最新15蛋白质合成及转运汇总
蛋白质合成及转运

但不能通过真核生物细胞膜 对70S核糖体的敏感性更高
四环素类抗生素对真核细胞核糖体也有抑制
氯霉素——广谱抗生素 与核糖体A位紧密结合,阻碍氨基酰tRNA进入 抑制肽酰转移酶活性,肽链延伸受到影响
50S大亚基蛋白组分
四环素族
氯霉素
链霉素和卡那霉素
嘌呤霉素
放线菌酮
抗生素抑制蛋白质生物合成的原理
eIF-6
促进核蛋白体分离成大小亚基
Met
40S
Met
Met
40S
60S
mRNA
eIF-2B、eIF-3、 eIF-6
①
elF-3
②
GDP+Pi
各种elF释放
elF-5
④
ATP
ADP+Pi
elF4E, elF4G, elF4A, elF4B,PAB
③
真核生物翻译起始复合物形成过程
Met-tRNAiMet-elF-2 -GTP
参与起始过程的蛋白质因子称起始因子(initiation factor,IF)。
肽链合成起始
在原核生物mRNA起始密码AUG上游,存在4~9个富含嘌呤碱的一致性序列,如-AGGAGG-,称为S-D序列。
A
C
B
又称为核蛋白体结合位点(ribosomal binding site,RBS)
能与16S核糖体RNA识别,以帮助从起始AUG处开始翻译。
ORF
每种AA都至少有一种tRNA负责转运 通常一种AA具有几种tRNA
01
tRNA转运活化的AA到模板上
02
反密码子位点
tRNA分子具有4个位点:
3’CCA-OH氨基酸接受位点
蛋白质合成及转运

2.肽链的起始:
三、核糖体
➢核糖体可以看作是一个大分子的机构,它具有许多 精密的配合部分,来挑选并管理参与蛋白质合成的各 个组分。它参与多肽链的启动,延长和终止的各种因 子的识别(许多结合位点)。
➢原核和真核生物均有大、小两个亚基,小亚基能与 mRNA结合形成复合物,再与tRNA结合;大亚基能与 tRNA结合,包括氨酰基位点(A位点)与肽酰基位点(P 位点)。
3、70S起始复合物
4、进位(EF-Ts,EF-Tu,GTP)
5、转肽
延长阶段
6、移位(EF-G,GTP)
7、识别终止密码子
8、水解
终止阶段
9、释放(RF,GTP)
1、30S-mRNA复合物的形成(IF3)
此反应须起始因子3(IF3)使已结束蛋白质合成的核 糖体30S和50S亚基分开。
2、30S预起始复合物(IF1,IF2,GTP)
四、真核生物蛋白质合成的某些特点
几个主要差别: 1、mRNA一般是单顺反子,一般只有一个起始 密码子,一个终止密码子; 2、一般与5’端的第一个AUG就是它的起始密 码子; 3、寻找AUG起始密码子的方式不同,真核靠 cap(帽子结合蛋白)就结合在5’端。
五、蛋白质合成的抑制剂
1、嘌呤霉素——抑制“转肽” 2、氯霉素、四环素、链霉素——抑制原核生 物(pro)翻译,不抑制真核生物(eu)翻译 3、亚胺环己酮——抑制eu,不抑制pro 4、白喉霉素——与EF-2结合,抑制“移位”
这些编码同一种氨基酸的多个密码子 称为同义密码子(synonymous codon)。
2、密码子的连续性
要正确阅读密码必须按一定的密码 框架(reading frame)从一个正确 的起点开始,一个不漏地挨着读下 去,直到碰到终止信号为止。
蛋白质的合成转运知识点整理

蛋白质的合成转运知识点整理●一、蛋白质合成的分子基础●(一)mRNA是蛋白质合成的模板●(1)mRNA以核苷酸序列的方式携带遗传信息,指导合成多肽链中的氨基酸的序列;●(2)每一个氨基酸可通过mRNA上3个核苷酸序列组成的遗传密码来决定,这些密码以连续的方式连接组成读码框架;读码框架之外的序列称作非编码区;●(3)读码框架5'端,是由起始密码AUG开始的,它编码一个蛋氨酸;在读码框架的3'端含有终止密码:UAA、UAG和UGA;●(4)mRNA分子的5'端序列对于起始密码的选择有重要作用,原核生物和真核生物有所差别。
●①原核生物中在mRNA分子起始密码子的上游含有一段特殊的核糖体结合位点序列,使得核糖体能够识别正确的起始密码AUG。
