细胞内蛋白质分选转运与细胞结构体系装配要求
细胞内蛋白质的分选和运输

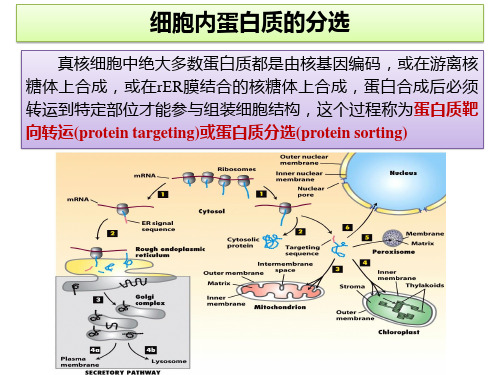
第十章细胞内蛋白质的分选和运输一个哺乳动物细胞含有近1万种、约1010个蛋白质分子。
除了少数蛋白质在线粒体核糖体上合成外,绝大部分蛋白质是由细胞核DNA编码、并在细胞质基质的核糖体上合成的。
蛋白质在细胞质基质中合成后,按其氨基酸序列中分选信号(sorting signal)的有无以及分选信号的性质被选择性地送到细胞的不同部位,这一过程称为蛋白质分选(protein sorting)和蛋白质靶向运输(protein targeting)。
另外,细胞外的蛋白质经胞吞作用进入细胞内部,也经历分选和靶向运输过程。
细胞中每一种蛋白质只有到达正确的位置才能行使其功能,例如激素受体必须进入细胞膜中才能识别相应的激素;RNA和DNA聚合酶必须送到细胞核中才能参与核酸的合成;酸性水解酶必须送到溶酶体才能进行大分子的降解作用。
因此,细胞内蛋白质的分选和运输对于维持细胞的结构与功能、完成各种细胞生命活动都是非常重要的。
第一节细胞内蛋白质的分选信号以及运输途径和方式细胞内各种蛋白质能够有条不紊地被运送到各自的目的地,关键是蛋白质在合成时带有分选信号,就象行李上贴上标笺一样注明了要到达的目的地,而在目的地有着能识别分选信号的受体,分选信号与相应受体的特异结合就引导蛋白质到达细胞的特定部位。
细胞内蛋白质的分选和靶向运输常常是多步骤的,有时要经过多次分选和运输才能完成。
目的地不一样,蛋白质运输的途径和方式也不一样。
一、信号假说和分选信号在20世纪70年代初有一个著名的实验:在体外无细胞系统中进行由mRNA编码的分泌蛋白在核糖体上合成,当合成系统中有微粒体存在时,所合成的蛋白质与体内合成的相同;如果在合成系统中去除微粒体,合成蛋白质的N-末端会多出一段短肽。
基于这一实验,1975年Blobel等人提出了信号假说(signal hypothesis),即在新合成蛋白质的N-末端有一段信号序列,称为信号肽(signal peptide),其作用是引导多肽链在合成过程中到内质网膜上,并在内质网中完成蛋白质的合成,而信号肽本身则在蛋白质合成完成前被内质网内的信号肽酶切除(图10-1)。
动物医学专业细胞生物学笔记5-6章

第五章物质的跨膜运输与信号传递物质的跨膜运输细胞通讯与信号传递思考题第一节物质的跨膜运输物质跨膜运输是细胞维持正常生命活动的基础之一●被动运输(passive transport)#特点:运输方向、跨膜动力、能量消耗、膜转运蛋白。
#类型:简单扩散(simple diffusion)、协助扩散(facilitated diffusion)#膜转运蛋白"载体蛋白(carrier proteins)——通透酶(permease)性质;介导被动运输与主动运输。
"通道蛋白(channel proteins)——具有离子选择性,转运速率高;离子通道是门控的;只介导被动运输。
类型:电压门通道(voltage-gated channel)配体门通道(ligand-gated channel)压力激活通道(stress-activated channel)●主动运输(active transport)#特点:运输方向、跨膜动力、能量消耗、膜转运蛋白。
被动与主动运输的比较#类型:三种基本类型*由ATP直接提供能量的主动运输"钠钾泵(结构与机制)"钙泵(Ca2+-ATP酶)"质子泵:P-型质子泵、V-型质子泵、H+-ATP酶。
*协同运输(cotransport):由泵与载体协同作用"共运输:动物中;植物中。
"对向运输:质子泵与逆向转运蛋白协同作用●胞吞作用(endocytosis)与胞吐作用(exocytosis)作用:完成大分子与颗粒性物质的跨膜运输,又称膜泡运输或批量运输(bulk transport)。
属主动运输。
#胞吞作用*胞饮作用(pinocytosis)与吞噬作用(phagocytosis)征**胞内体(endosome)的分选途径#胞吐作用*组成型的外排途径(constitutive exocytosis pathway)所有真核细胞连续分泌过程用于质膜更新(膜脂、膜蛋白、胞外基质组分、营养或信号分子)default pathway:除某些有特殊标志的驻留蛋白和调节的分泌泡外,其余蛋白的转运途径:粗面内质网→高尔基体→分泌泡→细胞表面*调节型外排途径(regulated exocytosis pathway)特化的分泌细胞储存——刺激——释放产生的分泌物(如激素、粘液或消化酶)具有共同的分选机制,分选信号存在于蛋白本身分选主要由高尔基体TGN上的受体类蛋白来决定*膜流:动态过程对质膜更新和维持细胞的生存与生长是必要的*囊泡与靶膜的识别与融合第二节细胞通讯与信号传递●细胞通讯与细胞识别#细胞通讯(cell communication)一个细胞发出的信息通过介质传到另一细胞产生相应的反应细胞间的通讯对于多细胞生物体的发生和组织的构建,协调细胞的功能,控制细胞的生长和分裂是必须的。
细胞内蛋白质分选的两条途径

