分子生物学 第七章
分子生物学 第七章 蛋白质的生物合成

7.1 遗传密码 7.2 tRNA 7.3 氨酰-tRNA合成酶 7.4 核糖体 7.5 与蛋白质合成有关的因子 7.6 蛋白质合成的生物学机制 7.7 蛋白质转运机制
比DNA复制和转录更为复杂的过程
氨基酸活化与转运----这个过程是在氨基酸活化酶和镁 离子作用下把氨基酸激活成为活化氨基酸。 起始----核糖体与mRNA结合并与氨酰基tRNA生成起 始复合物。 延伸----由于核糖体沿mRNA5’端向3’端移动,开始了 从N端向C端的多肽合成,这是蛋白质合成过程中速度 最快的阶段。 终止以及肽链的释放,核糖体从mRNA上解离,准备 新一轮合成反应。
(3)校正tRNA 校正tRNA分为无义突变及错义突变校正。 无义突变:在蛋白质的结构基因中,一个核 苷酸的改变可能使代表某个氨基酸的密码 子变成终止密码子(UAG、UGA、 UAA),使蛋白质合成提前终止,合成无 功能的或无意义的多肽。
错义突变是由于结构基因中某个核苷酸
的变化使一种氨基酸的密码变成另一种 氨基酸的密码。 错义突变的校正tRNA通过反密码子区的 改变把正确的氨基酸加到肽链上,合成 正常的蛋白质。(摆动原理现象)
结合的AA-tRNA分开。
7.1.3 遗传密码的特点
⑴连续性
⑵简并性与偏爱性
⑶通用性与专一性
(特殊性)
⑷终止密码
⑸密码子与反密码子
简并性: 由一种以上密码子编码同一个氨基酸的现象。 18种氨基酸都有一个以上的密码子 。
同义密码子。
只有精氨酸是个
例外,因为在真 核生物中CG双联
7.2 tRNA
7.2.1 tRNA的结构 由于链内的碱基互 补配对,tRNA呈现
分子生物学第七章原核生物基因表达调控

原核生物基因表达调控的特点
01
原核生物基因表达调控通常由特 定的转录因子、RNA聚合酶以及 其他调控蛋白介导,通过与DNA 的结合或解离来调节基因转录。
02
原核生物基因表达调控具有快速 响应环境变化的特点,能够在短 时间内调整基因表达模式,以适 应外界刺激和压力。
翻译后加工的调控
翻译后加工的调控
在翻译后加工阶段,新合成的蛋白质经过一系列修饰和加工,最终成为具有生物学活性的蛋白质。原 核生物通过控制翻译后加工酶的合成和活性来调控翻译后加工过程。此外,原核生物还可以通过控制 蛋白质的稳定性来影响其功能和表达水平。
总结
翻译后加工是基因表达调控的重要环节,原核生物通过控制翻译后加工酶的合成和活性,以及蛋白质 的稳定性来精细调控基因表达。
翻译延伸的调控
翻译延伸的调控
在翻译延伸阶段,核糖体沿着mRNA移动,将氨基酸组装成蛋白质。原核生物通过控制翻译延伸因子的合成和活 性,以及核糖体的合成和组装来调控翻译延伸。此外,原核生物还可以通过控制mRNA的结构和稳定性来影响翻 译延伸。
总结
翻译延伸是基因表达调控的重要环节,原核生物通过控制翻译延伸因子的合成和活性,以及核糖体的合成和组装, 以及mRNA的结构和稳定性来精细调控基因表达。
翻译起始的调控
原核生物通过控制翻译起始来调控基因表达。在翻译起始阶段, mRNA与核糖体结合,招募翻译所需的起始因子和其他成分。原 核生物通过控制起始因子的合成和活性,以及mRNA与核糖体的 结合来调控翻译起始。
总结
翻译起始是基因表达调控的重要环节,原核生物通过控制翻译起 始因子的合成和活性,以及mRNA与核糖体的结合来精细调控基 因表达。
第七章 真菌的分子生物学鉴定方法

RAPD是一种比较精细的分子标记技术,单个碱基的改 变有可能对扩增片断的多态性产生明显的影响。
国内有人利用RAPD对白色念珠菌、淋球菌、水稻白叶 枯病菌进行分型取得了很好的结果。 黄勃等人利用 RAPD 技术对拟青霉属菌株进行分类鉴定时,发现
RAPD 指纹图谱在拟青霉属不同种间具有明显的种的
特异性,可以区别所有形态近似的种类,但对比RAPD
(二)真菌线粒体DNA限制性片段长度多态性(RFLP)分析 限制性片段长度多态性分析(restriction fragment length polymorphism, RFLP),将目标DNA序列经一定数目和种类的限 制性内切酶(RE)进行酶切,由于不同生物体的基因组DNA在限制性 内切酶的酶切位点的遗传信息有差异, 限制性内切酶在其上的识别
例——捕食线虫真菌的分类发展
粘球和 非收缩环
节丛孢
收缩环
单顶孢
菌网
隔指孢
粘性分枝
分子生物学鉴定方法包括DNA碱基组成 (G+C)mol%、限制性片段长度多态性(RFLP)、 随机扩增多态性 (RAPD) 、Southerm印迹分析 脉冲电场凝胶电泳(PFGE)以及小亚基rDNA(或 rRNA)序列测定等。 过去依赖形态和生理生化等表型特征描述逐 渐引入分子生物学鉴定方法,按其亲缘关系和 客观的反映系统发育的规律对真菌进行自然分 类,达到人们追求已久的自然分类目的,无疑 是分类学发展过鉴定等的很好的分子 标记,已经被成功地用于动物、植物、人和微生物的遗 传多样性检测、基因定位、品系鉴定、医学诊断、遗传 图谱构建和系统学研究等。 在分析生物间系统发育关系、分析突变以及亲缘关系鉴 定等方面显示出卓越的功能,一些表型上不能反映的遗传 物质的细微变化都可以通过RAPD技术显示出来。
分子生物学复习总结题-第七章-基因表达调控

