南京师范大学 环境生态学(王国祥)
溢流堰影响下溶解氧与氨氮及CODMn的关系
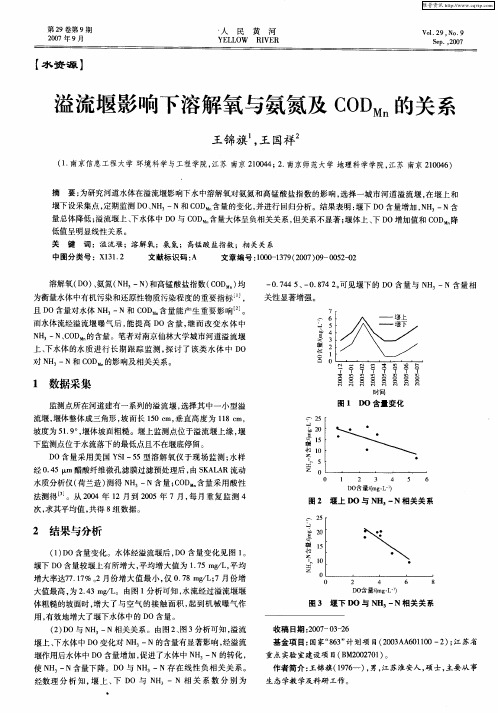
一
0 745 一 .7 o .4 、 0 842 可见堰下的 D O含量 与 N ,一N含量 相 H
关性 显著 增强。
一 . |、 如 h n壬 一 【1 l 一 J } 。
∞
N , N、O 的含量 。笔者对南京仙林 大学城市河道溢 流堰 H 一 CD
g
上、 下水体的水质 进行 长期 跟踪 监测 , 讨 了该类 水 体 中 D 探 O
重点 实验室建设项 目( M 02 0 ) B 2071 。
作者简 介 : 王锦旗( 96 ) 男, 17 一 , 江苏淮安人, 士, 硕 主要从事
生态学教 学及科研 工作。
维普资讯
第 9期
王锦旗等 : 溢流堰影 响下 溶解 氧与氨氮及 C D O 的关系
对 N 一N和 C D H, O 的影 响及相关关系 。
0
0
1 数据采集
监测点所在河道建有一系列的溢流堰 , 选择其 中一 小型溢
流堰 , 堰体整体成三角形 , 面长 10e 垂 直高度 为 18e 坡 5 m, 1 m, 坡度为 5 . 。堰 体坡 面粗糙 。堰上监测点位于溢流堰上缘 , 19 , 堰 下监测 点位 于水流落下 的最低点且不在堰底停留。 D O含量 采用美 国 Y I 5型溶 解氧仪 于现场监 测 ; 样 S 一5 水 经 0 4 m醋酸纤维微孔滤膜过滤预处理后 , S A A .5 由 K L R流动 水质分析仪( 荷兰造) 测得 N ,一N含量 ; O 含量采 用酸性 H CD 法测得 。从 20 04年 1 到 20 2月 0 5年 7月 , 月重 复监 测 4 每
增大率达7 . %。 71 2月份增大值 最小 , 07 gL7月份增 7 仅 .8m / ;
盐城海滨湿地不同盐沼土壤呼吸特征及温度响应

盐城海滨湿地不同盐沼土壤呼吸特征及温度响应徐杰;苏海蓉;余培培;王国祥;刘金娥【期刊名称】《生态与农村环境学报》【年(卷),期】2017(033)008【摘要】测定了盐城海滨湿地芦苇(Phragmites australis)盐沼、互花米草(Spartina alterniflora)盐沼和碱蓬(Suaeda salsa)盐沼土壤呼吸速率,结合盐沼土壤室内培养实验,比较3种盐沼土壤呼吸特征及温度响应.结果表明:(1)实验期内3种盐沼土壤呼吸均为CO2排放状态,芦苇盐沼、互花米草盐沼和碱蓬盐沼土壤呼吸速率平均值分别为323.67、499.51和150.41 mg·m-2·h-1.3种盐沼土壤呼吸均呈现明显的季节性差异,由高到低依次为夏季、春季和秋季.(2)野外实验中,3种盐沼土壤呼吸出现峰值时的土壤温度由高到低依次为芦苇盐沼(28℃)、互花米草盐沼(22℃)和碱蓬盐沼(20℃).室内培养条件下,土壤呼吸速率与土壤温度呈显著正相关(P<0.05),对温度变化的敏感性由大到小依次为芦苇盐沼(2.19)、碱蓬盐沼(2.15)和互花米草盐沼(1.58).(3)实验期内3种盐沼土壤呼吸累积排放量由大到小依次为互花米草盐沼(3400 g·m-2·a-1)、芦苇盐沼(2330 g·m-2·a-1)和碱蓬盐沼(1082 g·m-2·a-1),不同季节累积排放量由大到小依次均为夏季、春季和秋季.【总页数】7页(P715-721)【作者】徐杰;苏海蓉;余培培;王国祥;刘金娥【作者单位】南京师范大学环境学院/ 江苏省地理信息资源开发与利用协同创新中心, 江苏南京 210023;南京师范大学环境学院/ 江苏省地理信息资源开发与利用协同创新中心, 江苏南京 210023;南京师范大学环境学院/ 江苏省地理信息资源开发与利用协同创新中心, 江苏南京 210023;南京师范大学环境学院/ 江苏省地理信息资源开发与利用协同创新中心, 江苏南京 210023;南京师范大学环境学院/ 江苏省地理信息资源开发与利用协同创新中心, 江苏南京 210023【正文语种】中文【中图分类】Q148【相关文献】1.锡林河湿地土壤呼吸日变化及温度响应特征研究 [J], 王慧清;王云龙;杜金玲2.盐城海滨湿地土壤水分空间特征与植被关系研究 [J], 韩爽;张娇;陆大培;张华兵3.江苏盐城原生盐沼湿地表层水体Hg、As分布特征及评价 [J], 赵雪琴;赵善道;左平;腾厚锋;耿金菊;高翔4.盐城海滨湿地盐沼植被及农作物下土壤酶活性特征 [J], 毛志刚;谷孝鸿;刘金娥;任丽娟;王国祥5.盐城海滨湿地盐沼植被对土壤碳氮分布特征的影响 [J], 毛志刚;王国祥;刘金娥;任丽娟因版权原因,仅展示原文概要,查看原文内容请购买。
沉积物氮形态与测定方法研究进展

沉积物氮形态与测定方法研究进展刘波;周锋;王国祥;许宽;杜旭;凌芬;夏劲【期刊名称】《生态学报》【年(卷),期】2011(031)022【摘要】长期以来,国内外学者对沉积物中氮进行了大量的研究,在氮生物地球化学循环和生态学效应方面取得了重要进展.然而,现有关于氮赋存形态的研究主要集中在总氮和无机氮方面,还不能深入阐明沉积物氮的生物和生态学机制.分析了沉积物和土壤氮赋存形态划分和测定方法的研究进展,研究表明:沉积物氮的形态划分与测定方法基本上还是借鉴了土壤氮的研究方法;无机态氮的研究多集中在可交换态氮方面,对固定铵的研究相对较少;在可交换态氮提取方法上并没有针对沉积物与土壤的差异进行必要的论证和改进,沉积物中可溶态氮对可交换态氮测定的影响还不明确;有机氮的测定方法基本上是经验方法,目前还无针对有机氮生态学效应的分类及测定方法;连续分级浸提方法从生态学效应的角度对沉积物氮的研究进行了有益的探索,为深入揭示氮的生态学机制提供了新的思路,但是此类方法目前还集中在国内学者的相关研究中.