细胞内蛋白质分选的两条途径细胞是生命的基本单位,其中蛋白质是细胞最重要的组成部分之一。
在细胞内,蛋白质需要在不同的位置发挥不同的功能,因此需要进行分选。
目前已知有两种主要的细胞内蛋白质分选途径:囊泡转运和直接转运。
一、囊泡转运1. 什么是囊泡转运?囊泡转运是指通过形成、移动和融合小型液滴(即囊泡)来实现蛋白质分选的过程。
这些囊泡可由内质网、高尔基体、溶酶体等细胞器形成。
2. 囊泡转运的过程(1)合成:在内质网上合成的蛋白质被包裹在一个小型液滴中,形成一个囊泡。
(2)移动:这些囊泡随后通过微管道系统向高尔基体或其他目标位置移动。
(3)融合:到达目标位置后,这些囊泡与目标部位上的膜进行融合,并释放出其所携带的蛋白质。
3. 囊泡转运的特点(1)速度快:相对于直接转运,囊泡转运速度更快。
(2)可控性高:囊泡转运可以通过调节囊泡合成、移动和融合等过程来实现对蛋白质分选的精确控制。
(3)适用范围广:囊泡转运可以用于多种类型的细胞内蛋白质分选,例如从内质网到高尔基体、从高尔基体到溶酶体等。
二、直接转运1. 什么是直接转运?直接转运是指蛋白质在没有形成液滴的情况下,通过与其他蛋白质或分子相互作用实现分选的过程。
这些相互作用可能包括靶标蛋白识别、信号传递等。
2. 直接转运的过程(1)靶标识别:特定类型的蛋白质通过与目标位置上的特定靶标结合来实现定向传输。
(2)信号传递:一些蛋白质需要特定信号才能在细胞内进行分选。
例如,磷酸化可以作为一种信号来调节蛋白质在细胞内的分布。
3. 直接转运的特点(1)精确度高:直接转运可以通过靶标识别和信号传递等机制来实现对蛋白质分选的精确控制。
(2)适用范围窄:相对于囊泡转运,直接转运的适用范围较窄,只适用于特定类型的蛋白质分选。
结论:细胞内蛋白质分选是细胞内复杂的过程之一,目前已知有两种主要的分选途径:囊泡转运和直接转运。
这两种途径在速度、可控性、适用范围等方面存在差异,但都可以通过不同机制来实现对蛋白质分选的精确控制。
2023年高中生物竞赛课件:细胞内蛋白质的分选
新生肽链跨膜取向
新生跨膜蛋白的肽链中没 有N端信号肽,只有内部 信号序列(internal signal sequence)。内部信号序列 的转位方向主要取决于其 侧翼氨基酸残基电荷的分 布。一般而言,带正电荷 的氨基酸残基一侧朝向细 胞质基质一侧
一、信号假说与蛋白质分选信号
Ⅳ型:G蛋白偶联受体、葡 萄糖转运蛋白、电压门 Ca2+通道、CFTR(Cl-)通道
图6-4 内质网膜整合蛋白的拓扑学类型
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
ER单次跨膜 新生跨膜蛋白肽链中既有N端信号肽,又 蛋白的合成 有停止转移信号
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
1、分泌性蛋白在游离核糖体上起始合成 2、多肽链延伸至80个氨基酸残基时,N端内质网信号序列暴露,与 SRP结合,肽链延伸暂停,防止新生肽链N端损伤和成熟前折叠
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
3、SRP与ER膜上SRP受体结合,核 糖体新生肽复合物附着到ER膜,两 分子GTP分别与SRP-p54亚基和SRP 受体α亚基结合,相互作用加强
5、腔面信号肽酶切除信号肽并使之快速降解 6-8、肽链继续延伸,并在ER腔内折叠,移位子关闭
一、信号假说与蛋白质分选信号
(四)分泌性蛋白的合成与共翻译转运过程
一、信号假说与蛋白质分选信号
(五)ER膜整合蛋白的信号序列
➢ 开始转移序列(start transfer sequence) 作为内质网的信号序列,指导新合成的多肽向内
细胞内蛋白质的分选和转运机制

细胞内蛋白质的分选和转运机制作者:陈建坤来源:《中学生物学》2018年第03期蛋白质作为生命活动的执行者和体现者,与生物的遗传、疾病等都有着重要关联。
在细胞内,有些蛋白质是先合成再进行分选转运,如线粒体、叶绿体、细胞核等结构中的蛋白质;而有些是边合成边分选转运,如分泌蛋白、膜蛋白等。
细胞根据蛋白质是否携有分选信号(信号序列)以及分选信号的性质,有选择地将蛋白质运送到细胞的不同部位。
1分选信号的种类分选信号有两类:①信号肽:蛋白质多肽链上一段连续的特定氨基酸序列,一般位于新肽链的N端,属于一级结构。
完成分选任务后常被切除。
②信号斑:位于多肽链不同部位的几个特定氨基酸序列经折叠后形成的斑块区,是一种三维结构。
完成分选任务后,仍然存在。
2原核细胞中蛋白质分选转运途径原核细胞(如细菌)没有复杂的生物膜系统。
但是为了维持生命,原核细胞需要合成一些蛋白质分泌到细胞质或者转运到细胞外发挥作用。
原核生物中蛋白质的转运分泌途径主要包括3种:①一般分泌途径,即SEC途径;②双精氨酸移位酶途径,即TAT途径;③信号颗粒识别途径,即SRP途径。
除此之外,还有V型分泌途径、TPS分泌途径和分子伴侣引导分泌途径等。
①一般分泌途径,即SEC途径:SEC分泌途径是原核生物中蛋白质主要的跨膜运输机制,主要由SEC移位酶作为介导。
SEC途径可以转运多种蛋白质,包括毒性因子、菌毛、黏附素和蛋白酶等。
SEC途径主要功能是把尚未折叠完成的蛋白质转运到质膜外,在质膜外折叠成有活性的蛋白质。
SEC途径大致可分为3个过程:信号序列的识别与定位、跨膜转运和多肽的释放。
②双精氨酸移位酶途径,即TAT途径:TAT途径识别的肽链N端信号序列通常含有两个连续的精氨酸残基。
TAT途径主要转运已经折叠完成的蛋白质,而尚未折叠完成的蛋白质通常不能通过该系统分泌,从而避免未完成折叠蛋白在胞外被降解的命运,保证了分泌产物的结构和功能的准确性。
此外,TAT途径还可以将少数蛋白质整合到质膜中。
细胞内蛋白质分选转运的主要方式

