始,经过细胞分裂、分化、形态发生、组织形成和器官发育等过程,最终形成成熟个体的全部生物学过程。
2. 研究范围:- 细胞层面的发育:包括细胞周期、细胞分裂、细胞命运决定、细胞迁移等。
- 分子层面的发育:涉及基因表达调控、信号转导途径、转录因子和网络调控等。
- 形态发生和器官形成:研究生物体的形态变化、轴的形成、器官原基的诱导和分化等。
- 发育过程中的遗传和环境因素:探讨遗传变异、表观遗传学、环境因素如何影响发育过程。
- 发育异常和疾病:研究发育过程中的异常如何导致疾病和畸形。
二、发育生物学的发展历程1. 早期探索(17世纪- 19世纪):- 显微镜的发明使得科学家能够观察胚胎的早期发育。
- 卡尔·冯·林奈(Carl Linnaeus)和卡尔·恩斯特·冯·贝尔(Karl Ernst von Baer)等人的工作奠定了胚胎学的基础。
2. 胚胎学时期(19世纪末- 20世纪初):- 柏拉图生物学假说和重演论(Recapitulation theory)的提出。
- 奥古斯特·魏斯曼(August Weismann)提出了种质论,区分了体细胞和生殖细胞。
3. 细胞和分子生物学时期(20世纪中叶- 至今):- 发现DNA双螺旋结构,开启了分子生物学时代。
- 发育遗传学的发展,如同源框(homeobox)基因的发现。
- 克隆技术的应用,如克隆羊多莉(Dolly)的诞生。
三、发育生物学与其他学科的关系1. 与胚胎学的关系:- 发育生物学是胚胎学的延伸,两者都关注生物体的早期发育,但发育生物学更侧重于分子和细胞机制。
2. 与遗传学的关系:- 遗传学提供了理解发育过程中基因如何传递和表达的基础。
- 发育遗传学领域的研究揭示了基因如何控制发育过程。
八年级生物下册第八单元生物的生殖、发育与遗传第二十一章生物的生殖与发育第二节植物的有性生殖课件苏教版

植物的有性生殖和发育过程示意图
果实和;种子
花药 雄
花丝 蕊 花
花瓣 花萼
柱头 花柱
蕊
雌 蕊
花托
子房
花柄
图片上这些 昆虫在忙碌些 什么呢?
开花后,雄蕊花 药中的花粉通过不同 的途径传送到雌蕊的 柱头上的过程。
自花传粉
异花传粉
传粉的媒介
➢ 风媒: 靠风力传送花粉的传粉方式称风媒。 ➢ 虫媒: 靠昆虫为媒介进行传粉方式的称虫媒,多数
*果树的传粉受到了影响。
靠昆虫传粉的果树由于昆虫的飞行受 到了限制,因此传粉不足;
靠风传粉的果树由于阴雨导致花粉无 法随风传播,也造成减产。
人工授粉
➢农业生产上常采用人工辅助授粉的方法,以克服 因条件不足而使传粉得不到保证的缺陷,以达到预 期的产量。
纵切放大 子房壁
子房
珠被 胚 卵细胞 珠
受精 ——精卵结合
有花植物是依靠昆虫传粉的,常见的传粉昆虫有蜂 类、蝶类、蛾类、蝇类等。 ➢ 鸟媒:其他如借鸟类传粉的称鸟媒,传粉的是一些 小形的蜂鸟,头部有长喙,在摄取花蜜时把花粉传 开。 ➢ 水媒:水生被子植物中的金鱼藻、黑藻、水鳖等都 是借水力来传粉的。
水媒
风媒
图中植物的花是借助于什么媒介传粉的?
鸟媒
虫媒
在果树开花季节,如果遇到阴雨连绵的天 气,常会造成果树减产。这是什么原因?
