实验文昌鱼早期胚胎发育及组织结构观察
文昌鱼的胚胎

生物技术与生物制药的应用前景
开发新的生物技术手段
通过对文昌鱼胚胎发育的研究,可以开发出新的生物技术手段,如基因编辑、干细胞分化等,为生物 技术的进步提供支持。
挖掘新的药物靶点
文昌鱼胚胎发育过程中涉及的基因和信号通路可能与药物研发有关,通过对其深入研究,可以挖掘出 新的药物靶点,为生物制药领域的发展提供支持。
文昌鱼的胚胎发育
目录
CONTENTS
• 文昌鱼的生命周期 • 胚胎发育过程 • 胚胎发育的生物学特性 • 文昌鱼胚胎发育的生态影响 • 文昌鱼胚胎发育的研究价值与应用前景
01 文昌鱼的生命周期
生长阶段
01
02
03
生长阶段
文昌鱼从孵化后进入生长 阶段,通过摄取食物和吸 收营养,逐渐发育和成长。
生长特点
03
水质
水中的溶解氧、pH值、污染物等都会影响文昌鱼胚胎的发育,良好的
水质可以促进胚胎的正常发育,而不良的水质则可能导致发育受阻或死
亡。
生物因子对胚胎发育的影响
食物供应
胚胎发育过程中需要消耗大量的能量,充足的食物供应是 保证胚胎正常发育的重要条件。食物的种类和质量都会影 响胚胎的发育速度和健康状况。
种间关系
在自然环境中,文昌鱼可能会与其他生物竞争食物、空间 等资源,这种竞争关系可能会影响其胚胎的发育。
病原体与寄生虫
病原体和寄生虫是生物体的常见威胁,它们可能侵入文昌 鱼的卵或胚胎,对其发育造成严重影响。
胚胎发育与种群动态的关系
1 2
繁殖力
文昌鱼的繁殖力直接影响种群的数量增长和动态 变化。胚胎发育的速度和质量会影响繁殖力的发 挥,从而影响种群动态。
多细胞动物早期胚胎发育

实验2 多细胞动物早期胚胎发育一、实验目的1.通过对文昌鱼、海胆、海星、蛙等胚胎发育各个时期的观察,了解多细胞动物早期胚胎发育的一般过程。
2.认识动物个体发育的一般概念,从而加深对多细胞动物起源的理解。
二、实验材料1.牛精子涂片,蛙、昆虫卵切片和鸡蛋。
2.文昌鱼早期胚胎发育4个时期装片或切片:受精卵期、卵裂各期、囊胚期和原肠胚期。
3.海星、蛙早期胚胎发育模型。
4.三化螟、蛙、鱼生活史标本。
三、实验器具与药品显微镜、双目解剖镜、手提放大镜、培养皿、尖镊子、小瓷碗等。
四、实验内容与操作(一)精子构造观察用显微镜观察牛精子涂片,区分精子的头、颈和尾3部分(图2-1)。
头部前端可见染色较浅的帽状结构,称顶体。
头部其余部分染色较深,是细胞核,由于着色深,核内构造看不清楚。
细胞核和顶体的外面为染色较浅的薄层物质,是细胞质和细图2-1 人类精子的显微和超微结构A.精子的超微结构;B、C.精子的不同视角显微观察(自Vilee)胞膜,但这部分在涂片中不一定看得清楚。
颈部短小,其内部构造也不易看清楚。
尾部呈长鞭毛状。
(二)各种类型卵子的结构及卵裂形式观察注意比较它们卵黄的含量及其分布状况与细胞质和细胞核的位置关系。
1.少黄卵均黄卵 观察文昌鱼(或海胆)卵切片或装片及哺乳动物卵巢切片,对照挂图从卵巢切片中找到卵细胞。
可见卵黄含量少,其分布均匀,未受精卵核大而清晰,受精后不见。
其卵裂形式属完全均等分裂(图2-2)。
2.多黄卵偏黄卵 观察蛙卵切片,卵黄的分布偏植物极,其卵裂形式属完全不均等分裂,从第三次分裂(纬裂)开始不均等,偏向动物极。
端黄卵 取一新鲜鸡蛋观察。
用尖镊子小心从上面打开卵壳和两层壳膜(将鸡蛋横放),通过“天窗”观察鸡蛋的内部构造(图2-3),弄清楚卵细胞、卵黄和哪些结构属卵膜。
鸡卵为图2-2 海胆卵的等裂A .2胚胞;B .4胚胞;C .8胚胞;D .16胚胞;E .32胚胞。
(自江静波等)图2-3 鸟类的蛋(卵黄部分为卵子)(自丁汉波) 图2-4 昆虫卵的表裂A .几个核时期;B .核分裂的正常分布;C .核移至胚胎的外表面及胚盘的形成。
文昌鱼实验报告的实验原理

文昌鱼实验报告的实验原理文昌鱼是一种典型的观赏鱼类,常被用于实验研究。
在文昌鱼实验报告中,实验原理是描述实验所采用的方法和原理。
以下是关于文昌鱼实验报告的实验原理的详细描述,包括文昌鱼的生物学特点、实验目的、实验方法和实验结果分析。
一、文昌鱼的生物学特点文昌鱼(也被称为金鱼)是一种淡水鱼类,属于鲤科(Cyprinidae)。
它是人工繁殖的品种,具有较高的观赏价值。
文昌鱼通常具有鳞片多、体型饱满、外观美丽的特点。
文昌鱼的生命活动、生理功能和行为习性具有研究价值,因此被广泛应用于实验研究。
二、实验目的文昌鱼实验的目的是为了探索和研究文昌鱼在特定环境条件下的生物学特性和生理功能。
通过实验,我们可以了解文昌鱼在不同压力等环境条件下的适应能力、生长发育情况以及对外界刺激的反应。
三、实验方法1. 实验准备:选择健康的文昌鱼作为研究对象。
根据实验需要,可以针对不同的实验目的选择合适的样本数量和种类。
2. 实验环境:为了保证实验的科学性和可靠性,需要为文昌鱼提供适宜的实验环境。
包括恒温、恒湿和恒光的条件,并且保证水质的清洁和稳定。
3. 实验设计:根据实验目的,设计合理的实验方案。
可以设置不同的实验组和对照组,对文昌鱼进行不同处理,观察和记录实验结果。
4. 实验操作:将选定的文昌鱼放置于实验容器中,进行实验处理。
根据实验需求,可以注射药物、调节水温、改变光照强度等,观察文昌鱼的生理反应和行为表现。
5. 数据采集:在实验过程中,要对文昌鱼的生长发育和生理状况进行观察和测量。
包括记录文昌鱼的体长、体重、生长速率、饮食量等指标。
同时,可以使用适当的技术手段(例如显微镜、生物化学分析等)对样本进行进一步分析。
四、实验结果分析1. 实验数据处理:收集实验数据后,需要对数据进行整理和分析。
可以使用统计学方法(例如t检验、方差分析等)对数据进行比较和推断。
2. 结果解释:根据实验数据的结论,可以解释文昌鱼在实验结果中所展现的生理和行为变化。
实验文昌鱼

作业:
绘文昌鱼经过咽区的横切面图
文昌鱼皮肤很薄,在身体两侧可以透过皮肤看到分节排列的肌节,肌节成 “V”形,夹角向前,相邻的肌节之间有肌隔。在性成熟的个体,生殖腺呈圆块 状沿身体的两侧排列在腹褶上方。
文昌鱼(上雌下雄)
(2)内部构造:
用低倍镜观察文昌鱼整体透 明标本,首先在体前部的背方找 到脊索,脊索一般染成黄褐色, 带有横纹,纵贯全身,几乎与身 体等长,神经管位于脊索的背方 与脊索平行,但略短,是文昌鱼 的中枢神经系统,神经管的管壁 上有许多点子状的希氏小眼(一 般染成深褐色),它是感光器官。 在神经管的背方可以看到背鳍, 有许多方形或长方形的鳍条,鳍 条内有背鳍辐骨。
青岛文昌鱼自然保护区
文昌鱼外形及内部结构
文昌鱼全长为5-6厘米,液浸标本略带黄色。生活在海滨沙砾中,我国 青岛、厦门及海南岛等海滨均可采到。
台湾海峡地区的四种文昌鱼
白氏鳃口文昌鱼 Branchiostoma belcheri
马尔地夫侧殖文昌鱼 Epigonichthys maldivensis
两栖纲(蛙、大鲵)
爬行纲(龟鳖、蛇、鳄鱼)
鸟纲(白头鹰、皇冠鸟、猫头 鹰、火烈鸟)
哺乳纲(狗、山魈、狼、犀牛、狮子、考拉、驯鹿、海豹、人类)
文昌鱼:
在今天的海洋里,有一种生物能引起我们古老的回忆,那就是我们的远亲 文昌鱼。现代生物学家研究我们这个大家族的起源,大都从文昌鱼入手。现生 的文昌鱼是一种很原始的小型脊索动物,只分布在世界上很少的地方,如中国 的青岛、厦门和美国的佛罗里达。它们没有头化的现象,脑子、眼睛都处在很 原始的状态下,用生物学的手段才能鉴别出它有脑子,有类似眼睛的构造。它 们从云南虫的一支分化下来后就停止了演化,在温暖的浅海里找到了它们永久 的栖身之所。而云南虫的另一支 ——海口虫经过亿万年的演化,告别了鳍和鳃 丝,有了自由呼吸的肺和自由行走的双腿,发展出了高级智慧。
实验-文昌鱼早期胚胎发育及组织结构观察

文昌鱼发育所经历的主要阶段如下:
受精卵——桑椹胚——囊胚——原肠胚——神经胚——幼体——成体。 文昌鱼在六、七月间为产卵期,产卵和受精均在傍晚进行。受精卵 不久即开始分裂,由于卵黄少,为均黄卵(isolecithal egg),因此卵 的分裂为全分裂(holoblastic),且为等分裂。第一次和第二次分裂皆 为垂直分裂,分成四个细胞,第三次分裂为横分裂,分成上下八个细胞, 如此继续分裂下去,许多细胞结成一个实心的圆球,称桑椹胚 (morula)。
(三)幼体期
• 文昌鱼的胚胎发育进行很快,在傍晚受精卵开始 分裂,至次日早晨,胚胎即突破卵膜,成为全身 披有纤毛的幼体,在海水表层自由游泳,以后沉 落到海底进行变态。幼体在发育过程中,体形逐 渐增大并延长,鳃裂的数目由少增多,由不对称 到对称,口由偏左侧移向腹侧,鳃裂由直接开口 体外到通入新形成的围鳃腔中。 • 文昌鱼的幼体期持续约3个月。在6个月内体长可 达到18mm,一年以后可达38mm,这时性腺已 成熟。Fra bibliotek实验材料:
文昌鱼早期胚胎发育切片: 文昌鱼受精卵装片, 文昌鱼2细胞装片, 文昌鱼4细胞装片, 文昌鱼8细胞装片, 文昌鱼16细胞装片, 文昌鱼囊胚期装片。 实验仪器:光学显微镜 文昌鱼组织结构切片: 文昌鱼整体装片, 文昌鱼过口笠横切, 文昌鱼过咽横切, 文昌鱼过肠横切, 文昌鱼过性腺横切, 文昌鱼尾部横切。
作业:
1. 绘制文昌鱼早期胚胎发育图片,受精卵,2细胞, 4细胞,8细胞,16细胞,囊胚期任选一个。 2. 绘制文昌鱼整体装片,并附上详细说明。 3. 绘制文昌鱼的横切图一张(任选),并附上详 细说明。
(二)器官的发生
原肠胚期结束后,开始产生中枢神经系统,形成中 胚层,三个胚层进一步分化成不同的器官系统。 1. 神经管的形成; 2. 脊索的形成; 3. 中胚层的发生与器官的分化; 4. 内胚层。
文昌鱼的胚胎发育进程
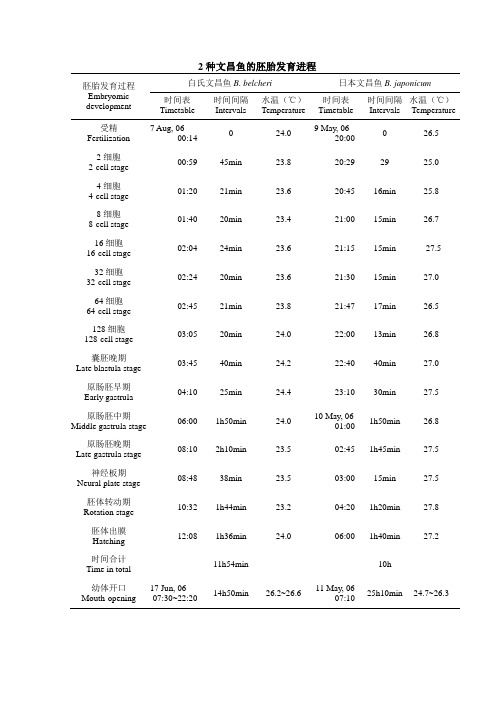
