第十一章 花朵大小的遗传
园林植物遗传育种学各章问答题

第一章种质资源试题五、简答题五、简答题1、阐述引种驯化的基本原理和引种驯化的基本原理:(1)、遗传学原理:简单引种:适应范围内引种;驯化引种:适应范围以外引种( 2)、生态学原理:气候相似论、主导生态因子(3)、生态历史分析理论限制引种成功的主导生态因子:(1)温度;(2)降水与湿度;(3)光照;(4)土壤;(5)生物因子*2、简述引种驯化的程序和步骤程序:(1)引种材料的收集(2)种苗检疫(3)登记编号(4)引种试验(5)推广试验步骤:3、简述引种成功的标准(1)、引种植物在引种地与原产地比较,不需要特殊的保护、能露地越冬或越夏,并生长良好。
(2)、没有降低原来的经济价值或观赏品质。
(3)、能够用原来的繁殖方法进行正常的繁殖。
(4)、没有明显或致命的病虫害。
(5)、对引种地不能构成生物入侵。
六、论述题2、在引种过程中应采取的方法有那些?(1)引种要结合选择(2)引种要结合有性杂交(3)选择多种地利条件实验(4)阶段训练与多代连续培育(5)试管苗培育(6)栽培技术研究第三章选种试题五、简答题3、简述芽变选择的程序和步骤.初选---目测预选。
A、对变异不明显或不稳定的都要继续观察,如果芽变范围太小,不足以进行分析鉴定,可通过修剪、嫁接等措施,使变异部分迅速增大,而后再进行分析鉴定。
B、对于变异性状十分优良,但不能证明是否为芽变的,可先将无性繁殖后代栽入移植鉴定圃再根据表现决定下一步骤。
C、对于有充分证据肯定为芽变的,且性状优良,但是还有些性状不是十分了解,可以不经移植鉴定圃进入选种圃。
D、对于有充分证据证明是十分优良的芽变,且没有相关劣变,可不经鉴定圃与选种圃,直接参加复选。
E、对于嵌合体形式的芽变,可采取修剪、嫁接、组织培养等方法,使变异性达到纯化和稳定。
2).复选---对初选芽变再次进行评选。
主要工作:a、对初选植株进行评选,繁殖成营养系;b、在选种圃中进行比较,也可结合生产试验进行复选出优良单株。
园林植物遗传育种课件:园林植物性状遗传

花径、重瓣性与数量性状
花的直径表现为数量性状,重瓣性在大多数情下也 表现为数量性状,其遗传遵循数量性状的遗传规律。
如:牡丹、菊花等。
返回 目录
增加花径的途径
花径是数量性状,其遗传遵循数量性状的遗传机理, 利用这一属性,通常采用下列方法来增加花径。
❖ 改进栽培条件。充足的水肥,适当的管理及精心培育能使 花径变得更大。
C类突变体表型示意图
返回 目录
花型的发育 花型的发育受基因控制。
返回 目录
第二节 花色与彩斑的遗传
1.花色的形成 2.花色遗传 3.彩斑的遗传
屈金香
兰花
目录
花色的形成
花色的形成与花部所含色素种类和花被片海棉组织层的 物理特性有关,而主要受所含色素种类的影响。
花的色素有三大类群:类胡萝卜素、类黄酮、花青素。每 一类色素均包括很多种类。植物的不同花色是由不同的色素组 成的。
返回 目录
第四节 株型遗传
1.株型的多样性 2.植物激素与株型 3.株型的遗传
目录
株型的多样性
❖ 按植物的分枝特性,可分为乔木、灌木、藤本。 ❖ 按植株的高矮, 有乔化、矮化之分。 ❖ 按枝条的姿势可分为直枝、垂枝、曲枝。
如将上述因子组合则有十几种株型。
返回 目录
植物激素与株型
植物的株型,主要与其自身激素平衡有关。 1.分支特性 乔、灌、藤之分主要是顶端优势的作用,而顶端
园林植物遗传育种学
目录
园林植物性状遗传
第一节 花的发育 第二节 花色与彩斑的遗传 第三节 花径与重瓣性遗传 第四节 株型遗传 第五节 抗性遗传
首页 结束
第一节 花的发育
植物的发育从胚开始: 地下部分(根) ← 根尖 ← 胚 → 茎尖 → 地上部分(茎、叶、花)
园林植物遗传育种ppt课件