细胞内蛋白质分选转运的主要方式细胞内蛋白质分选转运是细胞内重要的生物过程之一,它确保了蛋白质在细胞内的正确定位和功能发挥。
细胞内蛋白质分选转运的主要方式包括囊泡运输、膜蛋白介导的转运和核糖体直接转运。
一、囊泡运输囊泡运输是细胞内蛋白质分选转运的重要方式之一。
它涉及到细胞膜上的囊泡与细胞器之间的运输。
囊泡是由膜蛋白包裹的小泡状结构,可以在细胞内进行蛋白质的转运。
囊泡运输主要分为内质网-高尔基体-溶酶体途径和内质网-高尔基体-细胞膜途径两种。
内质网-高尔基体-溶酶体途径是一种常见的囊泡运输方式。
在这个过程中,新合成的蛋白质被翻译成多肽链后,通过内质网的蛋白质翻译复合物进入内质网腔。
内质网腔中的蛋白质经过修饰和折叠后,被囊泡包裹形成转运囊泡。
这些囊泡随后与高尔基体融合,将蛋白质运输到溶酶体进行降解或分泌到细胞外。
内质网-高尔基体-细胞膜途径是另一种囊泡运输方式。
在这个过程中,蛋白质通过内质网进入高尔基体,然后通过囊泡运输到细胞膜。
这种方式主要用于膜蛋白的转运,以及一些细胞外分泌蛋白的释放。
二、膜蛋白介导的转运膜蛋白介导的转运是细胞内蛋白质分选转运的另一种重要方式。
在这个过程中,膜蛋白起到了关键的作用,它们通过与其他蛋白质相互作用,将目标蛋白质从一个细胞器转运到另一个细胞器。
一个典型的例子是膜蛋白介导的线粒体蛋白质转运。
线粒体是细胞内的重要细胞器,它需要从细胞质中转运蛋白质进入。
这个过程中,线粒体膜上的特定蛋白质与目标蛋白质相互作用,将其引导到线粒体内部。
这种蛋白质介导的转运方式在细胞内的其他膜蛋白转运中也起到了重要的作用。
三、核糖体直接转运核糖体直接转运是一种相对简单的蛋白质分选转运方式。
在这个过程中,蛋白质在合成过程中直接从核糖体转运到目标细胞器。
这种方式主要适用于一些小分子蛋白质的转运,例如核糖体合成的核糖体蛋白质。
细胞内蛋白质分选转运的主要方式包括囊泡运输、膜蛋白介导的转运和核糖体直接转运。
这些方式相互配合,确保了细胞内蛋白质的正确定位和功能发挥。
高尔基体——精选推荐

⾼尔基体⾼尔基体、溶酶体及过氧化物酶体最早发现于1855年,1889年,Golgi⽤银染法,在猫头鹰的神经细胞内观察到了清晰的结构,因此定名为⾼尔基体。
20世纪50年代以后才正确认识它的存在和结构。
⼀、形态与组成是由数个扁平囊泡堆在⼀起形成的⾼度有极性的细胞器。
常分布于内质⽹与细胞膜之间,呈⼸形或半球形。
凸出的⼀⾯对着内质⽹称为形成⾯或顺⾯(cis face)。
凹进的⼀⾯对着质膜称为成熟⾯或反⾯(trans face)。
顺⾯和反⾯都有⼀些或⼤或⼩的运输⼩泡。
扁平囊直径约1m,单层膜构成,中间为囊腔,周缘多呈泡状,4~4~88个扁平囊在⼀起(某些藻类可达⼀⼆⼗个),构成⾼尔基体的主体,即⾼尔基体堆(Golgi stack)。
⼆、功能区隔1、⾼尔基体顺⾯的⽹络结构(cis Golgi network,CGN):是⾼尔基体的⼊⼝区域。
接受由内质⽹合成的物质并分类后转⼊中间膜囊。
2、⾼尔基体中间膜囊(medial Golgi):多数糖基修饰、糖脂的形成以及与⾼尔基体有关的糖合成均发⽣此处。
3、⾼尔基体反⾯的⽹络结构(trans Golgi network,TGN):由反⾯⼀侧的囊泡和⽹管组成,是⾼尔基体的出⼝区域,功能是参与蛋⽩质的分类与包装,最后输出。
4、⾼尔基体周围还存在⼤⼩不等的囊泡⾼尔基体各部分膜囊具有不同的细胞化学反应:①嗜锇反应:cis⾯膜囊被锇酸特异地染⾊;②焦磷酸硫胺素酶(TPP酶):可特异显⽰⾼尔基体的trans⾯的1~2层膜囊;③胞嘧啶单核苷酸酶(CMP酶):可显⽰靠近trans⾯上的⼀些膜囊状和管状结构,CMP酶也是溶酶体的标志酶。
④烟酰胺腺嘌呤⼆核苷磷酸酶(NADP酶):可显⽰⾼尔基体中间⼏层扁平囊;⾼尔基体与细胞⾻架关系密切,在⾮极性细胞中,⾼尔基体分布在微管的负端----微管组织中⼼(MTOC)。
⾼尔基的膜囊上存在马达蛋⽩(cytoplasmic dynein和kinesin)和微丝的马达蛋⽩(myosin)。
海南大学生物工程学院2021年《细胞生物学》考试试卷(34)