第七章基因表达调控一、选择单选:1. 关于“基因表达”的概念叙述错误的是A. 其过程总是经历基因转录及翻译的过程B. 某些基因表达产物是蛋白质分子C. 某些基因表达经历基因转录及翻译等过程D. 某些基因表达产物是RNA分子E. 某些基因表达产物不是蛋白质分子2. 关于管家基因叙述错误的是A. 在生物个体的几乎各生长阶段持续表达B. 在生物个体的几乎所有细胞中持续表达C. 在生物个体全生命过程的几乎所有细胞中表达D. 在生物个体的某一生长阶段持续表达E. 在一个物种的几乎所有个体中持续表达3. 目前认为基因表达调控的主要环节是A. 翻译后加工B. 转录起始C. 翻译起始D. 转录后加工E. 基因活化4. 顺式作用元件是指A. 基因的5’、3’侧翼序列B. 具有转录调节功能的特异DNA序列C. 基因的5’侧翼序列D. 基因5’、3’侧翼序列以外的序列E. 基因的3’侧翼序列5. 一个操纵子(元)通常含有A. 数个启动序列和一个编码基因B. 一个启动序列和数个编码基因C. 一个启动序列和一个编码基因D. 两个启动序列和数个编码基因E. 数个启动序列和数个编码基因6. 反式作用因子是指A. 对自身基因具有激活功能的调节蛋白B. 对另一基因具有激活功能的调节蛋白C. 具有激活功能的调节蛋白D. 具有抑制功能的调节蛋白E. 对另一基因具有功能的调节蛋白7. 乳糖操纵子(元)的直接诱导剂是A. 葡萄糖B. 乳糖酶C. β一半乳糖苷酶D. 透酶E. 别乳糖8. Lac阻遏蛋白结合乳糖操纵子(元)的A. CAP结合位点B. O序列C. P序列D. Z基因E. I某因9. cAMP与CAP结合、CAP介导正性调节发生在A. 葡萄糖及cAMP浓度极高时B. 没有葡萄糖及cAMP较低时C. 没有葡萄糖及cAMP较高时D. 有葡萄糖及cAMP较低时E. 有葡萄糖及CAMP较高时10. Lac阻遏蛋白由A. Z基因编码B. Y基因编码C. A基因编码D. I互基因编码E. 以上都不是11. 色氨酸操纵子(元)调节过程涉及A. 转录水平调节B. 转录延长调节C. 转录激活调节D. 翻译水平调节E. 转录/翻译调节12.基因表达的产物不包括A.蛋白质B. mRNAC. rRNAD. SnRNAE. tRNA13.真核基因调控中最重要的环节是A. 基因重排B. 基因转录C. DNA的甲基化与去甲基化D. mRNA的衰减E. 翻译速度14.RNA聚合酶结合于操纵子的A. 结构基因起始区B. 阻遏物基因C. 诱导物D. 阻遏物E. 启动子15. cAMP对转录的调控作用是通过A. cAMP转变为CAPB. CAP转变为CampC. 形成cAMP-CAP复合物D. 葡萄糖分解活跃,使cAMP增加,促进乳糖利用来扩充能源E. cAMP是激素作用的第二信使,与转录无关16. 原核生物与DNA结合并阻止转录进行的蛋白质称为A. 正调控蛋白B. 阻遏物C. 诱导物D. 反式作用因子E. 分解代谢基因激活蛋白17.增强子A. 是特异性高的转录调控因子B. 是真核生物细胞内的组蛋白C. 原核生物的启动子在真核生物中就称为增强子D. 是增强启动子转录活性的DNA序列E. 是在结构基因的5'-端的DNA序列18.关于色氨酸操纵子的错误叙述是:A.trpR参与阻抑调控B.色氨酸阻抑结构基因转录C.前导序列参与色氨酸操纵子的衰减调控D.色氨酰tRNA参与色氨酸操纵子的衰减调控E.前导序列的序列3和序列4形成衰减子结构多选:1、基因表达调控环节包括A.DNA复制B.转录起始C.转录后加工D. mRNA降解E.翻译2、关于原核生物基因表达A.每个原核细胞的一切代谢活动都是为了适应环境而更好地生存和繁殖B.操纵子是原核生物绝大多数基因的表达单位C.原核生物基因表达的特异性由 因子决定D.原核生物基因表达既存在正调控,又存在负调控E.转录起始是原核生物基因表达主要的调控环节3、原核生物基因的调控序列包括A.启动子B.终止子C.操纵基因D.增强子E.衰减子4、原核生物基因的调控蛋白包括A.特异因子B.起始因子C.延长因子D.激活蛋白E.阻抑蛋白5、乳糖操纵子包含以下哪些结构?cZB. lacAC. lacOD. lacPE. lacI6、关于乳糖操纵子的错误叙述是:A.乳糖操纵子编码催化乳糖代谢的3种酶cI促进乳糖操纵子转录C.别乳糖促进乳糖操纵子转录D.CAP促进乳糖操纵子转录E.cAMP抑制CAP的激活效应7、色氨酸操纵子的结构A.含trpYB.含trpAC.含trpOD.含trpPE.含前导序列8、与RNA聚合酶活性调控有关的成分有A.tRNAB.核糖体C.严谨因子D.鸟苷五磷酸E.鸟苷四磷酸9、以下关于cAMP对原核基因转录的调控作用的叙述,正确的A. 葡萄糖与乳糖并存时,细菌优先利用乳糖B. cAMP-CAP复合物结合于启动子上游C. 葡萄糖充足时,cAMP水平不高D. cAMP可与CAP结合成复合物E. 葡萄糖和乳糖并存时,细菌优先利用葡萄糖10、原核生物基因表达在翻译水平上的调控与那些因素有关?A.mRNA前体后加工B. mRNA稳定性C. SD序列D.翻译阻抑E.反义RNA11、以下哪些环节存在真核生物的基因表达调控A.DNA和染色质水平B.转录水平C. 转录后加工水平D. 翻译水平E. 翻译后加工水平12、与原核生物相比,真核生物的基因表达调控的特点是A.转录的激活与转录区染色质结构的变化有关B.转录和翻译分隔进行,具有时空差别C.转录后加工更复杂D.既有瞬时调控又有发育调控E.转录调控以正调控为主13、在真核生物基因表达调控过程中,DNA水平的调控包括哪些内容A.染色质结构改变B. DNA甲基化C. 基因重排D. 基因扩增E.染色质丢失14、关于真核生物基因表达转录水平的调控A.转录水平的调控实际上是对RNA聚合酶活性的调控B.RNA聚合酶Ⅱ是转录调控的核心C.转录水平的调控主要通过RNA聚合酶、调控序列和调控蛋白的相互作用来实现D.真核生物的调控序列又称顺式作用元件E.真核生物基因表达的调控蛋白即转录因子,又称为反式作用因子15、真核生物的调控序列有哪些?A.启动子B.终止子C.增强子D.沉默子E.衰减子16、哪些属于真核生物基因表达的调控蛋白A.转录因子B.反式作用因子C.通用转录因子D. 反式激活因子E.共激活因子17、哪些是真核生物调控蛋白所含的DNA结合域A.螺旋-转角-螺旋B.锌指C.富含脯氨酸域D.亮氨酸拉链E.螺旋-环-螺旋。
92184-分子生物学-第七章 蛋白质变异效应与短肽