有性生殖是指由亲代产生生殖细胞, 经过两性生殖细胞的结合,成为受精 卵,发育成新个体的生殖方式。
植物有性生殖的过程
1、当花的各部分成熟的时候,花就要开放,这就是 开;花
2、花粉从花药中散放出来,落到雌蕊柱头上的过程, 就叫 传;粉
第八章植物成花和生殖生理

→ 鳞片状叶; • 茎 : 攀缘→直立
图 8-2 常春藤幼年期和成年期叶形态的变化
图 8-2 植株基部通常是幼年期 , 顶端则是 成年的
• 1.2 缩短幼年期的方法 • (1) 长日照处理:桦树在连续长日照条件
下生长 , 幼年期由 5 ~ 10 年缩短到不足 一年。 • (2) 嫁接:果树、杉木。
• (3) 外施赤霉素 (GAs) ,缩短柏科、杉科 植物的幼年期。
• 第 2 节 春化作用 (vernalization) • 春化作用指低温对植物成花等的促进
作用。
8-3 低温对植物开花的影响
• 把吸胀萌动的种子进行低温处理后春播 , 结果当年抽穗开花。把这一措施称之为 “春化”。后来‘春化’一词扩展到除 种子以外的其它生育期植物对低温的反 应。
ቤተ መጻሕፍቲ ባይዱ
0 0 23 45 6 78 9
图 8-8 春化和 GA 处理对 Scrophylaria vernalis 开花和茎 伸长的差异
3 、一些与春化相关的基因
• Arabidopsis thaliana : FLC—— 迟化 , • FLC —— 无低温时抑制成花 • vrn1,vrn2—— 低温响应基因, fy,fpa,fve,f
℃ 。时间因植物种和品种不同(表 8-1 )而
异。
• 表8-1 冬小麦通过春化需要的温度和天数
类型
春化温度范围(℃)
春化天数
春性 半冬性
冬性
5~15 3~6 0~3
5~8 10~15 40~45
• 对一种植物来说,在可以通过春化的温度 下,温度越低,所需时间越短。
• 对不同类型的植物而言,通过春化作用所 需温度越低的植物,春化作用所需时间也 越长。
发育生物学8—17章 课后习题答案

第八章神经系统发育1、神经胚形成?答:神经胚形成:胚胎由原肠胚预定外胚层细胞形成神经管的过程。
神经胚:正在进行神经管形成的胚胎。
2、初级神经胚形成和次级神经胚形成?答:初级神经胚形成:由脊索中胚层诱导上面覆盖的外胚层细胞分裂,内陷并与表皮质脱离形成中空的神经管。
次级神经胚形成:外胚层细胞下陷进入胚胎形成实心细胞索,接着在细胞索中心产生空洞形成中空的神经管。
3、什么叫神经板,神经褶,神经沟?答:神经板:外胚层中线处细胞形状发生改变,细胞纵向变长加厚,形成神经板。
神经褶:神经板形成后不久,边缘加厚,并向上翘起形成神经褶。
神经沟:神经褶形成后在神经板中央出现的U型沟。
4、无脑畸形和脊髓裂?与哪些基因有关,如何避免?答:无脑畸形和脊髓裂均为人类胚胎的神经管闭合缺陷症。
人的后端神经管区域在27天时如不能合拢,则产生脊髓裂;若前端神经管区域不能合成,则胚儿前脑发育被停止,产生致死的无脑畸形。
它们与pax3、sonic hedghog和openbrain等基因有关。
约50%神经管缺陷可由孕妇补充叶酸加以避免。
5、斑马鱼的神经管如何形成?答:斑马鱼的神经管如何形成:鸟类,哺乳类,两栖类动物胚胎的后端神经管及鱼类的全部神经管形成均采用次级神经胚形成的方式,所以斑马鱼的神经管形成也如此。
6、三个原始脑泡的发育命运?答:前脑发育成为前端的端脑和后面的间脑,端脑最终形成大脑两半球,间脑形成丘脑和下丘脑区域及视觉感受区。
中脑腔最终形成大脑导水管。
菱脑再发育成前面的后脑和后面的髓脑,后脑形成小脑,髓脑形成延髓。
7、菱脑节?答:菱脑节:在神经管闭合后,后脑前后轴逐渐被划分为8节,成为菱脑节,每个菱脑节是一个发育单位,节内细胞可交换而节间不能交换(其是临时性结构,到发育后期逐渐消失,但部分由后脑产生的结构如颜面神经节仍保持分节性结构)。