海南大学生物工程学院2021年《细胞生物学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、判断题(35分,每题5分)1. 肿瘤细胞就是一类基因突变的干细胞。
()答案:错误解析:肿瘤组织中并非所有细胞都民间组织是干细胞,只有少数细胞是肿瘤干细胞。
2. 乳糖操纵子是一个超基因的功能单位,是由启动基因、操纵基因、调节基因和三个结构基因所组成。
()答案:错误解析:不包括调节基因。
3. 网格蛋白有被小泡中的“被”是指接合素蛋白。
()答案:错误解析:网格蛋白有被小泡的“被”还包括网格蛋白。
4. 叶绿体基质中的类囊体是彼此独立的由单位膜封闭形成的扁平小囊。
()答案:错误解析:叶绿体中相邻基粒类囊体经网管状或扁平状的基质类囊体相连,使类囊体腔彼此相通,因而一个叶绿体内一个全都类囊体实际上是的完整连续的封闭膜囊。
5. 酪氨酸磷酸化用来构建结合部位,用于其他蛋白质与受体酪氨酸激酶的结合。
()答案:正确解析:6. 核仁组织区就是核仁中负责组织核仁形成的纤维中心。
()答案:错误解析:核仁组织区是中期染色体位于次缢痕区的,是线粒体的一部分,与间期细胞核仁形成有关,但并非就是核仁凝胶的纤维中心。
7. IP3是直接由PIP2产生的,PIP2是从肌醇磷脂衍生而来的,肌醇磷脂没有掺入另外的磷酸基团。
答案:正确解析:PIP2带有三个磷酸基团,其中一个连接糖与二酰甘油酯。
IP3通过一个简单的水解反应产生。
2、名词解释(40分,每题5分)1. 细胞黏附分子(cell adhesion molecule)[浙江理工大学2019研]答案:细胞黏附分子是指促使细胞与细胞间的黏着或细胞与胞外基质间的黏着的分子,是基础理论细胞识别与黏着的分子基础。
主要类型有:钙黏蛋白、选择素、免疫球蛋白和整联蛋白王族后裔。
2015细胞生物学(翟中和第四版第8章 蛋白质分选(李绍军)

蛋白分选(蛋白质寻靶): 蛋白分子在蛋白内部的分选 信号指导下从细胞质运往各种目标细胞器或细胞表面。
• 细胞内合成的蛋白质、脂类等物质之所以能够定向的转运到特定的细胞器 取决于两个方面: – 其一是蛋白质中包含特殊的信号序列(signal sequence)。 – 其二是细胞器上具特定的信号识别装置(分选受体,sorting receptor)。
进入内质网腔的可溶性蛋白没有停止转移序列
含内在停止转移锚定序列(STA)和内在信号锚定序列(SA)的 蛋白在内质网膜上的拓扑特征
蛋定 膜 白序 整 膜列 合 定) 蛋 位和 白 中开 的 的始 内 作转 部 用移 停 序止 列转 (移 内序 部列 信( 号内 锚部 定停 序止 列转 )移 在锚
葡萄糖转运蛋白1等多跨膜α螺旋蛋白的合成和转运:多个开始转移序列(内部信号锚定序列,不切除的信号 肽)和停止转移序列(内部停止转移锚定序列)形成多次跨膜。
高尔基体在细胞分裂中的解体和分裂后重新组装Golgi assembly occurs in 2 stages upon mitotic exit: (a) Video frames illustrating post-mitotic Golgi assembly in mCherry(一种红色荧光蛋白)-Rab6(定位于高尔基体或后高尔基体区室上的GTP结合蛋白,调节膜 泡运输、融合) expressing RPE1(一种视网膜上皮细胞株) cells. Time zero marks approximate onset of telophase(末期开始). Boxed area is enlarged below. (b) Postmitotic Golgi particle size based on live imaging experiments in NT-control cells. Average fold increase of Golgi particles relative to time zero is shown. (c) Enlarged box from (a) showing Golgi mini-stack (red) clustering (6–9', blue and yellow arrows indicate two separate clusters) prior to re-location toward the centrosome (10'). Miller et al. Golgi-derived microtubules cluster and organize Golgi ministacks (G-stage of Golgi assembly). At the same time, centrosomal microtubules collect Golgi stacks in the cell center (C-stage of Golgi assembly). In CLASP高尔基 体膜上组织微管组装的一种蛋白)depleted cells that have no 细胞分裂末期高尔基体重新组装由两套微管体系完成——中心体微管体系(CGolgi-derived microtubules, G-
第八章蛋白质分选与膜泡运输
得到的免疫球蛋白分子的 N 端要比分泌到细胞外的免疫球
蛋白的N端多出一截。
第八章 蛋白质分选与 膜泡运输
本章主要内容
• 第一节 • 第二节 细胞内蛋白质的分选 细胞内膜泡运输
第一节 细胞内蛋白质的分选
细胞质中合成的上万种蛋 白质被分别运送到各自的 特定部位,说明细胞中必 定存在确保蛋白质准确分 选、转运的机制。这是通 过一定的结构和一系列复
杂的过程来完成的。
一、信号假说与蛋白于内质网膜上,直径约 8.5 nm 的通道蛋白,是中心有一直径为2 nm 通道的蛋 白复合体,参与新生蛋白的转运。
分泌蛋白等在内质网膜上的合成与转运
共转移 (cotranslocation):肽链边合成边转移 至内质网腔中的转运方式,也叫翻译同步转运。
跨膜疏水区
开始转移序列(Starttransfer Sequence),引 导肽链穿过内质网膜的 信号肽序列;停止转移 序列(Stop-transfer Sequence),与内质网膜 有很强的亲和力而结合 在脂双层之中,这段序 列不再转入内质网腔中 。
输入质体
输入过氧物酶体
Ser-Ser-Met-Ser-Ser-Leu-Ser-Leu-Ser-Ser-AsnSer-Phe-Leu- Gly-Gln-Pro-Leu-Ser-Pro-Ile-ThrLeu-Ser-Pro-Phe-Leu- Gln-Gly-
-Ser-Lys-Leu-COO+H 3N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-
细胞生物学第六章细胞内膜系统复习题