线 粒 体 呼 吸 链 膜 蛋 白 复 合 体 Ⅱ 的 三 维 结 构
HIV-1亚型膜蛋白V3环的氨基酸变异引起的效应
不同HIV-1亚型膜蛋白V3环的氨基酸序列特征及变异 特点,病毒V3顶端的四肽特征主要为: AE亚型GPGQ 48% 、B亚型GPGR 36%、其他形式16% 。
还发现DQDR、DQGQ等少见V3环顶端四肽组成形 式。
当tau蛋白发生高度磷酸化、异常糖基化、异常截 断作用以及泛素蛋白化时,tau蛋白就失去对微管 的稳定作用,神经纤维退化,功能丧失,引起中 枢神经系统变性疾病。
第三节 错误折叠蛋白质引起的变异效应
错误折叠蛋白质通常产生三种效应:
被细胞防御机制降解,导致功能缺失; 发生错误定位,导致细胞功能紊乱; 错误折叠蛋白质相互聚集形成淀粉样沉积
线粒体膜蛋白氨基酸突变引起一些疾病
人类有很多疾病诸如嗜铬细胞瘤、副神经节瘤和李氏症等 均与线粒体复合物II的翻译提前终止或氨基酸突变相关, 这些疾病多表现为氧自由基引起的神经性紊乱,而氧自由 基的产生与电子在线粒体复合物II中传递中的泄漏有关。
通过线粒体复合物II的结构,对已知的与人类疾病相关的 该复合物的突变位点进行了精确定位,发现这些突变位点 均位于电子传递体或醌结合位点的周围,它们的突变或影 响了电子传递体的结合,或影响了醌的结合,从而导致电 子传递的中断,逃逸到线粒体基质中或线粒体内膜中,产 生大量氧自由基进而导致肿瘤。
研究发现穿膜肽可以双向穿越细胞膜。它还可以介导如 DNA、多胺类,甚至比自身分子量大很多倍的寡聚核苷酸 进入细胞内。值得一提的是,穿膜时,分子是作为一个整 体插入,而不需要解折叠。该分子完全可以作为一种全新 的载体,它不需要能量,可以挂载的分子的类型多,分子 量大。
医学分子生物学 7 分子生物学常用技术

5′
3′
*********G —OH
*********C T T A A — P
3′
5′
5′
3′
P A A T T C*********
OH— G*********
3′ 5′
EcoR I: dATP+ [α-32P]-dTTP
19
根据反应原理确定同位素反应底物名称
• DNA的切口平移标记法 • 随机引物标记法 • DNA的3′末端标记
• 极高的特异性
缺点 半寿期短, 故要随用随标 放射性污染
9
1. DNA探针放射性标记方法
(1) 切口平移标记法 (2) 随机引物标记法 (3) PCR标记法 (4) 5′-末端标记 (5) 3′-末端标记
10
(1) DNA的切口平移标记法
5′
3 ′ 双链DNA
3′
5′
DNase I,Mg2+
② 反应产物的长度与加入的寡核苷酸引物的量呈反比。
当需要较长片段探针,可适当减少随机引物的加入量。
③ 所得到的标记产物为新合成的DNA单链。当采用单链
DNA片段或RNA作为模板时,必须注意得到的标记探针并 不是其本身.
④ 反应条件要控制在pH值6.6,抑制Klenow DNA聚合酶
的3′-5′外切酶的活性。
第七章 分子生物学常用技术
1
第一节 核酸分子杂交
2
*核酸分子杂交:
具有一定互补序列的不同来源的核苷酸单 链在一定条件下按照碱基配对的原则形成
杂交双链的过程。
实质: 核酸分子的变性与复性过程
变性:将双链DNA分子解聚成为单链的过程 复性:使单链聚合成双链的过程,又称为退火. 特点:高度特异性和灵敏性 杂交的双方: (靶,target) --待测序列
分子生物学复习7-9