8、脊髓背腹区域细胞的发育命运?各与哪些因子有关?答:脊髓背部区域依次产生6种中间神经元(dI1-dI6),腹部则形成运动神经元和4种腹侧神经元(V0-V3)。
花的发育——精选推荐

从营养生长到开花的转变发生在茎尖分生组织(Simpson et al. 1999;Parcy 2005)。
花诱导引起根尖分生组织产生花,花由一系列复杂的专门植物组成结构(Zeevaart 1976;伯尼尔1998)。
开花受信号的调节内源性和外源性。
内生信号包括昼夜节律,发育阶段和激素,而外部信号包括白天长度和温度。
有充分的证据表明,花的刺激是从树叶中转移过来的(加纳和阿拉德1920年;埃文斯1971;Yanovsky和凯2002;Searle and Coupland 2004)。
内生和外部信号使植物的生殖发育与外部信号同步环境(图10.1)。
形态学的变化首先发生在转变过程中从植物人到生殖阶段(Lang 1952;魏盖尔1995)。
当生殖发育启动,营养分生组织转化为原花序分生组织,依次产生拉长的花序轴具茎叶和花。
茎叶的腋芽发育分为次级花序分生组织。
花序分生组织不发达生长不稳定。
花的分生组织产生花,它们是由确定的生长形成的。
图10.1拟南芥开花的遗传途径。
箭头表示积极的交互作用和t线负交互作用10.2花器官发育的调控花分生组织产生四种不同类型的花器官,即萼片,花瓣,雄蕊和心皮。
在野生型拟南芥花中,这些器官是四个连续的轮生,第一个轮生有四个萼片,四个花瓣第二轮,第三轮六个雄蕊,第四轮两个心皮。
10.2.1与花发育相关的基因已经确定了三种调节花发育的基因。
这些包括分生组织鉴定基因、花器官鉴定基因和地籍鉴定基因(McSteen等人,1998年)。
分生组织标识和花器官标识基因编码转录因子(Busch et al. 1999)。
这些蛋白质控制的形成花器官。
地籍基因是花器官识别基因的调控因子。
分生组织身份基因作用于未成熟的原基上,形成花分生组织(西蒙等人,1996年)。
金鱼草属植物分生组织鉴定基因的突变(FLO)导致了没有产生的花序的形成说明FLO基因控制了花分生组织的测定的身份。
拟南芥中已鉴定出几种花分生组织同源基因;这些基因抑制了CONSTANS 1 (SOC1)、APETALA1 (AP1)和叶(LFY)。
第八章花2-chenxh

孢原细胞 平周分裂 周缘细胞 造孢细胞
增加珠心的层数
胚囊母细胞
㈡ 胚囊的形成与结构:
以单孢型胚囊(蓼型胚囊)为例:
周缘细胞
形成珠心一部分或退化消失。
(2N)
孢原细胞
(2N)
造孢细胞
(2N)
胚囊母细胞减数分裂四分体3个消失单核胚囊
1个发育
(2N)
(N)
(N)
直接发育
三次有丝分裂 7细胞或8核胚囊
卵细胞1个(N) 助细胞2个(N)
• ⑶压条:选取靠近地面的枝条,将其 压入土中,当产生不定根后,再从母 体上分离。
• ⑷嫁接:选取植物体的枝条或芽,移 接到另一株带根的植物体上,使二者 相互愈合,共同生长的方法。被接的 植物(带根的)称砧木,接上去的枝 条或芽称接穗。
• ①靠接: ②枝接: ③芽接:
嫁接仙人球
嫁接蟹爪兰
嫁接樱花
虫媒花 虫 媒:靠昆虫为媒介的传粉方式。 虫媒花:借虫媒传粉的花。 特点:具 色,香,味。
三、受精
精细胞和卵细胞互相融合的过程。
受精过程
识别反应 花粉萌发 花粉管伸长 释放内容物
(一) 花粉粒在柱头上的萌发
不亲和 不萌发 花粉 传粉 柱头 识别
亲和 吸水、萌发 花粉管
花 粉 粒 的 识 别
(二)花粉管的伸长
中央细胞1个(极核(N)2个或次生核1个)
反足细胞3个到多个(N—多N)