原核生物的mRNA通常是多基因的,分子内的核糖体结合位点使得多个基因可独立地进行读码框架的翻译;●②真核生物mRNA通常只为一条多肽链编码,mRNA5'末端的帽子结构可能对于核糖体进入部位的识别起到一定作用。
翻译的起始通常开始于从核糖体进入部位向下游扫描到的第一个AUG序列。
●(二)tRNA转运活化的氨基酸至mRNA模板上●关键部位:tRNA含有两个关键的部位:氨基酸结合部位,与mRNA的结合部位。
●接头的作用:tRNA在识别mRNA分子上的密码子时,具有接头的作用。
氨基酸一旦与tRNA形成氨酰-tRNA后,进一步的去向由tRNA来决定●(三)核糖体是蛋白质合成的工厂●1.核糖体的活性部位●A位=氨基酰位:结合氨基酰-tRNA●P位=肽酰位:结合肽酰tRNA●E位=出口位:释放已经卸载了氨基酸的tRNA●2.多核糖体●多核糖体是指分离核糖体时得到的若干成串的核糖体。
多核糖体是由一个mRNA分子与一定数目的单个核糖体结合而成的,形似念珠状。
每个核糖体可以独立完成一条肽链的合成,在多核糖体上可以同时进行多条多肽链的合成,提高了翻译的效率●二、蛋白质的生物合成●(一)原料●mRNA作为模板,tRNA作为特异的氨基酸搬运工具,核糖体作为蛋白质合成装配的场所,有关的酶与蛋白质因子参与反应、ATP或GTP提供能量●(二)酶●1.转肽酶●催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离;是一种核酶;肽基转移酶●2.氨酰-tRNA合成酶●催化氨基酸的活化●专一性:对氨基酸有极高的专一性,每种氨基酸都有专一的酶,只作用于L-氨基酸,不作用于D-氨基酸。
15蛋白质合成及转运 共46页
tRNA的关键部位: 氨基酸臂:AA结合部位 反密码环:mRNA结合部位
7
2019/7/11
8
(三) 核糖体是蛋白质合成的工厂 1.核糖体的组成
核糖体:无界膜,颗粒状, 大、小亚基组成
亚基:含不同的Pr、rRNA, 原核和真核生物不同
2019/7/11
9
多聚核糖体(polgsome)
41
信号肽的共同结构 10~40个AA残基组成;三个区 段
1)N端带正电荷的氨基酸:Lys、Arg等碱性AA
2)中间:10~15或更多的疏水AA区段:Leu、Ile等
利于分泌性蛋白进入膜结构
3)C端能被信号肽酶裂解的部位:Ala、Gly、Ser、Cys;
2019/7/1跨1 膜运输完成,信号肽酶识别并切割信号肽
3、转肽酶活性 催化肽键的形成
4、识别mRNA的位点 小亚基上,可容纳2个密码
2019/7/11
14
3、核糖体存在场所 粗面内质网(主要) 细胞质 线粒体、叶绿体
细菌细胞:约20,000个核糖体
真核细胞:106个
未成熟蟾蜍卵细胞:1012个
2019/7/11
15
密码子的特点
2配019对/7/1结1 合
23
形成起始复合体
原核生物
起始因子IF-3、IF-1结合于小亚基
(IF-3:促进上一轮蛋白质合成的核糖
体大小亚基解聚)
小亚基16S rRNA序列与mRNA的SD序列互补配对
使
小亚基结合于SD序列(真核生物无SD序列)
fMet-tRNAfMet与起始密码AUG配对结合,与GTP
亚基聚合:Hb的4个亚基聚合成四级结构
个别氨基酸的化学修饰
蛋白质合成后的加工及转运

The signal-recog整n理it课io件n particle (SRP)
14
③转移通道:存在与内质网膜上的跨膜通道。
④。 SRP受体(SPR receptor),是膜的整合蛋白, 为异二聚体蛋白,存在于内质网上,可与SRP特异结合。
⑤停止转移序列(stop transfer sequence),肽链上的 一段特殊序列,与转移通道蛋白亲合力很高,能阻止肽 链继续进入内质网腔。
第五节 蛋白质合成后的加 工及转运
整理课件
1
本节内容:
一、蛋白质合成后的细胞定位;
二、蛋白质合成后的转运;
三、蛋白质合成后的加工及修饰;
整理课件
2
一、蛋白质合成后的细胞定位:
1、蛋白质是在细胞中游离的核糖体上或者是在糙面内 质网上的核糖体上合成的。
2、蛋白质合成后需要运转到特定的位点起作用:
(1)、内质网驻留蛋白、高尔基体驻留蛋白质、溶酶 体蛋白质、分泌蛋白质、膜蛋白等这些蛋白是由位于 糙面内质网上的核糖体合成的。然后进入内质网腔或 内质网膜。