• 引种驯化〔introduction and domestication〕 就是把一种植物从现有的分布区域〔野生 植物〕或栽培区域〔栽培植物〕人为地迁 移到其他地域种植的过程。
• 8.1.2 引种驯化的意义
• 〔1〕引种可以迅速丰富和改善本地种类的 构造。
• 〔2〕引种省时、省力,可以迅速提高经济 效益
• 1〕控制花样的有关基因
• 〔1〕花样素基因
• 〔2〕控制花样素含量基因
• 〔3〕花样素分布基因
• 〔4〕助色素基因 助色素单独含于细胞中 时几乎无色,但它与花青素同时存在于细 胞中构成一种复合体时,就呈蓝色,与花 青素本来的颜色完全不同,这种复合体是 产生蓝色花的重要缘由。〔图6.1〕。
• 〔图6.2〕
• 一个种类引到新地域后,由于气候条件、 耕作制度与原产地不同,引入后能够有不 同的表现,所以必需进展引种实验。在实 验时,要求用当地有代表性的优良种类作 对照,同时实验地的土壤条件必需均匀, 管理措施力求一致,使引种资料能得到客 观的评价。步骤如下:
• 〔3〕引可以充实育种的基因资源,为其 他育种途径提供育种资料
• 8.1.3 我国引种驯化的概略
• 我国园林植物的引种历史悠久,早在汉代 张骞出使西域时,就引进了核桃、石榴、 葡萄、红花等经济作物。目前各大城市种 植较多的世界著名的行道树——悬铃木, 相传是晋代由鸠摩罗什引入陕西户县的。 我国幅员辽阔,土壤和气候条件非常复杂, 为引种任务提供了极为有利的条件,植物 引种有着宽广的前景。
• 2〕种质资源的利用
• 鉴定出具有优良性状的种质资料,可作为 亲本,经过杂交、人工诱变及其他手段发 明新的种质资源,为育种提供半废品,并 从其后代中选育出优良变异个体培育成新 种类;也可利用种质资源,直接从中选育 出优良个体培育废种类。目前,许多育种 家经过远缘杂交,将野生近缘植物的基因 导入植物种类,使其获得新的优良性状, 虽然有的不能直接利用与消费,但能够成 为有价值的育种资料。
备课素材:豌豆的花色遗传 高一下学期生物人教版必修2

豌豆的花色遗传先看一道试题:香豌豆的花色由两对独立遗传的等位基因B-b和R-r 控制,只有B、R同时存在时才表现为红花,其余表现为白花。
下列说法中错误的是()A.基因型为BbRR与bbrr的植株杂交,后代中红花植株与白花植株之比约为1:1B.基因型为BbRr的植株自交,后代中红花植株与白花植株之比约为9:7C.两个白花的品种杂交,后代不可能出现红花植株D.两亲本杂交,后代可能出现红花:白花为3:1或1:3的情况解析:基因型为BbRR与bbrr的植株杂交,后代中红花植株与白花植株之比约为BbRr:bbRr=1:1,A正确;基因型为BbRr的植株自交,后代中红花植株与白花植株之比约为9B R :(3B rr+3bbR +1bbrr)=9:7,B 正确;两个白花的品种杂交,后代也可能出现红花植株,如BBrr和bbRR杂交,C错误;基因型为BbRR与BbRR或BBRr与BBRr杂交,后代红花:白花为3:1;基因型为Bbrr与Bbrr或bbRr与bbRr杂交,后代红花:白花为1:3,D正确。
故答案为C。
这道试题以香豌豆的花色遗传为情境考查遗传规律。
豌豆的花色遗传遵循孟德尔分离定律,但是,花色的遗传有多种类型,基因也不一定是一对基因控制,由两对基因控制,甚至有三对基因控制。
因此,需要根据试题提供的情境去解决问题。
常见的花色遗传一般规律有哪些?1856~1863年孟德尔(G.J.Mendel)连续8年的豌豆杂交实验,奠定了花色遗传的理论基础。
他将纯系的紫花豌豆与白花豌豆杂交,F1全部开紫花。
F1自花授粉所得F2中,有705株开红花,224株开白花,紫花植株与白花植株之比接近于3∶1。