单 核 胚 囊
胚 囊 内 第 一 次 有 丝 分 裂
双 核 胚 囊
胚 囊 内 第 二 次 分 裂
反足细胞 中央细胞
卵细胞 助细胞
3个反足细胞
中央细胞 极核
卵细胞 卵器
助细胞
苏教初中生物七年级下册《8第8章 人的生殖与发育》教案 .doc

第八章人的生殖和发育第一节精卵结合孕育新的生命1、人的生殖系统的组成和功能⑴男性生殖系统由睾丸、输精管、前列腺、阴茎等器官组成。
女性生殖系统由卵巢、输卵管、子宫、阴道等器官组成。
⑵主要生殖器官男性——睾丸(产生精子,分泌雄性激素)女性——卵巢(产生卵子,分泌雌性激素)2、受精过程⑴精子和卵子结合形成受精卵叫做受精作用。
受精卵或精卵结合是新生命诞生的起点。
⑵场所:输卵管⑶结扎输卵管或输精管,能有效阻止生殖细胞精子和卵子的结合,但不会影响人的生殖器官的正常生理功能。
3、胚胎发育⑴受精卵开始细胞分裂的场所——输卵管⑵人的胚胎发育的主要场所——子宫(十月怀胎:280天左右)⑶胎盘的作用:胎儿通过胎盘从母体吸取养料和氧气,并排出二氧化碳等代谢废物。
第二节人的生长发育和青春期1、人的生长发育(起点:受精卵)阶段:婴儿期、幼儿期、童年期、青春期、成年期等。
2、青春期发育的特点⑴青春期身高和体重迅速生长生殖器官的生长发育⑵青春期的性发育第二性征的发育雄性激素和雌性激素男性第二性征:胡须生长,喉结突出,声音变粗,声调较低女性第二性征:乳房增大,声音变细,声调较高。
第三节人体概述1、实验:骨的成分与骨的特性之间的关系①骨的煅烧——烧掉的是有机物,剩下的是硬脆的无机物结论:骨中含有无机物②骨的脱钙——溶解掉的是无机物,剩下的是柔软的能打结的有机物。
结论:骨中含有有机物无机物(钙盐)——使骨脆硬骨的成分有机物(蛋白质)——使骨柔韧骨的特性既有一定的硬度又有一定的弹性2、人的各个系统是在神经系统和内分泌系统调控下,成为一个复杂的、协调、统一的整体。
第九章人的食物来自环境第一节人体需要的主要营养物质1、食物中的营养成分六大营养物质:蛋白质、糖类、脂肪、维生素、无机盐、水第七大营养物质:纤维素蛋白质:细胞的重要组成主要成分糖类:主要供能物质既是人体组成物质脂肪:贮存能量,保持体温,防止机械损伤又是人体供能物质维生素:需求量小,但有重要作用水:构成人体的重要物质(成人体内水60~70%)无机盐:对人体有重要作用2、缺乏症:Vc ——坏血病、牙龈出血V A——夜盲症V D——佝偻病V B1——脚气病缺钙——佝偻病、骨质疏松症缺铁——贫血(多吃含铁和蛋白质的食物)缺碘——地方性甲状腺肿大(多吃含碘盐、紫菜、海带等)第二节人体的消化与吸收1、消化系统的组成消化道:口腔、咽、食道、胃、小肠、大肠、肛门消化腺:唾液腺、肝、肠腺、胃腺、胰腺⑴消化道口腔→牙齿是人体最坚硬的器官(牙冠、牙颈、牙龈)龋齿的原因(吃甜食,不注意口腔卫生)咽→消化和呼吸的共同通道,食道胃→暂时贮存食物,初步消化蛋白质小肠→消化、吸收的主要场所大肠→形成粪便肛门→排出粪便⑵消化腺唾液腺→唾液(唾液淀粉酶)胃腺→胃液(胃蛋白酶)肠腺→肠液液(含多种消化酶)胰腺→胰液(含多种消化酶)肝(最大)→胆汁(不含消化酶),将脂肪乳化为脂肪微粒①细细咀嚼馒头有甜味,因为唾液淀粉酶能促进淀粉分解为麦芽糖。
植物生理学之成花PPT课件

图24.17 烟草的马里兰猛 犸(Maryland Mammoth) 突变体(右)与野生型烟 草(左)的对比。这两个 植株在夏天都是在温室中 生长的。
光周期的发现,使人们 认识到光不但为植物光合作用 提供能量,而且还作为环境信 号调节着植物的发育过程,尤 其是对成花诱导起着重要的作 用。
花熟状态是植物从营养生长转入生殖生长的标志。植物在开花之前,对环境的 反应相当敏感。对开花最有影响的环境因子是日照长度与温度。
