SOX_DMRT性别决定基因家族及其应用研究进展_李楠
【国家自然科学基金】_性别决定_基金支持热词逐年推荐_【万方软件创新助手】_20140731
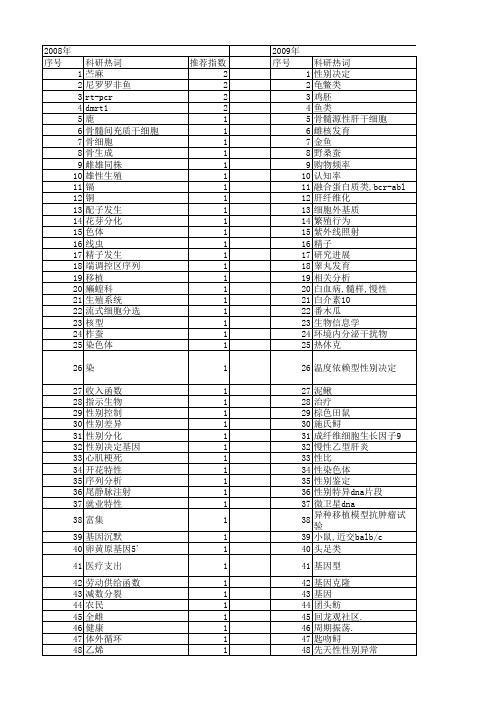
科研热词 苎麻 尼罗罗非鱼 rt-pcr dmrt1 鹿 骨髓间充质干细胞 骨细胞 骨生成 雌雄同株 雄性生殖 镉 铜 配子发生 花芽分化 色体 线虫 精子发生 端调控区序列 移植 癞蝗科 生殖系统 流式细胞分选 核型 柞蚕 染色体 染 收入函数 指示生物 性别控制 性别差异 性别分化 性别决定基因 心肌梗死 开花特性 序列分析 尾静脉注射 就业特性 富集 基因沉默 卵黄原基因5' 医疗支出 劳动供给函数 减数分裂 农民 全雌 健康 体外循环 乙烯 中国 东亚飞蝗 x、y精子 sry
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105
科研热词 推荐指数 性别决定 8 性别分化 3 雌核发育 2 转性 2 温度 2 性激素 2 微卫星 2 基因表达 2 sox基因 2 rt-pcr 2 鸡 1 鱼类 1 高敏c-反应蛋白 1 非混交雌体 1 间质干细胞 1 野生中国大鲵 1 重组抑制 1 重复序列 1 遗传因素 1 适应意义 1 输尿管梗阻 1 赌博任务 1 表观遗传学 1 血便量 1 董事会多元化 1 萼花臂尾轮虫(brachionus calyciflorus) 1 芦笋 1 肥胖 1 肝细胞生长因子 1 编码区序列 1 结果评价 1 结合分析 1 组织表达 1 简并pcr 1 稀有鮈鲫 1 稀有(鱼句)鲫 1 社会因素 1 生育 1 生殖细胞 1 甜瓜 1 环境因素 1 猫栉首蚤指名亚种 1 牛科 1 牛 1 爬行动物 1 滞育激素 1 滞育发生 1 温度依赖型性别决定 1 沼泽型水牛 1 河流型水牛 1 气候变化 1 植绥螨 1
第三章控制动物性别

9
10
11
实用文档
动物的性别
生理性别。在遗传性别控制下通过个体发育的生化过程而 形成的。
性腺性别。生理性别的基础取决于原始性器官的类型,该 基础性别称为性腺性别,再进一步分化为雌雄异体和雌雄 同体。
外部性别。鱼类的雌雄异形由第一和第二性征决定。第一 性征是直接与繁殖活动有关的性征,即性附属器官,如软 骨鱼类雄鱼的鳍脚、鳉科鱼雄鱼的生殖足、鰕虎鱼雄鱼的 臀突和鳑鲏鱼雌鱼的产卵管等。第二性征是与繁殖活动无 直接关系的性征,与性腺发育和分泌活动有关,如雄鱼的 婚姻装及珠星等。
实用文档
芳香化酶基因 在性激素产生过程中,有一系列的酶参与
实用文档
温度因素对鱼类性别决定分化的影响作用 常常大于其遗传因素。因而鱼类的性别趋向可 塑性是哺乳类和鸟类无法比拟的。由于自然水 体中温度因素变化是有规律的,和长期演化的 鱼类性别遗传因素相结,使温度对其性别影响 作用常常破忽视。养殖鱼类的经济价值常因为 其性别则不同差异很大,如乌苏里鱼拟雄鱼生 长快而味美,养殖价值远高于雌性;鲑科鱼和 鲟鱼的雌性由于生长较快且能生产鱼子酱,其 价值远大于雄性个体。
③XO/ XX 与ZO/ ZZ 型。除形态的差异外, 性染色体 也可能存在数目的差异, 即2 个性别相差1个性染色体, 一般而言, 在XO/ XX 型中,XX 型为雌性, 而XO型为雄 性, 即雄性缺少Y 染色体, 灯笼鱼科中的几种鱼类属于 此类型。在ZO/ ZZ 型中,ZZ 型为雄性,ZO 型为雌性, 即雌性缺少W 染色体。鲱形目的短颌鲚为此类型。
究竟是髓质部得以发育还是皮质部得以 发育,既取决于遗传因素,也受环境因素影 响。
实用文档
温度对鱼性别的影响
江黄颡鱼的性分化是属于 温度依赖型性别决定。
区块链系统中的分布式数据管理技术——挑战与展望