2种文昌鱼的胚胎发育进程胚胎发育过程Embryomic development白氏文昌鱼B. belcheri日本文昌鱼B. japonicum时间表Timetable时间间隔Intervals水温(℃)Temperature时间表Timetable时间间隔Intervals水温(℃)Temperature受精Fertilization 7 Aug, 0600:140 24.09 May, 0620:000 26.52细胞2-cell stage00:59 45min 23.8 20:29 29 25.04细胞4-cell stage01:20 21min 23.6 20:45 16min 25.88细胞8-cell stage01:40 20min 23.4 21:00 15min 26.716细胞16-cell stage02:04 24min 23.6 21:15 15min 27.532细胞32-cell stage02:24 20min 23.6 21:30 15min 27.064细胞64-cell stage02:45 21min 23.8 21:47 17min 26.5128细胞128-cell stage03:05 20min 24.0 22:00 13min 26.8囊胚晚期Late blastula stage03:45 40min 24.2 22:40 40min 27.0原肠胚早期Early gastrula04:10 25min 24.4 23:10 30min 27.5原肠胚中期Middle gastrula stage 06:00 1h50min 24.010 May, 0601:001h50min 26.8原肠胚晚期Late gastrula stage08:10 2h10min 23.5 02:45 1h45min 27.5神经板期Neural plate stage08:48 38min 23.5 03:00 15min 27.5胚体转动期Rotation stage10:32 1h44min 23.2 04:20 1h20min 27.8胚体出膜Hatching12:08 1h36min 24.0 06:00 1h40min 27.2时间合计Time in total11h54min 10h幼体开口Mouth-opening 17 Jun, 0607:30~22:2014h50min 26.2~26.611 May, 0607:1025h10min 24.7~26.3图5-1:白氏文昌鱼胚胎发育各阶段A:未受精卵;B:受精卵,受精膜举起;C:2细胞;D:4细胞;E:8细胞;F:16细胞;G:32细胞;H:64细胞;I:128细胞期,细胞之间界限还隐约可见,胚球表面较粗糙;J:囊胚晚期,细胞变小,胚球表面变光滑,植物极略变平坦(箭头所示);K:原肠早期,植物极细胞内陷(箭头所示);L:原肠中期,胚孔(箭头所示)直径约为胚胎直径的1/2;M:原肠晚期,胚孔(箭头所示)很小;N:神经板期,细箭头示神经板,粗箭头示胚体后端;O:神经胚转动期,粗箭头示神经胚前端,胚体后端的颜色较前端深;P:刚出膜的神经胚左侧面观,5个体节,箭头示胚体后端,颜色较前端为深;Q:神经胚左侧面观,具6体节;R:胚体逐渐伸长,具12体节;S:胚体继续伸长,鳃区扩大,体中部的神经管上出现第一个眼点(箭头所示),脊索纵贯全身;T:胚体逐渐转变成小鱼形;U:刚开口的幼体左侧面观,口开在幼体左侧(细箭头所示),口前窝(粗箭头所示)、尾鳍也开始出现,第一鳃裂形成中。
文昌鱼的解剖实训报告

一、实训目的通过本次文昌鱼解剖实训,使学生掌握文昌鱼的外部形态结构,了解其内部器官的形态、位置和功能,进一步理解无脊椎动物与脊椎动物在进化过程中的联系,提高学生的实验操作技能和观察能力。
二、实训时间2023年10月25日三、实训地点生物实验室四、实训材料文昌鱼、解剖刀、解剖剪、镊子、解剖盘、解剖针、显微镜、生理盐水、酒精、碘酒、解剖图谱等。
五、实训步骤1. 文昌鱼的外部观察首先,对文昌鱼进行外部观察,注意其体型、颜色、附着的物体等特征。
文昌鱼体型细长,呈圆柱形,体色透明,通常附着在岩石或珊瑚上。
2. 文昌鱼的内脏观察(1)解剖文昌鱼:用解剖剪在文昌鱼腹部中央剪开,注意不要伤及生殖腺。
(2)观察内脏:用解剖针轻轻挑开内脏,观察其形态、位置和功能。
(3)记录观察结果:将观察到的内脏器官及其功能记录在解剖报告中。
3. 文昌鱼生殖系统的观察(1)生殖腺的提取:用解剖剪在文昌鱼腹部中央剪开,轻轻挑开内脏,找到生殖腺。
(2)生殖腺的观察:将生殖腺放在解剖盘上,用显微镜观察其形态和结构。
(3)记录观察结果:将生殖腺的形态、结构和功能记录在解剖报告中。
4. 文昌鱼消化系统的观察(1)消化系统的提取:用解剖剪在文昌鱼腹部中央剪开,轻轻挑开内脏,找到消化系统。
(2)消化系统的观察:将消化系统放在解剖盘上,用显微镜观察其形态和结构。
(3)记录观察结果:将消化系统的形态、结构和功能记录在解剖报告中。
5. 文昌鱼循环系统的观察(1)循环系统的提取:用解剖剪在文昌鱼腹部中央剪开,轻轻挑开内脏,找到循环系统。
(2)循环系统的观察:将循环系统放在解剖盘上,用显微镜观察其形态和结构。
(3)记录观察结果:将循环系统的形态、结构和功能记录在解剖报告中。
6. 文昌鱼神经系统的观察(1)神经系统的提取:用解剖剪在文昌鱼头部中央剪开,轻轻挑开内脏,找到神经系统。
(2)神经系统的观察:将神经系统放在解剖盘上,用显微镜观察其形态和结构。
(3)记录观察结果:将神经系统的形态、结构和功能记录在解剖报告中。
实验-文昌鱼早期胚胎发育及组织结构观察

•
8、业余生活要有意义,不要越轨。20 20年12 月12日 星期六 1时36 分32秒1 3:36:32 12 December 2020
•
9、一个人即使已登上顶峰,也仍要自 强不息 。下午 1时36 分32秒 下午1时 36分13 :36:322 0.12.12
• 10、你要做多大的事情,就该承受多大的压力。12/12/
以后囊胚植物极的大细胞以内陷的方式向囊胚腔陷入,正像一个漏了
气的皮球,以至和上部动物极的细胞相贴近,囊胚腔被挤掉,而另形成一 个新的腔,称为原肠腔(archenteron)。原肠腔以原口(blastopore)与 外界相通。此时胚胎具备内外两层细胞,贴着原肠腔的一层细胞,称内胚 层(endoderm),与外界相接触的一层细胞,称为外胚层(ectoderm), 此阶段称原肠胚(gas- trula)。
作业:
1. 绘制文昌鱼早期胚胎发育图片,受精卵,2细胞, 4细胞,8细胞,16细胞,囊胚期任选一个。
2. 绘制文昌鱼整体装片,并附上详细说明。
3. 绘制文昌鱼的横切图一张(任选),并附上详 细说明。
(二)器官的发生
原肠胚期结束后,开始产生中枢神经系统,形成中 胚层,三个胚层进一步分化成不同的器官系统。
受精、卵裂及原肠胚的形成
桑椹胚一方面继续地分裂,一方面中心的细胞渐向表面迁移,因而变 成一个空心的圆球,此时期的胚胎称囊胚(blastula),中间的空腔内充满 胶状液体,称囊胚腔(blastocoel)。囊胚上端的细胞稍小,称动物极 (animal pole),下端的细胞稍大,称植物极(vegetative pole)。
• 13、无论才能知识多么卓著,如果缺乏热情,则无异 纸上画饼充饥,无补于事。Saturday, December 12, 2020
实验一文昌鱼形态结构

胚胎发育
01
02
03
卵裂与囊胚形成
受精卵经过迅速的卵裂形 成多个细胞,进而形成囊 胚。
原肠胚形成
随着胚胎发育,囊胚细胞 向内凹陷形成原肠胚,此 时胚胎开始分化出内胚层 和外胚层。
器官形成与孵化
原肠胚进一步发育,形成 各种器官和组织,最终孵 化出幼鱼。
实验一文昌鱼形态结构
目录
• 文昌鱼简介 • 文昌鱼形态特征 • 文昌鱼繁殖与发育 • 文昌鱼与人类的关系
01
文昌鱼简介
生物分类
生物分类
文昌鱼属于脊索动物门、头索动 物亚的动物。
特点
文昌鱼没有真正的头和脑,也没 有成对的器官和骨骼,但在进化 上具有重要地位,是研究脊椎动 物起源和演化的重要物种。
保护与环境问题
生态平衡
文昌鱼作为生态系统中的一部分,其数量的减少或增加可能对整个生态系统产生影响。
环境变化的影响
环境污染、气候变化等因素可能对文昌鱼的生存产生威胁,因此需要关注并采取措施进 行保护。
谢谢观看
地理分布
地理分布
文昌鱼主要分布在热带和亚热带的浅 海地区,包括我国南海、东海等海域 。
环境适应性
文昌鱼适应多种底质环境,从沙质底 到岩石底都有分布,但主要栖息在浅 海沙质底区域。
生态习性
食性
文昌鱼主要以浮游生物为食,如硅藻、蓝藻等。
繁殖
文昌鱼的繁殖方式是无性繁殖,通过分裂进行繁 殖,繁殖周期短,数量增长快。
骨骼结构
总结词
文昌鱼没有真正的骨骼,只有软骨和结缔组织构成的骨针。
详细描述
文昌鱼的软骨和结缔组织构成其支撑身体的骨针,这些骨针在显微镜下呈现为 细长的棒状结构,排列紧密,支撑着身体的主要部分。
文昌鱼受精卵实验报告

文昌鱼受精卵实验报告
文昌鱼的发育需经历受精卵一桑椹胚-襄胚- _原肠胚- 神经胚各个时期,才孵化成幼体。
受精卵进行几乎均等的全分裂( holoblastic) , 经过多次细胞分裂后,使许多细胞结成一个形似实心圆球的桑椹胚( morula)。
经过一系列的分裂分化,进而形成囊胚、原肠胚。
原肠胚胎已形成内、外两层细胞,分别称为内胚层( endoderm )和外胚层( ectoderm ) , 原肠胚的后期开始胚体延长、产生中枢神经、胚层进一步分化及各器言系统的形成神经胚( neurula )。
受精卵经过20多个小时后,文昌鱼的胚胎发育基本结束,全身披有纤毛的幼体就能突破卵膜,到海水中活动,此时有白天游至海底,夜间升上海面进行垂直洄游的生活规律。
幼体期约3个月,然后沉落海底进行变态。
幼体在生长发育和变态的过程中,身体日益长大, 出现前庭,鳃裂的数目因发生次生鳃条而增加了一倍,并由原来直接开口体外而变为通入新形成的围鳃腔中。
-龄的文昌鱼体长约40mm ,性腺发育成熟,可参与当年的繁殖。
有时,在某些特殊情况,幼体被卷入深海,变为深海的居住者。
身体不断地增长、很快地成为大型的幼体,甚至在变态为成体之前,它的生殖腺9早已开始发育了。
长满到11个月即开始成熟。
实验-文昌鱼早期胚胎发育及组织结构观察

精选课件
25
精选课件
16
作业:
1. 绘制文昌鱼早期胚胎发育图片,受精卵,2细胞, 4细胞,8细胞,16细胞,囊胚期任选一个。
2. 绘制文昌鱼整体装片,并附上详细说明。
3. 绘制文昌鱼的横切图一张(任选),并附上详 细说明。
精选课件
17
精选课件
18
精选课件
19
精选课件
20
精选课件
21
精选课件
22
精选课件
受精卵——桑椹胚——囊胚——原肠胚——神经胚——幼体——成体。
文昌鱼在六、七月间为产卵期,产卵和受精均在傍晚进行。受精卵 不久即开始分裂,由于卵黄少,为均黄卵(isolecithal egg),因此卵 的分裂为全分裂(holoblastic),且为等分裂。第一次和第二次分裂皆 为垂直分裂,分成四个细胞,第三次分裂为横分裂,分成上下八个细胞, 如此继续分裂下去,许多细胞结成一个实心的圆球,称桑椹胚 (morula)。