细胞生物学第六章细胞内膜系统复习题第六章细胞内膜系统基本概念题解学习要求:掌握细胞质基质和细胞内膜系统各部分的结构与功能。
掌握细胞内膜系统各个部分间的关系和细胞内蛋白分悬于细胞结构的装配及其相关知识。
基本概念:1.细胞质基质(cytoplasmic matrix):在真核细胞的细胞质中,除可分辨的细胞器以外的胶状物质,成为细胞质基质。
它是一种高度有序且不断变化的结构体系。
在确保与协调各种代谢反应、胞内物质运输与信息传递等方面,起重要作用。
2.内膜系统(endomembrane system) :指真核细胞内在结构、功能乃至发生上相关的由膜围绕的细胞器或细胞结构。
或者说是由膜分隔而形成的具有连续功能的系统,主要指核膜、内质网、高尔基体以及细胞质的各种囊泡。
而质膜、液泡膜以及溶酶体是这些内膜体系活动的最后产物,一般叶绿体、线粒体的膜也可直接或间接与内膜系统相联系但不包含在内膜系统内。
3.内质网(endoplasmic reticulum,ER):是分布于细胞质中由膜构成的网状管道系统,管道以各种形状延伸和扩展,成为各类管、泡、腔交织的状态。
内质网有两种:粗面内质网和滑面内质网。
前者指膜上附有核糖体颗粒。
后者膜外面不附有核糖体,表面光滑,主要是合成和运输类脂和多糖,它也可能是细胞之间通讯与传递系统。
细胞中内质网可以与细胞核的外膜相连,同时也可与细胞表面的质膜相连,而且还可能随同胞间连丝穿过细胞壁,与相邻细胞的内质网发生联系。
因此有人认为内质网构成了一个从细胞核到质膜,甚至与相邻细胞相连而直接贯通的管道系统。
4.易位子(translocon):指内质网膜上的一种蛋白质复合体,8.5nm,2nm的通道,其功能与新合成的多肽进入内质网腔有关。
5.高尔基体(Golgi body): 是由一些堆叠的扁平囊所组成。
主要功能是分泌活动、蛋白质加工以及合成多糖参与新细胞壁的形成。
6.溶酶体(lysosome): 是由单层膜围绕、内含多种酸水解酶类的囊泡状细胞器,其主要功能是进行细胞内的消化作用。
8.第八章 蛋白质的分选与膜泡运输

一、膜泡运输概观
➢内质网、溶酶体、分泌泡和细胞质膜及 胞内体也都具有各自特异的成分,这是 行使复杂的膜泡运输功能的物质基础。
➢在膜泡中又必须保证各细胞器和细胞间 隔本身成分特别是膜成分的相对恒定。
2021/5/15
2021/5/15
蛋白质的分泌与胞吞途径概观
二、 三种不同类型的包被膜泡具有不 同的物质运输作用
叶绿体蛋白质的运送及组装
2021/5/15
3、过氧化物酶体蛋白的分选
2021/5/15
第二节 细胞内膜泡运输
一、膜泡运输概观 膜泡运输是蛋白运输的一种特有的方
式,普遍存在于真核细胞中。在转运过 程中不仅涉及蛋白本身的修饰、加工和 组装,还涉及到多种不同膜泡定向运输 及其复杂的调控过程。
2021/5/15
2021/5/15
2021/5/15
图8-3分泌性蛋白的合成与其跨内质网膜的共翻译转运图解
2021/5/15
图8-3分泌性蛋白的合成与其跨内质网膜的共翻译转运图解
(2)跨膜蛋白的合成与转运过程
转运特点: ①信号肽(开始转移序列)分 N端信号肽和内部信号肽(信
号序列位于肽链内部)两种。 ②停止转移序列,即肽链上一段与内质网膜结合力很高的序
2021/5/15
(二)蛋白质转运四种基本类型
蛋白质的跨膜转运(transmembrane transport) 是指细胞基质中合成的蛋白质转运到内质网,线粒体、叶绿
体、过氧化物酶体等细胞器。 蛋白质的膜泡转运(vesicular transport)
是指从内质网到高尔基体进而分选至细胞的不同部位。
COPII包被膜泡 COPI包被膜泡 网格蛋白包被膜泡
2021/5/15
跨考专业课学824细胞生物学讲义-第6章