第44卷 第1期2021年1月计 算 机 学 报CHINESEJOURNALOFCOMPUTERSVol.44No.1Jan.2021收稿日期:2019 03 22;在线发布日期:2019 10 31.本课题得到国家重点研发计划项目(2018YFB1003404)、国家自然科学基金(U1811261,61672142)、辽宁省科学技术基金(20180550321)资助.于 戈,博士,教授,中国计算机学会(CCF)会员,主要研究领域为分布式数据库、分布与并行计算、区块链.E mail:yuge@mail.neu.edu.cn.聂铁铮(通信作者),博士,副教授,中国计算机学会(CCF)会员,主要研究方向为数据库、数据集成、区块链.E mail:nietiezheng@mail.neu.edu.cn.李晓华,博士,讲师,中国计算机学会(CCF)会员,主要研究方向为信息安全、区块链.张岩峰,博士,教授,中国计算机学会(CCF)高级会员,主要研究领域为分布式数据处理、云计算.申德荣,博士,教授,中国计算机学会(CCF)高级会员,主要研究领域为分布式数据库、数据集成.鲍玉斌,博士,教授,中国计算机学会(CCF)高级会员,主要研究领域为数据仓库、OLAP.区块链系统中的分布式数据管理技术———挑战与展望于 戈 聂铁铮 李晓华 张岩峰 申德荣 鲍玉斌(东北大学计算机科学与工程学院 沈阳 110169)摘 要 区块链是在数字加密货币的应用基础之上发展起来的一种分布式数据库技术.区块链系统具有去中心化、不可篡改、分布共识、可溯源和最终一致性等特点,这使其可以用于解决不可信环境下数据管理问题.区块链独特的数据管理功能已经成为各领域应用中发挥区块链价值的关键.本文基于对比特币、以太坊、超级账本等代表性区块链系统的研究分析,阐述区块链系统中分布式数据管理技术.首先,深入讨论区块链系统与传统分布式数据库系统之间的异同点,从分布式部署模式、节点角色、链拓扑结构等多个方面给出区块链的分类.然后,详细分析各类区块链系统所使用的数据存储结构、分布式查询处理与优化技术及其优缺点.最后,总结区块链系统的分布式数据管理技术在各专门领域应用中所面临的挑战和发展趋势.关键词 区块链;分布式数据管理;数据存储;查询处理中图法分类号TP311 犇犗犐号10.11897/SP.J.1016.2021.00028犜犺犲犆犺犪犾犾犲狀犵犲犪狀犱犘狉狅狊狆犲犮狋狅犳犇犻狊狋狉犻犫狌狋犲犱犇犪狋犪犕犪狀犪犵犲犿犲狀狋犜犲犮犺狀犻狇狌犲狊犻狀犅犾狅犮犽犮犺犪犻狀犛狔狊狋犲犿狊YUGe NIETie Zheng LIXiao Hua ZHANGYan Feng SHENDe Rong BAOYu Bin(犛犮犺狅狅犾狅犳犆狅犿狆狌狋犲狉犛犮犻犲狀犮犲犪狀犱犈狀犵犻狀犲犲狉犻狀犵,犖狅狉狋犺犲犪狊狋犲狉狀犝狀犻狏犲狉狊犻狋狔,犛犺犲狀狔犪狀犵 110169)犃犫狊狋狉犪犮狋 Blockchainisatechniqueofdistributeddatabasewhichisdevelopedwiththeapplicationsofdigitalencryptedcurrency.Ablockchainsystemhasthecharacteristicsofdecentralization,non tampering,distributedconsensus,provenanceandeventualconsistency,whichmakesitbeappliedtosolvedatamanagementproblemsoftheuntrustedenvironments.Thedatamanagementfunctionofablockchainsystemhasalreadybecometheimportantfeatureforplayingitsvalueintheapplicationsofdifferentdomains.Blockchainsystemsmakeeverynodecontainacompletecopyofledgerdata,andusedistributedconsensusalgorithmstoensuretheconsistencyofdata.Therefore,ablockchainsystemisanewkindofdistributeddatamanagementsystemscomparedwithtraditionaldistributeddatabasesystems.WithanalyzingtherepresentativeblockchainsystemsincludingBitcoin,EthereumandHyperledgerFabric,thispaperfocusesonthedistributeddatamanagementtechniquesinexistingblockchainsystems,whichcoversqueryprocessing,smartcontract,networkcommunication,anddatastoragelayersinthearchitectureofblockchainsystems.Thispaperfirstdiscussesthemaindifferencesandsimilaritiesbetweenablockchainsystemandatraditionaldistributeddatabasesystem.Justlikeadistributeddatabasesystem,ablockchainsystemhasfeaturesofdistribution,transparency,autonomyandscalabilityonmanagingdata,butitisalsodifferentfrommostofdistributeddatabasesystemsontopologic,datadistribution,queryprocessing,consistencyandsecuritymechanism.Then,thispaperpresentstheclassificationofblockchainsystemsondifferentaspectsofdistributeddeploymentstyles,noderolesandtopologicalstructures.Withthedevelopmentofblockchaintechnology,blockchainsystemsaredesignedtoadaptblockchainapplicationenvironments.Themodelsofpublicblockchain,consortiumblockchainandprivateblockchainareproposed,andfunctionsofblockchainareregroupedanddeployedtomakenodesplaydifferentrolesinasystem.Moreover,themultipletopologicstructuresofblockchainareproposed.Besidesthechainstructureoftraditionalblockchain,theDAGstructures,suchasTangleandLattice,areappliedtoimprovetheefficiencyofblockchainsystems.Thirdly,thispaperanalyzesthetechniquesofdistributeddatastoragemanagement,distributedqueryprocessingandoptimizationusedinblockchainsystemsanddiscussestheiradvantagesanddisadvantages.Specifically,thedatastoragetechniquesofexistingblockchainsystemsaredeeplyanalyzed,includingthedatastructuresofstorage,aswellastheorganizationofdatafilesandoptimizationtechniques.Key ValuedatabasessuchasLevelDBareusuallyusedinblockchainsystemstoimprovetheefficiencyofaccessingledgerdataandstatedata.Currently,moreresearchworksfocusonusingdifferentmethods,includingdatabase,indexanddistributedstorage,tooptimizethestorageofblockchain.Thispaperalsoanalyzesvariousqueriesintheexistingblockchainsystemsandclassifiesthemintothreetypes:accountquery,transactionqueryandcontractquery.Thedistributedqueryprocessingtechniquesusedinblockchainsystemsarediscussed.Fourthly,thispaperpointsoutthechallengesanddevelopmenttrendsofdistributeddatamanagementtechniquesforblockchainsystems,includingdistributedstorageforblockchaindata,efficientandsecureconsensusmechanismforblockchaintransactions,highavailablequeryprocessing,distributedmanagementofsmartcontracts,privacyprotectionforblockchaindata,dataauditandmonitoringinblockchainsystem.Finally,thispapershowsthedistributeddatamanagementofblockchainsystemsinvariousdomain specificapplications,suchasfinance,manufacture,networkstorage,creditandotherfields.犓犲狔狑狅狉犱狊 blockchain;distributeddatamanagement;datastorage;queryprocessing1 引 言在“互联网+”应用日益普及的大环境下,大量应用需要将发生的事件、行为、状态持久地记录在分布式环境中以用于日后的查询,即进行分布式记账.分布式记账已经逐渐成为互联网应用中的一项重要功能.以电子商务交易系统为例,客户需要执行提交订单、通过电子银行向商家支付货款、从物流公司收货等操作,商家需要执行接受订单、通过物流公司发货、通过电子银行收款等操作,电子银行方需要执行从客户收款、向商家付款等操作,物流公司需要执行从商家收货、收取物流款、向客户发货等操作.客户、商家、物流公司、电子银行共四方处于一个分布式环境中,在每一个环节都需要记录相关的操作和信息.由于各方之间并不存在完全信任关系,最终以哪一方记录的账目为确认信息是一个重要的问题.传统的方法采用由电子商务交易服务平台作为公正的第三方进行统一记账,所有的交易信息的查询操作全部在这个平台上进行处理,物流公司和银行的部分数据也以接入的方式添加至交易服务平台.在这种传统集中式记账方式里,主要的交易信息存储在单一的记账方,这是一种“逻辑”上的集中式存储模式,即交易数据存储在唯一的某业务参与方并由其负责管理.集中式记账方式存在的问题包括:(1)记账方为了保证可靠性需要存储数据的多个副本,从而造成了数据存储的性能瓶颈;(2)交易数据可能被记账方篡改且无法验证,因此各参与方需要完全信任记账方;(3)记账方受到攻击后数据难以恢复.因此,传统集中式记账方式存在着存储效率低、可信性差、易受攻击等弊端.为了解决以上难题,采用分布式记账方式的比921期于 戈等:区块链系统中的分布式数据管理技术———挑战与展望特币系统(Bitcoin)[1]在2008年被首次提出,并受到广泛关注.随后,区块链技术作为比特币系统所采用的底层技术逐渐引起工业界与学术界的重视,比特币系统所具有的分布共享性、共识性、不可篡改性、可溯源性和最终一致性等特点均来源于区块链技术.在基于区块链技术的分布式记账方式中,所有参与方都可以保存一份相同的完全账本,新加入的参与方可以下载完全账本并验证账本的正确性.这种方式降低了传统集中式记账方式中记账方的多副本数据维护成本,同时参与方也可以通过访问本地数据提高访问效率.此外,在区块链系统中,交易的账目采用数字签名和加密算法处理,从而提高了系统中数据的安全性,而区块之间通过哈希值串联的数据关联方式和基于共识算法确认区块的数据写入机制也使得区块链上的数据极难被篡改.起初,区块链技术所支撑的比特币系统仅是一个专用的交易系统,并不支持虚拟货币交易以外的其他功能,这严重限制了区块链技术在分布式数据管理上的应用.随着区块链技术的发展,产生了大量新型区块链系统.2014年由Buterin基于区块链技术推出了以太坊(Etherum)平台[2].以太坊提供了基于智能合约的编程功能,支持区块链应用的二次开发,这标志着区块链2.0时代的诞生.超级账本(HyperledgerFabric)[3]则是基于IBM早期贡献出的OpenBlockchain为主体搭建而成的Linux基金会的区块链项目,其主要目的是发展跨行业的商用区块链平台技术.在超级账本框架中,包括了HyperledgerFabric①、HyperledgerBurrow②、HyperledgerSawtooth③和HyperledgerIroha等多个项目,构成了完整的生态环境.区块链3.0时代[4]则是将区块链技术的应用范围扩展到各类应用之中,服务领域除金融、经济之外,还包括政府、健康、科学、文化等领域.区块链技术将支持各类资产交易与登记的去中心化可信处理,并与物联网等技术融合.未来,区块链技术将会与其他新兴技术相结合用于各类应用之中,诸如区块链+科学、区块链+医疗、区块链+教育、区块链+能源等应用将会迅速发展.目前,区块链技术已应用于多个领域之中.在数字货币服务领域,支持支付、兑换、汇款、交易功能;在金融服务领域,支持清算、结算、安全监管、反洗钱等功能;在B2C服务领域,支持无人管理的商亭等新业务;在P2P租赁管理领域,支持无需中介的货物交换、租赁等共享经济新业务;在供应链管理领域,支持物理资产签名、物流跟踪和交付等功能;在知识产权保护领域,用于建立不可篡改的权利和拥有权;在征信管理领域,支持身份认证、日志审计和监管等;在溯源管理领域,支持数据鉴别与存证、防伪溯源等功能.区块链技术是一种建立在多种技术之上的分布式共享账本技术,而区块链本质上是一种多方参与共同维护的分布式数据库.相对于集中式数据库管理系统,区块链系统采用去中心化或者弱中心化的数据管理模式,没有中心节点,所有参与节点均可以存储数据,而事务的持久性则依靠参与节点共同维护的不断增长的数据链和非集中式的共识机制予以实现,保证了数据在基于验证基础上的可信性.此外,相比于传统的分布式数据库和分布式数据存储系统,区块链系统的参与节点可以获得完整的数据副本,而非部分数据的副本.区块链系统的特殊数据存储机制和一致性共识机制是其不同于传统分布式数据库系统的主要原因.区块链的数据存储结构和数据组织方式不同于其他数据存储系统.区块链将数据记录组织成区块(Block),并在每个区块的区块头中通过记录前一区块的哈希值将区块组织成链式结构.这种结构使区块链的数据存储具有不易篡改性、可溯源性和可验证性.然而,区块链的存储结构和基于密码学算法的共识机制也为数据管理带来了交易确认效率低和查询不便等诸多弊端.例如在记录交易的吞吐量方面,使用区块链技术的比特币系统仅支持每秒处理7笔交易数,并且还需要经过1小时以上时间才可以确认写到区块(相关研究表明43%的比特币交易未能在一小时内得到处理④).此外,区块链的数据记录按时间顺序存储在区块中,这为交易数据的查询处理带来了挑战,当前很多数字货币系统的查询处理都要依赖于某种键值数据库系统.其次,区块链的共识机制也不同于分布式数据库系统.区块链系统为了在P2P网络环境下保证交易操作符合事务特性,需要维护数据一致性,并避免“双重支付”(DoubleSpends)的发生,这是区块链共03计 算 机 学 报2021年①②③④HyperledgerFabric.https://www.hyperledger.org/projects/fabricHyperledgerBurrow.https://www.hyperledger.org/projects/hyperledger burrowHyperledgerSawtooth.https://www.hyperledger.org/projects/sawtoothStudy:43%ofBitcoinTransactionsAren’tProcessedafterFirstHour.2017.https://www.ccn.com/43 bitcoin transactions not processed one hour study says识机制的主要考虑的问题.同时,由于区块链网络本身是一个去中心化的网络,参与节点完全自治,并没有统一的节点负责管理和维护,为此区块链节点之间需要使用P2P技术实现数据广播以更新节点的状态信息和账本信息.区块链系统公认的基础架构模型[5]主要分为6层,本文在其基础上增加了查询层,以便对区块链系统的查询处理机制进行分析.这样,区块链系统架构扩展为7层,如图1所示,主要包括:(1)应用层.基于区块链的各类应用,如数字货币、区块链金融、区块链征信等;(2)查询层.实现对交易账本数据的访问和验证,以及对账号状态的查询;(3)合约层.由脚本、算法机制和智能合约所构成的可编程基础框架;(4)激励层.负责为奖励记帐工作而进行货币发行、交易费用分配任务;(5)共识层.封装网络节点的PoW、PoS、DPoS和PBFT等各类共识算法,实现分布式共识机制;(6)网络层.封装P2P组网机制,数据传播机制和数据验证机制;(7)数据层.封装底层数据区块的数据结构和加密机制.当前的区块链系统大多基于该系统架构进行实现,其中数据层、网络层、共识层和查询层是区块链系统的必要元素.现有相关工作针对区块链系统不同层次的技术和区块链在各领域上的应用进行了大量研究与综述.对于区块链系统所包含的关键技术和研究现状,以及未来的发展趋势,袁勇等人[5]在区块链的基础架构模型方面对比特币的原理和技术进行了系统的阐述,何蒲等人[6]结合比特币系统介绍了区块链的概念和技术,并对前景进行了展望,邵奇峰等人[7]对比特币、以太坊和超级账本等多个区块链平台进行分析,总结了区块链的优势、劣势和发展趋势.在应用层方面,文献[8]对区块链在数字货币上的应用进行了全面的综述,刘敖迪等人[9]介绍了区块链技术在信息安全领域的研究现状和进展.由于区块链具有健壮的数据存储能力,因此相关研究工作在数据存储系统上进行区块链技术的应用[10 11].对于合约层,贺海武等人[12]结合多个领域应用场景对智能合约技术的概念、关键技术和面临的问题进行了阐述.此外,对于共识层、网络层和数据层,已有研究分别对区块链系统的共识机制[13 14]、安全机制[15 16]、网络协议[17]、可信数据管理[18]和查询处理[19]进行了整理和综述.区块链在设计之初就是以进行防篡改的数据存储和管理为目的,分布式数据管理是区块链系统的主要功能之一.区块链技术中涉及分布式数据管理的部分主要集中在区块链架构的查询层、合约层、网络层和数据层,其中查询层和合约层在区块链系统中负责实现对数据的处理操作,如图1所示.本文主要以分布式数据管理为视角,基于对当前主流的区块链系统分析,对比不同区块链系统在数据管理上的差异,对其中分布式数据管理所涉及的数据存储技术、查询处理机制和算法进行阐述和分析,并对区块链研究中涉及分布式数据管理的挑战进行探讨,对各领域的应用进行展望.本文第2节对区块链系统的分布式数据管理机制进行分析,对比区块链系统和传统分布式数据管131期于 戈等:区块链系统中的分布式数据管理技术———挑战与展望理系统的异同;第3节介绍区块链系统的分类;第4节介绍区块链系统中的数据存储技术,包括物理存储结构,对比不同区块链系统在物理存储机制上的差异,以及区块链系统所采用的数据存储优化技术;第5节介绍区块链系统的数据查询处理技术;第6节探讨区块链系统在分布式数据管理方面所面临的研究挑战和发展方向;第7节展望区块链所支持领域应用的场景和待解决的问题;第8节总结全文.2 区块链系统的分布式数据管理区块链系统作为一种分布式数据库管理系统,主要以解决数字货币的货币转移、兑换和支付功能而被提出.区块链的特征主要体现在数据的公开透明、不可篡改和网络结构的去中心化等几个方面.由于区块链主要面向的是不可信数据存储环境下的记账应用,因此在数据存储上采用了去中心化、全副本的分布式方式,即所有参与方均通过P2P网络结构连接,并可以存储完整的共享账本.由此可见,区块链系统在管理交易记账上虽然使用了分布式数据管理方式,但与传统的集中式数据管理和分布式数据库系统管理数据的方式均有所差别.本节主要将区块链系统与传统数据管理方式进行对比和分析,并阐述彼此间的共同点和差异性.图2 记账业务流程对比2 1 区块链与传统分布式数据库的共同点区块链技术主要是针对现有金融机构的集中式记账系统的信任问题而被提出的,其本身是由分布式存储、P2P网络、加密算法、共识机制等多种技术所构成的.中本聪基于区块链技术设计并发行了数字货币“比特币”,用以解决美国次贷危机中所展现的金融机构信任问题.相比于金融机构的集中式记账系统,基于区块链技术的交易记账系统具有公开透明、去中心化、可溯源查询和不可篡改等诸多的优势,从而避免了集中式记账方式中账本的真实性高度依赖于对记账方信任的弊端.这里以电子商务的交易记账应用为例,对传统基于清算中心的集中式记账方式和基于区块链的分布式记账方式的记账业务流程进行对比.传统集中式记账方式如图2(a)所示,交易相关的账目数据集中存储在清算中心的数据库中,交易的参与各方如果需要调用完整的交易信息需要访问清算中心,其弊端主要体现在完全依赖于对清算中心记账方的信任,一旦记账方失信或遭受攻击,其保存的数据也随之失去可信性.区块链的分布式记账方式如图2(b)所示,其中账本数据是整体共享的,以区块为单位通过密码学算法链接在一起,且网络中任何一个参与方均可以存储完整的共享账本副本,而数据的安全性则也是基于密码学算法予以保证.由于所有参与方均保存有共识后的共享账本,因此任何一个参与方进行双重支付或篡改账本数据的难度变得极大,从而保证账本数据在不可信环境中的可信性.区块链系统的分布式记账方式使其在数据存储管理的方式上与分布式数据库相同,即存储结构化的数据集合,这些数据逻辑上属于同一系统,物理上分布在计算机网络的各个不同场地上[17].区块链系统同样具有分布式数据库所具有的诸多特性:(1)分布性区块链系统与分布式数据库系统在数据的存储方面都是物理上分散、逻辑上统一的系统.区块链系统中具有全局统一的数据模式,数据以副本形式存储在参与节点中,每个参与节点存储的是数据模式相同且数据一致的共享账本.(2)透明性区块链系统在数据访问上具有透明性,用户看到的共享账本是全局数据模型的描述,就如同使用集中式数据库一样,在记录交易数据时也不需要考虑共享账本的存储场地和操作的执行场地.在数据复制方面,区块链系统的共享账本存储在各个参与节点上,并通过共识机制自动维护数据的一致性.23计 算 机 学 报2021年(3)自治性区块链系统的参与节点具有高度的自治性.在通信方面,参与节点可以独立地决定如何与其他参与者进行通信;在查询方面,参与节点本地就保存了完整的共享账本,可以在本地执行对账本数据的访问.(4)可伸缩性区块链系统支持参与节点规模的任意扩展.区块链系统允许参与节点在任意时刻加入和退出系统.而且,由于区块链的参与节点保存的是完整共享账本,因此对于参与节点重新加入区块链系统后,仅需要从其他节点更新缺失的区块数据即可完成数据的重新分布,不会影响整体的系统性能.2 2 区块链与传统分布式数据库的差异区块链系统原始的设计目的之一是解决非信任环境下数据的可信性问题.所谓的非信任环境是指负责数据存储的节点可能随意篡改数据而其他参与节点又无法识别,这将造成参与节点之间的互不信任问题.对于传统分布式数据库管理系统而言,系统建立在信任环境,其中参与节点采用统一管理的方式,节点之间具备完全相互信任的关系.因此区块链与传统的分布式数据库在数据管理方式上又具有显著的差异,如图3所示,具体体现在以下几个方面:(1)去中心化拓扑结构在参与节点的网络拓扑结构方面,区块链系统的去中心化结构采用了基于P2P的分布式模式,这种结构与基于P2P网络结构[20]的数据库系统(P2PDBS)[21 22]相似.如图3(b)所示,区块链节点通过通信控制器(CM)仅基于邻居地址进行通信,其加入和退出都是随意和动态的.传统分布式数据库虽然数据分布在不同的场地,但是通常采用中心化的主从结构,由全局的网络管理层存储各个局部数据库节点的地址和局部数据的模式信息,以用于查询处理时进行全局优化和调度,如图3(a)所示.(2)数据分布方式分布式数据管理的数据存储方式,通常分为两类[23]:①分割式.数据被划分成若干个不相交的分片,分别保存在不同的节点上,数据的划分方法分为水平分片和垂直分片;②复制式.同一个数据分片保存在一个以上的节点上,复制方式分为部分复制和全复制.分割式能够节省数据的存储空间,查询时需要在节点间传输数据,虽然使用半连接等算法可进行优化,但效率依然较低.复制式通过多节点的数据冗余存储可提高查询效率,但耗费存储空间且需要维护数据一致性.区块链系统的数据分布采用的是全复制式,即每个参与节点都在本地复制了具有全局模式的全部数据.因此,数据在区块链系统中是全局共享的,如图3(b)所示.相比于区块链系统,传统分布式数据库的分布方式主要基于在全局模式创建局部模式,再对数据进行垂直分片和水平分片,如图3(a)所示,每个节点存储的是全局数据分片的副本,再通过数据分片的元信息管理实现全局数据的访问和查询处理.当前很多基于分布式数据库技术的大数据存储系统,如HBase①等,均采用集中式的元信息管理节点管理数据副本的分布信息.图3 区块链系统与传统分布式数据库系统对比(3)数据查询处理区块链系统中对账本信息的查询处理通常在存331期于 戈等:区块链系统中的分布式数据管理技术———挑战与展望①ApacheHBaseReferenceGuide.http://hbase.apache.org/book.html储了完整共享数据的参与节点本地执行.由于区块数据采用基于文件的存储方式且本身缺少索引结构,因此在区块链上直接执行对账本查询只能使用顺序扫描的方式访问所有区块数据.目前区块链系统常用的查询优化方式是将账本记录存储在Key Value数据库中,以提高数据的访问效率.当前,比特币和以太坊等系统都使用了LevelDB①存储和检索数据.需要说明的是,在以太坊这类支持智能合约的第二代区块链系统中,智能合约代码的执行处理是嵌入在区块链记账功能中的.因此,对智能合约代码的调用是在所有参与进行共识验证的节点上执行.传统分布式数据库的查询处理主要基于数据副本的大小和分布场地进行优化[24],而在面向大数据的分布式数据库上则采用基于并行计算思想的查询优化方法[25].(4)数据一致性维护数据一致性是保证数据正确性和可信性的关键,区块链系统采用共识机制来保证各节点上数据的一致性.在数字货币的应用中通常采用工作量证明机制(PoW)通过算力竞争保证分布式的一致性[26],如解决基于SHA256、Ethash②等算法的数学难题,而从节约能耗的角度,则会采用权益证明机制(Proof of Stake,PoS)和授权权益证明机制(DelegatedProof of Stake,DPoS)等③方法.其中,使用工作量证明机制进行一致性维护的最大问题在于共识的效率过低,一个区块的一致性需要在其后生成一定长度的后续区块之后才能够被确认.分布式数据库系统通常采用包括实用拜占庭容错PBFT[27]、Paxos[28]、Gossip[29]、RAFT[30]等高效的算法维护数据的一致性,而这些算法也被一些面向联盟链应用的区块链系统所采用.(5)数据安全性机制区块链系统在安全性方面主要为用户提供了数据篡改验证、数据溯源和加密安全机制.数据的篡改可以通过校验前后区块的哈希值进行验证,因此要篡改数据并被所有参与者认可就需要在算力上付出高昂代价以重新生成区块,其难度相比传统的集中式和分布式数据库都要大很多.但是在数据的可访问性上,由于区块链的共享性,所有用户均可访问完整数据,而传统数据库管理系统则基于用户身份验证方式控制数据的访问.为了解决共享数据上的隐私安全性问题,区块链采用了基于非对称加密的交易方式实现匿名交易,其优点是很好地保护了用户隐私,缺点是一旦密钥丢失,用户的账号信息将无法恢复.综上所述,区块链系统相比传统分布式数据库系统,在记账方式上提供了更好的分布性、透明性和可信性,在功能上提供了防篡改验证机制和智能合约机制,因此更加适合在非可信环境下的匿名使用.另一方面,相比传统的分布式数据库系统,区块链系统在网络结构、数据存储和访问方式上也具有显著的差异.3 区块链系统的分类3 1 区块链系统部署方式的分类区块链系统根据其分布式部署方式和开放对象被划分为三种:“公有链”(PublicBlockchain)、“联盟链”(ConsortiumBlockchain)和“私有链”(PrivateBlockchain).三类区块链系统的对比如表1所示.表1 各区块链系统类型对比公有链联盟链私有链网络结构完全去中心化部分去中心化(多)可信中心节点规模无控制可控有限加入机制随时可以参加特定群体或有限第三方机构内部节点记账方任意参与节点预选节点机构内部节点数据读取任意读取受限读取受限读取共识机制容错性高、交易效率低(PoW或PoS等)容错性和交易效率适中(PBFT,RAFT)容错性低、交易效率高(Paxos,RAFT)激励机制有代币激励无代币激励无代币激励代码开放完全开源部分开源或定向开源不开源(1)公有链公有链是对所有人开放的,任何互联网用户都能够随时加入并任意读取数据,能够发送交易和参与区块的共识过程.比特币和以太坊等虚拟货币系统就是典型的公有链系统.公有链是完全去中心化的结构,其共识机制主要采用PoW、PoS或DPoS等方式,将经济奖励和加密算法验证相结合,以保证经济奖励和共识过程贡献成正比.此外,公有链中程序开发者对系统的代码是完全开源的,而且开发者无权干涉用户.在分布式数据管理方面,公有链系统的优势和缺陷主要包括以下几个方面:43计 算 机 学 报2021年①②③LevelDB.http://leveldb.org/RayJ.Ethash.https://github.com/ethereum/wiki/wiki/EthashBitshares.DelegatedProofofStake.http://docs.bits hares.org/bitshares/dpos.html。
动物的生殖器官与性别特征