孟德尔称F1表现的红花性状为显性性状,白花性状为隐性性状;F2显性性状与隐性性状同时分别出现,称为性状分离。
1.花色1910~1930年德国学者威尔施泰特(Will-statter)和瑞士学者卡勒(Karrer)从大量生物中分离出结晶的类胡萝卜素,存在于植物的花瓣、根、叶和果实中,它包含红色、橙色及黄色,是一大类色素类群,胡萝卜、番茄、南瓜、柿子所特有的颜色,均为类胡萝卜素的产物。
花径与重瓣性遗传

对立异位多基因系统模式: 在一个对立异位多基因系统中,某些成员具有加 性作用,另一些有减性作用,这一数量值是这些对 立基因平衡作用的结果。
同源等位基因之间不存在显隐性关系
P1 P2 F1 AABBCC aabbcc AaBbCc 26-4-4=18 10-2-2=6 18-3-3=12
第一种超亲现象。
A为显性,B、C基因之间不存在显隐性关系 P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 6+4+4=14
A 、B为显性,C基因不存在显隐性关系
P1 P2 F1 AABBCC aabbcc AaBbCc 6+6+6=18 2+2+2=6 6+6+4=16
2.2重瓣花的起源
• • • • 积累起源的重瓣花 雌蕊起源的重瓣花 花序起源的重瓣花 重复起源的重瓣花
1:2:1 1:4:6:4:1: 1:6:15:20:15:6:1 1:8:28:56:70210:252:210:120:45:10:1
0 1/64
1 6/64
2 15/64
3 20/64
4 15/64
5 6/64
6 1/64
1.3.3 多基因系统模式 等效异位多基因系统模式: 同源等位基因之间不存在显隐性关系 P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 4+4+4=12
A为显性,B、C基因之间不存在显隐性关系 P1 AABBCC 26-4-4=18 P2 aabbcc 10-2-2=6 F1 AaBbCc 26-3-3=20
矮牵牛花朵大小及相关性状遗传分析

河南农业科学,2024,53(2):118‐127Journal of Henan Agricultural Sciencesdoi:10.15933/ki.1004-3268.2024.02.013矮牵牛花朵大小及相关性状遗传分析张林霞1,2,张蔚1,2,张书婷3,孙苗苗1,张晓敏1,李志能2,刘国锋1(1.广州市林业和园林科学研究院,广东广州510405;2.西南大学园艺园林学院,重庆400715;3.湖北省农业科学院经济作物研究所,湖北武汉430064)摘要:为了探究矮牵牛花朵大小的遗传规律,以大花型和小花型矮牵牛高代自交系为亲本构建四世代遗传群体(P 1、P 2、F 1、F 2),对花朵大小遗传特征进行主基因+多基因混合遗传模型分析,并将F 1植株与中花型矮牵牛W115株系进行杂交,验证遗传规律。
同时以F 2群体为材料,对花径、萼片长、叶片长等23个表型性状进行测定,并研究其相关性。
结果表明,矮牵牛大花对小花性状符合2MG-A 模型,即由2对加性主基因控制,主基因遗传率为95.38%;大、小花杂交F 1与中花W115进一步杂交,后代出现大花与中花性状分离(1∶1),且中花植株的叶片和苞片叶绿素含量显著高于大花植株(P <0.01)。
大花×小花F 2群体的表型性状变异丰富,变异系数在7.67%~59.93%,平均22.38%。
相关性分析结果表明,花部性状、叶部性状以及两者之间均存在一定的相关性,其中花径与其他器官大小均呈显著正相关,与部分植株性状呈显著负相关。