【总页数】12页(P6947-6958)【作者】刘波;周锋;王国祥;许宽;杜旭;凌芬;夏劲【作者单位】南京师范大学地理科学学院,南京210046;南通大学地理科学学院,南通226007;南京师范大学地理科学学院,南京210046;南京师范大学地理科学学院,南京210046;南京师范大学地理科学学院,南京210046;南京师范大学地理科学学院,南京210046;南京师范大学地理科学学院,南京210046;南京师范大学地理科学学院,南京210046【正文语种】中文【相关文献】1.宜兴三氿沉积物氮形态分布及其影响因素 [J], 邹民虎;连斌;王晓昌;贺拓2.梯级水库建设对怒江与澜沧江沉积物氮形态分布的影响 [J], 朱晓声;郭小娟;王耀耀;何金艳;杨正健;纪道斌;马骏;刘德富3.外源添加对不同入湖河流沉积物氮形态变化规律的影响 [J], 韩年; 袁旭音4.黏性土弱透水层氮形态的赋存特征及迁移转化——以江汉平原沉湖沉积物为例[J], 张董涛;刘璐;马腾;邱文凯;刘锐5.不同湖泊表层沉积物氮形态的分布特征与影响因素 [J], 赵宝刚;张夏彬;昝逢宇;胡晓康;潘婷;刘敏因版权原因,仅展示原文概要,查看原文内容请购买。
都支杜鹃地理分布及种群现状分析

S HAO Ja . e 。 WA G G n in w n N ag ,WA G G ox n ( .Sh o o ega h cec ,N nig N u —i g, a ④ 1 c ol fG orp y S i e aj n n
,
N r a U i ri ,N nig2 4 om l nv sy aj 0 6,C ia .C l g fLf Sine ,A qn oma U iesy e t n 1 0 hn ;2 o eeo i c cs n ig N r l nvr t, l e e i A qn 4 0 , hn ; .C l g f i c ne , n u N r a U iesy Wu u 4 0 0, hn ) n i 2 6 1 C ia 3 o eeo f S i cs A h i om l nvr t, h 10 C ia , g 1 l Le e i 2 - ln e u.& E v o .2 1 , 1 3 : 3 9 , at s r .P R o ni n 0 2 2 ( ) 9 - 7 r
Absr c :On t e b sso o s li g d c me t n pe i n ta t hห้องสมุดไป่ตู้ a i fc n u tn o u n sa d s c me s,g o r p i a srbu in a d p p lto e g a h c lditi t n o u ai n o sau fRh d d nd o h n iFa g i o o e d o tt s o o o e r n s a i n n Rh d d n r n L.we e c mp e e sv l n e tg t d a d a ay e r o r h n ie yi v si ae n n l z d. a d as t e d n e e e e s v l a e n lo is n a g r d lv l wa e au td. Th e u t s o t a e r s ls h w h tR. s a i a rwl iti u e h ni s a n ro y d srb t d s e i si h b e Mo n an.is d srb to a g s o l i t d t o g a a o z ne wi n p c e n t e Da i u t i t it u in r n e i ny l e o a l n nd n r w o t a i mi h
好氧反硝化菌的研究进展

摘 要 综述了好氧反硝化菌的种类和特性 、好氧反硝化菌的反硝化作用机制和影响因素. 好氧反硝化菌主要包括假单胞菌属 ( Pseudom onas) 、产碱杆菌属 (A lca ligenes) 、副球菌属 ( Pa ra2 coccus)和芽孢杆菌属 (B acillus)等 ,属好氧或兼性好氧异养微生物. 好氧反硝化菌能在好氧条 件下进行反硝化 ,其主要产物是 N2 O ,并可将铵态氮直接转化成气态产物. 催化好氧反硝化菌 反硝化作用的硝酸盐还原酶是周质酶而不是膜结合酶. 溶解氧和 C /N 往往是影响好氧反硝化 菌反硝化作用的主要因素. 介绍了间歇曝气法 、选择性培养基法等好氧反硝化菌的主要分离 筛选方法. 概述了好氧反硝化菌在水产养殖 、废水生物处理 、降解有机污染物以及对土壤氮素 损失的影响方面的研究进展.
厌氧过程 [ 25 ] ,在反硝化作用过程中 , O2 被认为可抑 制反硝化还原酶 [ 10 ]. 另外 ,在有机物质氧化的过程 中 , O2 被普遍认为是首选的电子受体 [ 11 ] ,在有氧条 件下反硝化菌会优先使用溶解氧呼吸 ,这样就阻止 了使用 NO3 - 和 NO2 - 作为最终电子受体.
3 国家自然科学基金项目 (40471065) 、土壤与农业可持续发展国家 重点实验室基金项目 ( 055122 ) 、国家计委 、教育部科技创新工程重 大项目培育基金项目 ( 705824 ) 和江苏省重点科技专项资助 项目 (BM2002701) . 3 3 通讯作者. E2mail: zhongwenhui@ njnu. edu. cn 2006210225收稿 , 2007208209接受.