第七章基因的表达与调控(上)——原核基因表达调控模式(一)基本概念1.基因表达:细胞在生命过程中,把蕴藏在DNA中的遗传信息经过转录和翻译,转变成为蛋白质或功能RNA分子的过程称为基因表达。
2.基因表达调控:围绕基因表达过程中发生的各种各样的调节方式都统称为基因表达调控。
rRNA或tRNA的基因经转录和转录后加工产生成熟的rRNA或tRNA,也是rRNA或tRNA 的基因表达,因为rRNA或tRNA就具有在蛋白质翻译方面的功能。
3.组成型表达:指不大受环境变动而变化的一类基因表达。
如DNA聚合酶,RNA聚合酶等代谢过程中十分必需的酶或蛋白质的表达。
管家基因:某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因。
管家基因无论表达水平高低,较少受到环境因素的影响。
在基因表达研究中,常作为对照基因适应型表达:指环境的变化容易使其表达水平变动的一类基因表达。
应环境条件变化基因表达水平增高或从无到有的现象称为诱导,这类基因被称为可诱导的基因;相反,随环境条件变化而基因表达水平降低或变为不表达的现象称为阻遏,相应的基因被称为可阻遏的基因。
4.结构基因:编码蛋白质或功能性RNA的任何基因。
所编码的蛋白质主要是组成细胞和组织基本成分的结构蛋白、具有催化活性的酶和调节蛋白等。
原核生物的结构基因一般成簇排列,真核生物独立存在。
结构基因簇由单一启动子共同调控。
调节基因:参与其他基因表达调控的RNA或蛋白质的编码基因。
①调节基因编码的调节物质通过与DNA上的特定位点结合控制转录是调控的关键。
②调节物与DNA特定位点的相互作用能以正调控的方式(启动或增强基因表达活性调节靶基因,也能以负调控的方式(关闭或降低基因表达活性)调节靶基因。
操纵子:由操纵基因以及相邻的若干结构基因所组成的功能单位,其中结构基因的转录受操纵基因的控制。
(二)原核基因调控的分类和主要特点一、原核生物的基因调控特点:(1)基因调控主要发生在转录水平上,形式主要是操纵子调控.(2)有时也从DNA水平对基因表达进行调控,实质是基因重排。
分子生物学 第7章 RNA复制与逆转录ppt课件

精选ppt课件2021
19
tRNA
R U5 PB
gag
R′ U′5
R U5 PB
gag
R′ U′5
PB
gag
负链 DNA
An
pol
env
U3 R
以 tRNA 为引物开始合成 cDNA 负链
pol
env
U3 R An
RNase H 降解模板 R 和 U5
An
pol
env
U3 R
负链 DNA3′端与模板 R 区配对(第一次跳跃)
精选ppt课件2021
11
7.2.1 RNA复制酶类
由病毒自己编码的RNA复制酶主要是依赖RNA的RNA 聚合酶(RNA-dependent RNA polymerase,RdRP)。
RNA复制酶功能与特性: ⑴ 以“–RNA”为模板合成“+RNA”活性高,而以“+RNA” 为模板合成“–RNA”活性低; ⑵ 合成RNA链方向为5′→3′; ⑶ 能将互补的双链RNA分开; ⑷ 缺少核酸酶活性,校正功能低,差错率高; ⑸ 多数分布在细胞质中,少数在细胞核中; ⑹ 酶活性具有可调控性,复制时,RdRP抑制外壳蛋白表达, 而成熟期,外壳蛋白抑制RdRP活性。
这类病毒包括滤泡性口腔炎病毒、流感病毒、
副流感病毒、莴苣坏死黄化病毒、麻疹病毒、狂犬病毒
等。
脱壳
病毒
RdRP
加工
亲代 –RNA
+RNA ① RdRP
子代
mRNA ② 翻译
–RNA
GpppN GpppN
帽子 mRNA
③ 翻译
子代病毒颗粒 组装
RNA 复制酶 病毒外壳蛋白 致病蛋白
分子生物学第7章 DNA的重组与转座

Barbara McClintock 芭芭拉·麦克林托克) (芭芭拉 麦克林托克) 19021902-1992
C基因-Ds基因-Ac基因: Ac-Ds 控制系统 基因-Ds基因-Ac基因: Ac基因 基因 《玉米易突变位点的由来与行为》(1950),《染色体结构和基 玉米易突变位点的由来与行为》(1950), 因表达》 因表达》(1951)
7.2.2 细菌的特异位点重组
鼠伤寒沙门氏菌H1鞭毛蛋白和 鞭毛蛋 鼠伤寒沙门氏菌 鞭毛蛋白和H2鞭毛蛋 鞭毛蛋白和 白抗原表达的互变,称为鞭毛相转变, 白抗原表达的互变,称为鞭毛相转变, 这种转变是由H片段倒位决定的 片段倒位决定的, 这种转变是由 片段倒位决定的,此倒 位是由于特异重组位点hix发生重组后引 位是由于特异重组位点 发生重组后引 起的。 起的。
整合位点---attB、attP ◘ 整合位点 切除位点---attL、attR 切除位点 整合过程需要λ整合酶 ◘ 整合过程需要 整合酶 (integrase Int)( 编码) )(λ编码) )( 编码 和寄主的整合宿主因子IHF 和寄主的整合宿主因子 (integration host factor) ) 共同作用 ◘ 整合后的附着位点为 attL(BOP’) ( ) attR(POB’) ( )
Ⅰ型:两末端有相同的IS序列,或正向,或反向。 两末端有相同的IS序列,或正向,或反向。 IS序列 Ⅱ型:末端有反向重复序列。如TnA家族 末端有反向重复序列。 TnA家族
Tn A家族: A家族: 家族
TnA是复制型转座的转座子,长约5kb左右 TnA是复制型转座的转座子,长约5kb左右 。 是复制型转座的转座子 5kb 两端具有末端反向重复序列(而不是IS),长约38bp左右 IS),长约38bp左右, 两端具有末端反向重复序列(而不是IS),长约38bp左右, 任一个缺失都会阻止转座。 任一个缺失都会阻止转座。 中部的编码区编码三个基因:转座酶,解离酶和抗性基因, 中部的编码区编码三个基因:转座酶,解离酶和抗性基因, TnA家族都带有抗性标记 TnA家族都带有抗性标记 。 靶位点具有5bp的正向重复序列。 5bp的正向重复序列 靶位点具有5bp的正向重复序列。 解离位点( TnA家族特有的内部位点 家族特有的内部位点。 解离位点(res)是TnA家族特有的内部位点。
分子生物学 第七章试题