花诱导或成花诱导(floral induction) 外界环境条件作为一种信号诱导植物体细胞内发生开花所必需的一系列生理生化
变化,而后开始花芽分化。这一过程成为花诱导。
第3页/共68页
幼年期、温度、日照长短是控制植物开花 的三个重要因素 植物开花的三个阶段
通常将植物的开花过程分为三个阶段: 成花诱导(floral induction) : 接受信号诱导后,特异基因启动,使植物改变发育进程,进入了成花决定态;
成花启动(floral evocation):
指分生组织在形成花原基之前的一系列反应以及分生组织分化成可辨认的 花原基的全过程,也成为花的发端(initiation of flower);
第11页/共68页
8.2.1.3、春化刺激的感受和传递
1.感受低温的时期和部位 时期: 从种子萌发后到苗期都可以(冬性一年生植物) 植物营养体生长的苗期(两年生植物和多年生植物)
部位: 茎尖生长点或正在分生的组织。
拟南芥顶芽区域的营养组 织(A)与生殖组织的纵切 面(B)
2.春化效应的传递
植物完成了春化的感应状态只能随细胞分裂从一个细胞传递到另一个细胞, 传递时应有DNA的复制。还可以通过嫁接传递。如下页图。但在菊花中春化 刺激不能传递。
植物学--生殖器官——花 ppt课件全

ppt课件
27
花萼(calyx)类型
油菜花的离萼
ppt课件
28
花萼(calyx)类型
合萼的萼筒和萼裂片
ppt课件
29
花萼 副萼
一般花萼为一轮,若具两轮,外轮的
花萼叫副萼。
ppt课件
30
花萼(calyx)类型
棉 花 的 副 萼
ppt课件
31
Aconitum sanyoense
萼片大而鲜艳呈花瓣状——瓣状萼。
十字形、蝶形、唇形、管状、舌状、 漏斗状、高脚碟状、钟状、辐(轮)状
ppt课件
44
花冠
ppt课件
45
花冠的类型
ppt课件
46
十字形花冠
ppt课件
47
扁豆(蝶形花冠)
ppt课件
48
丹 参 ( 唇 形 花 )
ppt课件
49
管状花
ppt课件
50
黄鹌菜(舌状花)
ppt课件
51
曼陀罗(漏斗状花冠)
花托类型及子房位置
上位子房下位花
上位子房周位花 半下位子房周位花
上位子房下位花 下位pp子t课房件 上位花
壶状
杯状 壶状
23
花托类型
圆锥形花托
ppt课件
壶状花托
24
环状花托
花盘状花托
ppt课件
25
荷
玉
花
兰
花
花
托
托
桃 花 花 托
ppt课件
26
(二)花萼
概念:花的最外轮变态叶,由若干萼片(sepal)组 成,常呈绿色,不同植物的花萼形态大小、颜色不 同,构成花萼的萼片数目及其相互关系亦不同,是 植物种分类的依据之一,其结构与叶相似。
植物发育生物学第8章ppt课件

者经过低流量红光处理后再萌发,其萌发率极 大降低了。这说明,phyB主要负责调控这些条 件下的种子萌发。
➢ 红光诱导种子萌发,远红光抑制种子萌发 ,表明当phyB处于Pfr形式时,可以促进萌发 。 ➢ 为何某些种子在黑暗条件下也可以萌发, 原因还不清楚。可能是因为在胚胎发生过程中 ,由于phyB的合成和光转换,导致胚胎含有一 些PfrB。
➢ 6.2.1 光诱导的萌发 ➢ 6.2.2 幼苗黄化和光形态建成 ➢ 6.2.3 向光性 ➢ 6.2.4 开花的光周期调控
1.光敏色素与种子萌发
➢ 对拟南芥和其他被子植物来说,光可以促 进种子的萌发。
➢ 光诱导的种子萌发主要受光敏色素A和B的 调控。
野生型和光敏色素突变体的种子萌发率
((abc)) 野用在生低黑型流暗的量下种的浸子红湿即光培使照养在射几暗,天
2. 光形态建成的负调控因子
➢ 光受体下游的信号传导以两种方式诱导光 形态建成:
第一是感受光后,下游的正调控因子可以激 活光合作用的相关基因;
第二是光诱导的信号传导可以抑制光形态建 成的负调控因子。
➢ cop/det/fus突变体:在持续黑暗条件下呈
某种程度的光形态建成表型的拟南芥突变。
➢ 用去黄化(de-etiolated, det)或者持续光形 态建成(constitutively photomorphogenic, cop )来描述这种表型。在筛选紫色幼苗过程中, 还发现了具有cop/det表型的其他突变体,把这
些突变体称为fusca (fus,紫色)。
持续的UV-A/蓝光
黑暗
(d) 在黑暗下,野生型幼苗黄化,光形态建成受到抑制。多效的 cop/det/fus突变体在黑暗下有光形态建成的表型,表明 COP/DET/FUS基因是在黑暗下抑制光形态建成所必需的。
花是器官吗

花是器官吗白书农(✉)北京大学生命科学学院,北京,100871摘要:长期以来,在国内主流教科书和研究论文中,有关花的属性一直存在两种描述方式:一种是“花是变态枝条”;另一种是“花是生殖器官”。
花是一种客观存在,研究者应该对这种客观存在提出一个合理的描述,否则,很难想象在不合理的概念基础上能提出合理的问题,并对研究过程中的发现给出合理的解释。
值得注意的是,在国内主流教科书中为什么会长期存在两种含义如此不同的描述方式呢?这些描述来自哪里?它们各自的合理性在哪里?为找到这些问题的答案,作者对历史上的植物学教科书做了一点考证,发现“花是生殖器官”的描述源于19世纪末20世纪初一本在美国因写作优美轻松而被广泛采用,但在学术上是非主流的教科书。
随着20世纪植物解剖学和发育生物学的发展,早在19世纪末由德国植物学家提出的“花是变态枝条”的概念已经成为当今国际植物科学领域普遍认可的概念。
建议国内学界认真面对这个问题,对目前各学科、领域存在的一些彼此冲突的概念进行认真地梳理,以便我们在描述客观存在的自然现象时能使用尽可能准确的概念,起码不要在教科书中长期存在彼此不兼容的描述方式。
关键词:花,器官,枝条Is A Flower An OrganBAI Shu-nong (✉)College of Life Sciences,Peking University,Beijing 100871,China1问题的提出为了解释为什么要研究“器官形成”这个问题,首先应该给器官下一个定义。
我们实验室主要以雄蕊早期发育调控为研究对象,从形态建成的角度讲,雄蕊自然是一个植物器官。
可是这就带来一个逻辑问题:如果我们将雄蕊定义为器官,那么花是什么?在很多国内的植物学或植物生物学教科书中,都将花定义为生殖器官。
学过植物学的人都知道,花通常由四部分组成:萼片、花瓣、雄蕊和心皮。
显然,从形式逻辑上,我们不可能同时将整体(如花)和整体的一部分(如雄蕊)用同一个概念来定义。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
WUS 激活了 AG, 建立了花器官的径向模式; 然后,AG 抑制了WUS,使得花分生组织成为
有限模式。
(1)C类基因活性模式的建立
多数调控AG 表达的因子(LFY、AP2和LUG )都是
均匀地分布在整个花中。
- lfy突变体的花中AG转录延迟,表明LFY 促进了AG
的表达。
- 在ap2、lug 突变体中,AG在整个花中都有转录
,表明 AP2、LUG 抑制了 AG 在第1和2轮中的表达
。
- LUG 编码一种推测的转录共抑制子,该蛋白可通
起始,由边缘扩展到内部,最终发育成花器
官。
一个分生组织可以产生一个花序,称为花
序分生组织(inflorescence meristem)。它产
生的原基会直接发育成花,也就是原基本身
就是花分生组织。