要点二
包皮
覆盖在阴茎头部的皮肤,保护阴茎头免受外界刺激和损伤 。
雄性特有特征
雄性体型
通常比雌性更大、更强壮,有利于在求偶和竞争中占 据优势。
雄性毛色
部分动物雄性具有鲜艳的毛色或独特的标记,以吸引 雌性和宣示领地。
雄性行为
雄性动物在求偶、交配和育儿等方面表现出独特的行 为特征,如鸣叫、展示和争斗等。
雄性动物生殖器官,呈椭球形或长条形, 表面光滑,内部有大量的生精小管。
卵巢
雌性动物生殖器官,呈椭球形或不规则形 ,表面有卵泡和黄体等结构。
输精管与输卵管
分别为雄性和雌性动物的生殖管道,负责 输送精子和卵子。
阴茎与阴道
分别为雄性和雌性动物的交配器官,具有 特定的形态和结构以适应交配需求。
02
雄性动物生殖器官及特征
VS
性别展示
动物通过展示性别特征来吸引异性,如雄 性动物的鲜艳羽毛、强壮体魄等。性别展 示有助于增加动物的交配机会,进而繁衍 后代。
交配行为与生殖器官协同作用
交配行为
动物在繁殖季节进行交配,通过生殖器官的 接触实现精子和卵子的结合。交配行为需要 雄性和雌性动物的生殖器官协同作用,以确 保受精成功。
跨性别动物形成原因分析
遗传因素
基因突变、染色体异常等 遗传因素可能导致动物出
现跨性别特征。
环境因素
激素水平异常、外界化学 物质干扰等环境因素也可 能影响动物的性别发育。
生理因素
部分动物在特定生理阶段 会表现出跨性别特征,如 鱼类中的性别转换现象。
跨性别动物在自然界中地位
生态作用
跨性别动物在自然界中扮演着重 要的生态角色,如传粉、捕食、 被捕食等。
性别角色认知
两栖动物性别决定相关基因的研究进展

动物学杂志Chinese Journal of Zoology 2011,46(6):134 140两栖动物性别决定相关基因的研究进展刘佳李忻怡张育辉*陕西师范大学生命科学学院西安710062摘要:两栖动物的性别决定机制主要包括遗传性别决定(genetic sex determination ,GSD )和环境性别决定(environmental sex determination ,ESD )。
近年来,在两栖动物性别决定和性腺分化机制的研究中,运用分子生物学技术探讨性别决定相关基因及其相互关系方面的研究已获得新的成果。
本文通过对DMRT 1、DAX 1、SF 1、SOX 3、SOX 9、FOXL 2、CYP 19、CYP 17在两栖动物性别决定中作用的分析,显示DAX 1、SF 1、FOXL 2、SOX 3均参与芳香化酶基因转录的调节,其中FOXL 2、SOX 3促进了CYP 19的表达,DAX 1、SF 1则与CYP 17的表达调节有关。
这些结果提示,两栖动物性别决定相关基因通过作用于CYP 19、CYP 17的表达调控性别决定过程,基因和温度分别在GSD 和ESD 过程中通过影响雌、雄激素的水平而决定两栖动物性别。
关键词:两栖动物;性别决定;基因;温度中图分类号:Q953文献标识码:A文章编号:0250-3263(2011)06-134-07Sex Determination-related Genes in AmphibiansLIU JiaLI Xin-YiZHANG Yu-Hui *College of Life Science ,Shaanxi Normal University ,Xi'an710062,ChinaAbstract :The sexual phenotype of amphibians is determined either by chromosomal factors (genetic sex determination ,GSD ),or by environmental factors (environmental sex determination ,ESD ).Recently ,new findings on the sex determination-related genes and their interactions have obtained by utilizing molecular biology methods.Several genes such as DMRT 1,DAX 1,SF 1,SOX 3,SOX 9,FOXL 2,CYP 19and CYP 17have been found to play roles in determining the sexual phenotype of amphibians ,with DAX 1,SF 1,FOXL 2and SOX 3involved in transcriptional regulation of aromatase gene.FOXL 2and SOX 3promote CYP 19expression.DAX 1and SF 1can influence CYP 17expression.Sex-determination genes play their roles by acting on the expression of CYP 19and CYP 17.Both sex-determination related genes and temperature determine sex of amphibians by affecting estrogen and /or androgen levels.Key words :Amphibians ;Sex determination ;Gene ;Temperature基金项目国家自然科学基金项目(No.130770243);*通讯作者,E-mail :yu-huizhang@163.com ;第一作者介绍刘佳,女,硕士研究生;E-mail :liujia1986jj @sina.com 。
Ch4-性别决定与性别控制

• 间接免疫荧光法 将胚胎用H-Y 抗体处理30分钟,再用异硫氰酸盐 荧光素(FITC)标记的二抗处理,有荧光者为雄性, 无荧光者为雌性。 牛的雌性鉴定准确率为89%,猪为81%,绵羊为 85%。 • 囊胚形成抑制法 利用H-Y 抗体对雄性桑椹期向囊胚期发育具有可 逆性抑制的原理。 大鼠H-Y 抗体与大鼠桑椹期胚共同培养6小时, 形成囊胚者为雄性胚,未形成囊胚者为雌性胚。
3.控制发育条件
(1) 调节发育温度 对鱼类、两栖类、爬行类的性别发育有很大影响。 鳄鱼无性染色体,性别有胚胎发育的温度决定: 30℃以下→全是雌性;34℃以上→全是雄性; 32℃以下→有雌有雄,但雄性比雌性多4倍。 (2) 使激素处理 用性激素控制鱼类性别取得很大进展。一般认为, 在性分化即将开始而性别尚未决定之前,连续投放 有激素的饵料,可控制性别发育。但已完成性分化 的鱼苗,不能完成性反转,只能抑制副性征。
3
免疫学方法
利用H-Y抗原或H-Y单克隆抗体检测胚胎上是否 存在雄性特异性H-Y抗原,从而对胚胎进行性别鉴 定。 • 细菌毒性分析法 在胚胎培养液中加入H-Y 抗血清与补体,在 培养过程中继续发育者为H-Y- (雌性);卵裂期溶 解或不能发育到囊胚期者为H-Y+ (雄性)。此法因 破坏部分胚胎(雄性),很少使用。
第四章
性别决定与性别控制
一.性别及其控制的意义 二.性别发育的概念 三.性别决定 四. 胚胎性别鉴定 五. 性别控制
一. 性别及其控制研究的意义
1.不同性别家畜的生产性能和经济价值差异很大: 母鸡产蛋,母牛产奶,公畜役力强、皮毛优,公鹿 产鹿茸,雄蚕产丝多等。 2.增加选育强度 ,加快遗传进展:采用 MOET技术 (Multiple Overlation and Embryo Transfer),只需常规 育种所需25%的优良种畜,就可使遗传进展提高1.27 倍。 3.作为胚胎移植, 核移植的配套技术。
Dmrt基因在脊椎动物性别决定与分化中的研究进展_雷蕾