11期 王 薇等 :好氧反硝化菌的研究进展
2619
了在脱氮副球菌生长过程中 ,如果 O2 和 NO3 - 共同 存在 ,其生长速率会比二者单独存在时高. Bell等 [ 3 ] 证明 ,在 O2 存在的条件下好氧反硝化酶仍具有活 性. M eiberg等 [ 10 ] 报道 Hyphom icrobium X 能在好氧 条件下进行反硝化作用. 目前许多研究证明了好氧 反硝化菌的存在 [ 7, 17, 53 ] ,并发现了一些能在 O2 浓度 (氧分压 )很高情况下生存的反硝化菌 [ 51 ]. 好氧反硝 化菌的发现为生物脱氮等技术提供了崭新的思路. 本文从种类 、机制 、分离筛选 、应用等方面介绍国内
江苏省环境影响评价技术评审专家

21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
赵志强 翟由涛 蔡邦成 水建高 张磊 沈众 冯彬 凌虹 黄夏银 吴俊锋 孙卫红 陈婷 王向华 张彤晴 聂峰
环境保护部南京环境科学研究所 南京国环环境科技发展股份有限公司 南京国环环境科技发展股份有限公司 南京国环环境科技发展股份有限公司 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省环境科学研究院 江苏省淡水水产研究所 中国电力工程顾问集团华东电工 高级规划师 高工 高工 高工 研高 高工 高工 高工 高工 高工 高工 高工
览表(第二批)
职称获得年份 2000 2005 2004 1998 2003 2006 2010 2011 2010 2012 2007 2013 2007 2005 1998 2005 2010 2007 2005 2011 江苏省环境影响评价技术评审专家一览表
江苏省环境影响评价技术评审专家一览表(第二批)
序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 姓名 周钰明 傅大放 何建平 梅天庆 李澄 周迟骏 孙华 王强盛 王玉萍 方斌 李利 李时银 王国祥 鲁长虎 谢吉民 奚新国 马如宏 周海 孙勤芳 邹长新 工作单位 东南大学 东南大学 南京航空航天大学 南京航空航天大学 南京航空航天大学 南京工业大学 南京农业大学 南京农业大学 南京师范大学 南京师范大学 南京师范大学 南京师范大学 南京师范大学 南京林业大学 江苏大学 盐城工学院 盐城工学院 盐城工学院 环境保护部南京环境科学研究所 环境保护部南京环境科学研究所 职称 教授 教授 教授 副教授 教授 教授 教授 副教授 教授 教授 教授 教授 教授 教授 教授 副教授 教授 教授 研究员 副研究员
高职高专:【江苏城市职业学院】-环境监测与治理技术专业建设方案

项目组成员项目组组长:胡大华城市科学系系主任/副教授/博士项目组副组长:王国祥南京师范大学全国高职高专环保与气象类专业教指委秘书长/教授张祥志江苏省环境监测中心教授级高工王志良江苏省环境科学研究院江苏省环境工程重点实验室主任/研究员周剑峰江苏优联检测技术XXX 总经理韩承辉城市科学系系副主任/副教授/博士项目组校内成员:张晓教务处、校企合作处副处长/副教授/博士冯国刚城市科学系副处长/副教授/硕士刘海霞城市科学系讲师/硕士周丹城市科学系高级工程师潘涛城市科学系副教授/硕士戴朝霞城市科学系讲师/硕士刘霞城市科学系讲师/博士干方群城市科学系讲师/博士秦品珠城市科学系讲师/博士唐荣城市科学系讲师/博士陈海蓉城市科学系助教/硕士程婷城市科学系助教/硕士黄兆琴城市科学系助教/硕士项目组校外成员:王小平江苏省环境工程重点实验室高级工程师吕路南大金山环保科技XXX 总经理/高工龙超南大金山环保科技XXX 教授朱晓燕南京市鼓楼区环境监测站站长/高级工程师顾俊南京恒泰检测技术XXX 总经理赵恒星江苏优联检测技术服务XXX南京分公司经理丁曦宁江苏省环境监测中心工程师章勇江苏省环境监测中心工程师目录一、概述 (1)二、建设目标 (2)(一)总目标 (2)(二)具体建设目标 (4)三、建设背景和建设基础 (6)(一)建设背景 (6)(二)建设基础 (8)四、建设内容与改革举措 (14)(一)创新校企合作机制 (14)(二)夯实“校中厂”、“厂中校”为重点的实训基地建设 (14)(三)深化“三结合、三进阶”的人才培养模式 (15)(四)课程体系和课程建设 (17)(五)教师队伍建设 (20)(六)社会服务与辐射能力 (20)(七)建立第三方评价 (20)五、专业群建设 (21)(一)专业群现状 (21)(二)建设目标 (21)六、建设进度与项目预算 (22)七、预期效果 (30)一、概述江苏城市职业学院环境监测与治理技术专业是江苏省特色专业建设点,我院重点建设专业。
微生物在污染环境生物修复中的应用

生物修复(Bioremediation)又称为生物恢复(Biorestoration),是指利用生物特别是微生物的代谢潜能消 除或减少污染地区有害物质浓度的技术[5]。自1989年以来生物修复技术已被用于清除受污染农田、地下 水、河流、湖泊和海洋等环境中的污染物,这些污染物包括石化产品、多环芳烃、卤代烷烃、卤代芳烃[6 J以及 引起水体富营养化的其他成分等。虽然微生物不能降解重金属,但可转移重金属或降低其毒性。实践表 明,生物修复是治理大面积污染的一种实用性强的技术。生物修复通常分为原位修复和异位修复。原位修 复是指通过在污染地点投加刺激物或人工培养的微生物加速污染物质的分解,使污染物在当地降解。异位 修复则是指将被污染的土壤或水移至专门的反应器中进行修复。生物修复常以原位修复为主,因不需另建 处理设施,且修复的场地可以照常用于生产,并可与其他技术结合使用,降解过程迅速,费用低∞一】。 1 影响污染环境微生物修复的几个关键因素
苦草光合作用日变化对水体环境因子及磷质量浓度的影响

沉水植 物具有显 著提高水 质的作 用 ,其 机制 主 要包括影 响水质理化 参数 、营养盐 的 吸收 、改变沉 积物理化环 境等【 j目前研 究沉水植 物对水 质的影 】。 之 响大 多集 中在 长时 间( 月或一 年) 数 尺度上 进行 ,在 长时 间尺度 上沉 水植 物通 过改变 环境 因子 (H、溶 p 解 氧和 E h等) 式来影 响沉积物 和水体之 间磷 的 的方 交换 ,从而 改变水体 中营养盐 的水平 。但是 在一天 当 中沉水植 物对水质 的影 响研 究较少 ,从理 论上推 测 ,由于水生植 物在昼夜 和不 同天气条件下 具有不 同 的生理 过程 ( 白天 由于沉 水植 物 的光合 作 用提 如 高水 中的溶 解氧 ,而夜 晚沉 水植物 的呼 吸作 用耗氧 可能 降低 水 中的溶 解氧 ) 可能会 导 致水 体 中磷 质量
水体各形态磷 的质量浓度变化呈 白天降低 、 夜间升高趋势 , 水体中各形态磷 的质量浓度变化与苦草光合作用 日变化趋势相反。 实验结果表明 ,苦草光合作用引起的水体 中环境 因子的 日变化影响了沉积物磷释放 的动态平衡过程,从而引起水体磷质量浓