一、是非题1、Lac阻遏蛋白结合乳糖操纵子(元)的 P序列。
2、分解代谢物基因激活蛋白(CAP)对乳糖操纵子表达的影响是正调控。
二、单选题1、Lac阻遏蛋白由()A、Z基因编码B、Y基因编码C、 A基因编码D、 I基因编码2、DNA损伤修复的SOS系统()A、是一种保真性很高的复制过程B、LexA蛋白是一系列操纵子的阻遏物C、RecA蛋白是一系列操纵子的阻遏物D、它只能修复嘧啶二聚体3、色氨酸操纵子中的衰减作用最终导()A、DNA复制的提前终止B、在RNA中形成一个抗终止的发夹环C、在RNA中形成一个翻译终止的发夹环D、RNA polymerase从色氨酸操纵子的DNA序列上解离4、色氨酸操纵子的转录调控包括()A、阻遏系统B、阻遏系统和弱化系统C、诱导系统D、诱导系统和弱化系统三、多选题1、基因表达调控可以发生在( )A、转录水平B、复制水平C、转录起始D、翻译水平E、翻译后水平2、以下关于cAMP对原核基因转录的调控作用的叙述错误的是( )A、cAMP可与分解代谢基因活化蛋白CAP结合成复合物B、cAMPCAP复合物结合在启动子前方C、葡萄糖充足时,cAMP水平不高D、葡萄糖和乳糖并存时,细菌优先利用乳糖四、填空题1、Trp 操纵子的精细调节包括()机制及()机制等两种机制。
2、胰岛素在胰岛的β细胞表达、而在α 细胞不表达,称为基因表达的()特异性,又称为()特异性。
五、术语解释1、核糖体开关2、RNA干扰3、Operon:(操纵子)4、基因调控(gene regulation)六、简答题1、说明E.Coil 乳糖操纵子如何控制对乳糖的利用?2、说明E.coil 色氨酸操纵子如何控制其结构基因的表达?。
分子生物学课件第七章 RNA的生物合成

(for emergencies)
E. coli also has six alternative sigma factors that are used in special circumstances
Some of the sigma factors found in Bacillus subtilis
Control of sporulation in Bacillus subtilis also involves antisigma factors and anti-anti-sigma factors!
真核生物的启动子
三种RNA pol; 不同的RNA pol识别不同的启动子;
Promoter of RNAP II
原核生物RNA聚合酶可以被 利福霉素(rifamycin) 抑制。
利福霉素是从 链霉菌(streptomyces) 中分离得到的一 种抗生素,可以与细菌RNA pol的β亚基直接结合,抑制第 一个磷酸二酯键的形成。
人工合成的利福平 rifampicin,可供口服,用于细菌感染 的临床治疗。
真核生物叶绿体RNA聚合酶和线粒体RNA聚合酶对 -鹅膏蕈碱不敏感,但可以被利福霉素抑制。
RPB5 RPB6 RPB7
RPB8 RPB9 RPB10 RPB11 RPB12
220 Binds DNA
’
(and has CTD)
150 Binds NTPs
45
assembly
1
32
Not essential,
(stress response)
27
Target for activators
23
Efficient assembly
1分子生物学第七章基因作图和基因文库

原核生物重叠基因 • a. 一个基因完全在另一个基因里面。 • b. 部分重叠。 • c. 二个基因只有一个碱基对的重叠。 • 现在还不知道重叠基因在所有生物中是否具
有普遍性,仅在个别生物中发现有此现象, 例如流感病毒、大肠杆菌噬菌体G4,Qβ等
• 重叠基因的现象反映了原核生物利用有限的 遗传物质表达更多生物功能的能力。
• 一些物种之间的复杂性变化范围并不大,但 是C值却有很大的变化范围
• 这些问题的解决有待于进一步的研究。
基因组的基因数目
• 由于DNA中存在非编码序列,使计算产生 误差,故须先确定基因密度。
• 如果已知基因组的DNA序列,就可以获得 基因密度,包括ORF的平均长度和ORF间的 平均距离。
• 再根据已知基因数目及与已知基因相关的基 因数目来推测未知基因数目
• 虽然交换是一个普遍存在的遗传现象,但是,由 于当时还从来没有发现过交换发生在一个基因内 部的现象,因此在很长的一段时间里(直到40年 代中期)认为基因是三合一体,即既是一个功能 单位,也是一个突变单位和一个交换单位。
• (4)1944年Avery等首次用实验证实基因是 由DNA构成的。
• 基因是DNA
人
3.3×109 65000~8
0000
•真菌 •支原体(没有真正的细胞壁,格兰 氏阴性并且生长时需要类固醇)
•乳头(状瘤)多瘤空泡(形)病毒
•牛痘病毒
•细小病毒(小脱氧核糖核酸病毒)
Figure 1.36
The amount of
• 结构和功能越复杂,需要的基因产物的种类越 多,也就是说需要的基因越多,因而C值越大?
• 但是生物体复杂性和DNA含量之间并非线性关系(即 C值矛盾的本质)
分子生物学第7章RNA的生物合成