第22卷第3期2013年6月中国组织化学与细胞化学杂志CHINESE JOURNAL OF HISTOCHEMISTRY AND CYTOCHEMISTRYVol.22.No.3June.2013Dmrt基因在脊椎动物性别决定与分化中的研究进展雷 蕾 李忻怡 张育辉*(陕西师范大学生命科学学院,西安710062) 〔摘要〕 脊椎动物的Dmrt(double-sex and mab-3related transcription factor)是指与果蝇Dsx基因和线虫Mab-3基因同源的基因家族。
本文阐述了Dmrt家族中8个成员的基因和蛋白结构特征,并通过分析Dmrt在各类脊椎动物发育中的表达显示,Dmrt家族主要参与性别决定和性腺分化,也参与某些非性腺组织器官的发育:Dmrt1主要参与精巢分化,也与卵巢发育有关;Dmrt2主要参与体节的发生、分化以及非对称器官的形成;Dmrt3,4,5可共同参与或依次调控嗅器和脑的发育;Dmrt7为雄性配子发生所必须;Dmrt8属雄性偏向表达基因。
Dmrt1在各脊椎动物的遗传性别决定通路中的位置和作用位点不同。
〔关键词〕 Dmrt基因家族; DM保守域; Dmrt基因表达; 脊椎动物; 遗传性别决定通路〔中图分类号〕 R329 〔文献标识码〕 A DOI:10.3870/zgzzhx.2013.03.018Research Progress in Dmrt Genes in Sex Determination and Differentiation of VertebratesLei Lei,Li Xinyi,Zhang Yuhui*(College of Life Sciences,Shaanxi Normal University,Xi’an 710062,China)〔Abstract〕 Dmrt is a gene family in vertebrates,which are homologous genes with DoublesexinDrosophila melanogaster and Mab-3in Caenorhabditis elegans.The characteristics of genes and proteinsof Dmrt members were expounded in this review respectively.Analysis of the expression of the develop-ment in various vertebrates of showed the Dmrt genes are mainly involved in sex determination and gonaddifferentiation,and also associated with the development of certain non-gonadal tissues and organs:Dmrt1is mainly involved in testis differentiation,but sometimes also relevant to ovarian development;Dmrt2ismainly involved in somitogenesis,differentiation and formation of asymmetric organs;Dmrt3,4,5canjointly or sequentially regulate the development of olfactory organs and brain;Dmrt7is required for malegametogenesis;Dmrt8is a male-biased expression gene.Dmrt1plays different roles in the genetic sex de-termination pathway among various vertebrates,varying in its position and functional locus.〔Key words〕 Dmrt gene family; DM conserved domain; Dmrt genes expression; Vertebrates; Genetic sex determination pathway〔收稿日期〕2013-02-20 〔修回日期〕2013-03-25〔基金项目〕国家自然科学基金资助(30770243),中央高校基本科研项目资助(200902039)〔作者简介〕雷蕾,女(1988年),汉族,硕士研究生。
【真题】2022年9月17日事业单位考试《综合应用能力》试题及答案解析(C类)

【真题】2022年9月17日事业单位考试《综合应用能力》试题及答案解析(C类)材料一男性与女性在某些疾病的患病率和对某些药物的反应上都存在差异,那么这些差异是如何与性别联系起来的呢?以色列魏茨曼科学研究所的一项研究发现,数千个能够编码蛋白质的基因的表达情况存在两性差异。
这些基因中的有害突变倾向于在人群中积累,而且具有较高的基因频率。
这些基因的基因图谱已经发表在BMCBiology上,进一步说明了男性和女性经历了不同而又互相联系的演化历程。
几年前,魏茨曼科学研究所分子遗传所的ShmuelPietrokovski教授和Moran Gershoni博士意识到,人类某些特定疾病的发病率普遍较高。
他们关注的一个典型案例是,希望生育的夫妇中约15%被诊断为不孕不育,这一数据说明导致生育能力降低的突变较为普遍。
但这种现象与常识相违背——减少后代数量进而影响存活个体数的突变,应该在自然选择过程中很快被淘汰掉,但为什么这种疾病的患病率然如此之高呢?Pietrokovski和Gershoni发现,影响精子形成的特定基因突变能够保留下来的原因是:这些基因仅仅在男性中表达。
当一个突变只能影响种群中的一半个体,那么无论危害大小,它都能够通过另一半个体畅通无阻传递给下一代。
在进一步研究中,研究人员的分析范围由生殖必需的基因扩大到两性间表达不相同的基因。
为了确定这些基因,研究人员开展了GTEx(Genotype-TissueExpression,基因型-组织表达)项目的研究。
该项目拥有一座人类基因表达的数据库,这些基因表达数据来自近550名成年捐赠者提供的器官和组织样本,使得研究人员第一次能够绘制两性之间具有差异表达的基因的基因图谱。
Pietrokovski和Gershoni分析了大约两万个编码蛋白的基因,按照性别将它们分类,以找出那些存在差异表达的基因。
最终发现,大约6500个基因的表达活性与性别有关,且至少在人体某一个组织中存在差异。
Dmrt基因在脊椎动物性别决定与分化中的研究进展

Re s e a r c h Pr o g r e s s i n Dt o r t Ge n e s i n S e x De t e r mi na t i o n a nd Di f f e r e n t i a t i o n O f Ve r t e b r a t e s
ma i n l y i n v ol v e d i n s omi t o ge ne s i s,d i f f e r e n t i a t i on a nd f o r ma t i on of a s y m me t r i c or g a ns ;D t or t 3,4,5 c a n
o f D r t me mb e r s we r e e xp o und e d i n t hi s r e v i e w r e s p e c t i v e l y . Ana l y s i s o f t he e xp r e s s i o n of t h e d e v e l o p —
[ Ab s t r a c t ]
D m r t i s a g e n e f a mi l Y i n v e r t e b r a t e s . wh i c h a r e h o mo l o g o u s g e n e s wi t h Do u b l e s e  ̄ :i n
me n t i n va r i ous ve r t e b r a t e s o f s ho we d t he Dm r t g e ne s a r e ma i nl y i nv o l ve d i n s e x d e t e r mi na t i o n a n d g on a d di f f e r e nt i a t i 0 n,a nd a l s o a s s oc i a t e d wi t h t he de ve l op me n t o f c e r t a i n no n go n a d a l t i s s ue s a nd or g a ns :D r t l i s ma i nl y i nv o l ve d i n t e s t i s d i f f e r e n t i a t i o n. bu l s o n l e t i ue r s a l s o r e l e v a nt t o ov a r i an de ve l op me nt ;D m r t 2 i s
鸡性别决定及分化关键调控基因DMRT1研究进展