关键词:矮牵牛;花朵大小;遗传分析;相关性分析中图分类号:S681.6文献标志码:A文章编号:1004-3268(2024)02-0118-10收稿日期:2023-11-20基金项目:广东省自然科学基金-面上项目(2019A1515011840);广州市科技计划项目(穗财编[2023]1号)作者简介:张林霞(1996-),女,重庆人,硕士,主要从事园林植物遗传育种与生物技术研究。
孟德尔遗传定律的拓展及解题学习资料

长沙市一中 甘伟丰
基因分离定律的拓展及解题
【例1】紫茉莉的红花受A基因控制,白花受a基因控制,A、a是
一对等位基因。用开红花的紫茉莉品种与开白花的紫茉莉品种
杂交,F1全为粉红花。请回答: (1)F1自交,F2的表现型及比例为红花:粉红花:白花=1:2:1 。
(2)若让F2中的全部红花植物和粉红花植物群体内相互授粉,
个体相互授粉(杂交) AA与Aa构成的群体内
相互授粉:4/9 个体自花授粉(自交):1/2
基因分离定律的拓展及解题
【例1】紫茉莉的红花受A基因控制,白花受a基因控制,A、a是
一对等位基因。用开红花的紫茉莉品种与开白花的紫茉莉品种
杂交,F1全为粉红花。请回答: (1)F1自交,F2的表现型及比例为红花:粉红花:白花=1:2:1 。
雌配子:XA 、 Xa
基因分离定律的拓展及解题
类型
表现规律或遗传要点
不完全显性 配子致死 个体致死
显性纯合子与杂合子表现的性状不同 包括:显性基因配子或隐性基因配子致死;
雄配子或雌配子致死 包括显性个体致死和隐性个体致死
基因分离定律的拓展及解题
【例3】喷瓜有雄株、雌株和两性植株,G基因决定雄株,g 基因决定两性植株,g-基因决定雌株。G对g、g-是显性,g 对g-是显性,如:Gg是雄株,gg-是两性植株,g-g-是雌株。
究小组完成的三组杂交实验结果:
第一组
第二组
第三组
绿色雌株 × 金黄色雄株 ↓
绿色雄株
绿色雌株 × 金黄色雄株 ↓
绿色雄株 金黄色雄株 1∶1
绿色雌株 × 绿色雄株 ↓
绿色雌 绿色雄 金黄色雄 2∶ 1∶ 1
问:第一、二组没有出现雌株的最合理的解释是
园艺植物育种学:5 观赏植物主要性状的遗传

4-香豆酸盐 4-Coumarete
柚配质(黄酮 Naringenin
图1 花色素苷合成途径
3 花色遗传受一系列基因的作用和控制
花色素基因、花色素量的基因、花色素的分布基因、助色素基因和控制花瓣内部酸度的基因等;易变基因和基因的转座:常常造成花序或花朵上形成异质条纹、斑块(彩斑);不同花色杂交的显隐性(质量性状基因、基因互作)一般,带色花显性,白色花隐性;紫色花显性,红色花隐性;蓝色花显性,紫色花隐性。亦有例外。
毛华菊花朵直径大小的遗传变异
(三)增加花径的途径
改进栽培条件;倍性育种;增加花朵重瓣性;定向选择。
二、花重瓣性的遗传
1 重瓣花的形态起源
(一)概念:花朵重瓣性指观赏植物花瓣数量的多少。(二)重瓣花的遗传积累起源 雌雄蕊起源
花序起源重复起源(套筒起源)突变起源台阁起源
木槿
芙蓉
雌雄蕊起源
山茶雌雄蕊起源
观赏植物主要性状的遗传
花色彩斑花径与重瓣性
观赏植物主要性状
花
株型抗性
第一节花的发育
花是观赏植物的主要观赏器官,千奇百怪、万紫千红!植物学:植物的完全花是由花萼、花瓣、雄蕊、雌蕊等四轮构成的生殖器官。植物生理学:成年植物花的诱导需要一定的光、温周期,如二年生花卉大多需要经过低温的春化作用才能开花,多数菊花需要短日照处理才能开花。
仙客来
裂叶牵牛
百合
虞美人(罂粟科罂粟属)的美丽花边
花肋:沿中脉方向具放射性彩色条纹
紫脉吊钟
(二)不规则彩斑的遗传
花嵌合体、彩斑
叶部彩斑(“花叶”),变色叶
果实彩斑
2遗传机制
1常见类型
核内
核外
大象版科学六年级上册植物的遗传和变异课件

大象版科学六年级上册课件01新知讲解 Nhomakorabea 提出问题
植物是否也 有遗传和变 异的现象呢?