复制和转录的共同点:
DNA 模板 碱基配对规律 生成磷酸二酯键 链延长方向5'→3'
模板 原料
酶
产物
配对
复制和转录的不同点:
复制
转录
两股链均复制
dNTP DNA聚合酶 需要引物 子代双链DNA (半保留复制)
A -T,G-C
模板链转录(不对称转录) NTP
RNA聚合酶 不需要引物 mRNA , tRNA , rRNA
起始位点的嘌呤碱基 与模板+1位碱基配对
在模板指导下依 次引入NTP
延伸位点引入与模板+ 2位碱基配对的第二个 NTP,形成磷酸二酯键
新合成RNA链延伸约 6~10碱基后,σ因子从
全酶脱落
延伸阶段
图7-12 基因转录起始过程中第一个磷酸二酯键的形成
开放复合物内的RNA聚合酶往往会重复催化短RNA分子的 合成并释放它们,这样的合成称为“无效”合成。聚合酶的活 性中心最多能容纳8 nt,差不多等于“无效”转录物的长度。
n4CMP
产物RNA
7.1.2.2. 原核细胞RNA聚合酶的结构与功能
7.1.2.2.1 细菌的RNA聚合酶
核心酶(core enzyme):2
全酶(holoenzyme): 2
图7-2 E. coli RNA pol的体外组装
大肠杆菌RNA聚合酶各亚基的性质与功能
亚基 基因 rpo A
rpo B rpo C
7.1 DNA转录
7.1.1 转录的一般特征
(1) 转录发生在DNA分子上某些特定的区域 转录只发生在DNA分子上具有转录活性的区域,DNA两 条链并不是总会被转录。对某一特定基因来说,作为模板的 DNA链称为模板链、无义链、Waston链,与模板链互补的另 一条链称为编码链、有义链、Crick链。
分子生物学基础PPT第七章

第三节 真核基因表达转Байду номын сангаас水平的调控
2.锌指结构域 这类结构主要见于真核生物的调节因子。其结构如图7-4所示, 每个重复的“指”状结构约含23个氨基酸残基,锌以4个配价键与4个 半胱氨酸、或2个半胱氨酸和2个组氨酸相结合。
图7-4
蛋白质的锌指结构
第三节 真核基因表达转录水平的调控
3.亮氨酸拉链 亮氨酸拉链最初是通过比较酵母转录激活因子GCN4、 哺乳动物转录因子C/EBP(CAAT区及SV40增强子核心序列 结合蛋白)以及癌基因产物Fos、Jun和Myc的AA序列时发 现的。亮氨酸拉链中α螺旋的突出特点是每隔7个AA残基 出现一个疏水的亮氨酸残基。这些残基位于DNA结合域的C 端α-螺旋上,这样α-螺旋的侧面每两圈就会出现一个亮 氨酸,形成一个疏水的表面。结果α-螺旋的疏水表面间 就可相互使用,形成二聚体结构。早期的研究认为两个 α-螺旋疏水的亮氨酸残基交错排列,故称为亮氨酸拉链
图7-2 酵母中的交配型转换
第三节 真核基因表达转录水平的调控
一、真核基因转录与染色质结构变化的关系 DNA绝大部分都在细胞核内与组蛋白等结合成染色质, 染色质的结构影响转录,至少有以下现象: 1.染色质结构影响基因转录 在真核细胞中以核小体为基本单位的染色质是真核基 因组DNA的主要存在方式。DNA盘绕组蛋白核心形成核小体, 妨碍了与转录因子及RNA聚合酶的靠近和结合,使基因的 活性受到抑制。 2.组蛋白的作用 组蛋白H1及核心组蛋白共同参与核小体的组装与凝聚。 在特殊氨基酸残基上的乙酰化、甲基化或磷酸化等修饰, 可改变蛋白质分子表面的电荷,影响核小体的结构,从而 调节基因的活性。
第三节 真核基因表达转录水平的调控
4.螺旋-环-螺旋结构域(HLH) 螺旋-环-螺旋结合域总体上与亮氨酸拉链相似,只是 它的二个α-螺旋被一个螺旋的多肽分成二个单体蛋白,C 端α-螺旋一侧的疏水残基可以二聚化。与亮氨酸拉链一 样,螺旋-环-螺旋结构与经常与碱性结构域相邻,以形 成DNA结合蛋白所需的二聚体。这种特异型二聚体的形成 增加了转录因子所有组成成分的多样性和复杂性。其主要 特点是可形成两个亲脂性α-螺旋,两个螺旋之间由环状 结构相连,其DNA结合功能是由一个较短的富碱性氨基酸 区所决定的。其结构如图7-6所示。
分子生物学第七章原核生物基因表达调控

(三)、阻遏物 lac I 基因产物及功能
Lac 操纵子阻遏物 mRNA 是由弱启动子控制下组 成型合成的,该阻遏蛋白具有4个相同的亚基,每个亚 基均含347个氨基酸残基。
lacI 基因为组成型,通过启动子的上升突变体可获 得较多的阻遏蛋白;
阻遏物 2022/10/18
β-半乳糖苷酶 透过酶 转乙酰3酶2
2022/10/18
16
调节机理:
细胞中某一氨基酸或嘧啶的浓度发生改变
氨酰 – tRNA的浓度变化
核糖体在转录产物RNA上的结合位置不 同,使得RNA形成特定的二级结构 由RNA的二级结构判断基因能否继续转录
2022/10/18
17
3、降解物对基因活性的调节P252
葡萄糖效应或降解物抑制作用:细菌培养基中在 葡萄糖存在的情况下,即使加入乳糖、半乳糖等 诱导物,与其对应的操纵子也不会启动,这种现 象称为葡萄糖效应或降解物抑制作用。
这是通过阻止乳糖操纵子表达来完成的,这种 效应称为降解物抑制(catabolite repression)。
2022/10/18
35
(五)、cAMP与代谢物激活蛋白
葡萄糖
葡萄糖-6-磷酸
甘油 某些代谢产物抑制活性
腺苷酸环化酶
ATP
cAMP
编码
cAMP-CAP
Crp基因
代谢物激活蛋白 CAP
葡萄糖对其它糖的代谢抑制,是通过对 cAMP的抑制完成的。
2022/10/18
22
一、酶的诱导 ——
lac 体系受调控的证据
两种含硫的乳糖类似物:
异丙基巯基半乳糖苷
(IPTG)
巯甲基半乳糖苷(TMG)
E. coli 在不含乳糖的培养基生 长时,β-半乳糖苷酶含量极低;
分子生物学课件第七章 蛋白质的生物合成-翻译