畜牧兽医学报 2023,54(8):3152-3163A c t a V e t e r i n a r i a e t Z o o t e c h n i c a S i n i c ad o i :10.11843/j.i s s n .0366-6964.2023.08.003开放科学(资源服务)标识码(O S I D ):鸡性别决定及分化关键调控基因D M R T 1研究进展郑 钢,连 玲*(中国农业大学动物遗传资源与分子育种实验室,北京100193)摘 要:鸡是重要的农业动物之一,其所提供的鸡蛋是优质且廉价的动物蛋白来源㊂由于蛋鸡饲养中对母雏的需求偏好,使得早期性别鉴定成为蛋鸡生产中的重要一环,鉴定后会淘汰大量公雏,这不仅造成了巨大损失,还涉及到伦理道德问题㊂近年来,随着科学技术的发展使得人为控制畜禽性别成为可能,性别调控机制相关的研究挖掘出了很多候选基因,其中DM R T 1基因被认为是调控公鸡性腺形成的关键基因㊂本文系统的综述了DM R T 1基因在鸡性别决定及性腺分化中的作用㊁相关途径以及影响其表达的潜在因素,为后续深入研究家禽性别调控机制及开发人工性别控制技术提供参考㊂关键词:DM R T 1;鸡胚;性别决定;性腺发育 中图分类号:S 831.2 文献标志码:A 文章编号:0366-6964(2023)08-3152-12收稿日期:2022-12-30基金项目:国家重点研发项目(2022Y F F 1000204;2021Y F D 1300600);国家自然科学基金(32272865;U 1901206);国家蛋鸡产业技术体系(C A R S -40);家养动物种质资源库作者简介:郑 钢(1998-),男,安徽望江人,硕士生,主要从事家禽遗传育种研究,E -m a i l :j i u ji u 0820@163.c o m *通信作者:连 玲,主要从事家禽遗传育种研究,E -m a i l :l i a n l i n gl a r a @126.c o m R e s e a r c h P r o g r e s s o f K e y R e g u l a t o r y Ge n e D M R T 1i n C h i c k e n S e x D e t e r m i n a t i o n a n d D if f e r e n t i a t i o nZ H E N G G a n g ,L I A N L i n g*(L a b o r a t o r y o f A n i m a l G e n e t i c R e s o u r c e s a n d M o l e c u l a r B r e e d i n g ,C h i n a A g r i c u l t u r a l U n i v e r s i t y ,B e i j i n g 100193,C h i n a )A b s t r a c t :C h i c k e n i s o n e o f i m p o r t a n t a g r i c u l t u r a l a n i m a l s a n d t h e e g g s t h e y s u p p l i e d a r e a h i gh q u a l i t y a n d e c o n o m i c a l s o u r c e o f a n i m a l p r o t e i n .I n t h e l a y e r i n d u s t r y ,t h e e a r l y se x i d e n t if i c a t i o n i s i n d i s p e n s a b l e p r o c e d u r e d u e t o t h e p r e f e r e n c e f o r f e m a l e c h i c k s .L a r ge n u m b e r s of m a l e c h i c k s w e r e c u l l e d a f t e r s e x i d e n t i f i c a t i o n ,w h i c h n o t o n l y c a u s e s a h u ge l o s s b u t a l s o h a s e t h i c a l i s s u e s .I n r e c e n t y e a r s ,t h e d e v e l o p m e n t of s c i e n c e a n d t e c h n o l og yh a s m a d ei t p o s s i b l e t o a r t i f i c i a l l yc o n t r o l t h e s e x o f l i v e s t o c k a nd p o u l t r y .T he r e s e a r c h o n s e x r e gu l a t i o n m e c h a n i s m s h a s e x c a v a t e d m a n y i m p o r t a n t c a n d i d a t e g e n e s ,a m o n g w h i c h t h e DM R T 1g e n e i s c o n s i d e r e d t o b e a k e y g e n e i n t h e r e g u l a t i o n o f g o n a d f o r m a t i o n i n r o o s t e r s .T h i s p a pe r r e v i e w e d t h e r o l e of t h e DM R T 1g e n e i n chi c k e n s e x d e t e r m i n a t i o n a n d g o n a d a l d i f f e r e n t i a t i o n a s w e l l a s t h e r e l a t e dp a t h w a y s a n d t h e p o t e n t i a l f a c t o r s a f f e c t i n g i t s e x pr e s s i o n .T h i s r e v i e w p r o v i d e s a r e f e r e n c e f o r i n -d e p t h s t u d i e s o n t h e m e c h a n i s m o f s e x d e t e r m i n a t i o n a n d d e v e l o pm e n t o f f e a s i b l e a r t i f i c i a l s e x c o n t r o l a p pr o a c h e s .K e y wo r d s :DM R T 1;c h i c k e n e m b r y o ;s e x d e t e r m i n a t i o n ;g o n a d o g e n e s i s *C o r r e s p o n d i n g au t h o r :L I A N L i n g ,E -m a i l :l i a n l i n g l a r a @126.c o m8期郑 钢等:鸡性别决定及分化关键调控基因DM R T 1研究进展根据F A O (h t t p s :ʊw w w.f a o .o r g/f a o s t a t /z h /#d a t a /Q C L )统计数据,1961 2021年间,全球鸡蛋产量从1438万吨猛增至近8639万吨,中国是世界上最大的鸡蛋生产国,占全球生产总量的1/3左右㊂在规模化蛋鸡养殖产业中,雏鸡出生1日龄便对其进行性别鉴定,母雏全留,公雏被淘汰㊂据不完全统计,每年全球有将近60~70亿只公雏被扑杀[1],这不仅造成了巨大的经济损失,同时还面临严重的伦理问题㊂雏鸡的性别鉴定方法主要有翻肛法㊁羽色法和羽速法[2],这些方法都是通过对1日龄雏鸡进行人工性别鉴定,为了节省孵化成本,越来越多的学者致力于探索性别早期鉴定的方法,比如以D N A 检测为主的分子生物学方法㊁依据种蛋蛋形指数的蛋壳形态检测方法㊁以拉曼光谱和高光谱为主的光谱检测方法㊁提取尿囊液进行激素检测等,见表1[3]㊂但这些方法均存在一定的缺陷,如需要打开蛋壳等操作,存在影响胚胎发育的风险,同时上述方法也有检测准确率低㊁判定时间较晚等问题,此外分子生物学检测和光谱检测在应用上还存在操作复杂或检测成本高等问题,基于挥发物组成差异进行检测目前仅针对于京粉一号品种,且对试验环境和设备精度要求很高㊂随着生物技术的飞速发展,通过生物学手段人为控制或者逆转性别已成为可能㊂从性腺发育㊁分化以及性别决定基因入手挖掘性别控制的遗传基础已成为研究热点,其在科学研究以及产业应用上均具有重要价值㊂表1 鸡种蛋胚胎性别检测方法比较[3]T a b l e 1 C o m p a r i s o n o f e m b r y o s e x t e s t i n g me t h o d s [3]检测方法D e t e c t i o n m e t h o d判别时间/dJ u d gm e n t t i m e 准确率/%P r e c i s i o n 破坏性D e s t r u c t i v e n e s s 检测成本I n s pe c t i o n c o s t 分子生物学技术-100.00破坏高M o l e c u l a r b i o l o g y t e c h n i qu e s 15100.00破坏蛋壳形态学检测孵前-无损低E g g s h e l l m o r p h o l o gi c a l d e t e c t i o n 孵前80.21无损光谱检测法3.593.00破坏高S p e c t r o s c o pi c d e t e c t i o n 787.14无损1082.86无损1497无损激素检测法10-微创-H o r m o n e a s s a ys 16-破坏血线纹理特征-G A -B P 神经网络模型482.80无损低B l o o d -v e s s e l s t e x t u r e f e a t u r e -G A -B P n e u r a l n e t w o r k m o d e l透光性-光电检查法16~1884.00无损低T r a n s m i t t a n c e -p h o t o e l e c t r i c i n s p e c t i o n m e t h o d 挥发物组成差异算法分析(京粉一号)孵化早期100无损高A l g o r i t h m a n a l y s i s o f d i f f e r e n c e s i n v o l a t i l e c o m po s i t i o n - 代表无对应数据㊂表1引自参考文献[3],在其基础上有所改动 - r e p r e s e n t s n o c o r r e s p o n d i n g da t a .T ab l e 1i sc i t ed f r o m ref e r e n c e [3]w i t h m i n o r m o d i f i c a t i o n s 哺乳动物性染色体组成为X /Y ,雌性表现为同配(X X ),雄性表现为异配(X Y )㊂在奶牛上,通过流式细胞术分离携带X 或Y 的精子,能有效实现对后代性别的控制,但还存在受孕率低及高成本问题[4];而鸡和其他鸟类动物与此相反,雄性表现为同配(Z Z )而雌性表现为异配(Z W ),由于染色体组成的差异,无法在鸡上采取同样的精子分离方法进行有效的性别控制㊂早在1991年,研究人员就鉴定了哺乳动物中的Y 染色体连锁的睾丸决定基因S R Y (s e x d e t e r m i n i n g r e gi o n Y )[5],但目前为止,在包括鸡等鸟类动物中未能找到与S R Y 等效的性别决定基因,这提示鸟类动物存在不同的性别决定方式㊂鸟类动物的性别决定过程受到性染色体(Z Z/Z W )调控,这些性染色体中的一条或两条携带的基因在鸡胚发育早期能控制性别分化,在雄性个体中左右性腺都发育形成睾丸,而在雌性个体中,性腺发育左右不对称:左侧性腺产生功能性卵巢,而右侧性腺退化㊂鸡等鸟类动物的性别决定机制目前主要有3513畜牧兽医学报54卷两种假说[6]:一是Z染色体剂量效应假说:只有一条Z染色体的个体发育为雌性,有两条Z染色体的个体发育为雄性,该假说认为Z染色体剂量依赖机制是鸡性别决定的基础㊂其中Z连锁DM R T1(d o u b l e s e x a n d m a b-3r e l a t e d t r a n s c r i p t i o n f a c t o r1)基因的剂量效应是Z染色体剂量效应假说中最典型的证据,DM R T1在胚胎性腺发育中起关键作用,是调节睾丸发育形成的关键基因㊂二是W染色体显性效应假说:即W染色体上存在有显性的雌性发育相关基因能直接调控雌性性别决定过程,相关研究工作[7-9]发现一些W染色体连锁基因可能直接参与控制雌性的性别决定过程和表型的发生过程㊂S h a w等[10]和L i n等[11]发现,具有3A:Z Z W(A:常染色体A u t o s o m e)基因型的三倍体鸡表现为两性中间体,这些鸡有一个右侧睾丸和短暂出现的左侧卵巢(卵巢随年龄增长而逐渐退化),在发育成熟后表现为雄性外型㊂这表明,W染色体带有雌性性别决定相关基因,因为尽管存在两条Z染色体,仍可能形成一些卵巢组织,该发现也侧面佐证了Z染色体剂量对性别决定的重要影响㊂除上述两个假说外,激素调控以及细胞自主性识别(c e l l a u t o n o m o u s s e x i d e n t i t y,C A S I)假说也被广泛提及,激素调控性别假说认为由性腺分化而产生的性激素在性别分化中起调控作用,通过阻断或外源添加雌激素,能分别使雌鸡或雄鸡出现异性的性别特征,激素调控理论更多的是强调在性腺分化的基础上,性腺分泌的激素参与到性别分化进程㊂细胞自主性识别假说认为鸡等鸟类体细胞具有固有的性别识别方式,其性别分化本质上是细胞自主的[12],该假说认为雄性和雌性的差异主要不是由于激素作用的结果[13],且激素水平的改变不影响鸡等鸟类动物的体细胞自主性识别过程,这一点区别于人等哺乳动物激素水平对个体发育形态的影响,说明在鸡等鸟类动物中不以激素调节性别表型,提示这种特殊的调控机制背后涉及到复杂的基因调控㊂本文综述鸡性别决定及分化关键调控基因DM R T1的研究进展,从多个角度剖析DM R T1在鸡胚性别决定及分化中的重要作用以及影响DM-R T1表达的潜在因素,为深入研究DM R T1在鸡等鸟类动物性腺分化中的作用以及为研究鸡性别调控相关理论提供参考㊂1鸡胚性别决定及分化进程性别决定是指有性繁殖生物中,产生性别分化,并形成种群内雌雄个体差异的基础,性别分化指受精卵在性别决定的基础上,进行雌性或雄性性状的发育过程㊂脊椎动物的性别决定分为遗传型性别决定(g e n e t i c s e x d e t e r m i n a t i o n,G S D)和环境型性别决定(e n v i r o n m e n t d e p e n d e n t s e x d e t e r m i n a t i o n, E S D)[14]㊂哺乳动物和鸟类的雌雄两性个体具有明确的性染色体差异,其性别决定过程受性染色体的遗传调控[15]㊂性腺是由生殖细胞和体细胞两类细胞组成的,原始生殖细胞(P G C)来源于胚胎发育早期的中胚层,然后经过定向迁移到达生殖嵴(g e n i t a l r i d g e),与性腺体细胞共同发育为性腺㊂值得注意的是,与其他脊椎动物不同,鸡胚胎支持细胞(s e r-t o l i c e l l s)并非来自腔上皮,它们来自DM R T1(+)/ P A X2(+)/WN T4(+)/O S R1(+)间充质细胞群,在生殖细胞迁移到性腺的早期,这些细胞定植于未分化的生殖嵴[16]㊂另外,雌雄鸡胚早期都有两套原始生殖管道:一对沃尔夫氏管(W o l f f i a n d u c t,又称中肾管)和一对缪勒管(M u l l e r i a n d u c t,又称中肾旁管)㊂在雄性个体中,沃尔夫氏管发育形成雄性生殖管道,而缪勒管退化;而在雌性个体中,沃尔夫氏管退化,缪勒管则形成雌性生殖管道㊂鸡最早期性别决定发生在鸡胚受精卵形成时,由差异化的染色体组成(Z Z/Z W)决定㊂鸡胚性腺发育历程简化如图1所示[17]㊂鸡的胚原基(原始性腺)在胚胎发育E3.5d开始形成并发育成原始生殖嵴,有报道指出早在E2.0d时[18],侧板中胚层(L P M)腹内侧的细胞就被确定为性腺祖细胞(G P C),G P C在E2.0d-E3.0d时受H e d g e h o g信号激活进而分化形成性腺细胞并形成生殖嵴, E3.0d时腹内侧出现明显的增厚,这是鸡性腺发生的第一个迹象;在E3.5d-E4.5d时,还无法在形态上区分性腺的雌雄差异,在E5.5d时,性腺在形态上表现是外部上皮细胞层和密集的下层髓质中的细胞索,到E5.5d-E6.5d开始出现形态上的差异,逐渐分化形成雌/雄性腺㊂在雄性胚胎中,性腺皮质开始退化,生殖细胞与髓质相互作用形成生精小管索结构㊂而在雌性胚胎中,左侧性腺体细胞和生殖细胞在皮质内增殖,皮质层显著增厚,右侧性腺则开始退化;在E6.5d-E21.0d期间,两性性腺出现显著形态学差异:雄性睾丸对称且大小相似,两侧性腺组织中的皮质层进一步变薄,髓质形成管腔结构,生殖细胞与周围的支持细胞组成生精小管索㊂而雌性卵巢组织不对称,左侧性腺皮质层逐渐增厚且结构致45138期郑钢等:鸡性别决定及分化关键调控基因DM R T1研究进展密,髓质结构疏松,卵母细胞开始进行减数分裂,尽管右性腺也能类似于左性腺发生髓质空泡化,但它无法形成增厚的富含生殖细胞的皮质,右侧性腺逐渐发生萎缩㊂相关生殖管道的发育在鸡两性间也存在差别,雄性缪勒管在孵化E8.0d停止发育,并在E12.0d消退;而雌性左侧缪勒管发育形成输卵管,右侧导管在E12.0d后经历相对缓慢的退化后在孵化时完全消失[19]㊂图1鸡胚性腺性别分化示意图[17]F i g.1T h e s c h e m a t i c o f g o n a d a l s e x d i f f e r e n t i a t i o n i n c h i c k e m b r y o s[17]2鸡胚性别决定或者性别分化过程中涉及的重要基因鸡胚早期性别决定和性别分化过程中涉及很多相关基因的参与㊂目前研究较多的基因主要有DM R T1㊁S F1(o r p h a n n u c l e a r r e c e p t o r s t e r o i d o-g e n i c f a c t o r-1)㊁L HX9(L I M h o m e o b o x p r o t e i n)㊁D A X1(N R0B1,n u c l e a r r e c e p t o r s u b f a m i l y0 g r o u p B m e m b e r1)㊁S P I N1/S P I N1L(s p i n d l i n g 1/s p i n d l i n g1l i k e)㊁S O X9(s r y-b o x t r a n s c r i p t i o nf a c t o r9)㊁H E M G N(Z-l i n k e d m a l e f a c t o r,h e m o-g e n)㊁AMH(a n t i-M u l l e r i a n h o r m o n e)㊁WT1 (W i l m s t u m o r1)㊁C Y P19A1(c y t o c h r o m e P450 f a m i l y19s u b f a m i l y A m e m b e r1)㊁F O X L2(f o r k-h e a d b o x L2)㊁E S R1(e s t r o g e n r e c e p t o r1)㊁H I N TW(h i s t i d i n e t r i a d n u c l e o t i d e b i n d i n g p r o t e i n W)㊁U B E2I(u b i q u i t i n c o n j u g a t i n g e n z y m e E2I)㊁WN T4(W n t f a m i l y m e m b e r4)等㊂其中DM R T1㊁AMH㊁S O X9㊁H E M G N等在雄性发育中起关键调节作用,而C Y P19A1和F O X L2等在雌性发育中起关键的调节作用㊂上述基因中,DM R T1被广泛认为是调控公鸡睾丸形成的核心基因,DM R T1基因的剂量直接影响鸡性别决定进程㊂3D M R T1简介及胚胎期原位表达DM R T1最早在无脊椎动物中被发现,是一种古老的性别决定基因,是少数几个在鱼类㊁龟类㊁鳄鱼㊁两栖动物㊁鸟类和哺乳动物的代表性物种中被绘制的性别相关基因之一[20],DM R T1在许多进化物种中均表现出性别二态性(指同一物种不同性别之间的差别)表达,是脊椎动物性别决定通路中的保守成分[21-22]㊂鸡DM R T1基因位于Z染色体26.45~ 26.50M b之间,其编码的m R N A长度为1244b p,由6个外显子组成,经典蛋白产物长度是365a a (U n i p r o t)㊂D M R T1蛋白位于核内,是D M结构域(D M d o m a i n)家族转录因子之一,D M结构域在进化中高度保守,D M结构域基因是最早在脊椎动物和无脊椎动物门类中显示出性别二态性表达的基因[21]㊂DM R T1的表达具有很强的组织特异性,主要在性腺及相关组织中表达㊂O m o t e h a r a等[23]于2014年研究了鸡胚胎发育过程中D MR T1蛋白在雄性和雌性鸡胚的泌尿生殖系统(包括缪勒管)的表达模式(图2)㊂发现在性别决定的推测期(E4.5d)5513畜 牧 兽 医 学 报54卷之前,雄性未分化性腺的性腺体细胞与雌性相比表现出更强的DM R T 1的表达,在鸡胚性别决定之后的E 6.5d,雄性鸡胚形成睾丸索的支持细胞表达DM R T 1,同时发现在性别开始分化的E 4.5d 后,雄性和雌性的生殖细胞同样能表达DM R T 1,但在雄性鸡胚的表达是连续的,而在雌性鸡胚发育过程中,在左侧卵巢皮质生殖细胞中的表达不连续-从左侧卵巢皮层中央部分的生殖细胞向两侧边缘逐渐消失㊂同时DM R T 1也在E 4.5d -E 7.5d 的输卵管嵴(缪勒管的前体)中被检测到,在雌/雄鸡胚缪勒管的间充质和最外层的腔上皮都能检测到DM R T 1的表达,DM R T 1在雄性缪勒管退化前表达高,在E 8.0d ,雄性缪勒管消退,DM R T 1表达仅限于间充质区域,而雌性表达模式没有变化㊂尽管该研究对DM R T 1基因原位表达做了较为详细的论述,但该研究仅主要集中在E 4.5d -E 15d ,相关研究发现早在E 3.5d 的鸡胚生殖嵴中就已经有DM R T 1的表达[24],有关覆盖胚胎性腺发育全阶段DM R T 1的时空表达图谱还有待进一步的完善㊂线宽表示与异性相比的相对表达水平㊂阴影线表示性别决定的推定期T h e l i n e w i d t h r e p r e s e n t s t h e r e l a t i v e e x p r e s s i o n l e v e l c o m -p a r e d w i t h t h a t i n t h e o p p o s i t e s e x .A s h a d e d l i n e i n d i c a t e s t h e p r e s u m p t i v e p e r i o d o f s e x d e t e r m i n a t i o n 图2 D M R T 1在鸡胚泌尿生殖系统中的时空和性别二态性表达模式图[23]F i g .2 T h e s c h e m a t i c o f t h e s p a t i o t e m po r a l a n d s e x u a l d i m o r -p h i c e x p r e s s i o n p a t t e r n s o f D M R T 1i n t h e u r o ge n i t a l s y s t e m of t h e c h i c k e n e m b r yo [23]值得一提的是A y e r s 等[25]在2015年报道了DM R T 1不仅在早期形成的鸡胚缪勒氏管嵴中表达,还在导管形态发生期间经历从上皮细胞转变到在间充质细胞中表达,同时发现在鸡胚中敲低DM -R T 1会导致间充质层大大减少,阻断导管管腔上皮的尾部延伸,因此提出DM R T 1是缪勒管发育的早期必需步骤㊂然而I o a n n i d i s 等[26]在2019年研究DM R T 1基因敲除后的鸡胚(Z -W )发育时,发现其能形成正常左侧缪勒管,该结果对DM R T 1在缪勒管发育早期的必要性提出了质疑㊂值得注意的是O m o t e h a r a 等[27]在2017年研究发现,部分表达DM R T 1的皮质细胞仅在左侧睾丸发育开始后对左侧睾丸髓质中的支持细胞有贡献,结果表明DM -R T 1在睾丸中的表达存在不对称性㊂4 雄性鸡睾丸形成依赖于D M R T 1基因剂量早在1999年,R a ym o n d [28]就发现DM R T 1在鸡性别分化之前的生殖嵴和沃尔夫氏管中表达,且在Z Z 型胚胎中的表达水平高于Z W 胚胎;S h a n 等[24]在2000年的研究中发现,DM R T 1在E 3.5d的雄性生殖嵴中的表达高于雌性;N a n d a 等[29]在2000年基于染色体同源性研究中发现,DM R T 1基因与人类上的直系同源物和人类X Y 性别逆转相关,提出DM R T 1是脊椎动物性别决定最早期的剂量敏感基因㊂O r a l 等[30]在2002年通过对性腺分化前后相关基因表达趋势的分析,发现在E 5.0d 时DM R T 1在整个性腺中表达,但由于原位杂交技术的局限性,未在此阶段揭示该基因表达是否存在明确的性别二态性,但在E 6.0d 的鸡胚中,雄性比雌性显著高表达㊂随后,E 7.0d 的雄性性腺中出现AMH 和DM R T 1的高表达(DM R T 1在雄性性腺中比雌性性腺表达高两倍以上)以及S O X 9的开始表达,且同时伴随着睾丸索的形成;除上述基因外,其他基因不呈现显著的性别二态性㊂另外,Y a m a m o t o 等[31]在2003年发现DM R T 1表现出雄性特异性表达模式,提出DM R T 1㊁S O X 9和AMH在E 5.5d -E 8.5d 时与睾丸形成有关㊂S m i t h 等[32]2003年在使用芳香酶抑制剂F A D (f a d r o z o l e)诱导的雌性反转为雄性的鸡胚中分析DM R T 1的表达,发现性反转胚胎DM R T 1表达水平升高,该现象与具有两个Z 染色体拷贝的正常雄性个体相似,基于此结果,他认为睾丸发育中确实涉及DM R T 1基因上调,但两个拷贝DM R T 1并非是必须的;此外郑江霞和杨宁[33]在2007年,利用经芳香化酶抑制剂处理产生的性反转鸡胚进行试验,也发现D M R T 1的上调表达与睾丸形成有关㊂2009年,65138期郑钢等:鸡性别决定及分化关键调控基因DM R T1研究进展S m i t h等[34]给出了有关DM R T1表达与鸡性别决定有关的最直接证据,他们使用R N A干扰技术(R N A i)敲低早期鸡胚中的DM R T1基因,发现试验组的雄性表现出部分性别逆转,导致遗传雄性(Z Z)胚胎出现性腺的雌性化,包括左性腺显示雌性样组织㊁睾丸索紊乱㊁睾丸标志物S O X9下降㊂该研究还发现,在性腺雌性化的Z Z鸡胚中,卵巢标志物芳香酶被异位激活,而且相比较于左侧性腺,右侧性腺DM R T1和异位芳香酶活化的变化更大,表明左右性腺对DM R T1的敏感性不同㊂随后L a m b e t h等[35]在2013年使用逆转录病毒载体R C A S B P在鸡胚中异位表达芳香酶基因C Y P19A1,发现雄性鸡胚胎中过表达C Y P19A1诱导了雄性性腺向雌性的反转,此外还发现雄性性腺发育的关键基因DM R T1㊁S O X9和AMH的表达受到抑制,性反转雄性个体的生殖细胞分布和雌性相似㊂同年,F a n g等[36]的研究指出在性别决定和性腺分化期间,由外源雌激素诱导雄性到雌性的性反转胚胎中,在E3.0d-E5.0d期间DM R T1表达活性较低㊂随后2014年L a m b e t h[37]的研究还发现,DM R T1在雄性胚胎性腺中表达上调是出现在H E M G N㊁S O X9和AMH表达之前的,这表明DM R T1在胚胎雄性性别决定中处于更靠前的位置,另外过表达DM R T1会诱导雄性发育相关基因的表达并拮抗胚胎性腺中的雌性相关途径,而在雌性性腺中异位表达DM R T1能诱导局部AMH㊁S O X9㊁H E M G N等雄性相关基因的表达㊂2017年H i r s t等[38]在鸡胚发育E3.0d时外源注射F A D至胚胎中,最终诱导了雏鸡雌性到雄性的性反转,在反转个体性腺中发现芳香酶活性显著丧失并伴随着性腺出现雄性化特征,但W染色体连锁的H I N TW㊁F A F和F E T1基因的表达水平在雌性鸡性反转前后没有差异,据此,提出是Z连锁的DM R T1而不是W性染色体调控鸟类的性别分化过程㊂DM R T1剂量对鸡性别分化影响的最直接的证据是I o a n n i d i s等[26]在2021年的报道,基于C R I S P R-C a s9的单等位基因靶向方法产生具有DM R T1基因靶向突变的鸡,发现由此产生的具有DM R T1单一功能拷贝的染色体雄性(Z+Z-)鸡发育形成卵巢,直接证明鸡性别决定机制基于DM-R T1剂量,结果说明了在雄性个体中,DM R T1剂量(两个拷贝)对睾丸形成命运的决定作用,有趣的是突变鸡(Z+Z-)的外形㊁生长速度和肌肉发育等和野生型的公鸡更为相似,该结果也支持了鸟类动物的C A S I假说㊂同年L e e等[39]也做了相似的工作,采用C R I S P R-C a s9破坏DM R T1起始密码子,发现在雄性鸡胚发育的早期阶段,DM R T1的破坏诱导性腺雌性化,在激素合成紊乱的情况下,雌性的功能性生殖能力无法实现㊂5D M R T1作用途径有关DM R T1在性别决定和分化中如何发挥作用尚未有完善的试验性结果,但大多数研究推测认为DM R T1主要是与F O X L2拮抗抑制雌激素通路从而调节性腺分化的进程㊂S n c h e z和C h a o u i y a[40]2018年依靠已发表的试验数据组装了一个基因网络,形成了一个整合Z染色体剂量效应和W染色体显性效应的假设逻辑模型(图3)㊂该模型表明,鸡性腺的命运是由DM R T1和F O X L2之间存在相互抑制的关系引起的,其中DM R T1产物的初始量决定了性腺的发育㊂H i r s t等[41]在2018年的研究中指出,性腺中表达的DM R T1可以激活睾丸发育中的S O X9㊁AMH和H E M G N基因,同时发现DM R T1还抑制卵巢通路基因,如F O X L2和C Y P19A1,但雌性性腺中较低水平的DM R T1表达与卵巢通路的激活是可以同时发生的㊂M a j o r等[42]在2019年的报道中指出,雄性鸡胚性腺中F O X L2的错误表达会抑制睾丸发育途径,消除雄性发育相关基因DM R T1㊁S O X9和AMH的局部表达的同时会抑制支持细胞发育,也验证了F O X L2与DM R T1潜在的拮抗关系㊂I o a n n i d i s等[26]在2021年较为系统的阐述了DM R T1和F O X L2雌激素相关通路在调控性腺分化中的相互作用(图4)㊂该研究指出,在DM R T1表达正常的Z+Z+(两个拷贝)鸡胚胎中,DM R T1抑制F O X L2的表达,进而引起芳香化酶的表达减少和雌激素合成减少,在Z+W胚胎中,DM R T1的量不足以抑制F O X L2的表达,从而导致雌激素E2对睾丸发育通路的抑制㊂另有研究发现,在DM-R T1基因突变的Z-W胚胎中,虽然发育形成了卵巢结构,但与Z+W相比,性腺在形态上更小且皮质层薄,不能正常进行减数分裂,这表明DM R T1的存在对于雌性性腺正常发育也是必须的;在E2合成受阻的Z+W和Z+Z-鸡胚胎中,雄性相关发育通路未被抑制,发育形成了睾丸结构,同时在对照组野7513畜 牧 兽 医 学 报54卷Z 1和Z 2表示任意一条Z 染色体,而W 表示W 染色体;粗㊁细实线箭头分别表示促进和抑制作用,虚线箭头表示间接或潜在的作用Z 1a n d Z 2r e p r e s e n t e a c h Z c h r o m o s o m e ,w h e r e a s W r e pr e -s e n t s W c h r o m o s o m e ;T h i c k ,t h i n s o l i d a r r o w s r e pr e s e n t p o s i t i v e a n d n e g a t i v e i n t e r a c t i o n s ,r e s p e c t i v e l y ,an d d a s h e d a r r o w s i n d i c a t e i n d i r e c t o r p r o po s e d i n t e r a c t i o n s 图3 控制鸡早期性别决定的简化基因调控网络图[33]F i g .3 T h e s i m p l i f i e d g e n e r e g u l a t o r y n e t w o r k c o n t r o l l i n gc h i c k e n p r i m a r ys e x d e t e r m i n a t i o n [33]2x 和1x 表示2个拷贝和1个拷贝;虚箭头和实箭头分别表示雄性和雌性的作用,线宽表示作用的强弱2x a n d 1x r e p r e s e n t 2c o p i e s a n d 1c o p y ,r e s p e c t i v e l y;T h e d o t t e d a n d s o l i d l i n e a r r o w s r e pr e s e n t t h e e f f e c t i n m a l e s a n d f e m a l e s ,r e s p e c t i v e l y ,a n d l i n e w i d t h r e p r e s e n t s t h e s t r e n gt h o f t h e e f f e c t图4 雄/雌鸡胚生殖系统中D M R T 1基因网络调节示意图[26]F i g .4 T h e s c h e m a t i c o f D M R T 1g e n e n e t w o r k r e g u l a t i n g th e m a l e /f e m a l e c h i c k e m b r y o r e p r o d u c t i v e s ys t e m [26]生型Z W 胚胎的雌激素合成阻断试验中,性腺发育形成睾丸,但在E 2合成受阻的Z -W 鸡胚胎中性腺髓质类似于卵巢,结果充分表明了DM R T 1的剂量效应对雌性发育的影响依赖于雌激素,DM R T 1在雌性个体形成卵巢的发育过程中也起到了非常重要的作用㊂目前相关研究发现,DM R T 1直接或间接激活雄性相关因子H E M G N ㊁S O X 9和AMH 的表达[43],但其在禽睾丸发育过程中的直接转录靶标目前尚不清楚㊂DM R T 1促进雄性胚胎中的睾丸发育而抑制雌性胚胎中的卵巢发育,DM R T 1很可能在鸡胚性腺中既能充当转录激活因子又能充当转录抑制因子,但这些还缺乏试验性数据支持㊂DM R T 1基因在其他物种上的研究也值得借鉴,如在幼年小鼠睾丸中,采用染色质免疫共沉淀结合高通量测序技术(C H I P -s e q)和R N A 表达分析测定DM R T 1全基因组靶标,发现D M R T 1蛋白大约能结合到1400个近端启动子区域,这里还包括DM R T 1本身[44]㊂L i n d e m a n 等[45]在小鼠体内和体外细胞培养中发现,S O X 9和DM R T 1在颗粒细胞重编程成支持细胞中协同发挥作用,提出DM R T 1可作为先驱因子开放染色质从而引起S O X 9的结合㊂另外,G a o 等[46]于2005年在斑马鱼上的研究发现,S O X 5结合DM R T 1启动子并抑制其表达,指出DM R T 1和S O X 5之间的拮抗关系,同时也表明DM R T 1在早期胚胎发生中存在潜在的转录调控机制;L e i 等[47]在2009年对大鼠的研究发现,F O X L 2通过3.2k b /2.8k b 调控区抑制DM R T 1启动子,为颗粒细胞中的DM R T 1转录沉默提供了潜在原因;T a n g 等[48]和W e i 等[49]在2019年发现,DM R T 1通过直接结合S O X 30和S O X 9B 启动子内的特异性C R E 正调控罗非鱼S O X 30和S O X 9B 的转录㊂上述物种上的研究为解析DM R T 1在鸡等鸟类动物上复杂的分子机制提供了很好的参照㊂6 D M R T 1的潜在调控机制研究6.1 m i R N A s 的调控m i R N A s 是机体内调控基因转录后表达的重要途径,m i R N A s 通过靶向目的基因m R N A s 的3'U T R 区域沉默m R N A s 表达或者降解m R N A s ,以达到对基因表达的抑制作用㊂相关研究有证据支持m i R N A s 在鸡胚胎性腺发育中发挥作用,而关于m i R N A s 和DM R T 1的研究提示m i R N A s 可作为DM R T 1潜在的调控靶点㊂B a n n i s t e r [50]使用雌激素诱导鸡胚雄性性腺到雌性的反转后,M I R 202*的85138期郑钢等:鸡性别决定及分化关键调控基因DM R T1研究进展表达降低到正常雌性鸡胚的表达水平,而用芳香酶抑制剂诱导雌性到雄性的性反转后,M I R202*表达增加,研究还发现M I R202*表达降低与睾丸相关基因DM R T1表达减少相关,而M I R202*表达增加与F O X L2和芳香酶的下调以及DM R T1和S O X9的上调相关㊂结果证实,M I R202*的上调与胚胎鸡性腺的睾丸分化方向一致㊂此外,C u t t i n g 等[51]2012年通过对性腺中m i R N A s的表达分析发现一些m i R N A s在性腺中表达的二态性,也提出了m i R N A s能潜在调节DM R T1的表达㊂W a r n e f o r s等[52]在2017年对雌/雄鸡不同组织m i R N A s进行测序,研究结果发现了很多具有明显性别偏向性表达的m i R N A s,其中m i R-2954-3p编码基因位于Z染色体上,表现出雄性偏向性表达,且显示出对鸡Z染色体上剂量敏感基因的保守偏好㊂同时依据他们的测序结果,笔者发现g g a-m i R-30e-3p㊁g g a-m i R-2954-3p㊁g g a-m i R-202-3p㊁g g a m i R-153-5p㊁g g a-m i R-6562_M2-3p㊁g g a-m i R-6562_M1-3p㊁g g a-m i R-138-2-3p和D M R T1也存在靶向关系,同时在性腺中也具有显著的性别偏向表达,可作为后续DM R T1基因研究的潜在靶点㊂此外,P r a s t o w o和R a t r i y a n t o[53]使用3个在线数据库(即m i R D B㊁T a r g e t S c a n和m i c r o T-C D S)挖掘靶向鸡DM R T1的m i R N A s,共得到78个靶向DM R T1的3'U T R 的m i R N A s,在最少两个数据库中发现了8个m i R-N A s㊂这些研究结果提示,m i R N A s在DM R T1的表达调控中发挥重要作用,但深入的机制解析还有待开展㊂m i R N A s作为药物治疗已经有广泛的应用,筛选潜在作用于DM R T1基因的m i R N A s可能是实现性别调控的重要手段㊂6.2鸡雄性高甲基化区域(c MH M)调控由于在鸡上没有直接证据表明Z染色体随机失活现象的存在,但鸡Z连锁基因在雌雄个体间的平均表达差异是1ʒ1.4~1.8[41]而不是1ʒ2,这说明有相关机制起到了一定的剂量补偿作用,其中雄性Z染色体上的MHM以及其附近区域的转录活性较低,对邻近一些基因表达产生了一定的抑制作用,所以MHM也被认为是剂量补偿效应的一个解释㊂MHM最早由T e r a n i s h i等[54]在鸡Z染色体的短臂上发现,位于鸡Z染色体的27.140~27.398M b区域内,其包含有200多个长度为2.2k b的串联重复序列,在雄性体内高甲基化且没有转录活性,但MHM在雌性个体中低甲基化并能转录形成长链非编码R N A(l n c R N A)[54-55]覆盖Z染色体㊂另外T e r a n i s h i等[54]还发现,在Z Z Z三倍体细胞中,所有3个Z染色体的MHM区域都是高甲基化和无活性的,而在Z Z W三倍体中,MHM区域是低甲基化并且两个Z染色体都能转录,这些结果提示W染色体或许存在某些特殊的调节方式调控MHM区域的表达㊂Y a n g等[56]在2010年为了分析MHM对性别依赖基因表达的调控作用,构建含有鸡MHM的外源质粒并注射到13周龄公鸡的左侧睾丸中,结果发现用外源性p E G F P-N1-c MHM质粒处理的公鸡睾丸中DM R T1表达下调㊂R o e s z l e r等[55]在2012年的研究中发现,在胚胎阶段雄性个体中的MHM的错误表达会导致DM R T1的表达受损;另外C a e t a-n o等[57]发现,在同一发育阶段,雌性中的MHM上调而DM R T1下调,从而提出MHM可能在卵巢发育中起作用,这些结果充分表明MHM n c R N A对DM R T1存在潜在调节作用㊂S u n等[58]在2019年对鸡Z染色体研究发现了两个雄性高甲基化位点MHM1(之前报道的MHM)和MHM2,MHM1㊁MHM2分别位于染色体27.140~27.398M b和73.160~73.173M b区域㊂在鸡的整个发育阶段,与大多数体细胞中的Z 染色体其余部分相比,位于MHM1或MHM2附近(25~32M b,72.5~73.5M b)基因的表达(指雄性ʒ雌性的相对表达量)会降低,笔者发现DM R T1基因刚好位于Z染色体26.45~26.50M b之间,其表达可能受到MHM1的影响,但S u n等[58]未在性腺组织上采样进行比较,因此,在性腺组织中DM R T1转录活性是否也会受到MHM1影响还有待探索㊂S h i o d a等[59]也报道了与之类似的表观遗传修饰对性腺分化的影响,其将遗传雄性(Z Z)鸡胚用外源性雌激素刺激处理后发现,在孵化时性腺暂时出现雌性化,性腺雌性化的Z Z个体在1年内被雄性化回退形成睾丸㊂该研究指出,性腺雌性化的Z Z 个体中,可能其体细胞群和生殖细胞群都保持着遗传性别的转录组和表观遗传记忆㊂这种表观遗传记忆是否与DM R T1基因功能有关也还需进一步研究㊂7结论与展望从对鸡DM R T1基因的研究进展来看,DM-9513。
鱼类性别决定及分化相关基因研究进展