度 的 日变化 。 关键 词 :苦 草 ;环境 因子 ;磷 ; 日变 化 ;光 合 作 用 中 图分 类 号 :X13 7 文 献标 志 码 :A 文 章 编 号 : 17 —9 6( 0 0)1-6 90 6 450 2 1 2 6 —6 1
=
植 物 叶 片对 环 境 的光 合 响 应提 供 了植 物在 不 同光 条 件下 生 存 和 生长 的 能力 以及 对不 断变 化 的 环境条件适 应能力 的信息L 植物 叶绿素荧 光信号 3 。
能快 速 灵 敏地 反 映植 物 生 理状 态 及 其 与环 境 的关
1960年以来太湖水生植被演变

1960年以来太湖水生植被演变赵凯;周彦锋;蒋兆林;胡娟;张秀山;周洁;王国祥【摘要】太湖的富营养化污染日益严重,针对太湖水生植被的研究工作非常重要,然而全面的太湖水生植被调查已经有将近二十年未见报道.基于2014年夏季全湖水生植被调查结果,结合历史资料,比较分析1960年以来太湖水生植被演变情况.结果表明,1960年以来,共有23种水生植物从太湖消失,其中1981、1997和2014年分别消失7、4和12种.从分布区面积来看,1960年以来太湖水生植被总体呈北部湖区水生植被消失,东北部、东部及南部湖区水生植被分布区面积持续扩张的态势,1981年全湖水生植被分布区面积占8%,到2014年已经有33.82%的水面有水生植被分布.从生物量组成来看,太湖水生植被先升后降,从1960年的10×104 t,持续上升到1988年的44.72×104 t,1997年下降到36×104 t,2014年进一步下降到29.09×104 t.但挺水植被以外的水生植被,尤其是浮叶植被的生物量一直保持上升态势.总生物量的下降与东太湖挺水植被大面积消失有关,到2014年全湖挺水植被生物量比重仅占5.15%,东太湖沼泽化问题已不复存在.从群落组成变化情况来看,苦草(Vallisneria natans)群落分布区面积锐减,马来眼子菜(Potamogeton malaianus)和荇菜(Nym-phoides peltatum)分布区持续扩张.目前太湖水生植被管理面临的主要问题是北部湖区水生植被恢复和东部湖区水生植被过量生长.%Comprehensive investigations on aquatic vegetation have been scarcely implemented in Lake Taihu during the past two decades, which is of special importance with the severe eutrophication and pollution. The change in aquatic vegetation in Lake Taihu since 1960 were analyzed based on the exhaustive survey of vegetation in summer, 2014. The results indicated that 23 spe-cies disappeared in Lake Taihu since 1960, in which 7species disappeared in 1981, 4 species disappeared in 1997, and 12 spe-cies disappeared in 2014. From the perspective of spatial distribution, aquatic vegetation showed a trend of expansion in the east part and a trend of shrinking in the west of the lake. The coverage rate of aquatic vegetation to the entire lake area was 8% in 1981, whereas, this value increased to 33.82% in 2014. The biomass of aquatic vegetation increased from approximately 1×105 t in 1960 to 4.472×105 t in 1997, and the n decreased from 3.6×105 t in 1997 to 2.909×105 t in 2014, respectively. However, excluding the emergent Phragmities australis community, the biomass of other aquatic vegetation, particularly that of the floating-leafed vegeta-tion, continuously increased. A rapid decrease of biomass was closely related to the diminishing of emergent plants in the eastern lake, which accounted for only 5.15% of the total vegetative biomass in Lake Taihu in 2014. It suggests that now the trouble of swampiness does not exist in this area any longer. The change in community composition revealed that the area of Vallisneria natans sharply decreased, while that of Potamogeton malaianus and Nymphoides peltatum progressively increased. Currently, the main is-sues of aquatic vegetation management in Lake Taihu are restoring the aquatic vegetation in the northern lake and controlling the o-vergrowth of aquatic vegetation in the eastern lake.【期刊名称】《湖泊科学》【年(卷),期】2017(029)002【总页数】12页(P351-362)【关键词】太湖;水生植被;植被演变;植被管理;水草收割【作者】赵凯;周彦锋;蒋兆林;胡娟;张秀山;周洁;王国祥【作者单位】南京师范大学环境学院,南京 210023;安庆师范大学资源环境学院,安庆 246011;中国水产科学研究院淡水渔业研究中心,无锡 214081;江苏省太湖渔业管理委员会办公室,苏州 215104;安庆师范大学资源环境学院,安庆 246011;安庆师范大学资源环境学院,安庆 246011;安庆师范大学资源环境学院,安庆 246011;南京师范大学环境学院,南京 210023【正文语种】中文水生植被作为湖泊生态系统的主要初级生产者,在维持水生态健康过程中扮演着至关重要的角色[1-3]. 