作出假设
植物也有像人那样的的遗传和变异。
同一种花有 许多种颜色
……
同一棵树上 的叶子不同
调查
实地观察,在同 类植物上能不能 找到两片完全相 同的叶子。
尚未成熟的植物个体通 常具有植株低矮、叶片 嫩且颜色浅等特征,这 些是生长发育的问题。
亲代
亲代芦荟 叶片形状……
叶片边缘 有刺、 绿色、 簇生……
子代芦 叶片形状
子代
事实证据
亲代凤仙花
播种凤仙花, 观察子代凤仙花 成熟后与亲代凤 仙花的异同。
亲代凤仙花 和子代凤仙花叶 片形态差不多, 都是互生。只是 红花更多了。
杂交水稻就是选用在遗传上有一 定差异,同时它们的优良性状又能互 补的水稻品种进行杂交,培育出同时 具有两种水稻优势的新品种。
杂交优势是生物界的普遍现象, 利用杂交优势提高农作物产量和品 质是现代农业科学的主要成就之一, 袁隆平是中国杂交水稻育种专家, 被誉为“杂交水稻之父”。
小麦的杂交
03
课堂讨论
得出结论
植物子代与亲代非常相似, 但也有一些细微的不同,这是 遗传和变异共同作用的结果。
04
延伸拓展
一、判断题
√ √ √ √ ×
第十二章 花朵大小的遗传040628

第十二章花朵大小的遗传第一节增加花朵直径的途径一、栽培措施的作用二、增加花朵直径的遗传学途径增加花朵直径的途径是指创始性的途径:1.诱发多倍体2.诱发突变3.增加重瓣性4.发掘多基因系统的潜力图12-1 Nicotiana longiflora的花径遗传(East,1916)第二节花朵直径与多基因系统伊斯特(East,1916)曾用两种花朵大小不同的烟草自交系(Nicotiana longiflora)做杂交。
其中一个亲本类型的花为长筒状,另一个为短筒状(图12-1)。
花朵大小这一性状在烟草中具有相对较高的遗传力,他对各代群体的花冠长度做了详细记录,并按照每3mm为一组进行分类。
数据列于表12-1。
在上述试验中,大花和小花类型的杂交,产生了较大的F1和F2群体。
从表现型不同的若干F2植株又产生了若干不同的F3家系,其中有些家系还一直追踪到F4和F5。
象前面描述的做法一样,每个家系和其中的个体的花朵直径都做了记载和分组,画出它们的频率分布图。
表12-1 Nicotinana Longiflora品种间杂种花冠长度的遗传(Grant 1975,from East)所有这些特点都证明品种间杂交花朵大小的遗传方式符合前面所说的数量性状的遗传规律,既花径遗传是受微效多基因系统控制的。
表12-2 种间杂种Nicotiana forgetiana×N.alata花冠长度的频率分布(Grant 1975 from East)表12-3 杂交实验中花冠长度变异的数据分析根据Grant(1975)的总结,花朵大小的遗传符合多基因假说,其要点主要如下:1.当两个花朵大小不同的自交系(或纯系)亲本互相杂交时,杂种F1群体的花朵表现型应当是整齐一致的,即不发生分离;2.F2群体的变异幅度远远大于F1的变异幅度;3.亲本类型应当在F2群体中重新出现。
如果在F2中未出现,应该在随后的后代中出现,如F3、F4等;4.在F2群体的频率分布曲线上不同点的个体所产生的相应的F3群体,应当表现出显著不同的花径平均值;5.F2中不同个体产生的F3群体具有大小不同的变异范围;6.F2以后各代家系的变异范围,应当以F2亲本的数量值为中心;7.在F2以后的各世代中,任何家系的遗传力可以小于但不会大于产生它的那个群体的遗传力。
植物花发育和花色的遗传学研究

植物花发育和花色的遗传学研究随着科技的不断进步,生物学领域的研究也在不断地深化,其中植物花发育和花色的遗传学研究成为了热点话题。
本文将从植物花的发育过程入手,探讨花色遗传机制的研究,以及对遗传学研究的未来展望。
一、植物花的发育过程植物的花是植物体中的生殖器官,产生后代的重要场所。
在植物的生长发育过程中,花的形成被称为花发育,是一个复杂的过程。
花发育分为各个阶段:萌芽期、原基分化期、萼片期、花瓣期、雄蕊期、雌蕊期和果实期。
在每个阶段,不同的基因负责不同的发育程序。