鱼类性别决定及分化相关基因研究进展作者:路畅,苏利娜,朱邦科来源:《湖北农业科学》 2014年第13期路畅1,2,苏利娜1,朱邦科2(1.华中农业大学水产学院,武汉430070;2.宁波大学海洋学院,浙江宁波315211)摘要:综述了Sox、DMRT、芳香化酶、FTZ-F1、FOXL2、Pod1、GSDF、Fanconianemia/BRCA等一些与鱼类性别决定及分化相关的基因的研究动态与进展。
旨在为系统研究鱼类性别决定机制提供参考。
关键词:鱼类;性别决定基因;Sox;DMRT;芳香化酶基因;FOXL2中图分类号:S965;Q132.1文献标识码:A文章编号:0439-8114(2014)13-2981-05鱼类是脊椎动物中最低等但却是分布最广、种类最多的一类生物。
鱼类在进化上比较原始,与高等脊椎动物相比,其性别决定的遗传力较小。
鱼类的性别发育以遗传因素为基础,并受到外界环境和自身内分泌调节的影响,是三者相互作用的结果,因此鱼类的性别决定机制复杂多变,而且没有一个普遍的模式。
鱼类中存在从雌雄同体到雌雄异体,从遗传决定型到环境决定型的各种性别决定类型,性逆转在鱼类中也是较为常见的现象,基本上具有所有脊椎动物的性别决定方式,因此鱼类是一个极好的研究性别决定机制进化的模型。
了解鱼类性别决定特点对于研究者寻找鱼类性别决定相关基因及性染色体有很大的帮助。
鱼类的性染色体组成形式多样,且其性染色体的出现与否及性染色体的种类与它们的系统进化地位无关,在亲缘关系较近的种类中,可能会具有完全不同的性染色体类型,如尼罗非鱼(Tilapianilotica)为XX/XY型,而奥利亚罗非鱼(Tilapiaaureus)为ZW/ZZ型[1]。
据统计资料显示,在所研究的鱼类中,仅有10%左右的鱼类具有异型性染色体,只在个别鱼类中,如青鳉(Oryziaslatipes)[2]已鉴定出性别决定候选基因DMY,可以认为其性别是由性别决定基因控制的。
水产动物性别决定的相关因素