跨考专业课学824细胞生物学讲义-第6章跨考专业课-2022年考研北京大学824细胞生物学讲义-第6章考研全程辅导专家跨考教育专业课全力助你备考2022年第六章细胞质基质与细胞内膜系统第一节细胞质基质概念经典细胞学:光镜下,细胞除去可见的细胞器和内含颗粒的透明部分――细胞液细胞生物学:1、电镜下,除去可见的细胞器及亚微结构以外的细胞质部分――细胞质基质;(观察角度)2、分级离心后,除去所有的细胞器和颗粒剩下的清液部分――胞质溶胶(生化角度)这个概念是存在以下争议:有人认为细胞骨架不属于细胞质基质,是细胞器;有人认为细胞骨架是细胞质基质的主要结构体系,是其他成分锚定的骨架,经常处于装配和解聚的动态平衡中,解聚的亚单位仍保持在液相中。
(观点占多数)细胞质基质是高度有序的体系(p160)细胞质骨架纤维贯穿于蛋白质胶体中;蛋白质与骨架直(间)接结合,或与生物膜结合,完成特定的生物学功能;功能相关的酶通过弱键结合在一起形成多酶复合物,定位在特定部位,催化一系列反应;大分子之间通过弱键相互作用,并处于动态平衡之中。
细胞内膜系统内膜:电镜下可见的细胞质内的膜相结构,区分于质膜(细胞质膜)。
内膜系统(endomembrane system)是、高尔基体、细胞核、溶酶体和液泡(含内体和分泌泡)5类细胞器,它们的膜是相互流动的,处于动态平衡之中,功能上也相互协同。
内膜系统的共同结构特点:都是单位膜结构;仅存在于真核细胞中;处于动态平衡中,膜之间有转化现象。
内膜系统和质膜的结构区别:单位膜的层次不如质膜明显;厚度稍薄,6~7nm;膜上的抗原不同。
第二节内质网endoplasmic reticulum,ER概述(P164)K. R. Porter(1945)发现于培养的小鼠成纤维细胞,是位于细胞质内部的网状结构,故名内质网。
ER是由封闭的膜系统及其围成的腔形成的互相沟通的网状结构。
存在于真核细胞中,占细胞膜系统总面积的一半左右。
细胞内蛋白质的分选和运输细胞生物学

胞外
胞内
细胞膜
高尔基体
内质网
三、蛋白质从细胞质基质到内质网的运输
蛋白质进行的是一种穿膜运输转运在蛋白质合成的 过程中进行即共翻译转运此运输过程不需要直接能量
1.可溶性蛋白质到内质网腔的运输
转运子的中央孔与核糖体大亚基的中央通道对齐 供多肽链穿入
信号肽引导多肽链通过内质网膜上蛋白质转运子进入内质网
细胞根据蛋白质是否携有分选信号以及分选 信号的性质选择性地将其送到细胞不同的 部位这一过程称为蛋白质分选Tri des protéines 和蛋白质靶向运输l’adressage des protéines
大多数蛋白质带有分选信号
第一节 细胞内蛋白质的分选信号 以及运输途径和方式
吞噬作用
吞饮作用
二、胞吞途径 voies d‘endocytose
细胞摄入胆固醇的方法 LDL颗粒-受体介导的胞吞
有被小泡
遗传性高胆固醇血症原因之一
受体缺陷:不能形成有被小泡
溶酶体的形成与胞吞密切相关
早期内体
晚期内体
溶酶体
高尔基体
三、生物合成-分泌途径 从细胞内到细胞表面的蛋白质分选和运输
细胞不同部位的蛋白质存在不同的运输方式如通过核孔复 合体进出核的运输为门控运输;到内质网、线粒体、过氧化物酶 体的运输为穿膜运输;而从内质网到高尔基体高尔基体的各个 区室之间以及由高尔基体到溶酶体、细胞膜或重新运回内质网 的运输是由运输小泡介导的
门控运输为通过核孔复合体进出核的运输是一个信号识别 核输入、输出受体分别识别核输入、输出信号和载体介导 的主动运输过程具有选择性、双向性、耗能的特点蛋白在 核质间的转运受严格的控制
高尔基体到细胞表面的蛋白质分选运输
8-2 蛋白质的分选与膜泡运输 细胞生物学

微管
膜泡运输的关键步骤:
1.供体膜的出芽、装配和断裂,形成不同的包被转运膜泡。 2.在细胞内由马达蛋白驱动,以围观为轨道的膜泡运输。 3.转运膜泡与特定靶膜的锚定和融合。(SNARE、RabGTP)
四、细胞结构体系的组装
生物大分子的组装方式
装配具有重要的生物学意义
生物大分子的组装方式
自我装配(self-assembly):信息存在于装配 大分子复合物亚基本身,细胞提供装配环境。 协助装配(aided-assembly):在大分子复合 物装配过程中,需要形成最终结构的亚基、其 他组分的介入;或者对装配亚基进行修饰,保 障装配过程或正确行使功能。 直接装配(direct-assembly):某些亚基直接 装配到预先形成的基础结构上。
在细胞合成与分泌途径中,不同膜组分之间 三种不同的膜泡运输方式: 1. 网格蛋白有被小泡:介导从高尔基体TGN 质膜和胞内体及溶酶体的运输; 2. COPII有被小泡:介导顺向运输,从内质网 高尔基体的运输; 3. COPI有被小泡:介导逆向运输,即在高尔基 体内膜囊间和从cis膜囊、CGN RER。 (物质沿内吞途径的转运未表示在图中)。
跨膜受体
被运输蛋白
装配:
Sar1-GDP
Sec12
膜泡运输