自从湖泊二相理论[4-6]被广泛应用到富营养化湖泊治理中以来,越来越多的生态学家相信维持水生植被的生长是健康水体生态修复的关键手段及目标所在[7-8].太湖作为一个得到广泛关注的富营养化水体,其水生植被的研究工作也得到了广泛的开展. 历史上针对太湖的全面植被调查大体分为三个阶段:第一阶段是建国后为了开发利用太湖水生生物资源进行的综合性调查[9];第二阶段是改革开放以后从渔业生产的合理开发角度进行的水生植物生产力调查*曹萃禾.东太湖水生维管束植物调查报告(铅印本),1981.*许兆明.西太湖水生维管束植物调查报告(铅印本),1981.[10-11];第三阶段是1990s以后随着富营养化污染的日益严重而进行的富营养化机理研究中涉及的植被调查[12]. 此后的太湖植被调查仅限局部湖区[13-17],针对全湖水生植被的全面系统调查已经有近二十年未见报道. 在太湖水污染现状日益严峻的今天,作为太湖水生态过程中的重要环节之一,针对太湖全湖的水生植被现场调查紧迫而且必要.本文基于2014年夏季太湖全湖水生植被调查结果,结合历史资料,比较分析1960年以来太湖水生植被演变情况,试图全面了解太湖水生植被的分布及演变,为太湖水资源管理提供理论依据.1.1 断面设置调查时间为2014年8月3-14日. 调查范围仅包括太湖岸线以内常年被水淹没的区域,不包括沿岸湖滨带的芦苇群落. 由于太湖面积太大,为便于对植被分布状况进行描述,将全湖划分成9个调查区域分别进行调查,每个调查区内根据预调查结果设置断面,要求断面尽量均匀分布,且能穿过典型水生植被分布区(图1). 调查区内,每条直线视为一个断面,共设置91条断面,总长度899.63 km(表1). 此外,考虑到太湖中大型岛屿周边可能存在水生植物分布,在西洞庭山、乌龟山、三山、漫山等岛屿周边另设样线绕岛航行.1.2 调查方法调查范围为湖区所有水生植物物种及群落组成,另有少量湿生植物也在湖区内有分布,一并纳入调查范围. 水生植物与湿生植物区别在于:水生植物是指能在淹水环境中完成整个生活史过程的植物;湿生植物是指能在过湿环境中完成整个生活史过程的植物,部分物种因其较强的耐涝性可在淹水环境正常生长,但不能完成整个生活史过程. 根据水生植物各层片的优势种组成对群落进行命名,同一层片的优势种之间用“+”相连,不同层片之间的优势种用“—”相连.利用船载法对水生植被进行调查,尽量沿断面行船,根据水深及风浪条件判断可能的水生植物分布区,在目测无水生植物分布的水域,每5 km设1个采样点,测量水深、透明度、浊度等环境因子,然后用自制铁钩拖行100~200 m判断水下有无植被分布. 发现有植物分布的水域直接用GPS打点标记,估算盖度,并在典型水生植物群落分布区通过样方法调查物种及群落组成. 挺水植被样方大小为1 m×1 m,直接用镰刀收割样方内所有物种的地上部分,记录物种组成,估算盖度,称取生物量;挺水植被以外的水生植被通过水草采样夹(0.15 m2)进行群落调查. 每个调查区内每种群落设置的样方数至少为5个.在ArcGIS软件中完成植被图的绘制并计算各群落分布区面积,具体方法为:将全湖标记的485个样点(图1)导入软件中,根据每个样点的水生植被分布情况进行群落分类,先用直线将相邻断面上相同的植物群落连接,再通过同时期遥感影像进行校正,若遥感影像无法分辨群落差别则通过水深、透明度、风浪等水文条件估测可能的水生植被分布区边界. 生物量集中在水底的沉水植物往往无法通过目测估计其盖度,采取的办法为默认其盖度为100%,根据多个样方生物量平均值乘以总分布区面积得出该群落的总生物量. 生物量集中在水面附近的浮叶植物和沉水植物,其水平结构上往往不是均匀分布的,采取的办法为在生物量最大处设置样方,再根据样方处的盖度和总植物群落盖度换算出单位面积生物量,乘以该群落的分布区面积得出总生物量.2.1 物种组成本次调查共记录到维管植物22科31属39种(表2),包括蕨类植物3科3属3种,双子叶植物11科12属14种,单子叶植物8科17属22种. 其中挺水植物11种,沉水植物13种,浮叶植物5种,漂浮植物7种,湿生植物3种.2.2 群落组成太湖水生植被总体呈北部湖区和西部湖区裸露,东北、东部、南部湖区广泛分布的格局,湖心仅西竹岛附近有零散的马来眼子菜群落分布,另北部湖区梅梁湖近湾口偶见单株马来眼子菜. 根据水生植物群落优势种组成,将太湖水生植被划分成10种群落类型(图2). 由西向东以马来眼子菜群落逐步过渡到荇菜-马来眼子菜群落,再过渡到其他群落类型,单位面积生物量逐渐增加,群落组成也逐渐复杂.以挺水植物为主要优势种的群落类型主要分布在东部湖区的东太湖和胥湖近岸区,其他湖区近岸区也有小面积分布,主要优势种有芦苇、菰和莲.浮叶植物为主要优势种的群落类型以荇菜-马来眼子菜群落最为多见,以南部湖区分布面积最大,东部湖区和东北部湖区也有大面积的片状分布. 荇菜+菱-聚草-金鱼藻群落分布于东部湖区的东太湖苏州湾以西水域、冲山以东水域及胥湖航道北侧水域,适宜淤泥底质的静水水域.以沉水植物为主要优势种的群落类型包括马来眼子菜群落、苦草群落、微齿眼子菜群落、马来眼子菜-微齿眼子菜群落、微齿眼子菜+黑藻群落、马来眼子菜+聚草-金鱼藻群落. 其中,马来眼子菜群落分布区面积最大,从东北部湖区一直到南部湖区均有大面积连片分布,但该群落或呈零星分布,或呈斑块状分布,总体盖度一般小于1%. 苦草群落分布区位于东西山之间水域. 微齿眼子菜群落分布区位于贡湖东部的羊湾水厂取水口附近水域. 微齿眼子菜+黑藻群落及马来眼子菜-微齿眼子菜群落分布于胥湖近岸水域,生物量较低. 马来眼子菜+聚草-金鱼藻群落大面积分布于东太湖及东西山之间水域.2.3 生物量分布2014年夏季太湖湖区水生植被分布区面积共计790.76 km2,占太湖总水域面积的33.82%,夏季总生物量分别为29.09×104 t(鲜重)、2.96×104 t(干重). 各植物群落分布区面积及生物量分布情况见表3.从生态型来看,挺水植物群落分布区面积9.87 km2,占太湖水域总面积的0.42%;浮叶植物群落分布区面积168.84 km2,占太湖水域总面积的7.22%;沉水植物群落分布区面积为612.05 km2,占太湖水域总面积的26.18%.从群落类型分布情况来看,马来眼子菜群落分布区面积最大,占总植物分布区面积的62.64%,但大多分布于与敞水区交界的区域,多成零散分布,总盖度仅为0.3%,因此其生物量总量不大,仅占总生物量的6.21%. 荇菜-马来眼子菜群落是太湖现存植被中生长最为旺盛的群落类型,二者常以面积不等的斑块在水平结构上镶嵌排列,分布区面积居第二位,占总植物分布区面积的18.37%,生物量却达到总生物量的43.84%,居第一位. 马来眼子菜+聚草-金鱼藻群落分布区面积居第三位,占总分布区面积的2.91%,占总生物量的6.48%,也居第三位. 