输入内质网
-Leu-Ala-Leu-Lys-Leu-Ala-Gly-Leu-AspIle-
+H3N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-PheLys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-LeuLeu-
-Ser-Lys-Leu-COO-
整理课件
34
(四)、叶绿体的蛋白质转运
转运到基质的前体蛋白具有典型的N端序列。转运到 叶绿体内膜和类囊体膜的前体蛋白含有两个N端信号序 列,第一个被切除后,暴露出第二个信号序列,将蛋白 导向内膜或类囊体膜。
蛋白质合成、加工和转运的过程

一、蛋白质的合成1、核糖体是合成蛋白质的机器,其功能是按照mRNA的指令由氨基酸合成蛋白质。
2、游离核糖体游离于胞质中,合成细胞内的基础蛋白质;附着核糖体,附着在内质网表面,构成粗面内质网的核糖体,合成分泌蛋白和膜蛋白。
3、蛋白质合成的一般过程:1)氨基酸的活化。
氨基酸和tRNA在氨酰—tRNA合成酶作用下合成活化的氨酰—tRNA。
2)起始、延伸和终止。
3)蛋白质合成后的加工。
肽链N端Met的去除;氨基酸残基的化学修饰,乙酰化、甲基化、磷酸化等;肽链的折叠;二硫键的形成。
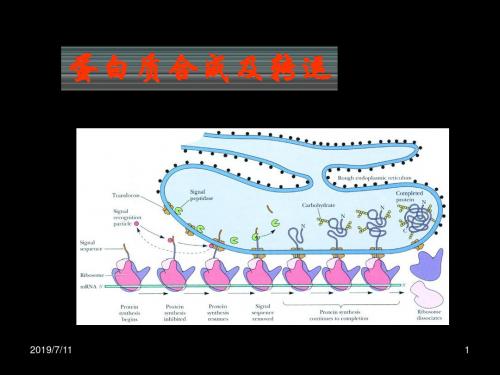
二、蛋白质的分泌合成、加工修饰和转运1、信号肽介导分泌性蛋白在粗面内质网的合成。
1)信号肽是蛋白质合成中最先被翻译出来的一段氨基酸序列,通常由18-30个疏水氨基酸组成,能指引核糖体与内质网结合,并引导合成的多肽链进入内质网腔。
2)新生分泌性蛋白质多肽链在胞质中的游离核糖体上起始合成。
当新生肽链N端的信号肽被翻译后,可立即被细胞质基质中的信号识别颗粒(SRP)识别、结合。
3)与信号肽识别结合的SRP,识别结合内质网膜上的SRP-R,并介导核糖体锚泊附着于内质网膜的通道蛋白移位子上。
而SRP则从信号肽—核糖体复合体上解离,返回细胞质基质中重复上述过程。
4)在信号肽的引导下,合成中的肽链,通过由核糖体大亚基的中央管和移位子蛋白共同形成的通道,穿膜进入内质网网腔。
随之,信号肽序列被内质网膜戗面的信号肽酶且除,新生肽链继续延伸,直至完成而终止。
最后完成肽链合成的核糖体大、小亚基解聚,并从内质网上解离。
2、跨膜驻留蛋白的插入和转移决定了蛋白质的两种去处:1)穿过膜进腔,为可溶性蛋白质,包括分泌蛋白和内质网驻留蛋白。
2)嵌入内质网膜中,形成膜蛋白。
3、粗面内质网与外输性蛋白质的分泌合成、加工修饰和转运过程密切相关。
1)新生多肽链的折叠与装配,与合成同时发生。
内质网为新生多肽链正确的折叠和装配提供了有利的环境。
分子伴侣通过对多肽链的识别结合来协助它们的折叠组装和转运。
《蛋白质的合成与运输》 知识清单

《蛋白质的合成与运输》知识清单一、蛋白质的合成蛋白质是生命活动的主要承担者,其合成过程复杂而精密。
1、遗传信息的转录在细胞核中,以 DNA 的一条链为模板,按照碱基互补配对原则,合成 RNA 的过程称为转录。
这就像是根据一份蓝图制作出一份施工说明书。
DNA 双链解开,碱基暴露,游离的核糖核苷酸与 DNA 模板链上的碱基互补配对,在 RNA 聚合酶的作用下,依次连接形成 RNA 单链。
最终合成的 RNA 包括 mRNA(信使 RNA)、tRNA(转运 RNA)和rRNA(核糖体 RNA)等。
2、遗传信息的翻译mRNA 从细胞核通过核孔进入细胞质,与核糖体结合,开始翻译过程。
翻译过程中,tRNA 起着重要的作用。
tRNA 一端携带特定的氨基酸,另一端有反密码子,能与 mRNA 上的密码子互补配对。
核糖体沿着 mRNA 移动,读取密码子。
一个密码子决定一个特定的氨基酸。
当核糖体读取到起始密码子时,携带相应氨基酸的 tRNA与之结合,随后第二个携带氨基酸的 tRNA 进入,两个氨基酸在核糖体中形成肽键,不断重复这一过程,氨基酸依次连接形成多肽链。
二、蛋白质合成的场所核糖体是蛋白质合成的“工厂”。
核糖体由大、小两个亚基组成。