信号假说与蛋白质分选信号 蛋白质分选的基本途径与类型 膜泡运输 细胞结构体系的组装
一、信号假说与蛋白质分选信号 一、信号假说与蛋白质分选信号
信号假说(Signal hypothesis) 信号假说 G.Blobel et al:Signal hypothesis,1975 . : 膜蛋白的形成 导肽( 导肽(Leader peptide)与后转移(Post translocatio)
跨膜运输( 跨膜运输(transmembrane transport); ) 膜泡运输( 膜泡运输(vesicular transport) ) 门控运输(gated transport); 门控运输( ) 细胞质中蛋白质的转运
三.膜泡运输
膜泡运输是蛋白运输的一种特有的方式, 膜泡运输是蛋白运输的一种特有的方式,普遍存在 于真核细胞中。在转运过程中不仅涉及蛋白本身的修饰、 于真核细胞中。在转运过程中不仅涉及蛋白本身的修饰、 加工和组装, 加工和组装,还涉及到多种不同膜泡定向运输及其复杂 的调控过程。 的调控过程。 三种不同类型的包被小泡具有不同的物质运输作用 膜泡运输是特异性过程,涉及多种蛋白识别、装配、 膜泡运输是特异性过程,涉及多种蛋白识别、装配、 去装配的复杂调控
信号假说
信号密码→信号肽 识别→SRP-核糖体复合体 蛋 核糖体复合体→蛋 信号密码 信号肽→SRP识别 信号肽 识别 核糖体复合体 白质合成暂时终止→SRP-核糖体复合体与内质网膜上 核糖体复合体与内质网膜上 白质合成暂时终止 受体结合 核糖体与核糖体结合蛋白(易位子 的 SRP受体结合 核糖体与核糖体结合蛋白 易位子 结 受体 结合→核糖体与核糖体结合蛋白 易位子)结 解离→蛋白质合成重新开始 合→SRP解离 蛋白质合成重新开始 信号肽通过易位 解离 蛋白质合成重新开始→信号肽通过易位 子进入内质网→信号肽切除 肽链合成完成→蛋白质游 子进入内质网 信号肽切除→肽链合成完成 蛋白质游 信号肽切除 肽链合成完成 离于内质网腔中→核糖体脱离内质网。 离于内质网腔中 核糖体脱离内质网。 核糖体脱离内质网
细胞生物学_10真核细胞内膜系统、蛋白质分选与膜泡运输结果