萌芽期,植物生长点经过物质的影响,形成原芽,原芽先是扩大,之后从花蕾基部膨胀,形成苞片和花轴。
而在原基分化期,根据不同植物的特点,原基逐渐分化成不同的花部构造,如花萼、花瓣、雄蕊和雌蕊等。
花发育与花色的具体形成过程,实际上是一个基因调控的过程,影响多个生长因子,同时还与环境、植物内分泌和激素等生理过程有关。
二、花色遗传机制的研究植物花色是受到基因调控的,多数与花瓣颜色相关的遗传因子都位于花革属中,在研究花色遗传机制时,科学家往往先通过遗传分析的方法来确定染色体区域,以及探究某一个或多个全能基因对于花色的调控作用。
花色基因分类上分为革兰氏算法和根据翻译后氨基酸序列的颜色序列树方法。
1、花色素生物合成途径的研究1939年Grierson指出,植物花色与生物合成途径相关的酶和代谢物有关。
目前研究中发现,花色素生物合成途径涉及到多个生物化学反应,这些化学反应使花革素从前花革素B基质发生反应被激活然后转变成茜素酸化合物后再次被激活,最后形成不同的花色素分子。
例如在向日葵中,花色素生成途径中的96%来源于外接源,8个花色素基因分别对应着4种花色素,每一种花色素都有一个可变的基因和一个基本的合成酶基因。
3’-羟基茜色素有一定的特殊性质,可以转化成不同的涂层,从而形成不同的花色素,这是花色基因控制了花瓣颜色的原因。
在花色素生物合成途径上,具有花色素生成反应的酶,如黄黴素氧化还原酶(flavonoid oxidase/reductase)和花革素-双苯酮氧化脱氢酶(anthocyanidin synthase,ANS),是维持花朵颜色的主要因素之一。
花发育的遗传调控

• 2)春化作用对成花的影响和机理
• 春化作用:低温诱导或促进植物成花的效 应
• 春化处理可诱导某些基因的启动或关闭, 最终诱导花的形成。 • 春化基因的分离
2.2内部因子对成花的影响 叶对开花有重要影响。促进开花的物质在叶中 产生并运输到芽中,使叶芽转变为花芽。 2.3生长调节物质对开花的影响 IAA等生长素在短日下大幅度抑制菊花开花 赤霉素可使一些需冷植物在常温下抽苔开花, 使一些短日植物在长日下开花。 激素的平衡变化可导致与成花有关的基因解除 阻遏 成花诱导由自身遗传因子和外界环境因素决定, 是开花基因在时间和空间上顺序表达的结果。 • 1)植物应对外界环境信号,开花时间基因控制 从营养生长转向生殖生长。 • 2)外界环境信号激活花分生组织决定基因。 • 3)花芽分生组织决定基因活化不同区域的花器 官决定基因 • 4)花器官决定基因激活下游的器官构造基因, 形成各轮花器官。
3、花转变的顺序和基因对成花的控制
3.1植物开花的遗传调控
1)开花时间决定基因
在植物开花过程中有一类基因可以不受日照长度的影响,它们的 表达足够触发开花,这一类基因被称为开花时间决定基因 CO是从拟南芥中分离出的开花时间决定基因. CO基因的表达可足 够触发开花
2).花序分生组织决定基因
EMF\TFL1、CEN基因均与花序发育有关,决定了顶端分生组织的属 性 EMF抑制生殖生长,使营养生长得到保持,其突变体emf失去了这 种能力 TFL1早期抑制、后期维持花序发育,其突变体tfl1叶片减少,花序 提前,无限花序变为有限花序。 CEN突变体cen花芽减少,顶端成花。
• ABCD模型 • ABCDE模型 • 四因子模型
4、成花逆转现象
成花逆转:植物完成成花诱导,其顶端分 生组织开始稳定的生殖生长,但如果环境 条件不适应,植物可能从生殖生长逆转回 营养生长。 类型: 1)花逆转 2)花序逆转 3)部分开花
21.2植物的有性生殖课件2023--2024学年苏教版生物八年级下册
莲蓬
植物的有性生殖
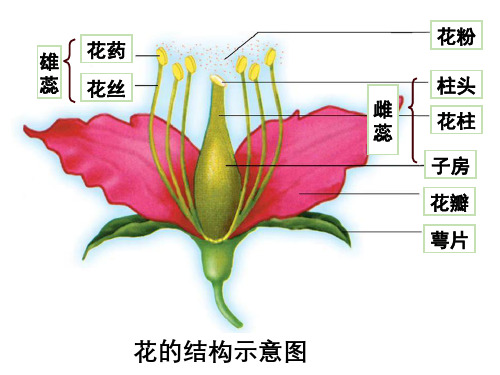
一、开花与结果
柱头 凋落
雌蕊 花柱 子房壁
发育成
果皮 果
花
子房 胚珠
珠被 卵细胞
种皮 胚
种子 实
雄蕊 花粉(精子) 花丝
花瓣、花柄、花托
(3)其他动物
花粉进入柱头后发生 了什么?