在真核生物中,核糖体有的游离在细胞质基质中,合成胞内蛋白;有的附着在内质网上,合成分泌蛋白、膜蛋白等。
三、蛋白质的加工与修饰新合成的多肽链往往是没有生物活性的,需要进一步加工和修饰。
1、内质网的加工内质网能对多肽链进行初步加工,比如切除部分氨基酸序列,形成一定的空间结构等。
2、高尔基体的加工高尔基体对来自内质网的蛋白质进行进一步的加工、分类和包装。
3、其他修饰蛋白质还可能会进行磷酸化、甲基化、乙酰化等化学修饰,从而改变其活性、稳定性或功能。
四、蛋白质的运输蛋白质合成并加工完成后,需要被运输到特定的部位发挥作用。
1、胞内运输在细胞内,一些蛋白质通过细胞质中的微管、微丝等细胞骨架进行运输。
蛋白质的合成、转运、修饰

蛋白质的合成蛋白质的种类是由基因决定的,也就是说人类基因组有多少个基因,人体就有多少种蛋白质,只是蛋白质表达的时期和部位不同.根据人类基因组计划分析得知:全部人类基因组约有2.91Gbp,约有39000多个基因;也就是说人体蛋白质的种类有39000多种蛋白质生物合成可分为五个阶段,氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰一.氨基酸的活化分散在胞液中的各种氨基酸需经特异的氨基酰-tRNA合成酶催化,ATP供能,并需Mg2+或Mn2+参与在氨基酸的羧基上进行活化,生成中间复合物()后者再与相应的tRNA作用,将氨基酰转移到tRNA分子的氨基酸臂上,即3′末端腺苷酸中核糖的3′(或2′)羟基以酯键相结合形成氨基酰-tRNA【氨基酰tRNA的生成】tRNA各种tRNA的一级结构互不相同,但它们的二级结构都呈三叶草形三叶草形结构的主要特征是:含有四个螺旋区、三个环和一个附加叉四个螺旋区构成四个臂,其中含有3′末端的螺旋区称为氨基酸臂,因为此臂的3′-末端都是C-C-A-OH序列,可与氨基酸连接三个环分别用Ⅰ、Ⅱ、Ⅲ表示环Ⅰ含有5,6二氢尿嘧啶,称为二氢尿嘧啶环(DHU环)环Ⅱ顶端含有由三个碱基组成的反密码子,称为反密码子环;反密码子可识别mRNA分子上的密码子,在蛋白质生物合成中起重要的翻译作用环Ⅲ含有胸苷(T)、假尿苷(ψ)、胞苷(C),称为假尿嘧啶环(TψC环);此环可能与结合核糖体有关tRNA在二级结构的基础上进一步折叠成为倒“L”字母形的三级结构起始因子原核起始因子只有三种(IF1、IF2、IF3)真核起始因子(简称为eIF)种类多且复杂,已鉴定的真核起始因子共有12种延长因子原核生物(简称EF)由三部分组成:EF-Tu,EF-Ts,和EF-GEF-Tu它介导氨酰-tRNA进入核糖体的空位EF-Ts充当EF-Tu亚基的鸟嘌呤核苷酸交换因子,催化EF-Tu释放GDPEF-G催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来真核生物(简称eEF)真核生物中分为:eEF-1和eEF-2eEF-1有两个亚基,α和βγα相当于原核生物中的EF-Tu亚基,它介导氨酰-tRNA进入核糖体的空位Βγ相当于原核生物中EF-Ts,核苷酸交换因子α,催化GDP从α上释放eEF-2相当于原核生物的EF-G,催化tRNA的移位和多肽延伸的每个循环后期mRNA从核糖体上掉下来终止因子(释放因子)原核生物细胞的释放因子(简称RF):识别终止密码子引起完整的肽链和核糖体从mRNA 上释放的蛋白质释放因子1(RF1):能识别终止密码子UAA和UAG而终止蛋白质合成的细菌释放因子释放因子2(RF2):能识别终止密码子UAA和UGA而终止蛋白质合成的细菌释放因子释放因子3(RF3):与延长因子EF-G有关的细菌蛋白质合成终止因子当它终止蛋白质合成时,它使得因子RF1和RF2从核糖体上释放真核生物细胞只有一种终止因子(称为eRF)能识别所有的终止密码子因为它没有与GTP结合的位点,所以它不能帮助完成合成的多肽从P位点的tRNA的释放在真核生物内可能还存在能与eRF合作、帮组多肽从核糖体释放的蛋白质核糖体的活性部位单个核糖体上存在四个活性部位,在蛋白质合成中各有专一的识别作用1.A部位:氨基酸部位或受位:主要在大亚基上,是接受氨酰基-tRNA的部位2.P部位:肽基部位或供位:主要在小亚基上,是释放tRNA的部位3.