2020/10/28
38
2020/10/28
39
原核生物翻译过程中核蛋白体结构模式:
P位:肽酰位 (peptidyl site)
A位:氨基酰位 (aminoacyl site)
E位:排出位 (exit site)
2020/10/28
40
蛋白质生物合成体系
mRNA、tRNA、rRNA n 氨基酸
大多数简并性表现在密码子的第三个核苷酸上,即 第一、二个核苷酸确定后,第三个核苷酸可变。
色氨酸
意义: 简并密码子越多,生物遗传的稳定性越大,
氨基酸出现频率越高
2020/10/28
18
(3)摆动性(wobble)
转运氨基酸的tRNA的反密码需要通过碱基互补与 mRNA上的遗传密码反向配对结合,但反密码与密码间 不严格遵守常见的碱基配对规律,称为摆动配对。
氨基酰-tRNA + AMP +PPi
2020/10/28
31
(2)氨基酰tRNA合成酶(amino acyl-tRNA synthetase,AARS)
存在于胞液中,催化一个特定的aa结合到相应的 tRNA分子上。
每种氨基酰tRNA合成酶对相应氨基酸以及携带氨 基酸的数种tRNA具有高度特异性,保证tRNA能够 携带正确的氨基酸对号入座。
UUU,UUG,UGU,GUU, GGG,GGU,GUG,UGG。 • U和G随机加入到三联体中,这样按比例各个位置 上进入U和G的概率不同,如氨基酸测定结果:
2020/10/28
7
• 如UUU:UGG=(555):(511)
= 25 : 1
• 同理UUU:UUG =5 :1,
• 根据检测结果推测:
分子生物学-13-4-第七章原核基因表达调控-Arb