子房
纵切放大
子房的结构
子房壁
珠被 珠心(内有1个
卵细胞和2个极核)
胚 珠
极核
卵细胞 珠孔
受精的过程
当花粉落到雌蕊的柱头上,受到柱头 分泌的黏液的刺激,花粉就会萌发出 花粉管。花粉管穿过柱头、花柱,直 达子房里的胚珠。从珠孔进入胚珠后 ,花粉管末端破裂,其中的精子释放 出来,与卵细胞结合成为受精卵。 植物的精子与卵细胞结合形成受精卵 的过程,叫做受精。受精卵是植物体 发育的起点。 植物的有性生殖:由两性生殖细胞的 结合,成为受精卵,发育成新个体。
自花传粉
同一朵花的雄蕊花药中的花粉传给雌蕊
异花传粉
一朵花的雄蕊花药中花粉传给另一朵花的雌蕊
传粉的媒介
(1)风 风媒花
花小,花瓣颜色不鲜艳,无香味,花粉 量多而轻,容易被风吹走。花的柱头大,常 分叉,带有黏液,易接受花粉。
杨树
水稻
玉米
(2)昆虫 虫媒花
一般较大,花瓣具有鲜艳的颜色,有香味 或甜美的花蜜。花粉量少而有粘性。子房 的基部有蜜腺,能产生花蜜。
花的哪些结构发育成 果实和种子?
二、果实和种子的形成
➢受精作用完成后,花萼、花冠、雄蕊 以及雌蕊的柱头和花柱一般都会逐渐凋 落,只有雌蕊的子房发育成果实。
果实的各部分结构分别是由子房 的哪一部分发育而成的?
花发育成果实和种子
花蕾到花朵的变化过程

花蕾到花朵的变化过程一、花蕾的形成和发育花蕾是指花的未打开的状态,它是由植物的生长点包裹在保护鳞片中形成的。
花蕾形成的过程称为花蕾发育,这个过程受到环境、激素和基因等因素的调控。
1.分生组织:花蕾的发育始于植物的分生组织,特别是位于茎尖的分生锥。
分生锥中的分生细胞持续不断地分裂,从而形成新的组织和器官,其中就包括未来的花蕾。
2.包被鳞片:在花蕾的形成过程中,花蕾细胞逐渐增多并伸长,最后形成一个原始花蕾。
同时,茎的外部包被形成保护花蕾的鳞片。
鳞片是由茎叶的修饰器官演化而来的,其主要功能是保护花蕾及其内部组织。
3.花蕾分化:原始花蕾内部的细胞会进一步分化成不同的组织和器官,比如雄蕊(花药和花丝)、雌蕊(子房、花柱和柱头)以及花瓣和花萼等。
4.花蕾大小和形态的变化:花蕾的大小和形态会随着时间的推移而发生变化。
这一过程受到遗传和环境因素的影响,比如温度、光照等。
花蕾通常会在适宜的环境条件下迅速膨大,并逐渐展开。
二、花蕾的开放过程当花蕾达到一定的大小和成熟度时,它们会开始开放,形成鲜艳的花朵。
花蕾开放的过程涉及许多生物学和物理学机制。
1.花蕾壁的软化:在花蕾开放之前,花蕾壁中的细胞会发生生理和化学变化。
其中,细胞壁会变得柔软和薄弱,以促进花蕾的展开。
2.花蕾上的分离层:花蕾上通常存在一层称为分离层的细胞组织。
分离层中的细胞具有特殊的形态学和生理学特征,它们会在花蕾展开的过程中分离,从而促使花瓣和花萼的展开。
3.花蕾壁的撕裂:当分离层分离后,花瓣和花萼中的细胞会膨胀,使花蕾壁撕裂。
这个过程通常是由水分和激素的作用引起的。
4.花瓣和花萼的展开:一旦花蕾壁撕裂,花瓣和花萼中的细胞会快速吸水膨胀,从而使它们迅速张开。
花瓣和花萼的展开过程称为展花。
三、花朵的成熟和凋谢一旦花瓣和花萼展开,花朵会进一步发育和成熟。
成熟的花朵具有完整的雄蕊和雌蕊,能够吸引传粉媒介如昆虫或风力进行传粉。
1.雄蕊和雌蕊的生殖细胞发育:花朵的成熟过程包括了雄蕊和雌蕊内花粉和胚珠的发育。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
176个群体中出现一个亲本类型,因此 决定这一性状的基因约为3~4个。
(2) A为显性,B、C基因之间不存在显隐 性关系 P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 6+4+4=14
(3)A 、 B为显性,C基因不存在显隐性 关系 P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 6+6+4=16
三、多基因系统的作用机理
(一)多基因系统的组成 多基因系统的基因数 多基因的连锁关系 遗传力 基因互作 基因互作的相对强度 基因成员的作用方向 显性作用 等效和加性效应
多基因系统在F2群体中分离比理论值
等位基因 对的数量 1 2 3 4 5 比 率 分母数 4 16 64 256 1024
Gilia capitata 柱头长度的遗传
等位基因 对的数量 1 2 3 4 5
比
率
分母数
1:2:1 1:4:6:4:1: 1:6:15:20:15:6:1 1:8:28:56:70:56:28:8:1 1:10:45:120:210:252:210:120:45:10:1
4 16 64 256 1024
(4)三个基因全部表现显性
P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 6+6+4=18
等 效 异 位 多 基 因 系 统 模 式
2.