肽基转移酶部位(肽合成酶),简称T因子:位于大亚基上,催化氨基酸间形成肽键,使肽链延长4.GTP酶部位:即转位酶(EF-G),简称G因子,对GTP具有活性,催化肽键从供体部位→受体部位核糖体上还有许多与起始因子、延长因子、释放因子以及各种酶相结合的位点核糖体的大小是以沉降系数S来表示,S数值越大、颗粒越大、分子量越大原核细胞与真核细胞核糖体的大小亚基是不同的二.核糖体循环(肽链合成)1.肽链启动阶段在蛋白质生物合成的启动阶段,核蛋白体的大、小亚基,mRNA与一种具有启动作用的氨基酸tRNA共同构成启动复合体。
蛋白质的生物合成及转运

第十二章蛋白质的生物合成及转运蛋白质的生物合成在细胞代谢中占有十分重要的地位。
目前已经完全清楚,贮存遗传信息的DNA并不是蛋白质合成的直接模板,DNA上的遗传信息需要通过转录传递给mRNA。
mRNA才是蛋白质合成的直接模板。
mRNA是由4种核苷酸构成的多核苷酸,而蛋白质是由20种左右的氨基酸构成的多肽,它们之间遗传信息的传递与从一种语言翻译成另一种语言时的情形相似。
所以人们称以mRNA为模板合成蛋白质的过程为翻译或转译(translation)。
翻译的过程十分复杂,几乎涉及到细胞内所有种类的RNA和几十种蛋白质因子。
蛋白质合成的场所是核糖体,合成的原料是氨基酸,反应所需能量由A TP和GTP提供。
蛋白质合成的早期研究工作都是用大肠杆菌的无细胞体系进行的,所以对大肠杆菌的蛋白质合成机理了解最多。
真核细胞蛋白质合成的机理与大肠杆菌的有许多相似之处。
第一节遗传密码任何一种天然多肽都有其特定的严格的氨基酸序列。
有机界拥有1010~1011种不同的蛋白质,构成数目这么庞大的不同的多肽的单体却只有20种氨基酸。
氨基酸在多肽中的不同排列次序是蛋白质多样性的基础。
目前已经清楚,多肽上氨基酸的排列次序最终是由DNA上核苷酸的排列次序决定的,而直接决定多肽上氨基酸次序的却是mRNA。
不论是DNA还是mRNA,基本上都由4种核苷酸构成。
这4种核苷酸如何编制成遗传密码,遗传密码又如何被翻译成20种氨基酸组成的多肽,这就是蛋白质生物合成中的遗传密码的翻译问题。
一、密码单位用数学方法推算,如果mRNA分子中的一种碱基编码一种氨基酸,那么4种碱基只能决定4种氨基酸,而蛋白质分子中的氨基酸有20种,所以显然是不行的。
如果由mRNA 分子中每2个相邻的碱基编码一种氨基酸,也只能编码42=16种氨基酸,仍然不够。
如果采用每3个相邻的碱基为一个氨基酸编码,则43=64,可以满足20种氨基酸编码的需要。
所以这种编码方式的可能性最大。
应用生物化学和遗传学的研究技术,已经充分证明了是293三个碱基编码一个氨基酸。
十五章蛋白质的合成转运和加工

起始和延伸Met-tRNAs的区别
⑴两种tRNA本身有差别; ⑵氨基酸的状态不同。
Formyl Met
AA AC AC AA CA GC
A A A
A
Met A C C A GC GC
G
A CAU
A* CAU
3 0 S -m R N A 复 合 体
IF -1 IF -3 核 糖 体
30S
IF -2
GTP FM et
fM et
fM et
IF -1 IF -2 IF -3
G D P Pi
起始反应的产物
70S核 糖 体
图 15- 原 核 生 物 翻 译 起 始 复 合 体 的 装 配
Only fMet –tRNAf enters partial P site on 30S subunit bound to mRNA
三真核生物蛋白质合成的起始
因子
eIF-3 eIF-4F(CBP-Ⅱ) eIF-4E(CBP-Ⅰ) eIF-1 eIF-4B eIF-4A eIF-6 起 eIF-5 始 eIF-4C eIF-2α eIF-2β eIF-2γ eIF-1A eIF-3A
表15-3 真核生物蛋白质合成中涉及的各种辅助因子
原核生物翻译起始因子
表 15-2 E.coli蛋 白 质 合 成 起 始 所 需 的 三 种 起 始 因 子
因 子质量因 子 /核功 能
( KDa)糖 体
IF3 23
25% 亚 基 解 离 与 mRNA的 结 合
IF2 97.3 ?