8.1 花的发育
8.2 花的径向轴
8.3 金鱼草花的近-远轴
花的径向轴,即花萼、花瓣、雄蕊和心皮 着重于拟南芥花的发育。拟南芥的花有4个
。
LFY 转录因子也能结合到AG 基因的这个区域 上,然后WUS 和LFY 共同激活AG 基因在花的中
心表达。
(2)A类基因活性模式的建立
AP2 在所有四轮花器官中都有表达,因此将 AP2限制在第1和2轮中需要其他因子。
有一个候选基因就是花分生组织特性决定基因
AP1 , AP1 最初在整个花分生组织中都有表达,
- SUP 可能以非细胞自主性的方式来抑制第4轮细 胞中AP3 的表达。
B类基因表达模式的建立
(a)LFY/AP1 和 UFO 促进了花分生组织中B类基
因 AP3 和 PI 的转录。 AP3 在第2和第3轮中表达
,反映了UFO的表达区域。还不清楚为何 PI最
初在第2、3和4轮中都有表达。
(b)在分生组织后面的发育中, AP3 和 PI 的表
达通过AP3-PI异源二聚体的作用得以维持。因
为在第4轮中没有异源二聚体,PI的表达在第4
轮中中止。SUP是阻止AP3扩散至第4轮中表达
所必需的。
2、建立A类和C类基因活性的模式
ABC模型的核心是A类和C类基因功能之间的互
相抑制,这点也暗示出A类和C类基因参与彼此活
性模式的建立。
A类和C类基因表达模式的建立
MADS结构域蛋白以多聚体复合体的形式起
作用。
- AP3、PI、AG、SEP 都编码MADS转录因子。
- AP3 和PI 蛋白形成一个异源二聚体,与SEP3 及AG 蛋白相互作用。同样,AG 蛋白可与SEP1 、SEP2 和SEP3 蛋白相互作用。
四聚体模型
8.1.1 ABC 模型
8.1.2 A,B和C活性模式的建立
第八章 花
8.1 花 8.2 花的径向轴 8.3 金鱼草花的近-远轴
花最普遍的形态是辐射对称,其他形态是 花包含四轮花器官,外轮是萼片(花萼)
由辐射对称进化而来的,如左右对称。
,其次是花瓣(花冠),花冠里面有一轮雄
蕊和一个包含一个或多个心皮(雌蕊)的中
心轮。
从营养发育向生殖发育的转化牵涉到茎顶
过与转录因子相互作用来抑制转录。
LFY、AP2 和LUG 在整个花中都有表达。因此, 将AG 基因的表达限制在第3和4轮中必须依靠其他
的沿花径向轴不对称分布的因子?
与在茎顶端分生组织中一样, WUS 在花分生组
织的中心表达,而且WUS 是维持分生组织活性所 必需的。
WUS 编码一种同源异型结构域转录因子,它 能结合到AG 基因第二个内含子中的调控序列上
1、建立B类基因活性的模式
B类基因表达模式的建立
(1)B类基因转录的起始 1)部分是通过花分生组织特性的基因来建立的。
从营养性发育向开花状态转变的核心是在花分
生组织形成处LFY 基因表达的增加,以及随后在发 育中的花分生组织中AP1 的转录。
在 lfy,ap1 突变体的花中 AP3、P1 的转录大
大减少,这表明LFY 和AP1 在花中是AP3、P1
转录所必需的。
所有这些基因都编码转录因子。
LFY 和AP1 只在花发育的早期阶段表达。
LFY 均一地在花分生组织中表达。
AP1 最初在整个花分生组织中都有表达,但
后来局限在第1、2轮中。
B类基因表达模式的建立
2)UFO 基因
ufo 突变体通常缺失花瓣和雄蕊 ,这说明 UFO 是B类基因转录所必需的。
类和B类基因的活性联合决定;雄蕊由B类和 C类基因的活性联合决定;心皮则只由C类
基因来决定。
-
两个假设:
假设A类和C类基因互相抑制对方的活性。 假设C类基因的功能是确定花发育所需要的。
3、ABC 模型成功地预测了所有同源异型转
变
拟南芥花中的同源异型转变(1)
拟南芥花中的同源异型转变(2)
轮的同心模式是如何发育的?