以后囊胚植物极的大细胞以内陷的方式向囊胚腔陷入,正像一个漏了
气的皮球,以至和上部动物极的细胞相贴近,囊胚腔被挤掉,而另形成一 个新的腔,称为原肠腔(archenteron)。原肠腔以原口(blastopore)与 外界相通。此时胚胎具备内外两层细胞,贴着原肠腔的一层细胞,称内胚 层(endoderm),与外界相接触的一层细胞,称为外胚层(ectoderm), 此阶段称原肠胚(gas- trula)。
精选课件
7
受精、卵裂及原肠胚的形成
精选课件
8
桑椹胚一方面继续地分裂,一方面中心的细胞渐向表面迁移,因而变 成一个空心的圆球,此时期的胚胎称囊胚(blastula),中间的空腔内充满 胶状液体,称囊胚腔(blastocoel)。囊胚上端的细胞稍小,称动物极 (animal pole),下端的细胞稍大,称植物极(vegetative pole)。
《实验文昌鱼》课件

显微镜、切片机、载玻片、盖玻片。
步骤
1. 将文昌鱼肌肉组织固定在载玻片上。
2. 使用切片机将肌肉切成薄片。
文昌鱼神经系统的观察
实验三
显微镜、染色剂、吸水纸。
准备工具
步骤
1. 对文昌鱼神经系统进行染色处理。
2. 用吸水纸吸干多、染色剂、培养皿。
准备工具
经济价值
由于文昌鱼的肉质鲜美,营养丰富,因此被视为美味佳肴。在一些国家和地区,文昌鱼是一种非常受欢迎的海鲜食品。此外,文昌鱼还具有观赏价值,许多人将它们养作为宠物鱼。
实验准备
显微镜
解剖工具
培养皿和培养基
离心机和相关试剂
01
02
03
04
用于观察文昌鱼的组织结构和细胞形态。
包括解剖刀、镊子、钳子等,用于解剖文昌鱼。
生活习性
繁殖习性
文昌鱼是一种雌雄异体的鱼类,繁殖方式是卵生。雌鱼会在海底或珊瑚礁上产卵,然后雄鱼受精。受精卵孵化后成为仔鱼,仔鱼经过一段时间的生长和发育,最终成为成鱼。
食性
文昌鱼主要以小鱼、甲壳类动物和浮游生物为食。它们具有锐利的牙齿和强壮的嘴巴,能够轻松地咬碎和吞食猎物。
生态价值
文昌鱼在海洋生态系统中扮演着重要的角色。它们是海洋食物链中的重要一环,通过捕食小鱼和甲壳类动物来控制这些物种的数量,从而维持生态平衡。
为了更深入地研究文昌鱼的繁殖机制,建议进一步探究影响文昌鱼繁殖的环境因子和遗传因子。
感谢观看
THANKS
结果观察和记录
02
01
03
04
05
实验操作
文昌鱼解剖观察
实验一
文昌鱼肌肉组织切片
实验二
文昌鱼神经系统的观察
实验十 青蛙早期胚胎发育的观察

枣庄学院
• 实验作业 • 绘蛙早期胚胎的囊胚期发育简图, 注明各部分结构名称。
图10-2 文昌鱼囊胚(自曲漱惠等)
• 4.原肠期 • 胚胎继续发育转入原肠期。囊胚较大的一 端植物极发生内陷,并向内部迁移,内陷 继续进行,结果形成具有两层细胞的胚体, 称原肠胚。组成原肠胚的外层细胞称外胚 层,内层细胞称为内胚层。内外两胚层之 间残留的空腔为原来的囊胚腔,由内胚层 细胞包围的腔称原肠腔。内胚层细胞在胚 胎发育过程将形成消化道的上皮组织,故 称原肠。原肠腔与外界相通的小孔叫胚孔 或原口(图10-3)。
• 文昌鱼受精卵的第1、2次卵裂是从动 物极向植物极纵裂的,第3次呈水平分 裂,共形成8个分裂球,进入8细胞期, 因为细胞排列不在同一平面上,观察 时必须及时转动细调焦器才能看清楚。 以后再交替地分裂下去则形成16细胞 期、32细胞期等。
• 3.囊胚期
• 图10-2文昌鱼囊胚(自曲漱惠等)
• 文昌鱼的受精卵经过多次的卵裂后, 分裂的细胞群逐渐向胚胎的表面迁移, 结果发育成由单层上皮细胞构成的中 空的球形胚体,称囊胚期。观察文昌 鱼囊胚期装片,注意单层上皮细胞称 囊胚层,中央的空腔称囊胚腔,腔内 有细胞分泌的囊胚液(图10-2 )。
• 原口的位置是胚胎的后端,相反一端 是前端,因此,胚胎发育到原肠期就 已经确定了胚胎的对称面。文昌鱼的 胚胎发育继原肠期之后,内胚层继续 分化出脊索和中胚层,外胚层分化出 神经管等,最终完成整个胚胎发育过 程。
•
图10-3 文昌鱼原肠作用过程 1 外胚层;2 中胚层;3 内胚层;4 脊索;5 囊胚腔; 6 胚孔背唇;7 胚孔腹唇;8 原肠腔;9 胚孔 (自曲 漱惠等)
•
图10-1 文昌鱼早期卵裂 A 受精卵;B 2细胞时期;C 4细胞时期;D 8细胞时期; E 16细胞时期;F 32细胞时期 (自曲漱惠等)
文昌鱼Gsc基因在早期胚胎发育中的功能

第62卷 第5期厦门大学学报(自然科学版)V o l .62 N o .5 2023年9月J o u r n a l o f X i a m e nU n i v e r s i t y (N a t u r a l S c i e n c e )S e p.2023 h t t p :ʊjx m u .x m u .e d u .c n d o i :10.6043/j.i s s n .0438-0479.202209032文昌鱼G s c 基因在早期胚胎发育中的功能商留珂1,2,刘惠敏2,石成刚2,李 光2,王义权2*(1.洛阳职业技术学院医学技术学院,河南洛阳471000;2.厦门大学生命科学学院,福建厦门361102)摘要:同源异型盒基因G o o s e c o i d (G s c )在脊椎动物胚胎 组织者 和脊索前板形成中具有重要功能.为探究该基因在脊索动物胚胎发育中的功能演化,通过基因过表达㊁原位杂交等手段对其在文昌鱼胚胎发育中的功能展开分析.结果显示:向文昌鱼胚胎中注射G s c m R N A 及其转录抑制形式E N -G s c (H D )m R N A ,均导致胚胎发育出现相似的体轴发育畸形,说明G s c 在文昌鱼胚胎发育中具有重要功能,且主要以抑制型转录因子形式发挥作用.基因表达分析显示,G s c 过表达导致原肠胚期中胚层发育相关基因B r a c h y u r y ㊁W n t 8和背部标记基因C h o r d i n ㊁L e f t y 的表达量下降,腹部标记基因V e n t 表达范围向背部扩张;神经胚中期B r a c h y u r y 在后端尾芽的中胚层祖细胞中表达量下降,体节标记基因m -a c t i n 标记的体节数减少,神经管和尾芽标记基因W n t 3在尾芽处的表达量下降而在神经管处表达范围变宽.以上结果表明,文昌鱼G s c 基因在胚胎早期发育过程中的体轴建立㊁中胚层形成及后期尾芽功能维持中均有重要功能.关键词:文昌鱼;G s c 基因;中胚层形成;胚胎发育中图分类号:Q953 文献标志码:A 文章编号:0438-0479(2023)05-0858-07收稿日期:2022-09-21 录用日期:2022-12-27基金项目:河南省卫生健康委员会医学教育研究项目(W jl x 2020262);国家自然科学基金面上项目(32070458,32070815)*通信作者:w a n g y q@x m u .e d u .c n 引文格式:商留珂,刘惠敏,石成刚,等.文昌鱼G s c 基因在早期胚胎发育中的功能[J ].厦门大学学报(自然科学版),2023,62(5):858-864. C i t a t i o n :S H A N GLK ,L I U H M ,S H I CG ,e t a l .F u n c t i o n o f G s c g e n e i n e a r l y e m b r y o g e n e s i s o f a m ph i o x u s [J ].JX i a m e nU n i v N a t S c i ,2023,62(5):858-864.(i nC h i n e s e)组织者 是指动物胚胎发育早期位于背部的一组具有 指导 或 组织 其他细胞分化能力的细胞,最早由德国科学家H a n sS pe m a n n 在两种蝾螈(T r i t o n c r i s t a t u s 和T .t a e n i a t u s)胚胎移植实验中发现并命名[1].G o o s e c o i d (G s c )基因最早由C h o 等[2]在爪蛙(X e n o p u s l a e v i s )胚胎的背唇同源异型盒基因文库中发现,其转录本存在于 组织者 区域,在爪蛙胚胎发育的4细胞时期,向腹部两个分裂球中注入G s c m R N A 可诱导胚胎形成第二体轴,显示该基因具有 组织者 的功能.G s c 基因广泛存在于无脊椎动物和脊椎动物中,在脊椎动物胚胎发育早期,该基因在 组织者 [2-3]及脊索前板[4]区域特异性表达.G s c 基因突变的小鼠原肠胚期胚胎发育正常,但出生后不久即死亡,突变体小鼠颅面㊁颌骨发育不正常[5-7],体轴发育亦受影响[8].果蝇(D r o s o p h i l am e l a n o ga s t e r )中与G s c 基因同源的D -gs c 基因也有类似功能,可挽救果蝇经紫外光照射引起的体轴异常[9].敲低爪蛙胚胎中G s c基因表达,爪蛙胚胎腹部中胚层扩大[10].由上可见,G s c 基因参与胚胎的中胚层图式化和背腹轴的形成.文昌鱼是头索动物亚门(C e ph a l o c h o r d a t a )动物的总称,是最接近脊椎动物直接祖先的现生动物[11-13],其成体的躯体构筑方式与脊椎动物相似但更简单[14-15],是研究脊椎动物胚胎发育机制和起源演化的理想模型.1962年T u n g 等[16]开展的文昌鱼移植实验及2007年Y u 等[17]在文昌鱼中发现的脊椎动物 组织者 同源基因,均证实文昌鱼存在类似脊椎动物的组织者 区域.文昌鱼G s c 基因空间表达谱数据显示[18]:原肠胚期该基因的表达集中在背部中内胚层,该区域对应于脊椎动物 组织者 ;神经胚早期该基因表达位于轴中胚层,将来发育成脊索;神经胚中期,除近尾端区域外,该基因在轴中胚层的表达完全消失.而在脊椎动物中,G s c 基因在原肠胚晚期的头部组织区域表达,随后在脊索前板特异表达[2,19],Y a s u o k a等[20]认为G s c 基因表达模式的改变导致脊索动物进化过程中头部的发育.文昌鱼脊索贯穿全身,前端缺乏调节脊椎动物大脑发育的脊索前板,其大脑结构十分简单,仅神经管最前端膨大形成脑泡;表达谱数据提示文昌鱼G s c 基因的功能与脊椎动物既有共同之处Copyright ©博看网. All Rights Reserved.第5期商留珂等:文昌鱼G s c 基因在早期胚胎发育中的功能h t t p :ʊjx m u .x m u .e d u .c n 又有差异,但尚缺乏实验证据.