因此只有在没有葡萄糖的时候,同时又有半乳糖的时候,启动子1才是开放的为什么gal 操纵子需要两个转录起始位点?(涉及半乳糖在细胞代谢中的双重功能)半乳糖两个作用: 可以作为唯一碳源供细胞生长; 与之相关的物质--尿苷二磷酸半乳糖(UDPgal )是大肠杆菌细胞壁合成的前体。
而启动子也有两个: galP1起始的转录——无内源葡萄糖、有外源半乳糖时进行,以保证碳源的供应。
galP2起始的转录——有内源葡萄糖、无外源半乳糖时进行,以保证细胞壁的合成需要。
生理功能(可以理解为生物学意义?)无论从必要性和经济性考虑,都要有一个不依赖于cAMP-CAP 的启动子(s2) 进行本底水平的组成型合成,以及一个依赖于cAMP-CAP 的启动子(s1),进行高水平的调节,这样既可以满足细胞最基本的需要(细胞壁),又可以满足在没有葡萄糖而有半乳糖时,细胞能够利用半乳糖进行生长。
进一步解释:gal P2是不依赖于cAMP-CRP 的,相反: cAMP-CRP 对gal P2还起到一种抑制作用,这是因为其与结合位点的结合,会影响到RNA 聚合酶对gal P2的利用。
因此教材上(page257)认为:只有S2活性完全被抑制时,(S1)的调控作用才是有效的。
7.4.2 阿拉伯糖操纵子(arabinose operon)araB 基因、araA 基因和araD, 形成一个基因簇,简写为araBAD三个基因的表达受到ara 操纵子中araC 基因产物AraC 蛋白的调控。
C 蛋白有三个结合位点O2、O1和 I 。
I BADCRPO2O1C结构基因调节基因P BADaraC 基因是araBAD 的调节基因L核酮糖激酶L阿拉伯糖异构酶L核酮糖-5-磷酸-4-差相异构酶结合到ara I 的时候,由于araBAD的启动子本身与ara I有部分重叠,另外还可以引起上游序列回折弯曲,使得AraC同时与O2结合,从而使CRP 聚合酶也不能结合到启动子上,araBAD基因不转录。
分子生物学2009-6
中去除5′→3′ 切酶活性的Klenow片段。该酶常用于测序、
探针标记等。
Taq DNA聚合酶:Taq DNA聚合酶使是一种耐热的依赖
于DNA的DNA聚合酶。最佳作用温度为72℃ ~80℃。常用 于PCR反应和测序反应。
逆转录酶: 反转录酶是以RNA为模板的DNA聚合酶。常
用于合成互补DNA(cDNA)。
分子生物学
杨孝朴
第七章
1 基因工程工具酶
基因工程
2 分子克隆载体与宿主细胞 3 DNA建立 4 目的基因的获得5 目的基因与载体的连接
6 重组DNA分子导入受体细胞 7 重组体的筛选
8 克隆基因的表达
第七章
基因工程
基因工程兴起于20世纪70年代初。1972年Berg P和他的同事将 首次构建出DNA的重组体。由于SV40能使动物致癌,出于安 全的 考 虑 , 这 项 工作没有进行下去 。 第二年 ,Cohen S和 得到基因的分子克隆,由此产生了基因工程。
衔接头可以使DNA片段的平末端转变为黏性末端,或由一种
黏性末端转变为另一种黏性末端,并且无需用限制酶切,在
基因工程中十分有用。
例如:EcoRⅠ-SmaⅠ平末端衔接头 5′-AATT CCCGGG-3′ 3′-GGGCCC-5′ EcoRⅠ-XmnⅠ-BamHⅠ衔接头 5′-AATT CGAACCCCTTCG-3′
DNA聚合酶(常用DNA聚合酶Ⅰ的大片段Klenow酶)填补缺
口,最后由T4DNA连接酶连接(见图)。
另外的一种连接方式是粘平末端的连接,即要连接的DNA片 段的一端为粘端、另一端为平端。产生“粘-平”方式的末 端有两条途径,即用一个产生平性末端的限制性内切酶(如 Sma Ⅰ , HpaⅠ ) 与 一 个 产 生 粘 性 末 端 ( 如 EcoR Ⅰ, BamHⅠ)的内切酶切割目的基因与载体;或先用一个产生 粘性末端的内切酶酶切DNA,然后用修饰酶补平或消平粘性
分子生物学第7章RNA加工和核糖核蛋白
➢内切酶识别切割5’端的前导区和2个3’端核苷 酸。
➢切除
剪 切
(a)
添 加 (C)
剪 接
(b)
碱 基
(d) 修
饰
(2) (1)
碱基修饰的方式:
(1)
(1)甲基化 如:A m A
一、原核生物 tRNA 加工
3.tRNA加工步骤
➢初始转录物折叠形成特异性的茎环结构。 ➢RNase E或F开始对3’端切割 ➢RNase D对3’端修剪至比成熟长度多2个2 nt。 ➢RNase P切割5’端生成成熟的5’端。 ➢最后,RNase D切除两个3’端的残基 ➢碱基修饰:通过甲基化酶,硫醇酶,假尿嘧啶核苷
20S pre-rRNA
45S pre-rRNA 41S pre-rRNA
32S pre-rRNA
Methylation takes place at over 100 sites to give 2´-O-methylribose
and this is now known to be carried out by a subset of small nuclear RNP particles which are abundant in the nucleolus. 细胞中的一种叫核 仁小核糖核蛋白颗粒(small nucleolar RNA, snoRNA)的短RNA分子指 导着rRNA分子特定位点的修饰作用。
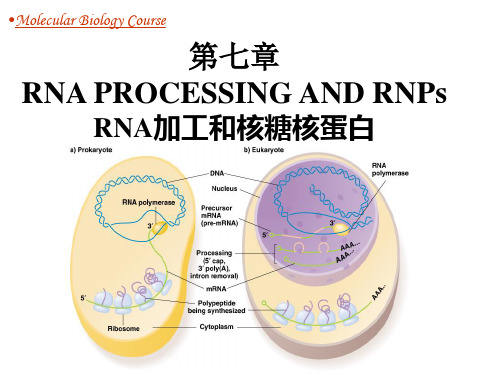
•Molecular Biology Course
第七章 RNA PROCESSING AND RNPs
RNA加工和核糖核蛋白
教学要求
了解RNA加工类型 理解并掌握真核生物mRNA加工的机制 掌握原核生物rRNA和 tRNA的加工
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Molecular Biology
Fifth Edition
Robert F. Weaver Chapter 7 Operons: Fine Control of Bacterial Transcription
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
Control of the lac Operon
• The lac operon is tightly controlled, using 2 types of control
– Negative control, like the brake of a car, must remove the repressor from the operator - the “brake” is a protein called the lac repressor – Positive control, like the accelerator pedal of a car, an activator, additional positive factor responds to low glucose by stimulating transcription of the lac operon
• As long as no lactose is available, the lac operon is repressed
7-6
Negative Control of the lac Operon
7-7
Inducer of the lac Operon
• The repressor is an allosteric protein
7-17
Mechanism Summary
• Two competing hypotheses of mechanism for repression of the lac operon
– RNA polymerase can bind to lac promoter in presence of repressor
7-15
The Mechanism of Repression
• The repressor does not block access by RNA polymerase to the lac promoter • Polymerase and repressor can bind together to the lac promoter • Polymerase-promoter complex is in equilibrium with free polymerase and promoter
– Causing the repressor to change conformation that favors release from the operator
• The inducer is allolactose, an alternative form of lactose
7-8
Inducer of the lac Operon
7-16
lac Repressor and Dissociation of RNA Polymerase from lac Promoter
• Without competitor, dissociated polymerase returns to promoter • Heparin and repressor prevent reassociation of polymerase and promoter • Repressor prevents reassociation by binding to the operator adjacent to the promoter • This blocks access to the promoter by RNA polymerase
7-11
Effects of Regulatory Mutations:
Wild-type and Mutated Repressor
7-12
Effects of Regulatory Mutations:
Wild-type and Mutated Operator with Defective Binding
7-5
lac Repressor
• When the repressor binds to the operator, the operon is repressed
– Operator and promoter sequence are contiguous – Repressor bound to operator prevents RNA polymerase from binding to the promoter and transcribing the operon
7-10
Discovery of the Operon
• Using merodiploids or partial diploid bacteria carrying both wild-type and constitutive alleles distinctions could be made by determining whether the mutation was dominant or recessive • Because the repressor gene produces a repressor protein that can diffuse throughout the nucleus, it can bind to both operators in a meriploid and is called a trans-acting gene because it can act on loci on both DNA molecules • Because an operator controls only the operon on the same DNA molecule it is called a cis-acting gene
• All 3 genes are transcribed together producing 1 mRNA, a polycistronic message that starts from a single promoter
– Each cistron, or gene, has its own ribosome binding site – Each cistron can be translated by separate ribosomes that bind independently of each other 7-3
– Binding of one molecule to the protein changes shape of a remote site on that protein – Altering its interaction with a second molecule
• The inducer binds the repressor
7-13
Effects of Regulatory Mutations:
Wild-type and Mutated Operon binding Irreversibly
7-14
Repressor-Operator Interactions
• Using a filter-binding assay, the lac repressor binds to the lac operator • A genetically defined constitutive lac operator has lower than normal affinity for the lac repressor • Sites defined by two methods as the operator are in factes of the lac Operon
• The genes are grouped together:
– lacZ = b-galactosidase – lacY = galactoside permease – lacA = galactoside transacetylase
7.1 The lac Operon
• The lac operon was the first operon discovered • It contains 3 genes coding for E. coli proteins that permit the bacteria to use the sugar lactose
7-18
lac Operators
• There are three lac operators
• The inducer of the lac operon binds the repressor • The inducer is allolactose, an alternative form of lactose
7-9
Discovery of the Operon
During the 1940s and 1950s, Jacob and Monod studied the metabolism of lactose by E. coli •Three enzyme activities / three genes were induced together by galactosides • Constitutive mutants need no induction, genes are active all the time • Created merodiploids or partial diploid bacteria carrying both wild-type (inducible) and constitutive alleles