起 始 tRNA的 结 合 与 GTP水 解
第五节蛋白质合成后的加工及转运课件

在蛋白质合成过程中,信号识别颗粒与新生蛋白质结合,引导蛋白 质向内质网定位。
作用
确保新生蛋白质正确地转运到内质网,进行进一步的加工和修饰。
跨膜运
跨膜运输
是指蛋白质通过生物膜的运输过 程,包括通过细胞膜、线粒体膜 、叶绿体膜等。
运输方式
包括主动运输和被动运输,其中 主动运输需要消耗能量,而被动 运输则不需要。
多肽链在核糖体上合成的同时,通过 信号肽的引导,进入内质网腔或跨膜 转运至高尔基体、溶酶体和细胞骨架 。
03
膜泡运输
通过形成囊泡的方式,将已经折叠好 的蛋白质从一个膜结构转运到另一个 膜结构。例如,从内质网到高尔基体 ,或从高尔基体到溶酶体。
PART 04
蛋白质合成后的加工和转 运的调节
蛋白质合成后的加工的调节
蛋白质的乙酰化
在蛋白质合成后,某些赖氨酸残基可被乙酰化,从而调节 蛋白质的稳定性。这一过程由乙酰转移酶催化。
蛋白质转运的调节
01
核孔复合体的调节
核孔复合体是细胞核膜上的转运孔道,可选择性地将蛋白质从细胞质转
运到细胞核内或从细胞核转运到细胞质。核孔复合体的转运活性受到多
种因素的调节。
02
囊泡转运的调节
2023-2026
ONE
KEEP VIEW
蛋白质合成后的加工 及转运课件
REPORTING
CATALOGUE
目 录
• 蛋白质合成后的加工 • 蛋白质的转运 • 蛋白质分选的信号和途径 • 蛋白质合成后的加工和转运的调节 • 蛋白质合成后的加工及转运异常与疾病的关系
PART 01
蛋白质合成后的加工
蛋白质二硫键的形成
在蛋白质合成后,某些氨基酸残基需要经过氧化形成二硫 键,以稳定蛋白质的高级结构。这一过程由特异的二硫键 异构酶催化。
《蛋白质的合成与运输》 知识清单

《蛋白质的合成与运输》知识清单一、蛋白质的合成(一)遗传信息的转录1、定义转录是指以 DNA 的一条链为模板,按照碱基互补配对原则,合成RNA 的过程。
2、场所主要在细胞核中进行。
3、过程(1)解旋:DNA 双螺旋解开,碱基暴露。
(2)配对:游离的核糖核苷酸与 DNA 模板链上的碱基互补配对。
(3)连接:在 RNA 聚合酶的作用下,相邻的核糖核苷酸连接形成RNA 链。
(4)释放:合成的 RNA 从 DNA 链上释放,DNA 双螺旋恢复。
4、产物包括 mRNA(信使 RNA)、tRNA(转运 RNA)和 rRNA(核糖体RNA)。
(二)遗传信息的翻译1、定义翻译是指以 mRNA 为模板,合成具有一定氨基酸顺序的蛋白质的过程。
2、场所细胞质中的核糖体。
3、密码子(1)概念:mRNA 上决定一个氨基酸的三个相邻碱基。
(2)特点:简并性(一种氨基酸可以由多种密码子决定);通用性(几乎所有生物共用一套密码子)。
4、反密码子(1)概念:tRNA 一端与密码子互补配对的三个碱基。
(2)作用:识别密码子,转运相应的氨基酸。
5、过程(1)起始:核糖体与 mRNA 结合,起始密码子位于核糖体的 P 位。
(2)进位:tRNA 携带相应的氨基酸进入核糖体的 A 位。
(3)脱水缩合:在核糖体中,相邻氨基酸形成肽键。
(4)移位:核糖体沿着 mRNA 移动,原在 A 位的 tRNA 离开核糖体,新的 tRNA 携带氨基酸进入 A 位。
(5)终止:当核糖体遇到终止密码子,翻译停止,多肽链释放。
二、蛋白质的运输(一)内质网中的运输1、内质网的功能内质网是由膜围成的管状、泡状或扁平囊状结构,具有加工蛋白质和合成脂质的功能。
2、内质网中的运输过程新合成的多肽链进入内质网腔,进行折叠、修饰等加工。
内质网通过出芽形成囊泡,将加工后的蛋白质运输到高尔基体。
(二)高尔基体中的运输1、高尔基体的结构和功能高尔基体由扁平膜囊和大小不等的囊泡组成,主要对来自内质网的蛋白质进行加工、分类和包装。
《蛋白质的合成与运输》 知识清单

《蛋白质的合成与运输》知识清单一、蛋白质的合成蛋白质是生命活动的主要承担者,其合成过程是一个极其复杂而精细的过程。
首先,我们来了解一下蛋白质合成的场所——核糖体。
核糖体就像是一个“蛋白质生产工厂”,由大小两个亚基组成。
在真核细胞中,核糖体可以分为游离核糖体和附着核糖体。
游离核糖体主要合成细胞自身所需的蛋白质,而附着核糖体则与内质网结合,合成一些分泌蛋白和膜蛋白。
蛋白质合成的“蓝图”是信使 RNA(mRNA)。
DNA 中的遗传信息通过转录形成 mRNA,mRNA 从细胞核中出来,进入细胞质,与核糖体结合。
在核糖体上,还有转运 RNA(tRNA)的参与。
tRNA 像是一个个“搬运工”,它们一端携带特定的氨基酸,另一端有反密码子,可以与mRNA 上的密码子互补配对。