本研究通过活体注射m R N A 过表达文昌鱼G s c 基因,观察相应的胚胎形态结构并检测相关基因的表达情况,探讨文昌鱼G s c 基因在早期胚胎发育中的功能.1 材料与方法1.1 材 料佛罗里达文昌鱼(B r a n c h i o s t o m a f l o r i d a e )最初引种于Y u 实验室,在厦门大学王义权教授实验室已连续繁育多代.利用光周期结合热诱导的方法诱导性成熟的雌雄个体产卵排精[21].运用王义权教授实验室已建立的方法进行显微注射[22],以同一雌性个体所产生的部分卵不经注射直接人工受精作为对照.1.2 方 法1.2.1 目的基因质粒构建以王义权教授实验室已构建的G s c -pG E M -T 质粒为模板,以特异引物B b -G s c -P C R -F 1(5 -C G G G G T A C C A T G A C T A C C G T C A C C A G -3 ,下划线表示K pn Ⅰ酶切位点)和B b -G s c -P C R -R 1(5 -C G G A C T A G T T T A C T G T C C A T C G C T A G G -3,下划线表示S p e Ⅰ酶切位点)扩增同属的白氏文昌鱼(B .b e l c h e r i )G s c 基因的完整编码区序列;P C R 产物经双酶切回收后,连接到p X T 7载体的T 7启动子下游多克隆位点中,构建B b -G s c -p X T 7质粒.将果蝇E n gr a i l e d 基因编码转录抑制区域(E N )的序列[23-24]与编码G s c 蛋白D N A 结合区域(H D )的序列重组,由其转录出的m R N A 可以翻译得一嵌合蛋白,该嵌合蛋白能起到转录抑制因子的作用[25].以果蝇基因组D N A 为模板,用特异引物D m E n -E x p-F 2(5 -g gG G T A C C A T G G C C C T G G A G G A T C G C T G C A G -3 ,下划线表示K p n Ⅰ酶切位点)和D m E n -E x p -R 2(5 -g A C T A G T A G A T C T G A A T T C C T C G A G A G A T C C C A G A G C A G A T T T C T C T G -3,下划线分别表示S pe Ⅰ和X h o Ⅰ酶切位点),扩增E N 序列,P C R 产物经K p n Ⅰ和S pe Ⅰ双酶切回收后连接到p X T 7载体的T 7启动子下游多克隆位点中,构建E N -p X T 7质粒.以G s c -pG E M -T 质粒为模板,用引物F u s e d B b G s c -F 1(5 -c c gC T C G A G A T C G G C C A G G C G AC C C A T C C C G -3 ,下划线表示X h o Ⅰ酶切位点)和B b -G s c -P C R -R 1(5 -C G G A C T A G T T T A C T G T C C A TC G C T A G G -3 ,下划线表示S p e Ⅰ酶切位点),扩增G s c (H D )序列,P C R 产物纯化回收后连接到经相同的限制性内切酶(X h o Ⅰ及S pe Ⅰ)切割的E N -p X T 7载体中,构建E N -G s c (H D )-p X T 7质粒.1.2.2 m R N A 的合成重组质粒线性化后体外转录合成m R N A ,用其注射文昌鱼未受精的卵,随着胚胎发育进程可翻译成蛋白并发挥作用[22].将测序无误的B b -G s c -pX T 7和E N -G s c (H D )-pX T 7质粒用B a m HⅠ线性化,线性化产物经纯化回收后按照m E S S A G E m M A C H I N E 试剂盒(A m b i o n 公司)说明书操作,体外转录合成带帽的m R N A .经1%(质量分数)琼脂糖凝胶电泳检测,合成所得G s c m R N A 质量浓度为500n g /μL ,E N -G s c (H D )m R N A 质量浓度为200n g /μL .1.2.3 胚胎观察㊁固定及拍照活体观察比较注射G s c m R N A ㊁E N -G s c (H D )m R N A (注射组)和未经注射(对照组)的胚胎发育情况.选取数枚胚胎置于滴加1%(质量分数)甲基纤维素溶液的载玻片上,于倒置显微镜(I X 71,O l y m p u s 公司)下拍照记录.根据最新的文昌鱼胚胎发育时期划分标准[26],在胚胎发育的G 5原肠胚期和N 4神经胚期分别取G s c m R N A 注射组和对照组的胚胎数枚,用4%(质量分数)多聚甲醛-3-(N -吗啉基)丙磺酸固定,于4ħ冰箱保存备用.1.2.4 文昌鱼整胚原位杂交王义权教授实验室前期已制备所涉及基因的地高辛标记反义探针.原位杂交方法参照S k y y u 等[27]的报道稍作修改,将预杂交时间改为4h 以上,染色底物更换为B MP u r pl e (R o c h e 公司),待胚胎染色适宜后用含0.1%(体积分数)吐温-20的磷酸盐缓冲液洗涤终止染色,之后转移至80%(体积分数)丙三醇中,倒置显微镜下观察拍照.杂交后的胚胎长期保存于4ħ冰箱中.2 结果与分析2.1 G s c 基因过表达导致文昌鱼胚胎发育畸形如图1所示,G s c m R N A 注射并不影响文昌鱼早期胚胎发育,但当胚胎发育至G 5原肠胚期时,经过注射的胚胎开始呈现一致的异样表型,主要表现在:与对照组相比,胚胎发育明显滞后,且原肠胚的外胚层与中内胚层分离,贴合不紧密[图1(a )和(a ')];在N 1神经胚期注射组的胚体后端向背面弯曲[图1(b)和(b ')];随着胚胎发育,在N 1至N 4神经胚期,注射组胚胎体长明显小于对照组胚胎体长,并且胚胎前端膨大[图1(c )~(d ')];至T 1尾芽期,绝大多数注射组胚㊃958㊃Copyright ©博看网. All Rights Reserved.厦门大学学报(自然科学版)2023年h t t p :ʊjx m u .x m u .e d u .c n 胎不能形成口[图1(e )和(e ')];随着胚胎发育至L 幼体期,头部和尾部畸形严重,身体卷曲,无法形成鳃裂等器官[图1(f )~(g ')],最终由于胚胎严重畸形而逐渐死亡.图中标尺均为50μm ,(a )中标尺适用于(a )~(e '),(f )中标尺适用于(f )~(g').图1 注射G s c m R N A 后胚胎在发育不同时期的表型F i g .1P h e n o t y p e s o f t h e e m b r y o s i n j e c t e dw i t hG s c m R N Aa t d i f f e r e n t d e v e l o p m e n t a l s t a ge s 2.2 G s c 基因参与文昌鱼胚胎背腹轴建立及中胚层和尾芽发生G s c 基因过表达(G s c m R N A 注射)导致胚胎发育自G 5期开始出现内外胚层分离,提示其可能影响胚层分化.为进一步了解文昌鱼G s c 基因表达与胚层分化的关系,采用原位杂交方法检测G s c 基因过表达后,文昌鱼G 5期胚胎轴中胚层发育相关基因B r a c h y u r y和W n t 8[28]㊁背部标记基因C h o r d i n [17]和L e f t y [29],以及腹部标记基因V e n t [30-31]的表达情况.如图2所示:G s c 基因过表达导致B r a c h y u r y ㊁C h o r d i n ㊁L e f t y 和W n t 8基因表达水平下降[图2(a )~(h ')];而V e n t 基因表达水平上升,表达范围向背部扩张[图2(i )~(j')].上述结果提示G s c 基因可能在文昌鱼胚胎发育早期的背腹轴建立和中胚层发生过程中有重要作用.当文昌鱼胚胎发育至神经胚期后,在尾芽处不断形成新的体节和神经组织等,胚胎体长得以持续增加.G s c 基因过表达引起文昌鱼神经胚体长变短且后端弯曲,推测其可能还影响尾芽的发育.为此在G s c 基因过表达的神经胚中期(N 4期)胚胎中,分别检测体节标记基因m -a c t i n [32]㊁背部神经管和尾芽标记基因W n t 3[33],以及在进入神经胚期后又可作为脊索和尾芽标记基因的B r a c h y u r y 表达情况.如图3所示:在G s c m R N A 注射组胚胎中m -a c t i n 标记的体节数(5个)明显少于对照组胚胎的体节数(7个)[图3(a)和(a ')];B r a c h y u r y 和Wn t 3在G s c m R N A 注射组胚胎尾芽处的表达水平明显低于对照组胚胎[图3(b )~(c ')],W n t 3在G s c m R N A 注射组胚胎背部神经管的表达范围变宽[图3(c )和(c ')].2.3 文昌鱼G s c 可能为抑制型转录因子将E N -G s c (H D )m R N A 注射到文昌鱼未受精的卵中,受精后待受精卵分别发育至N 4㊁T 0和L 0期时,观察E N -G s c (H D )m R N A 注射对文昌鱼胚胎发育的影响.如图4所示,与G s c m R N A 注射相似,E N -G s c (H D )m R N A 注射同样引起胚胎弯曲㊁变短㊁前端膨大等表型,暗示文昌鱼G s c 基因可能编码抑制型转录因子.3 讨 论3.1 文昌鱼G s c 蛋白的转录调控活性G s c 作为抑制型转录因子,在爪蛙胚胎发育中可以抑制其下游基因W n t 8的表达,维持胚胎前端正常发育[34].果蝇E n gr a i l e d 也是一种抑制型转录因子,将其转录抑制区域[23-24]与目的转录因子D N A 结合域融合构成嵌合蛋白,可用于分析目的转录因子是抑制型㊃068㊃Copyright ©博看网. All Rights Reserved.第5期商留珂等:文昌鱼G s c 基因在早期胚胎发育中的功能h t t p :ʊjx m u .x m u .e d u .cn 图中标尺为50μm .各图右下角数字含义:呈如图所示表达模式的胚胎数/该组用于原位杂交的胚胎总数(图3同).