在非细胞系统中蛋白质的翻译过程与 P、DP和微粒体的关系
细胞类至少存在两类蛋白质分选的信号:
①信号序列:存在于蛋白质一级结上的线性序列,通常1560个氨基酸残基,有些信号序列在完成蛋白质的定向转移 后被信号肽酶切除。 ②信号斑:存在于完成折叠的蛋白质中,构成信号斑的信号 序列之间可以不相邻,折叠在一起构成蛋白质分选的信号。 ③每一种信号序列决定特殊的蛋白质转运方向。
由内质网到高尔基体的蛋白转运中,大多数跨膜蛋白是直 接结合在COP II衣被上,但是少数跨膜蛋白和多数可溶性蛋 白通过受体与COP II衣被结合,这些受体在完成转运后,通 过COP I衣被小泡返回内质网。
COP II衣被所识别的分选信号位于跨膜蛋白胞质面的结构 域,形式多样。
COPⅡ小泡的装配需要一种称为Sar1的G蛋白的参与。 当Sar1中GDP与GTP进行了交换,诱导Sec23和Sec24蛋 白的结合,接着是Sec13和Sec31蛋白的结合,最后由一 种结合在ER表面的大蛋白质,Sec16与Sec23/Sec24复 合物、Sec13/Sec31复合物相互作用,装配成一个完整的 小泡。
四、细胞结构体系的组装
目前机理尚不清楚。 组装方式:自我组装、协助组装、直接组装。 组装具有重要的生物学意义: ⒈减少和校正蛋白质合成中出现的错误。 ⒉可大大减少所需的遗传物质信息量。 ⒊通过组装与去组装更容易调节与控制多种生物学过程。
分子伴侣:细胞中的某些蛋白质分子可以识别正在合成 的多肽或部分折叠的多肽并与多肽的某些部位结合,从 而帮助这些多肽转运、折叠或组装,这一类分子本身并 不参与最终产物的形成。
每种转运小泡都有特异的v-SNARE,能识 别并与靶膜上t-SNARE相互作用。通过vSNARE与t-SNARE两类蛋白间的互补性和相 互作用,决定供体膜泡在靶膜上的锚定与融合。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
共转移(信号肽)
主要是在粗面内质网 上合成的蛋白;
特点:蛋白质边合成 边转移
内质网腔
后转移(post translocation) 1 分子伴侣结合
线粒体,叶绿体等的蛋 胞质中多肽
白;
2 导肽结合线粒体
基本特征:
上的受体
蛋白完全在细胞质基质 中合成,然后再转移到 某些细胞器中。
3 当多肽转移时, 伴随ATP水解,胞 质中的分子伴侣 脱离
②膜泡运输(vesicular transport):蛋白由不同类 型转运小泡从其ER合成部位转运至高尔基体进 而分选运至细胞的不同部位。
③孔门运输(gated transport):细胞质基质中合成 的蛋白质通过核孔复合体到核内或相反。
④细胞质基质中蛋白质的转运:和细胞骨架相关。
二 膜泡运输
普遍存在于真核细胞中,是蛋白运输的一种 特有方式。在转运过程中涉及蛋白本身的修 饰、加工和组装,及不同膜泡定向运输和复 杂的调控过程。 >10种运输小泡参与完成胞内的膜泡运输, 其上有特殊标志,可以保证转运物质到达特 定部位。
蛋白质跨膜转移需
4 线粒体中的分子
ATP使多肽去折叠,及一 伴侣结合转运中的
些蛋白的帮助(如Hsp70) 多肽
使其正确折叠。
5随ATP水解
而释放,多肽在
线粒体中折叠
2.蛋白质分选的四种基本类型:
①跨膜转运(transmembrane transport): 在细胞 质基质中合成的蛋白质转运到内质网、线粒体、 质体(包括叶绿体)和过氧化物酶体等细胞器。 蛋白以非折叠态跨膜。
组成: COPI包被含有8种蛋白亚基, 其依赖ARF(GTP 酶)调节包被的装配与去装配;
功能: ① 负 责 回 收 、 转 运 内 质 网 逃 逸 蛋 白 ( escaped proteins)返回内质网(Golgi →ER,逆行转运); ②行使从Golgi →ER-Golgi中间组分→ ER的物质 运输。 ③在组成性分泌过程中,其在非选择性的批量运 输中行使功能。
内质网:信号肽; 线粒体:导肽; 叶绿体:转运肽; 过氧化物酶体:(Peroxisomal-targeting signal,
PTS),C端PTS1,N端PTS2; 核定位蛋白:核定位信号,(nuclear localization signal,
NLS); 溶酶体水解酶:ER CGN, 信号斑。
装配:
跨膜受体
被运输蛋白
Sar1-GDP Sec12 Sar1-GTP(活化)
露出一条脂肪酸的尾巴,插 入内质网膜,促进其他衣被 蛋白的活化和组装。
激活磷脂酶D,将一些磷 脂水解,使形成衣被的蛋 白牢固地结合在膜上。
形成运输小泡 当衣被小泡从膜上释放后,GTP水解,衣被解体。
3) COPI包被小泡
内体
1.参与膜泡运输的三种不同类型的有被小泡
网格蛋白有被小泡; COPII有被小泡; COPI有被小泡;
网格蛋白 接合素蛋白
1)网格蛋白包被小泡
受体
结构:
网格蛋白组成外层结构骨架;
接合素构成内壳。
功能:
货物分子
a.负责蛋白质从高尔基体TGN到质膜、胞内体或溶酶体 和植物液泡的运输。
b.在受体介导的细胞内吞途径中负责将物质从质膜运到 细胞质,以及从胞内体到溶酶体的运输。
① 网格(笼形)蛋白: 由3个重链和3个轻链形成的具 有3个曲臂形状(triskelion)的蛋白。笼形蛋白的 曲臂部分可相互交织,形成5/6边形网孔的“笼子”。
网格蛋白的 结构,A电 镜照片,B 分子模型, C衣被模型。
重链 轻链
重叠臂 曲臂末端
接合素
②接合素/衔接蛋白(adaptors) 介导网格蛋白与膜受体之间的连接。
内质网中保留及回收蛋白质的 两种机制:
①运输泡将应被保留的驻留蛋 白排斥在外,防止出芽转运;
②通过识别驻留蛋白C-端的 回收信号的特异性受体,以 COPI-包被小泡的形式捕获逃 逸蛋白。
内质网蛋白的回收信号:
网 L膜 L任具 蛋 或 内 狱由ye腔蛋意su有白回质”于-((的白氨L回发收网内yoK蛋(基sp收生回比D质-e白如酸XnE信逃来喻网-Lp:)SX号逸,成)的rR(i。L,,所“sP;驻yo开K受sn即也以K留-放)A体X使会有蛋的。sX)有保人p,白监-:G的留将Xl:u-
②膜组分的相对稳定有重要 作用:
高尔基体依靠膜囊上结合 的动力蛋白和微管相互作 用,向微管组织中心聚集, 从而维持极性;(秋水仙 素)
细胞内蛋白质的分选转运与细胞结构 体系的装配要求
绝大多数蛋白均在细胞质基质中的核糖 体上开始合成,然后转运至细胞的特定 部位,并装配成结构与功能的复合体, 参与细胞生命活动, 此过程称蛋白质的定 向转运(protein targeting)或分选(protein sorting)。
蛋白定位?
指导蛋白定位运转的信号:
在生物合成及分泌 途径中三种包被小 泡参与的膜泡运输。
粗面内质网: 物质供应站 高尔基体: 集散中心
vesicular tubular clusters
三种有被小泡的比较
Sec12
2膜泡运输的特异性
膜上受体和配体特异结合 决定膜泡运输的特异性;
①膜泡识别和融合是特异性 的:供体膜蛋白与受体膜 蛋白的特异性相互作用。
一 蛋白质分选的基本途径与类型
1.蛋白质分选的两条途径:
完全在细胞质基质合成的蛋白, 转运至膜性细
胞器(线粒体、叶绿体、过氧化物酶体、细胞
核)及细胞质基质的特定部位; •(后转移)
合成起始后转移至RER合成的蛋白质,经高尔基
体运至溶酶体、细胞膜或分泌到胞外或驻留在
Байду номын сангаас
内质网与高尔基体内。
•(共转移)
类型: AP1:高尔基体→胞内体的蛋白运输; AP2:质膜→胞内体的蛋白运输; AP3:高尔基体→溶酶体的蛋白运输。
③网格蛋白有被小泡介导的选择性运输
2) COPII 包被小泡
功能:介导从内质网到高尔基体的物质运输。
结构组分: Sar1:GTP酶,结合GDP失活,结合GTP活化;调节包被的
装配和去装配;召集其他包被蛋白形成包被; Sec23/Sec24复合体:和Sar一起构成包被内层; Sec13/Sec31复合体:构成包被外层; Sec16:可能作为骨架蛋白起作用; Sec12:Sar1的GDP-GTP交换因子。