非等效异位多基因模式 是指具有加性效应但作用强度不等 的多基因,其中有些产生较强的效果, 另一些则较弱。
(1) 同源等位基因之间不存在显隐性关系
P1 AABBCC 9+4.5+4.5=18 P2 aabbcc 3+1.5+1.5=6 F1 AaBbCc 6+3+3=12
(2) A为显性,B、C基因之间不存在显隐 性关系 P1 AABBCC 9+4.5+4.5=18 P2 aabbcc 3+1.5+1.5=6 F1 AaBbCc 9+3+3=15
第十一章 花朵大小的遗传
一、增加花朵直径的途径
1. 栽培措施的作用 2. 增加花朵直径的遗传学途径 诱发多倍体 诱发突变 增加重瓣性 发掘多基因系统的潜力
二、花朵直径与多基因系统
伊斯特(East,1916)曾用两种花朵大小不 同的烟草自交系做杂交。其中一个亲本类型的 花为长筒状,另一个为短筒状。他对各代群体 的花冠长度作了详细记录并画出它们的频率分 布图。 这个试验证明品种间杂交花朵大小的遗ቤተ መጻሕፍቲ ባይዱ 方式符合前面所说的数量性状的遗传规律,及 花径遗传是受微效多基因系统控制的。
烟草花径的遗传
花朵大小遗传的多基因假说: Grint(1975)
当两个花朵大小不同的自交系(或纯系)亲本互相杂 交时,杂种F1群体的花朵表现型应当是整齐一致的, 即不发生分离; F2群体的变异幅度远远大于F1的变异幅度; 亲本类型应当在F2群体中重新出现。如果在F2中未出 现,应在随后的后代中出现; 在F2群体的频率分布曲线上不同点的个体所产生的相 应的F3群体,应当表现出显著不同的花径平均值; F2中不同个体产生的F3群体具有大小不同的变异范围; F2以后各代家系的变异范围,应当以F2亲本的数量值 为中心; 在F2以后的各世代中,任何家系的遗传力不会大于产 生它的那个群体的遗传力。
(3)A 、 B为显性,C基因不存在显 隐性关系 P1 AABBCC 9+4.5+4.5=18 P2 aabbcc 3+1.5+1.5=6 F1 AaBbCc 9+ 4.5 +3=16.5 (4) 当ABC均为显性时,群体明显偏向 显性亲本。
非 等 效 异 位 多 基 因 模 式
3.对立异位多基因系统模式 在一个对立异位多基因系统中,某些成 员具有加性作用,另一些有减性作用,这 一数量值是这些对立基因平衡作用的结果。
1:2:1 1:4:6:4:1: 1:6:15:20:15:6:1 1:8:28:56:70:56:28:8:1 1:10:45:120:210:252:210:120:45:10:1
0 1/64
1 6/64
2 15/64
3 20/64
4 15/64
5 6/64
6 1/64
(二) 多基因系统模式
1. 等效异位多基因系统模式 (1) 同源等位基因之间不存在显隐性关系 P1 AABBCC 6+6+6=18 P2 aabbcc 2+2+2=6 F1 AaBbCc 4+4+4=12
杂种优势现象。
(3)A 、 B为显性,C基因不存在显隐性 关系。 P1 AABBCC 26-4-4=18 P2 aabbcc 10-2-2=6 F1 AaBbCc 26-4-3=19 (4) 当ABC均为显性时,群体与显性亲本重 合。
对 立 异 位 多 基 因 系 统 模 式
四、多基因系统的鉴定
(1)同源等位基因之间不存在显隐性关系
P1 AABBCC 26-4-4=18 P2 aabbcc 10-2-2=6 F1 AaBbCc 18-3-3=12 第一种超亲现象。
(2) A为显性,B、C基因之间不存在显隐 性关系 P1 AABBCC 26-4-4=18 P2 aabbcc 10-2-2=6 F1 AaBbCc 26-3-3=20