图2 G s c m R N A 注射后G 5期胚胎标记基因的表达模式F i g .2M a r k e r g e n e e x p r e s s i o n p a t t e r n s i n t h e e m b r y o s i n j e c t e dw i t hG s c m R N Aa t G 5s t a ge 以上均为侧面观,图中标尺为50μm .图3 G s c m R N A 注射后N 4期胚胎标记基因的表达模式F i g .3M a r k e r g e n e e x p r e s s i o n p a t t e r n s i n t h e e m b r y o s i n j e c t e dw i t hG s c m R N Aa tN 4s t a ge 还是激活型.使用类似方法,本研究发现文昌鱼G s c 基因过表达与胚胎显微注射E N -G s c (H D )m R N A 均可导致文昌鱼胚胎出现一致的表型,暗示文昌鱼G s c 基因编码抑制型转录因子,在胚胎发育早期对下游基因的表达可能有负调控作用.3.2 文昌鱼G s c 基因参与胚胎中胚层形成和后端生长文昌鱼胚胎发育过程与脊椎动物类似,也会形成㊃168㊃Copyright ©博看网. All Rights Reserved.厦门大学学报(自然科学版)2023年h t t p :ʊjx m u .x m u .e d u .cn 图中标尺为100μm .图4 注射E N -G s c (H D )m R N A 后不同发育时期的胚胎表型F i g .4P h e n o t y p e s o f t h e e m b r y o s i n je c t e dw i t h E N -G s c (H D )m R N Aa t d if f e r e n t d e v e l o p m e n t a l s t a ge s 由中胚层细胞分化而来的脊索和体节等组织器官,通过尾芽不断分化形成新体节㊁神经管等组织,使体长不断增加(即后端生长).但与脊椎动物不同的是,文昌鱼胚胎不形成明确的脊索前板及与之相关的大脑[35].基于爪蛙㊁斑马鱼(D a n i o r e r i o )等脊椎动物模型的研究表明,G s c ㊁B r a c h y u r y ㊁W n t 8和骨形成蛋白(B M P )信号是促进脊椎动物胚胎上述结构发生的重要参与者.在文昌鱼胚胎中,B r a c h y u r y 和W n t 8基因的表达,以及B M P 信号的活性区域与其在脊椎动物中的同源基因或信号相似,集中在中胚层祖细胞及其衍生组织中,暗示它们在文昌鱼胚胎中胚层祖细胞分化及其衍生组织的形成中具有保守的功能.本研究中G s c m R N A 过表达结果显示,在文昌鱼胚胎中G s c 基因对B r a c h y u r y 和Wn t 8基因的表达同样具有抑制作用.然而,虽然G s c 基因在文昌鱼原肠胚及更早时期胚胎中的表达谱与脊椎动物胚胎的表达谱相似(均表达于胚胎背侧),但是在原肠胚之后并没有特异性地出现在脊索前板,而是持续表达于尾芽部分细胞中[18].这种表达差异提示G s c 基因获得脊索前端的表达可能是脊椎动物产生脊索前板及复杂大脑结构的关键,同时说明G s c 基因在文昌鱼尾芽功能维持中扮演重要角色.本研究在文昌鱼胚胎中过表达G s c 基因可导致B r a c h y u r y 和Wn t 3基因在尾芽处的表达下降,胚胎后端延伸无法正常发生(体节数减少),以及神经管膨大.脊椎动物胚胎后端生成主要依赖尾芽处的神经中胚层祖细胞,B r a c h y u r y 基因功能异常会导致该类群细胞向中胚层分化的能力减弱,向神经外胚层分化的能力增强[36];而文昌鱼同样存在后端生长,且B r a c h y u r y 基因特异表达于文昌鱼尾芽细胞中[28,37],推测文昌鱼后端延伸机制可能与脊椎动物类似,都依赖B r a c h y u r y 基因,而G s c 基因过表达引起的文昌鱼后端延伸异常很可能是通过其对B r a c h y u r y 基因表达的抑制引起的.但需要注意的是,上述结果也可能是由于G s c 基因过表达对早期中胚层分化影响而间接造成的,阐明确切机制仍需更多精细研究.3.3 文昌鱼G s c 基因参与胚胎背腹建立的机制与脊椎动物不同位于原肠胚背侧背唇区域的组织者 是调控脊椎动物背腹轴建立的信号中心.G s c 基因是脊椎动物 组织者 最特异的标记基因之一,也是赋予 组织者 强大功能的关键基因.在爪蛙和斑马鱼等脊椎动物胚胎中过表达G s c 基因可导致 组织者 变大,背侧组织向腹侧扩张(背部化);而敲低(或敲除)G s c 基因表达则会使 组织者 变小,腹侧组织向背侧扩张(腹部化).机制分析显示,过表达G s c 基因引起B M P 信号抑制因子C h o r d i n 表达区域从背侧向腹侧扩张,使得腹侧的B M P 信号活性减弱,最终导致胚胎出现背部化表型;而敲低(或敲除)G s c 基因表达则引起相反的结果[38].本研究结果显示,文昌鱼也有类似脊椎动物的 组织者 区域,在文昌鱼胚胎中过表达G s c 基因同样会影响其背腹轴建立,但效果却与脊椎动物相反:C h o r d i n (及L e f t y )在原肠胚背侧( 组织者 区域)表达量下降,而B M P 信号通路下游基因V e n t 的表达从胚胎腹侧向背侧扩张.引起这种差异的原因目前尚不清楚,推测这很可能是在两个类群生物中G s c 对C h o r d i n 基因表达调控机制上有差异所致:在脊椎动物中G s c 促进C h o r d i n 表达,而在文昌鱼中G s c 抑制C h o r d i n 表达.这种同源基因在头索动物和脊椎动物两个不同进化类群中的作用方式相反的现象,是否反映出进化上两个动物类群分化时选择了不同的体轴调控方式,或反映出调控胚胎体轴发生的信号网络的演化,都是有待深入研究的科学问题.参考文献:[1] S P E M A N N H ,M A N G O L D H.Üb e ri n d u k t i o n v o n㊃268㊃Copyright ©博看网. All Rights Reserved.第5期商留珂等:文昌鱼G s c 基因在早期胚胎发育中的功能h t t p :ʊjx m u .x m u .e d u .c n e m b r y o n a l a n l a g e nd u r c h i m p l a n t a t i o na r t f r e m d e ro r ga n i -s a t o r e n [J ].A r c h i vf ür M i k r o s k o pi s c h e A n a t o m i eu n d E n t w i c k l u n gs m e c h a n i k ,1924,101(1):458.[2] C H OK W Y ,B L U M B E R GB ,S T E I N B E I S S E R H ,e t a l .M o l e c u l a r n a t u r eo fS p e m a n n o r ga n i z e r :t h er o l eo f t h e X e n o pu s h o m e o b o x g e n e G o o s e c o i d [J ].C e l l ,1991,67(6):1111-1120.[3] G A U N TS J ,B L U M M ,D ER O B E R T I SEM.E x pr e s s i o n o f t h em o u s e G o o s e c o i d g e n ed u r i n g m i d -e m b r y o ge n e s i s m a y m a r k m e s e n c h y m a lc e l l l i n e a g e si nt h ed e v e l o p i n gh e a d ,l i m b sa n db o d y w a l l [J ].D e v e l o p m e n t ,1993,117(2):769-778.[4] S C H U L T E -M E R K E R S ,H A MM E R S C HM I D T M ,B E U C H L E D ,e ta l .E x p r e s s i o no fz e b r a f i s h G o o s e c o i d a n d n o t a i l g e n e p r o d u c t s i nw i l d -t y pe a n dm u t a n t n o t a i l e m b r y o s [J ].D e v e l o pm e n t ,1994,120(4):843-852.[5] R I V E R A -P ÉR E Z JA ,M A L L O M ,G E N D R O N -M A G U I R E M ,e t a l .G o o s e c o i d i sn o t a ne s s e n t i a l c o m po n e n t o f t h e m o u s e g a s t r u l ao r g a n i z e rb u t i s r e qu i r e df o r c r a n i o f a c i a l a n dr i bd e v e l o p m e n t [J ].D e v e l o pm e n t ,1995,121(9):3005-3012.[6] Y A M A D A G ,M A N S O U R I A ,T O R R E S M ,e t a l .T a r ge t e dm u t a t i o nof t h em u r i n e G o o s e c o i dg e n e r e s u l t s i n c r a n i o f a c i a l d e f e c t s a n d n e o n a t a l d e a th [J ].D e v e l o pm e n t ,1995,121(9):2917-2922.