荇菜+菱-聚草-金鱼藻群落见于东部湖区的东太湖、胥湖及冲山附近水域,分布区面积居第四位,但植被生长非常茂盛,占总植被分布区面积虽然只有2.98%,生物量占比却能达到17.97%,居第二位.3.1 物种组成变化1960年的调查共记录66种维管植物[9],1981年为61种,1997年为66种[12],对照物种名录将这些物种进行生态型划分:1960年调查到49种水生植物、17种湿生植物,1981年有45种水生植物、16种湿生植物,1997年水生和湿生植物分别有43和23种. 本次调查(2014年)共记录水生植物36种,湿生植物3种.对比1960年以来太湖水生植物物种组成变化情况,根据《中国植物志》的分类体系将不同时期调查到的太湖高等植物物种名录进行重新划分,将佛朗眼子菜(Potamogeton franchetii)并入眼子菜(Potamogeton distinctus)[18],其他种名和学名也进行相应调整. 在此基础上对1960年以来太湖水生植物物种分布情况进行比较分析(表4). 结果表明,共有26种水生植物分布情况发生变化,其中挺水植物10种,浮叶植物4种,漂浮植物2种,沉水植物10种. 这26种植物中,有23种为逐渐消失的物种,其中有7种在1981年调查时消失,4种在1997年调查时消失,12种在2014年调查时消失. 有3种植物为外来迁入物种,其中伊乐藻为1986年人工引入[19],水盾草为外来物种入侵[17],篦齿眼子菜迁入原因不明,鉴于该物种较强的耐污性,可能与水质恶化有关.3.2 生物量变化从总生物量变化情况来看,1960年以来,太湖水生植被生物量保持先升后降的态势,从1960年的10×104 t上升到1981年的36.82×104 t,增加3倍多,到1987年进一步上升到44.46×104 t,1988年维持在44.72×104 t,1997年下降到36×104 t,2014年进一步下降到29.09×104 t. 但从生物量组成来看,挺水植被变化趋势与总生物量一样,先升后降,挺水植被以外的沉水植被和浮叶植被则一直保持上升的态势(图3).1960年的调查中,挺水植被共8×104 t,占总水生植被生物量的80%;1981年挺水植被生物量达30.17×104 t,占比达81.94%;1987和1988年分别为34.8×104和33.02×104 t,分别占78.27%和73.83%;1997年,挺水植被生物量减少到22.5×104 t,占比62.5%;到2014年,挺水植被生物量下降到2.91×104 t,仅占湖区水生植被生物量的10.00%. 挺水植被以外的水生植被在1960年仅2×104 t,占总生物量的20%;到1981年上升至6.65×104 t,占比18.06%;1987和1988年达到9.66×104和11.70×104 t,分别占总生物量的21.73%和26.16%;1997年进一步上升至13.5×104 t,占37.50%;2014年更是达到26.18×104 t,占总生物量的90.00%.从水生植物分布区范围来看,1960年仅在东部湖区的东太湖和东西山之间有大面积水生植被分布[9]. 到1988年,东部湖区的胥湖、光福湾、镇湖湾,东北部湖区的贡湖,北部湖区的梅梁湖、竺山湖等水域均出现不同面积的水生植被,水生植被分布区面积占总水域面积的8%[11]. 1997年北部湖区的水生植被明显退化,仅竺山湖有少量残存,东北部湖区及东部湖区水生植被反而持续扩张,盖度>1%的植被分布区面积占全湖水域面积的19%[12]. 2014年夏季,北部湖区植被完全消失,东部湖区水生植被则进一步扩张,胥湖和东西山之间几乎全部有水生植被分布,南太湖更是扩张明显,几乎全部水域都有水生植被分布,甚至蔓延到西部湖区的东侧,全湖水生植被分布区面积达到33.82%.3.3 群落分布变化结合历史资料绘制1960年以来历年太湖水生植被空间分布图(图4). 从植被分布区来看,沉水植被和浮叶植被的增加趋势并没有生物量这么明显,这是由盖度差异导致的. 例如,1960年东太湖以外的湖区水生植被分布区范围并不小,但几乎全部为单株或零星分布,生物量不足15 g/m2[9].从东太湖水生植被群落组成来看,1960年东太湖优势种为挺水植被菰和芦苇,沉水植被优势种为马来眼子菜和苦草. 1981年挺水植被中菰的优势度上升,芦苇仅占少量,沉水植被中马来眼子菜优势度下降显著,但苦草分布区面积显著上升. 1987年和1988年菰分布区面积和生物量均远大于芦苇,沉水植被中苦草和马来眼子菜为共优种. 1997年东太湖芦苇和菰的分布区面积均有所上升,沉水植被优势种变为微齿眼子菜,浮叶植被荇菜和菱也开始成为建群种,但分布区范围非常有限. 从2002年开始,东太湖菰和芦苇分布区面积锐减,浮叶植被和沉水植被分布区面积及生物量显著上升,荇菜、伊乐藻、马来眼子菜等成为优势种[20]. 2014年东太湖挺水植被分布区面积进一步下降,仅沿岸有少量芦苇群落分布,菰和莲仅在围网外围有人工栽培,其他区域以马来眼子菜和荇菜分布最为广泛.东太湖以外水域在1960年仅在近岸区的芦苇群落外围有少量马来眼子菜和苦草分布. 1981年,北部湖区的竺山湖和东部湖区的杨湾附近水域均出现大量苦草群落,马来眼子菜群落分布区面积则锐减,生物量仅为1960年的11%,同时,在围网外围开始出现少量荇菜群落. 1988年苦草、聚草和马来眼子菜成为优势种,其中苦草优势度最大,集中分布在北部湖区的竺山湖和东部湖区的杨湾附近水域,聚草则分布于东部湖区的胥湖,马来眼子菜在各湖区零散分布. 1997年北部湖区水生植被基本消失,东北部湖区、东部湖区及南部湖区以马来眼子菜和微齿眼子菜为主要优势种. 2014年北部湖区无水生植被分布,在东部、东北部及南部湖区马来眼子菜群落呈现出由东向西扩张的态势,成为分布区最大的水生植物,而荇菜更是一跃成为分布区第二广泛的水生植物,南部湖区、东部湖区、东北部湖区,均可见一望无垠的荇菜群落,场面蔚为壮观. 微齿眼子菜仍然是主要优势种之一,但分布区转移到了东北部湖区的贡湖和东部湖区的胥湖. 此外,菱、聚草、金鱼藻和黑藻均在东部湖区有成片分布.自1960年以来,一共有23种水生植物从太湖消失. 从消失的物种组成来看,不仅包括了水蕨、莼菜这样的珍稀濒危物种,也包括了矮慈菇、野慈菇、眼子菜、石龙尾、茶菱这样的广布种. 其余物种如萍蓬草、水车前、乌苏里狐尾藻、狐尾藻、水马齿等曾经都是分布非常广泛的物种,如今在整个长江下游地区都已变得非常罕见,由此推断,太湖水生植物物种多样性的丧失具有一定的普遍性. 此外,莼菜和水葱在太湖周边有大量人工栽培,茶菱、矮慈菇、野慈菇、鸭舌草在周边的农田水网仍有大量分布. 这说明部分消失的水生植物可以通过人工引种和生境修复恢复生长. 在所有消失的物种中,有7种在1960-1981年间消失,4种在1981-1997年间消失,12种在1997-2014年间消失. 鉴于不同时期太湖环境变迁的主要驱动力不同,1960-1981年间消失的物种可能是大规模围湖造田和建闸后水位变化趋于稳定造成的,而1981年以后消失的16个物种则是水质持续恶化和围网养殖、围堰取土、人工水位调控等人为干扰共同导致的.