蛋白质合成的过程大致可以分为三个阶段:起始、延伸和终止。
起始阶段,核糖体小亚基先与 mRNA 结合,然后再结合起始 tRNA,形成起始复合物。
接着,核糖体大亚基结合上来,蛋白质合成正式开始。
延伸阶段,按照 mRNA 上的密码子顺序,携带相应氨基酸的 tRNA 依次与核糖体结合,在核糖体的催化下,氨基酸之间形成肽键,逐渐延长肽链。
终止阶段,当核糖体遇到 mRNA 上的终止密码子时,蛋白质合成停止,释放出合成好的多肽链。
二、蛋白质的加工与修饰新合成的多肽链通常没有生物活性,需要经过一系列的加工和修饰才能成为具有特定功能的成熟蛋白质。
在内质网中,多肽链会进行折叠和初步修饰。
例如,一些蛋白质会进行糖基化修饰,即在蛋白质上添加糖链。
高尔基体也参与蛋白质的加工和修饰。
在这里,蛋白质可能会进一步糖基化,或者进行磷酸化、甲基化等修饰,还会被分选和包装,运送到细胞的不同部位。
此外,一些蛋白质还会在细胞质或其他细胞器中进行加工,比如某些蛋白酶会切除多肽链的一部分,使其成为有活性的蛋白质。
三、蛋白质的运输蛋白质合成和加工完成后,需要被运输到特定的部位发挥作用。
分泌蛋白通过内质网形成的囊泡运输到高尔基体,经过进一步加工后,再由高尔基体形成的囊泡运输到细胞膜,通过胞吐的方式分泌到细胞外。
蛋白质合成及转运-生科

延伸因子识别tRNAMet
蛋白质合成及转运-生科
● 原核生物中的第一个蛋氨酸要进行甲酰化 修饰---甲酰Met:
fMet - tRNAiMet
蛋白质合成及转运-生科
5、翻译起始于mRNA与核糖体的结合:
● 真核生物mRNA分子的5’端有核糖体进入部位: 帽子结构帮助识别mRNA分子与核糖体的结合位点. 核糖体沿着 mRNA分子5’ → 3’扫描至起始密码AUG.
蛋白质合成及转运-生科
①IF1、IF3与30S小亚基结合:
IF3防止30S亚基与50S 亚基过早结合.
mRNA与小亚基结合
蛋白质合成及转运-生科
② fMet–tRNAiMet进入
蛋白质合成及转运-生科
③50S大亚基的结合
蛋白质合成及转运-生科
A:新进来的氨基酸结合位点. P:肽链结合位点. E:出口(大部分在大亚基上).
无法释放,抑制70S核糖体的解离.
蛋白质合成及转运-生科
四环素(土霉素、金霉素) ①作用于细菌30S小亚基,抑制起始复合物形成; ②抑制氨酰tRNA进入核糖体A位,阻滞肽链延伸; ③影响终止因子与核糖体的结合 四环素类抗生素对真核细胞核糖体也有抑制. 但不能通过真核生物细胞膜. 对70S核糖体的敏感性更高.
蛋白质合成及转运-生科
● 蛋白质的加工修饰:
1、肽链末端的修饰:N-端fMet或Met的切除 2、信号序列的切除. 3、二硫键的形成. 4、部分肽段的切除. 5、个别氨基酸的修饰. 6、糖基侧链的添加. 7、辅基的加入.
蛋白质合成及转运-生科
●胰岛素原的加工:
间插序列(C肽区)
HS SH
HS SH HS
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
真核生物核糖体:4种rRNA、约82个Pr,Mw
420万
原核生物 真核生物
小 rRNA 16S-rRNA 18S-rRNA
亚
基 蛋白质
21种
33种
rRNA 5S-rRNA 5S-rRNA
大
23S-rRNA 28S-rRNA
亚
5.8S-rRNA
基 蛋白质
34种
49种
2020/8/5
11
• 原核生物5S rRNA可与tRNA互补,与23S rRNA 互补
➢ GTP水解释放能量,促使大亚基结合成完
整的核糖体,IF-1、IF-2、IF-3脱离复合体
➢ m202R0/N8/5A、fMet-tRNAfMet和核糖体构成三元起始复
原核生物能将tRNAiMet的氨基酸甲酰化
2020/8/5
20
1)转运Met的tRNA:2 种
(1)tRNAiMet(原核: tRNAifMet): 被起始因子识别,与起始密码AUG配对,在肽链N端
掺入Met,启动蛋白质合成
(2)tRNAMet:被延伸因子识别,与mRNA之间AUG配 对,掺入Met(延伸)
2)Met的活化产 物
(1)起始位置:Met-
2020/8/5
tRNAiMet
原核:fMet-tRNAMet
(2)中间位置:Met-
tRNAMet
21
2、翻译开始于mRNA与核糖体的结 合
原核生物的mRNA存在富含嘌呤碱基的SD序列
核糖体小亚基上的16S rRNA3’端的富含嘧啶序列可与SD
序列
2配020对/8/5结合
3、转肽酶活性 催化肽键的形成