[7] B E L OJA ,L E Y N SL ,Y A M A D A G ,e t a l .T h e p r e c h o r d a lm i d l i n e o f t h e c h o n d r o c r a n i u m i s d e f e c t i v e i n G o o s e c o i d -1m o u s em u t a n t s [J ].M e c h a n i s m s o f D e v e l o p m e n t ,1998,72(1/2):15-25.[8] F I L O S AS ,R I V E R A -P ÉR E ZJA ,G ÓM E Z A P ,e t a l .G o o s e c o i d a n d HN F -3βge n e t i c a l l y i n t e r a c tt or e g u l a t e n e u r a l t u b e p a t t e r n i n g d u r i n g m o u s e e m b r y o ge n e s i s [J ].D e v e l o p m e n t ,1997,124(14):2843-2854.[9] H A H N M ,J ÄC K L EH .D r o s o ph i l a G o o s e c o i d p a r t i c i p a t e s i n n e u r a l d e v e l o p m e n t b u t n o t i n b o d ya x i s f o r m a t i o n [J ].T h eE M B OJ o u r n a l ,1996,15(12):3077-3084.[10] S T E I N B E I S S E R H ,F A I N S O D A ,N I E H R SC ,e t a l .T h er o l e o f G s c a n d B M P -4i nd o r s a l -v e n t r a l p a t t e r n i n g of t h em a rg i n a l z o n e i n X e n o pu s :a l o s s -o f -f u n c t i o ns t u d y u s i n g an t i s e n s eR N A [J ].T h eE M B OJ o u r n a l ,1995,14(21):5230-5243.[11] B L A I RJE ,H E D G E SS B .M o l e c u l a r p h y l o g e n y an d d i v e r ge n c e t i m e s of d e u t e r o s t o m e a n i m a l s [J ].M o l e c u l a r B i o l og y an dE v o l u t i o n ,2005,22(11):2275-2284.[12] D E L S U CF ,B R I N K M A N N H ,C H O U R R O U TD ,e t a l .T u n i c a t e s a n d n o t c e p h a l o c h o r d a t e s a r e t h e c l o s e s t l i v i n gr e l a t i v e s o fv e r t e b r a t e s [J ].N a t u r e ,2006,439(7079):965-968.[13] P U T N A M NH ,B U T T ST ,F E R R I E RDEK ,e t a l .T h e a m ph i o x u s g e n o m ea n dt h ee v o l u t i o no ft h ec h o r d a t e k a r y o t y pe [J ].N a t u r e ,2008,453(7198):1064-1071.[14] B E R T R A N DS ,E S C R I V A H.E v o l u t i o n a r y cr o s s r o a d s i n d e v e l o p m e n t a l b i o l o g y :a m p h i o x u s [J ].D e v e l o p m e n t ,2011,138(22):4819-4830.[15] S H I M E L DSM ,H O L L A N DND .A m ph i o x u sm o l e c u l a r b i o l o g y :i n s i g h t s i n t ov e r t e b r a t ee v o l u t i o na n dd e v e l o p-m e n t a lm e c h a n i s m s [J ].C a n a d i a nJ o u r n a lo fZ o o l o g y ,2005,83(1):90-100.[16] T U N G Y F ,Y E H Y .E x pe r i m e n t a ls t u d i e so nn e u r a l i n d u c t i o n i n a m p h i o x u s [J ].S c i e n t i a S i n i c a ,1961,11(6):805-820.[17] Y U J K ,S A T O U Y ,H O L L A N D N D ,e ta l .A x i a l p a t t e r n i n g i nc e ph a l o c h o r d a t e s a n d t h e e v o l u t i o no f t h e o r ga n i z e r [J ].N a t u r e ,2007,445(7128):613-617.[18] N E I D E R T A H ,P A N O P O U L O U G ,L A N G E L A N DJ A .A m ph i o x u s G o o s e c o i d a n d t h e e v o l u t i o no f t h eh e a d o r ga n i z e r a n d p r e c h o r d a l p l a t e [J ].E v o l u t i o n &D e v e l o p m e n t ,2000,2(6):303-310.[19] S U D O U N ,Y A M A M O T OS ,O G I N O H ,e t a l .D yn a m i c i n v i v o b i n d i n g o f t r a n s c r i p t i o n f a c t o r s t o c i s -r e g u l a t o r ym o d u l e s o f c e r a n d g s c i n t h e s t e pw i s e f o r m a t i o no f t h e S p e m a n n -M a n g o l d o r g a n i z e r [J ].D e v e l o pm e n t ,2012,139(9):1651-1661.[20] Y A S U O K A Y ,T A N D O Y ,K U B O K A W A K ,e ta l .E v o l u t i o n o f c i s -r e g u l a t o r y mo d u l e s f o r t h e h e a d o r g a n i z e r g e n e G o o s e c o i d i n c h o r d a t e s :c o m p a r i s o n s b e t w e e n B r a n c h i o s t o m a a n d X e n o pu s [J ].Z o o l o g i c a l L e t t e r s ,2019,5:27.[21] L IG ,S H UZH ,W A N GY Q .Y e a r -r o u n d r e pr o d u c t i o n a n d i n d u c e d s p a w n i n g o f C h i n e s e a m p h i o x u s ,B r a n c h i o s t o m a b e l c h e r i ,i nl a b o r a t o r y [J ].P L o S O n e ,2013,8(9):e 75461.[22] L I U X ,L IG ,F E N GJ ,e t a l .A n e f f i c i e n tm i c r o i n je c t i o n m e t h o df o r u n f e r t i l i z e d eg g s o f A s i a n a m ph i o x u s B r a n c h i o s t o m ab e l c h e r i [J ].D e v e l o pm e n t G e n e s a n d E v o l u t i o n ,2013,223(4):269-278.[23] J A Y N E SJB ,O F A R R E L LP H.A c t i v e r e pr e s s i o no f t r a n s c r i p t i o n b y t h e e n gr a i l e d h o m e o d o m a i n p r o t e i n [J ].T h eE M B OJ o u r n a l ,1991,10(6):1427-1433.[24] H A N K ,M A N L E Y J L .F u n c t i o n a ld o m a i n so ft h eD r o s o p h i l aE n g r a i l e d p r o t e i n [J ].T h eE M B OJ o u r n a l ,1993,12(7):2723-2733.