萼片、4个花瓣、6个雄蕊和2个融合的心皮 。
拟南芥花
8.1.1 ABC 模型
8.1.2 A,B和C活性模式的建立
1、花的同源异型突变体(floral homeotic
mutants)
花同源异型突变体:花一个位置上的花器
官为另一个位置上的器官所取代,如花瓣为
雄蕊所取代。
端分生组织,或者腋分生组织。 - 整个分生组织的细胞分裂速率加快,尤其是 中心区以前分裂较慢的细胞。 - 这使得分生组织变得更大,而且经常引起分 生组织形状的改变,变成高度圆顶形,或者 变得更扁,或者二者都有,先圆后扁或先扁 后圆。
一个分生组织可以产生一朵花,称为花分
生组织(floral meristem)。它从花器官原基
2轮种的表达。
(b) AP1 初始在整个花分生组织中转录但是它 的表达受到 AG 的抑制。因此,一旦 AG 开始表
达,AP1的表达就被限制在第1和2轮,在那里
AP1发挥其A类基因的功能。 AG 也会抑制 WUS
的表达。因为WUS是维持分生组织活性所必需
的,这就引发了花发育的有限模式。
(3)WUS 和AG 组成了一种负反馈环
5、SEP 基因
拟南芥SEP 基因功能丧失会影响花瓣、雄蕊
和心皮的发育,形成只包含萼片的嵌套的花。
SEP 基因是B、C类基因活性所必需的,它的
有三个 SEP 基因( SEP1 、 SEP2 和 SEP3 ),它
表达主要限制在花内部的三轮中。
们编码亲缘关系相近的MADS结构域转录因子 。
6、多聚体模型
发育的早期阶段表达,因此,B类基因转录的维持
还需要其他机制?
PI 最初在花分生组织的第2,3和4轮中都有转录
,到后来才局限在第2和3轮中。第4轮中PI沉默的
机制?
AP3 和PI 蛋白相互结合形成一个异源二聚体,
这个异源二聚体是维持B类基因转录所必需的。
因为AP3 不在第4轮中表达,所以,在这个位
AP2 (A类)在包括所有花器官轮的整个植 株中都有表达,这表明 AP2 的A类基因活性
限制在萼片和花瓣部位必须有其他因子的参 与。
A类、B类和C类基因的转录会持续到整个
花基本成熟。在花器官发育晚期丧失A类、B 类或C类基因活性。这表明,ABC表达在整 个器官发育过程中对于维持器官特性都是必 需的。
4、拟南芥中的A、B、C基因
A类基因: AP2 。
B类基因: AP3 和PI 。 C类基因: AG 。
AP3 , PI 和 AG (B、C类)编码MADS结构 域蛋白转录因子家族的成员。 AP2 (A类)
编码一种没有亲缘关系的转录因子。
AP3 和PI (B类)在花瓣轮和雄蕊轮中转 录,AG (C类)在雄蕊轮和心皮轮中转录。
-
瓣代替了雄蕊,第二朵花代替了心皮。
2、ABC 模型内容
根据花同源异型突变体定义了ABC基因。
- A类基因是萼片和花瓣(1轮和2轮)的发育 所必需的。 - B类基因是花瓣和雄蕊(2轮和3轮)的发育 所必需的。 - C类基因是雄蕊和心皮(3轮和4轮)的发育 所必需的。
萼片只由A类基因的活性来决定;花瓣由A
置没有AP3-PI 异源二聚体,PI 在第4轮内的初始
转录就不能维持。
AP3 不在第4轮中表达的维持需要SUP 基因。
- sup 突变体的花分生组织中,AP3 最初在第2和 3轮内表达,但是后来AP3 的表达扩展至第4轮。 - SUP 编码一个推测的转录因子,在第3轮细胞紧
邻第4轮边界的窄带中表达。
UFO 的表达局限在第2和第3轮中。AF3 转录 的区域性模式似乎是由UFO 基因决定的。
UFO
编码一种与蛋白降解调控相关的蛋白,
有一种可能是UFO 的表达破坏了那些对AP3 转
录负调控的蛋白。
(2)B类基因转录的维持
B类基因表达模式的建立
建立PI 和AP3 表达的LFY、AP1 和 UFO 等只在花
但是后来局限在第1和2轮中。
这种限制是由C类基因AG 决定的。 因此,AP1 可能与 AP2 协同作用来决定萼片和
花瓣的特性。有时把AP1认为是一种A类基因。
A类和C类基因表达模式的建立
(a)LFY 和 WUS 促进C类基因AG的表达, AP2 和
LUG 抑制它的表达。一个推测的不对称分布的 抑制因子可能与AP2和LUG作用阻止AG在第1和