水生生态系统包括水质浑浊的藻型和水质清洁的草型两种稳态,当水体营养盐上升到一定的阈值后会发生草型湖泊向藻型湖泊的转换,从而造成水环境恶化、生态功能丧失[5]. 大量文献报导了富营养化导致太湖水生植被消失[14,16,22],而太湖的富营养化污染更是已经持续二十余年并保持加剧态势[23-24]. 本研究结果表明,自1960年以来,伴随着愈演愈烈的水体富营养化过程,太湖的水生植被不仅没有消失,相反其分布区面积和生物量却持续上升. 但是,从植被分布区变化格局来看,水生植被分布区上升仅发生在水质相对较好的东部湖区,北部湖区水生植被在1997年以后已经完全消失. 湖泊二相理论从1993年提出以后受到了多方面的挑战,该理论的提出者Scheffer本人于2007年对其进行了完善,提出生态系统的二相转换是受多方面因素调控的,除了营养盐水平之外,还应包括空间异质性、水位变化、湖泊面积、气候变化等因素[6]. 对于太湖这样的超大型湖泊而言,其复杂的空间异质性是毋庸置疑的. 同时,太湖的开阔湖面非常广阔,风浪也成为影响植被分布的重要影响因子之一[12]. 此外,太湖地处经济活跃的苏南平原,人为活动密集,除营养盐输入之外,增殖放流、渔业生产、蓄水调水、水草收割等都对太湖水生植被分布格局造成影响. 因此,对于太湖这样的超大型湖泊而言,其水生植被的分布格局应该是非常复杂的,准确描述其水生植被演变的机制还有待进一步研究.在2002年之前的研究工作中,东太湖沼泽化一直是被广泛关注的热点问题[15-16,25]. 历史上东太湖一直是一个受人为干扰较小、保持自然演替格局的荒芜水域,挺水植被芦苇分布广泛. 1960年以后开始的大规模围湖造田使得水域面积下降,芦苇群落大面积消失. 围湖造田造成的水流减缓和随后人工引种栽培的菰群落大量繁衍,加速了东太湖的沼泽化进程,到1980s,东太湖挺水植被生物量已超过20×104 t. 2000年以后,随着大规模围网养殖,东太湖水生植被遭到严重破坏,尽管沼泽化趋势仍然持续,但生物量已经显著下降[9]. 2009年以后,规范化水产养殖使得东太湖苏州湾以西水域围网面积显著下降,挺水植被在此区域几乎完全消失,浮叶和沉水植被分布区得到部分恢复. 2012年开始,地方政府陆续在东太湖苏州湾及以东水域挖深取土以供城市建设,使得该水域水深普遍达到4 m以上,局部区域甚至达到9 m. 这一系列人为干扰对东太湖水生植被群落组成造成了显著影响,广袤的挺水植被已不复存在,苏州湾及以东水域几无水生植被分布,以西部分几乎全被浮叶和沉水植被占据. 到2014年夏季,东太湖挺水植被仅9.18 km2,占东太湖总水域面积的7.01%,东太湖沼泽化问题已不复存在.到调查时为止,太湖已经成为一个典型的“草藻混合型湖泊”,即东部、东北部和南部湖区以草型湖泊为主,有大面积水生植被分布,其他湖区则为藻型湖泊,水草消失,蓝藻水华恶性暴发. 综合分析来看,太湖的水生植被演变的主要驱动力包括围湖造田、渔业生产、人工水草收割、富营养化污染和人工水位调控等. 围湖造田的影响主要是在1960s-1990s期间,以东太湖为例,1960-1997年间,围湖造田导致东太湖面积减少120 km2,导致沿岸芦苇群落大面积衰退,湖区的浮叶植物和沉水植物开始新的演替过程[12]. 渔业生产对水生植被的影响体现在1980s后太湖范围内广泛的围网养殖、草食性鱼类的大量投放及破坏底泥的捕捞作业等. 围网养殖导致湖区风浪减小,有利于水生植物生长的同时也加速了湖泊沼泽化进程[12],竺山湖1995-1998年的草食性鱼类养殖可能是导致该水域水草消失的主要原因[12],本次调查发现的光福湾水生植被消失可能与该水域大量投放中华绒螯蟹(Eriocheir sinensis)有关. 人工水草收割对太湖水生植被的影响主要体现在1960s-1980s,此时周边百姓大量捞取太湖水草用作绿肥,芦苇也被大量收割,这直接导致了太湖水生植被生物量的严重下降,1983年以后太湖严禁水草打捞,水生植被开始有所恢复[10]. 此外,受目前东部湖区水生植被大量生长的影响,水生植被集中腐烂,导致水质快速恶化,影响水资源利用的现象时有发生[17]. 2012年开始,太湖苏州部分重新开启了大规模水草打捞,捞草船和人工打捞同步进行,这对太湖水生植被的影响评估尚未见报道. 水位通过影响水生植被的光合作用对其分布格局存在显著影响[26-27],2014年8月对122个样点进行的水深测定显示,太湖平均水深为243.12 cm,对水生植被分布区内74个样点的水深测定结果显示,水生植被分布区平均水深209.22 cm,这个数值已经逼近水生植被分布范围的临界值[28],这可能会使得太湖的水生植被对水位变化变得非常敏感,针对该假设的研究还有待进一步开展.最近十几年来,遥感技术取得了长足进步,并被应用到太湖植被调查中[29-30],对于长期监控植被演变具有重要意义. 该技术不需要繁重的野外实地调查,仅需进行简单的现场校正即可,可弥补野外实地调查中断面之间水域植被分布情况完全未知的缺陷. 但该技术受天气、光照、风浪、季节等的影响较大,受光谱分辨率的影。
沉水植物苦草对上覆水各形态磷浓度的影响

沉水植物苦草对上覆水各形态磷浓度的影响陈秋敏;王国祥;葛绪广;王立志【摘要】沉水植物是影响湖泊磷营养状态的重要因素.在室内模拟了"水苦草沉积物"生态系统,在沉水植物苦草生长过程中分析上覆水中的总磷(TP)、溶解性总磷(TDP)、溶解性有机磷(DOP)、溶解性活性磷(SRP)、颗粒态总磷(PP)的变化,探讨了苦草对上覆水磷赋存形态的影响.结果表明,苦草在生长过程中减缓了沉积物磷的相对释放速率使上覆水中TP、SRP、DOP、PP均呈现不同程度的下降.上覆水TP、SRP、DOP、PP分别下降了37.5%、74.6%、42.0%、19.5%;苦草主要通过吸收上覆水和间隙水中的磷及对环境因子的影响,使得上覆水中各形态磷浓度保持在较低的水平,降低沉积物向水体释放磷的速率.【期刊名称】《水资源保护》【年(卷),期】2010(026)004【总页数】5页(P49-52,56)【关键词】苦草;上覆水;磷;沉积物【作者】陈秋敏;王国祥;葛绪广;王立志【作者单位】南京师范大学地理科学学院,江苏,南京,210046;江苏省环境演变与生态建设重点实验室,江苏,南京,210046;南京师范大学地理科学学院,江苏,南京,210046;江苏省环境演变与生态建设重点实验室,江苏,南京,210046;南京师范大学地理科学学院,江苏,南京,210046;江苏省环境演变与生态建设重点实验室,江苏,南京,210046;南京师范大学地理科学学院,江苏,南京,210046;江苏省环境演变与生态建设重点实验室,江苏,南京,210046【正文语种】中文【中图分类】X173磷是影响湖泊富营养化的关键性限制因素之一[1-2],同时也是植物生长发育的必需元素。
水体中各种磷形态之间的转化,尤其是其他形态磷化合物向正磷酸盐转化速率是决定湖泊营养状态和上覆水生产力的重要因素[3-4]。