[25] C O N L O NFL ,S E D G W I C KSG ,W E S T O NK M ,e t a l .I n h i b i t i o n o f X b r a t r a n s c r i pt i o n a c t i v a t i o n c a u s e s d e f e c t s i nm e s o d e r m a l p a t t e r n i n g a n dr e v e a l sa u t o r e gu l a t i o no f X b r a i nd o r s a lm e s o d e r m [J ].D e v e l o p m e n t ,1996,122㊃368㊃Copyright ©博看网. All Rights Reserved.厦门大学学报(自然科学版)2023年h t t p :ʊjx m u .x m u .e d u .c n (8):2427-2435.[26] C A R V A L H OJE ,L A H A Y EF ,Y O N GL W ,e t a l .A nu p d a t e ds t a g i n g s y s t e mf o rc e p h a l o c h o r d a t ed e v e l o p m e n t :o n e t a b l e s u i t s t h e ma l l [J ].F r o n t i e r s i nC e l l a n dD e v e -l o p m e n t a l B i o l o g y,2021,9:668006.[27] S K Y Y UJK ,H O L L A N DLZ .A m ph i o x u sw h o l e -m o u n t i n s i t u h y b r i d i z a t i o n [J ].C o l dS p r i n g H a r b o rP r o t o c o l s ,2009,2009(9):p d b .pr o t 5286.[28] H O L L A N DP W ,K O S C H O R ZB ,H O L L A N DLZ ,e ta l .C o n s e r v a t i o n o f B r a c h y u r y (T )g e n e s i n a m p h i o x u s a n dv e r t eb r a t e s :d e v e l o p m e n t a l a n de v o l u t i o n a r y i m pl i -c a t i o n s [J ].D e v e l o p m e n t ,1995,121(12):4283-4291.[29] O N A IT ,Y UJK ,B L I T ZIL ,e t a l .O p p o s i n g No d a l /V g 1a n d B M P s i g n a l s m e d i a t e a x i a l p a t t e r n i n g in e m b r y o s o f t h eb a s a l c h o r d a t e a m p h i o x u s [J ].D e v e l o p -m e n t a l B i o l o g y,2010,344(1):377-389.[30] P A N O P O U L O UGD ,C L A R K MD ,H O L L A N DLZ ,e ta l .A m p h i B M P 2/4,a na m p h i o x u sb o n e m o r p h o ge n e t i c p r o t e i nc l o s e l y r e l a t e dt o D r o s o p h i l ad e c a p e n t a p l e gi c a n d v e r t e b r a t e B M P 2a n dB M P 4:i n s i g h t s i n t o e v o l u t i o n o fd o r s o v e n t r a la x i ss p e c i f i c a t i o n [J ].D e v e l o pm e n t a l D yn a m i c s ,1998,213(1):130-139.[31] S C H U B E R T M ,H O L L A N D L Z ,P A N O P O U L O U GD ,e ta l .C h a r a c t e r i z a t i o n o fa m p h i o x u s A m ph i W n t 8:i n s i g h t s i n t o t h e e v o l u t i o n o f p a t t e r n i n g o f t h e e m b r yo n i c d o r s o v e n t r a l a x i s [J ].E v o l u t i o n&D e v e l o p m e n t ,2000,2(2):85-92.[32] K U S A K A B E R ,K U S A K A B E T ,S A T O H N ,e ta l .D i f f e r e n t i a l g e n e e x pr e s s i o n a n di n t r a c e l l u l a r m R N A l o c a l i z a t i o n o f a m p h i o x u s a c t i n i s o f o r m s t h r o u gh o u t d e v e l o p m e n t :i m p l i c a t i o n s f o r c o n s e r v e d m e c h a n i s m so f c h o r d a t e d e v e l o p m e n t [J ].D e v e l o pm e n t G e n e s a n d E v o l u t i o n ,1997,207(4):203-215.[33] S C H U B E R T M ,H O L L A N DLZ ,S T O K E SM D ,e t a l .T h r e e a m p h i o x u s W n t g e n e s (A m p h i W n t 3,A m p h i W n t 5,a n d A m ph i W n t 6)a s s o c i a t e dw i t h t h e t a i l b u d :t h e e v o l u t i o n o f s o m i t o g e n e s i s i nc h o r d a t e s [J ].D e v e l o p m e n t a lB i o l o g y ,2001,240(1):262-273.[34] Y A O J ,K E S S L E RDS .G o o s e c o i d p r o m o t e s h e a d o r g a n i z e r a c t i v i t y b y d i r e c tr e p r e s s i o n o f X w n t 8i n S p e m a n n s o r g a n i z e r [J ].D e v e l o p m e n t ,2001,128(15):2975-2987.[35] F E R R A NJ L ,I R I M I A M ,P U E L L E S L .I st h e r eap r e c h o r d a l r e gi o n a n d a n a c r o t e r m i n a l d o m a i n i n a m p h i o x u s ?[J ].B r a i n ,B e h a v i o r a n dE v o l u t i o n ,2022,96(4/5/6):334-352.[36] K O C HF ,S C H O L Z E M ,W I T T L E RL ,e t a l .A n t a g o n i s t i c a c t i v i t i e s o f S o x 2a n d B r a c h y u r y co n t r o l t h e f a t e c h o i c e o fn e u r o -m e s o d e r m a l p r o g e n i t o r s [J ].D e v e l o p m e n t a l C e l l ,2017,42(5):514-526.[37] Y U A N L ,W A N G Y Q ,L IG .D i f f e r e n t i a le x pr e s s i o n p a t t e r n o f t w o B r a c h y u r y g e n e s i na m p h i o x u s e m b r y o s [J ].G e n eE x pr e s s i o nP a t t e r n s ,2020,38:119152.