4、识别mRNA的位点 小亚基上,可容纳2个所 粗面内质网(主要) 细胞质 线粒体、叶绿体
细菌细胞:约20,000个核糖体
真核细胞:106个
未成熟蟾蜍卵细胞:1012个
2020/8/5
14
密码子的特点
4种碱基,编码20个氨基酸
2020/8/5
18
2、每个氨酰-tRNA合成酶可识别一个特定的AA和 与此AA对应的多个tRNA的特定部位
氨酰-tRNA合成酶具有校对功能,如果产物不对应, 则启动校对活性,水解非正确组合的氨基酸和 tRNA之间形成的共价联系。
2020/8/5
19
(二)起始阶段
1、所有蛋白质翻译起始为甲硫氨酸 一个特殊的tRNA启动了蛋白质的合成
翻译(蛋白质合成)起始的必须步骤——确定阅读框架
按照不重叠的三联体密码子翻译产生对应的AA并形成肽键
终止密码子,合成结束,肽链释放(终止密码连续出现2~3个)
UAA、UGA、
AUG 5’
UAG
3’
5’端非编码区
编码区
3’端非编码区
2020/8/5
3
mRNA:蛋白质合成的 模板
mRNA分子中每三个相邻碱基组成一个密码子 真核生物mRNA的成熟需要经过剪切修饰,只 编码一条肽链,转录和翻译发生在不同的空间和 时间 原核生物mRNA转录和翻译几乎同时进行,可 以编码多条肽链
一种tRNA只与一种AA结 合 一种AA可与几种tRNA结 合 2t02R0/N8/5A约50余种
tRNA的关键部位: 氨基酸臂:AA结合部位 反密码环:mRNA结合部位
6
2020/8/5
7
(三) 核糖体是蛋白质合成的工厂 1.核糖体的组成
核糖体:无界膜,颗粒状, 大、小亚基组成
亚基:含不同的Pr、rRNA, 原核和真核生物不同
15蛋白质合成及转运
蛋白质的生物合成:
以mRNA为模板合成蛋白质的过程
将mRNA中核苷酸顺序转变为蛋白质分子中氨基酸顺序的过 程,即将mRNA中4种核苷酸的语言解读为蛋白质中20种氨 基酸的语言——翻译(Translation)
2020/8/5
2
一、蛋白质合成的分子体系
(一) mRNA:蛋白质合成的模 板 辨认起始密码子(AUG):
2020/8/5
8
多聚核糖体(polgsome)
mRNA和多个核蛋白体的聚合物,一般间隔40个核苷酸结合 一个核糖体。
(一个mRNA分子同时有多个核蛋白体在进行蛋白质的合成)
2020/8/5
9
多聚核糖体的电镜照片
2020/8/5
原核生物的 转录翻译同 步进行
10
rRNA与蛋白质构成核糖 体
细菌核糖体:3种rRNA、57个Pr,Mw 270万
2020/8/5
15
二、翻译的步骤
5个阶段 合成方向:N→C端 翻译方向:5’→3’
2020/8/5
16
2020/8/5
在核糖体上进行, 且是一个循环过程, 因此也成为核糖体
循环。
17
(一)氨基酸的活化 1、氨基酰-tRNA合成酶:使AA结合到特定的 tRNA上
AA活化(能量) tRNA携带AA到mRNA指定部位(专一 性)
• 连续性:两个密码之间没有任何碱基隔开。 • 简并性:64个密码子对应20个氨基酸,同一种氨基酸有
两个或多个密码子的现象为密码子的简并性。 • 变偶性:专一性取决于头两个碱基,tRNA上的反密码子
与mRNA密码子匹配时,第一二碱基严格配对,第三碱基 可以变动。反密码子第一位可为I,可识别更多简并密码 子。这样32种tRNA即可识别61个编码氨基酸的密码子。 • 通用性:共用一套遗传密码。线粒体编码方式有所不同。
22
形成起始复合体
原核生物
➢ 起始因子IF-3、IF-1结合于小亚基
(IF-3:促进上一轮蛋白质合成的核糖
体大小亚基解聚)
➢ 小亚基16S rRNA序列与mRNA的SD序列互补配对
使
小亚基结合于SD序列(真核生物无SD序列)
➢ fMet-tRNAfMet与起始密码AUG配对结合,与GTP
结合
的起始因子IF-2同时结合,形成五元起始复合物
2020/8/5
4
(二)tRNA:转运活化的AA至mRNA 模板
1、引导氨基酸进入核糖体 2、将mRNA的碱基序列翻译为氨基酸序列 3、tRNA的反密码子与mRNA的密码子碱基互补
2020/8/5
5
70~90个核苷酸,分子较小 三叶草形结构 1)含稀有碱基 2)反密码环有三个反密码子 3)氨基酸臂有CCA结构
• 16S rRNA的3’端ACCUCCUUA与mRNA的SD 序列互补,翻译起始定位;与23S rRNA互补,大 小亚基结合
• 23S rRNA与起始tRNA互补
2020/8/5
12
2、核糖体的结构与功能
1、P位点(肽酰基位点) 结合肽基-tRNA的位点
2、A位点(氨酰基位点) 结合氨基酰-tRNA的位点