展 望
水产动物性别决定至今还没有一个普遍的 模式。但随着分子生物学和分子遗传学的 发展、人类基因组和后基因组计划的实施, 水产动物性别决定的分子遗传研究领域也 在不断地拓宽。水产动物性别决定基因的 发掘必将成为研究的热点和重点。
参考文献
[1]楼允东,吴萍,温度在水产动物性别控制中的作用.上海水 产大学学报.2008年7月 [2]张翠敏,杨永刚,姚青梅,胡 山,海产育苗养殖水温控制 系统设计.制造业自动化.2011 [3]勤超,人工诱导黄颡鱼多倍体技术研究.华中农业大学硕士 学位论文.2010.06 [4]孙业盈,半滑舌鳎性腺差异表达基因的克隆鉴定与表达分析 ,中国海洋大学博士学位论文.2008.06 [5]龚军辉.环境与性别分化.高等函授学报.2005.06 [6]麻艳群,鱼类性别决定基因研究综述.动物科学.2011 [7]聂振昌 陈开健,性别决定基因(Sox9,DMRT 1)在水产动物 的研究与应用探讨,河北渔业.2009年第4期 [8]王志勇, 蔡明夷, 刘贤德, 姚翠鸾, 林利民, 谢芳靖, 刘家富.大黄鱼人工雌核发育和性别控制的研究.2007
温度
激素 环境因子
人工诱导 雌雄核发育
盐度、光照、 水质、PH值和 食物丰度等
人工诱导 多倍体
温度
温度诱导水产动物性别分化的机理尚处于探索中, 那么,温度如何影响水产动物的性别分化?就鱼类 来说,较为认同的看法是温度影响体内性激素(雄 激素和雌激素)的产生。在雄激素的作用下,鱼会 趋向雄性分化;而在雌激素的作用下,鱼则趋向 雌性分化。
欢迎提问指正!!!
2n n 已抑制第二极体 排放的卵子 精子
精子保留
2n n 3n
行为对性别分化的影响
性别决定基因(Sox9,DMRT1)在水产动物的研究与应用探讨

的 内分 泌 腺— — 促 雄 性 腺 ( n rg ncga d 也 a do e i ln )
有 举 足轻重 的作 用 。在 已报 道 的虾蟹 类性 别决定 机制 中, 遗传 ( 染 色 体 ) 有 性 决定 型 、 境 ( 化 环 环 理 境 或 社会环 境 ) 决定 型 以及 遗 传 与 环 境 ( 因子 ) 多 共 同决定 型 。这些 都说 明 甲壳 动物 的性 染色体或
X XY 型性 别 决 定 ; 大 部 分 两 栖 类 动 物 的性 X/ 但
染 色体 尚未 出现异 型 质分 化 。 龟类 是 具有 约两 亿年 进化 史 的一支 古老 的爬
以通 过 其行 为控 制 其 他 个 体 的性 别分 化 , 时提 同
行动 物 。 自 2 0世 纪 7 O年 代 以来 , 龟类 的性 别 决
性 别决 定机 制仍 处 于进 化 的早 期 阶段 。 两 栖类 动物 是从 水 生到 陆生 进化 上 的一 个关
l 水 产 动 物 的性 别 决 定 机 制 [ ] 1
1 1 鱼 类 的 性 别 决 定 机 制 .
键 环节 , 同时 , 栖类 动 物 的染 色 体代表 着染 色体 两
龟类 动物 都缺 乏 性 染 色 体 的分 化 , 别决 定 受 环 性
境 温度 的影 响 。
因是 生 物 体 的 性 别 遗 传 物 质~ 性 染 色 体 ( e sx
c r mo o ) 外 就 是性 腺 性 别 尚未 分 化 前 那 h o sme , 些影 响 分 化 的环 境 因 素 。另 外 , 甲壳 动 物 所 特 有
摘 要 性 别 决 定 (e eemiain 涉 及 遗 传 、 育 和进 化 等 领 域 , sxdtr n t ) o 发 开展 性 别 决 定机 制 的 研 究对 认 识 动 物 性 分 化异 常 等 多 种 与 性 别 有 关 的疾 病 、 现 动 物 性 别 的 人 为 控 制 等都 有 着 重 大 的现 实 意 义 , 时还 可 为 生 实 同 物进 化 的 分 析 提 供 新 的 线 索 。 本 文 主 要 阐述 两 大性 别决 定基 因 家 族 (o 。 S x DMR 中 S x 。 MR T) o 9 D TI在 水 产 动
DMRT家族生物信息学分析及Dmrt4Dmrt6表达与功能初步研究的开题报告