[38] S A N D E R V ,R E V E R S A D E B ,D E R O B E R T I S E M.T h e o p p o s i n g ho m e o b o x g e n e s G o o s e c o i d a n d V e n t 1/2s e l f -r e g u l a t e X e n o pu s p a t t e r n i n g [J ].T h eE M B OJ o u r n a l ,2007,26(12):2955-2965.F u n c t i o no fG s c g e n e i n e a r l y e m b r y o g e n e s i s o f a m ph i o x u s S H A N GL i u k e 1,2,L I U H u i m i n 2,S H I C h e n g g a n g 2,L IG u a n g 2,W A N GY i qu a n 2*(1.S c h o o l o fM e d i c a l T e c h n o l o g y ,L u o y a n g V o c a t i o n a l&T e c h n i c a l C o l l e g e ,L u o y a n g 471000,C h i n a ;2.S c h o o l o f L i f e S c i e n c e s ,X i a m e nU n i v e r s i t y ,X i a m e n 361102,C h i n a )A b s t r a c t :T h e G o o s e c o i d (G s c ),a h o m e o b o x g e n e ,p l a y s a n i m p o r t a n t r o l e i n t h e f o r m a t i o no f "o r ga n i z e r "a n d p r e c h o r d a l p l a t e i n v e r t eb r a t e s .T o i n v e s t i g a t e i t s f u nc t i o n a l e v o l u t i o n i n c h o rd a te ,w e a n a l y z e d t h ef u n c t i o n o f G s cg e n e i n e m b r y o g e n e s i s o f a m ph i o x u s v i a o v e r -e x p r e s s i o n a n dw h o l e i n s i t u h y b r i d i z a t i o n .O u r r e s u l t s s h o w e d t h a t t h e i n je c t i o nof G s c m R N Ao r E N -G s c (H D )m R N A c o d i ng a t r a n s c r i p t i o n r e p r e s s o r y i e l d e d s i m i l a r b o d y a x i s d e f o r m i t y ,i n d i c a t i n g th a t G s c p l a y s a n e s s e n ti a l r o l e i n a m p h i o x u s e m b r y o g e n e s i s a s a r e p r e s s i v e t r a n s c r i p t i o n a l f a c t o r .G e n ee x p r e s s i o na n a l y s i ss u g g e s t e dt h a t G s c o v e r e x p r e s s i o ne x p a n d e dt h ee x pr e s s i o no f V e n t m a r k i n g t h e v e n t r a l i d e n t i t i e s a n dr e d u c e d t h e e x p r e s s i o no f B r a c h y u r y a n d W n t 8r e l a t e d t om e s o d e r mf o r m a t i o n ,a sw e l l a s t h e e x p r e s s i o no f C h o r d i n a n d L e f t y ma r k i n g t h e d o r s a l s t r u c t u r e s a t g a s t r u l a t i o n s t a g e .G s c m R N A i n j e c t i o n a l s o r e d u c e d t h e e x p r e s s i o n o f B r a c h y u r y inm e s o d e r m p r o g e n i t o r c e l l s o f t a i l b u d ,s o m i t em a r k e r g e n e m -a c t i n i n s o m i t e s a n dn e u r a l t u b e a n d t a i l b u dm a r k e r g e n e W n t 3i n t a i l b u d ,b u t e x p a n d e d t h e e x p r e s s i o no f W n t 3i nn e u r a l t u b e .T h e a b o v e r e s u l t s d e m o n s t r a t e t h a t G s c g e n e p l a ys a n e s s e n t i a l r o l e i n b o d y a x i s e s t a b l i s h m e n t a n dm e s o d e r mf o r m a t i o n ,a n d a l s o i nm a i n t a i n i n g t h e f u n c t i o n o f t a i l b u d i n a m p h i o x u s .K e yw o r d s :a m p h i o x u s ;G s c g e n e ;m e s o d e r mf o r m a t i o n ;e m b r y o g e n e s i s (责任编辑:徐婷婷)㊃468㊃Copyright ©博看网. All Rights Reserved.。
《文昌鱼的胚胎》课件

多种调控机制参与文昌鱼胚胎的基因表达,包括DNA甲基化、组蛋白修饰和 microRNA的调控等。这些调控机制共同作用,确保胚胎发育过程中的基因表 达的精确性。
细胞分裂与分化
细胞分裂
在文昌鱼胚胎发育过程中,细胞通过 有丝分裂的方式进行增殖,形成更多 的细胞。
细胞分化
胚胎中的细胞通过分化成为特定的组 织或器官。在文昌鱼中,细胞分化的 过程受到多种信号分子的调控,如生 长因子、激素和细胞因子等。
器官发生
文昌鱼胚胎发育过程中, 各器官逐渐形成,但与脊 椎动物相比,其器官发生 过程存在差异。
文昌鱼胚胎发育与脊椎动物的联系
神经管形成
文昌鱼胚胎发育过程中,神经管的形成是其与脊椎动物相似的重 要特征之一。
细胞分化
文昌鱼胚胎在发育过程中出现了高度细胞分化,这与脊椎动物的细 胞分化过程具有一定的相似性。
提供生物进化的证据
通过研究文昌鱼胚胎发育的特点,可以发现其在生物进化过程中的地位和作用,为生物进化提供有力 的证据。
胚胎发育机制研究
探索胚胎发育的奥秘
文昌鱼胚胎发育过程中涉及多种复杂的生物学过程,研究这些过程有助于深入了解胚胎发育的机制。
发现新的发育生物学规律
通过对文昌鱼胚胎发育的研究,以发现一些新的发育生物学规律,为人类和其他动物的胚胎发育研究提供借鉴 。
囊胚阶段
囊胚阶段是胚胎发育的一个重 要阶段,此时胚胎细胞开始分 化成不同的组织。
囊胚呈球形,细胞数量增多, 胚胎内部的细胞开始出现不同 的分化趋势。
囊胚阶段的发育对于后续胚胎 的发育至关重要,是决定胚胎 未来发育方向的关键时期。
原肠胚阶段
01
原肠胚阶段是胚胎发育的一个重要转折点,此时胚胎开始形成 三个胚层。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
实验材料:
文昌鱼早期胚胎发育切片: 文昌鱼受精卵装片, 文昌鱼2细胞装片, 文昌鱼4细胞装片, 文昌鱼8细胞装片, 文昌鱼16细胞装片, 文昌鱼囊胚期装片。 实验仪器:光学显微镜 文昌鱼组织结构切片: 文昌鱼整体装片, 文昌鱼过口笠横切, 文昌鱼过咽横切, 文昌鱼过肠横切, 文昌鱼过性腺横切, 文昌鱼尾部横切。
受精、卵裂及原肠胚的形成
桑椹胚一方面继续地分裂,一方面中心的细胞渐向表面迁移,因而变 成一个空心的圆球,此时期的胚胎称囊胚(blastula),中间的空腔内充满 胶状液体,称囊胚腔(blastocoel)。囊胚上端的细胞稍小,称动物极 (animal pole),下端的细胞稍大,称植物极(vegetative pole)。 以后囊胚植物极的大细胞以内陷的方式向囊胚腔陷入,正像一个漏了 气的皮球,以至和上部动物极的细胞相贴近,囊胚腔被挤掉,而另形成一 个新的腔,称为原肠腔(archenteron)。原肠腔以原口(blastopore)与 外界相通。此时胚胎具备内外两层细胞,贴着原肠腔的一层细胞,称内胚 层(endoderm),与外界相接触的一层细胞,称为外胚层(ectoderm), 此阶段称Байду номын сангаас肠胚(gas- trula)。
作业:
1. 绘制文昌鱼早期胚胎发育图片,受精卵,2细胞, 4细胞,8细胞,16细胞,囊胚期任选一个。 2. 绘制文昌鱼整体装片,并附上详细说明。 3. 绘制文昌鱼的横切图一张(任选),并附上详 细说明。
文昌鱼发育所经历的主要阶段如下:
受精卵——桑椹胚——囊胚——原肠胚——神经胚——幼体——成体。 文昌鱼在六、七月间为产卵期,产卵和受精均在傍晚进行。受精卵 不久即开始分裂,由于卵黄少,为均黄卵(isolecithal egg),因此卵 的分裂为全分裂(holoblastic),且为等分裂。第一次和第二次分裂皆 为垂直分裂,分成四个细胞,第三次分裂为横分裂,分成上下八个细胞, 如此继续分裂下去,许多细胞结成一个实心的圆球,称桑椹胚 (morula)。
实验原理
文昌鱼是脊索动物门头索动物亚门的重要 代表,成体文昌鱼具备脊椎动物的三大特征: 脊索、背神经管和咽鳃裂。
实验原理
研究文昌鱼的胚胎发育具有很重要的意义,因为 文昌鱼是以简单而典型的形式代表着脊椎动物的发育, 把文昌鱼这一简明图案了解清楚,对于了解高等脊椎 动物较复杂的发育有很大的方便。 另一方面,文昌鱼的早期发育又与棘皮动物很相 似,由个体发育可以看到它与棘皮动物的关系。