DMRT家族生物信息学分析及Dmrt4Dmrt6表达与功能初步研究的开题报告一、研究背景DMRT (Doublesex/Mab-3 Related Transcription factor) 家族是一类高度保守的转录因子,在多个物种中都具有重要的调节性别发育的作用。
该家族在多种组织和器官中表达,并参与了多个生物学过程,如性别分化、生殖调节、胚胎发育、异型分化和肿瘤发生等。
DMRT家族成员的相互作用和相关的信号通路在不同物种和不同组织中存在重要的差异,这使得这个家族成为研究性别特异性的一个重要的调控因素。
Dmrt4和Dmrt6是DMRT家族的两个成员,在多种物种中都广泛表达,但其生物学功能尚不完全清楚。
研究表明,它们可能在配子形成、胚胎发育和生殖调节等过程中发挥重要作用。
二、研究目的1. 通过生物信息学分析,对不同物种中DMRT家族成员的进化关系、结构特征、功能功能特点进行分析。
2. 对在小鼠和人类组织中Dmrt4和Dmrt6的表达模式、时空分布进行初步探究,以及探讨它们可能的调控作用。
3. 通过建立Dmrt4和Dmrt6基因敲除模型,对其对生物学过程的影响进行初步分析。
三、研究方法1. 生物信息学分析:通过NCBI数据库的物种比对和序列比对,分析DMRT家族成员的进化关系、结构特征、功能功能特点。
2. 组织特异性实验:在小鼠和人类中,通过实时荧光定量 PCR 和免疫组织化学技术分析Dmrt4和Dmrt6在不同组织中的表达情况及时空分布特点。
3. 基因敲除实验:通过 CRISPR/Cas9技术建立Dmrt4和Dmrt6基因敲除小鼠模型,并通过形态学、生殖细胞调查和生殖激素水平检测等方法分析基因敲除对小鼠生物学特征的影响。
四、研究意义通过对DMRT家族成员Dmrt4和Dmrt6的生物信息学分析和实验研究,可以进一步了解它们在生殖发育和生殖调节等方面的作用,为解决某些生殖疾病的分子机制提供新的思路。
此外,本研究还将为揭示生物性别特异性的分子调控机制提供有益的信息和启示。
大绿蛙Sox基因和Dmrt基因的序列分析

g n ssa d s x d tr n t n . h e h i u fP R r s d i h sp p rt mp i h o e e a d Dmr g n f e e i n e - e e mi a i o T e tc nq e o C we e u e n t i a e o a l y t e S x g n n f t e eo
p p rs p l s mo e u a a a frsu yn h e — e e i i g me h n s o a a l i a a d t e e ou in o o e e a e u p i l c l rd t o t d i g t e s x d t r n n c a im fR n i d n h v l t fS x g n e m v o
Abs r c Th o e efmi n tg n mi c o a t oe nted v lp n rc se f al mb y - t a t: eS x g n l a dDmr e ef l a ti r n lso h e eo me t o e s so r e ro a y a y mp t r p e y
陈启 龙 ,马 文 丽 h , 文 岭 郑 ,姚 文娟 聂 刘 旺 ,
( . 海 大学电子生物技 术 中心 ,上海 20 7 ; . 山学院生物 系,安徽 黄 山 2 5 2 ; 1上 00 2 2黄 4 0 1
性别决定相关基因SOX9研究进展

性别决定相关基因SOX9研究进展
侯林;崔英霞;姚兵
【期刊名称】《中国优生与遗传杂志》
【年(卷),期】2007(15)8
【摘要】SOX9是一种位于17号常染色体上的基因,其基因表达蛋白SOX9包含SRY样HMG框序列,它们在骨骼形成和睾丸发育调控中扮演着重要角色。
本文总述了SOX9基因及SOX9蛋白在个体发育中的调控作用。
【总页数】3页(P1-2)
【关键词】SOX9;骨骼;性别决定
【作者】侯林;崔英霞;姚兵
【作者单位】南京军区南京总医院生殖遗传研究室
【正文语种】中文
【中图分类】R394
【相关文献】
1.鱼类性别决定及分化相关基因研究进展 [J], 路畅;苏利娜;朱邦科
2.蜜蜂性别决定相关基因的研究进展 [J], 李淑云;王子龙;颜伟玉
3.哺乳动物性别决定相关基因研究进展 [J], 廖勤丰;何航;陈亚强;杨延辉;李文娟;程文超
4.哺乳动物性别决定及相关基因研究进展 [J], 谢晓刚;贾燕青;仇薪鑫;罗艳;张振仓;李丹;马乃祥;权富生
5.硬骨鱼类性别决定与分化相关基因研究进展 [J], 李永婧;吴利敏;李学军
因版权原因,仅展示原文概要,查看原文内容请购买。
SOX9基因与人类性别分化

SOX9基因与人类性别分化
朱敏;刘智
【期刊名称】《国外医学:遗传学分册》
【年(卷),期】1999(022)001
【摘要】近年来的研究表明SOX9基因同时参与了控制胚胎期骨骼发育和性别分化的过程,本文拟就SOX9基因的研究进展及其与人类性别分化的关系作简要综述。
【总页数】5页(P14-18)
【作者】朱敏;刘智
【作者单位】湖南医科大学分子生物学研究中心;湖南医科大学分子生物学研究中心
【正文语种】中文
【中图分类】Q344.2
【相关文献】
1.ApoE基因与人类老年性疾病的关系及ApoE基因敲除小鼠在人类老年疾病研究中的应用 [J], 程春华;杨继红
2.人类基因组多样性(HGD)与人类基因组多样性计划(HGDP)(2) [J],
3.人类基因组多样性(HGD)与人类基因组多样性计划(HGDP)(1) [J],
4.人类基因组多样性(HGD)与人类基因组多样性计划(HGDP)(2) DNA、RNA和蛋白质研究的有关资料 [J],
5.人类基因组多样性(HGD)与人类基因组多样性计划(HGDP)(2) 小核糖核酸(miero-RNA)和RNA干扰(RNAi) [J],
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
SOX 、DMRT 性别决定基因家族及其应用研究进展*李楠1,2王秀利1**仇雪梅1(1大连水产学院生命科学与技术学院,大连 116023;2黑龙江八一农垦大学生命科学技术学院,大庆 163319)摘 要: SO X 基因家族是在动物中发现的一类新的编码转录因子的基因家族,其产物具有一个HM G 基序保守区,参与诸如性别决定等多种早期胚胎发育过程。
到目前为止,在XX -XY 染色体性别决定系统中,只发现了两个性别决定基因:一个是SRY ,它主要在哺乳动物性别决定中起作用;一个是DM Y,它是在青鳉(Or y z ias latip es )中发现的。
SRY 属于SOX 基因家族,而DM Y 则属于另一个普遍参与脊椎动物性别决定过程的D M T R 基因家族。
本文综述了这两大性别决定基因的研究进展,并探讨了它们在水产养殖动物性别决定基因研究中的意义和价值。
关键词: 性别控制 SO X 基因 DM R T 基因 动物 水生动物The Research Advances and Applications of SOX andDMRT Gene FamilyLi Nan 1,2 Wang Xiuli 1 Qiu Xuemei 1(1College of L if e scie nce and Biotec hnology ,Dalian Fisher ie s Unive rsity ,Dalian 116023;2Colleg e of L if e science and Biotechnolog y ,H eilongj iang A ug ust First L an d Rec lamation Univ er sity ,Daqing 163319)Abstract: T he Sox g ene family of transcriptio n factor s ar e fo und thro ug hout t he animal kingdom.T hey ar e char acter ized by the presence of a H M G domain,involved in the r egulation of such diverse dev elo pmental pro cesses of early embry o -g enesis as sex deter minat ion.In the XX -XY sex determining chro mosomal sy stem,there are only tow sex deter mined gene has been found:one is the SRY g ene,which play an im po rtant ro le in mammals;the ot her is the D M Y g ene,which has been found in Or y z ias latip es .T he SR Y g ene belonged to the SO X g ene fam ily ,and DM Y belonged t o the DM T R gene family,w hich is inv olv ed in sex det erminatio n of v ertebrates.T his r eview mainly discusses the research trends and development o f SOX gene family,and DM T R gene family,and po int out the re -search value in searching for sex deter mined g ene in aquatic animals.Key words : Sex -determining co nt rol SOX DM RT A nimal Aquat ic animal动物的性别控制是指通过对精子或胚胎的性别进行鉴定,从而达到调控子代性别的目的。
随着胚胎冷冻、胚胎切割、体细胞克隆技术的日渐成熟,使得动物的性别控制显得更为重要。
多少年来人们一直在不停地探索如何提高动物性别控制的准确性。
随着科学技术的发展,人们不仅在细胞水平上发现了高等动物的性别决定于性别分化过程,而且还通过染色体的组型和核型分析,找到了与性别紧密相关的染色体)))性染色体。
分子遗传学原理和分子生物技术的飞速发展,使得人们在基因水平上研究动物的性别控制有了可能。
如果我们掌握了某些动物的性别决定基因,利用现代分子标记技术,便可极大的提高性别控制的准确性和有效性。
1990年哺乳动物性别决定基因SRY 的发现[1],是哺乳动物性别控制领域的一项重大突破。
近年来人们又发现和确定了一些与哺乳动物性别决定相关的基因(DM -RT 1、DAX1、WT1等),并提出几种基于哺乳动物性别决定基因的分子调控模型。
人们在研究和应用哺收稿日期:2005-04-07 *基金项目:大连水产学院科研基金项目**通讯作者:王秀利(1964-),男,博士,教授。
E -mail:xiuliw ang417@sin 生物技术通报#综述与专论#BIOTECHNOLOGY BU LLETIN2005年第3期乳动物性别决定基因的同时,也致力于探索水生动物的性别决定基因,但到目前为止,仅在青鳉(Or y-z ias latip es)中发现了鱼类性别决定基因DM Y。
在已知的与哺乳动物和脊椎动物性别决定有关的基因中,以SOX基因家族和DM RT基因家族最具有代表性。
1SOX基因家族1.1SRY基因Sinclair等人[1](1990年)在人类和小鼠中克隆了睾丸决定因子SRY(sex determining region on Y chrom oso me,SRY)基因,进一步研究发现SRY基因产物含有一段高迁移率蛋白质群(hig h mobility gro up,H M G),并发现它具有高度保守性[1][2]。
根据该H M G的保守性,发现了一类新的编码转录因子的SOX(SRY-type H MG box,SOX)基因家族[2]。
目前,人们仅在哺乳动物中,就发现了30多种SOX基因,它们广泛参与诸如性别决定、骨组织的发育、血细胞生成、神经系统的发育、晶状体的发育等多种早期胚胎发育过程。
属于SOX基因家族的众多已知基因中,目前只有SRY基因的被确定为哺乳动物性别决定基因。
研究发现X染色体上的SOX3基因是与SRY基因相似程度最高的基因,一般认为SRY基因与SOX3基因原本是同一基因的不同等位基因,只是在进化过程中SRY基因获得了性决定的功能[3],而SOX3的表达则与性别决定无关。
Koichiro等[4](2004)通过对老鼠胚胎性别分化过程中SRY表达量和SRY的5c端甲基化程度的研究,认为在老鼠的性腺发育过程中,机体可以利用DNA甲基化的方式调控SRY基因表达。
哺乳动物性别决定和性别分化依靠于Y染色体上的SRY基因。
SRY基因的瞬间表达产物能激发支持细胞分化和使双功能的胚胎性腺发育成睾丸。
1.2哺乳动物性别决定的其他SOX基因SOX9基因是与哺乳动物性别决定直接相关的基因,SOX9在XY雄性鼠胚胎的睾丸形成中表达,而在雌鼠中不表达[5],这表明SOX9基因在其性别决定过程中起作用。
该基因可能是靠近SRY基因的下游基因,并且与人类的睾丸生成过程有关。
人的SOX9基因位于17q上,在骨骼发育和性别决定中都有作用,它的突变可导致骨骼形成异常综合症(Compom dic Dysplasia,CD)和性反转现象,患有CD 的XY性染色体型的患者多数为性反转女性[6]。
目前,一般认为,SOX9基因既是CD基因,参与胚胎早期骨骼形成和发育的过程,同时又是一个常染色体上与性别决定有关的基因,参与调控睾丸发育过程。
近来有研究表明SOX8可以促进SOX9在睾丸形成中的决定作用。
人的SOX8基因(5.4kb)位于16p13.3上,编码一个466个氨基酸组成的蛋白[7]。
SOX8是哺乳动物睾丸决定途径中的一个关键基因,与SOX9一样也在性别决定时期的鼠的睾丸中表达[8,9]。
目前已经证实SOX8能部分补偿骨骼形成异常综合症中已经削弱的SOX9活性[9]。
有人预言在许多物种中当SOX9不表达或表达的太晚,以至于不能参与性别决定和调节米勒管抑制因子(M u-l lerian inhibitor y substance,M IS)表达时,SOX8可能是SOX9的替代物[8,9]。
SOX8在性别决定及其他发育方面的许多重要作用,仍需要我们不断的去探索。
除了SRY、SOX8和SOX9的表达与哺乳动物性别决定有关外,还有一些SOX基因在性腺的生殖细胞中表达,如SOX5、SOX6在小鼠的减数分裂后的生殖细胞中表达,尤其是在精细胞阶段[10]。
此外,爬行类温度依赖型性别决定(Temperature-depend-ant sex determ ination,TSD)的海龟中,也发现SOX9参与其T SD过程[11]。
由于SOX基因广泛参与动物的性别决定和性别分化等多种早期胚胎发育过程,所以人们在鱼类的性别决定基因研究中,不得不十分重视它。
1.3鱼类SOX基因的探索与研究鱼类,是良好的研究细胞发育的试验模型,同时也是遗传研究上不可多得的好材料。
鱼类存在着复杂的性别控制机制(XX-XY,ZZ-ZW,X1X2Y, XY1Y2和ZW1W2性染色体系统)[12]。
鱼类性别控制的研究,对水产养殖来说,具有重大的实用价值和意义。
因为许多养殖鱼类其生物学性状诸如生长率、成熟年龄、繁殖方式、体色、体型、个体大小等雌雄鱼之间存在较大的差异。
由于SOX基因家族在哺乳动物性别决定中独特的地位,所以,人们在探索鱼类性别决定基因的过6生物技术通报Biotechnology Bulletin2005年第3期程中,首先考虑到的就是鱼类中是否在也存在像SRY、SOX9这样的SOX基因。
如果鱼类中存在这类基因,那么它们的作用又是如何。
为了解决这些疑问,人们做了大量的研究工作,但结果并不是十分理想。
现已研究证实在一些鱼类中确实存在SRY 基因[13]、SOX9基因[14],但其并不具备性别专一性表达的特性。
有趣的是,用northern blot和原位杂交分析青鳉中的SOX9,发现它主要在成年鱼的卵巢中表达[14]。
然而,在精巢中,仅能用RT-PCR法检测到,这说明在鱼鳉中的性腺发育中,SOX9有着与其它脊椎动物的不同作用[14]。