沉水植物是湖泊生态系统的一个重要组成部分和主要的初级生产者之一,对湖泊生态系统中物质和能量的循环起着重要作用,沉水植物占据了湖泊中水和沉积物的主要界面,是水体两大营养库间的有机结合,对湖泊生产力及湖泊生态系统过程具有重要的影响[5],沉水植物在生长发育、衰亡及季相交替过程中,通过光合、呼吸、残体分解等作用,可以对上覆水的DO、pH、氧化还原电位(Eh)以及水动力等产生复杂的影响,并通过这些影响因子的变化进而影响到磷素的迁移转化[6-9]。
芦苇和蓝藻混合分解对水质的影响

芦苇和蓝藻混合分解对水质的影响曹勋;王国祥;丁新春【摘要】开展室内模拟实验,探讨芦苇和蓝藻水华混合分解对水质的影响.结果表明:蓝藻藻浆和芦苇混合分解过程中,水质的变化表现出明显的阶段性,0~8天,芦苇和蓝藻分解较快,迅速释放C、N、P元素,第8天水体的TDN、TDP、TOC均达到最大值,分别可达41.88、0.63、294.82 mg/L,8~64天TDN、TDP、TOC总体上呈现下降趋势;混合分解过程中,水体NH4+-N浓度在0~32天保持相对较高的水平,第4天L、M、H组最高浓度可迭15.6、36.72、41.30 mg/L,32 ~64天呈现下降趋势;无机氮中NH4+-N占有绝对优势,占比大于90%;整个分解过程中,NO3--N 和NO2--N都保持较低的浓度.【期刊名称】《四川环境》【年(卷),期】2019(038)001【总页数】7页(P6-12)【关键词】芦苇;蓝藻;分解;水质【作者】曹勋;王国祥;丁新春【作者单位】南京大学盐城环保技术与工程研究院,江苏盐城224000;南京师范大学环境学院,南京210046;南京大学盐城环保技术与工程研究院,江苏盐城224000【正文语种】中文【中图分类】X7031 前言蓝藻水华的分解易引起水体局部厌氧,在特地的水文和气象条件下,蓝藻会和底泥中的有机质发生反应,形成“湖泛”[1],恶化生态环境的同时,威胁市民的饮水安全。
而太湖近岸区芦苇丛是蓝藻最易聚集的区域,大量芦苇残体和藻华在近岸区堆积衰亡,芦苇和蓝藻释放到水体中的氮磷营养物质可以为湖滨带大型水生植物提供必要的营养盐来源,对整个生态系统的物质循环具有积极的作用,但是同时,蓝藻分解释放的大量氨氮、磷酸根、有机氮、有机磷可以被微生物和浮游植物直接利用,成为维持水华的营养来源,为湖泛提供条件。
另外,过量的芦苇和蓝藻分解会引起二次污染,加快湖泊的富营养化进程。
因此,芦苇和藻华的分解过程对太湖物质循环过程和环境质量产生影响[2-3],而两者的混合衰亡少见报道。
苦草(Vallisneria natans)对沉积物微生物群落结构的影响

苦草(Vallisneria natans)对沉积物微生物群落结构的影响∗张亚朋;章婷曦;王国祥【摘要】A sediment-water-Vallisneria natans system was built within a simulating lake system, where phospholipid fatty acids ( PLFAs) method was used to determine the changes in sediment microbial community structures in the submerged plant V. natans at different growth stages. The results showed that during the growth of V. natans, the organic matter content decreased first and then increased. The concentrations of total phosphorus, organic phosphorus and inorganic phosphorus decreased by 8. 97%, 7. 81% and 10. 28%, respectively, during the growth period. The microbial activity and total phosphorus in sediments showed ex-tremely significantly negative correlation. In the early exuberant period, microbial activity in the experimental group was greater than the control group, while the decline in the microbial activity of the control group was slightly higher than the experimental group. Microbial community structure in the sediments of different periods showed significant differences. Bacteria were the main part of the total sediment microbes, accounting for 76% to 84% of the total microbial. As to bacterial distribution, gram-positive bacilli were predominant. The percentage of gram-positive bacteria and fungi increased with the growth of the V. natans.%模拟湖泊系统构建了“沉积物水苦草( Vallisneria natans)”系统,应用磷脂脂肪酸( PLFAs)法测定在沉水植物苦草不同生长时期沉积物表层微生物群落结构的变化,探讨沉水植物对沉积物中的微生物群落结构的影响.结果表明,从苦草生长初期到旺盛期再到衰亡期,沉积物中有机质含量先下降后上升;总磷、有机磷、无机磷分别下降了8.97%、7.81%、10.28%;沉积物微生物的活性与总磷呈极显著负相关,在苦草生长初期和旺盛期,实验组的沉积物微生物活性大于对照组,而在衰亡期对照组的沉积物微生物活性略高于实验组;不同时期沉积物中微生物群落结构发生了明显变化,组成结构差异显著,微生物组成中细菌占主要成分(占微生物总量的76%~84%);细菌中革兰氏阳性菌占主要优势,且革兰氏阳性菌百分含量随苦草生长呈上升趋势,革兰氏阴性菌呈下降趋势;真菌的百分含量呈上升趋势.【期刊名称】《湖泊科学》【年(卷),期】2015(000)003【总页数】6页(P445-450)【关键词】沉积物;苦草;微生物群落结构;磷脂脂肪酸【作者】张亚朋;章婷曦;王国祥【作者单位】南京师范大学地理科学学院,南京210023;南京师范大学地理科学学院,南京210023; 江苏省碳氮循环过程与污染控制重点实验室,南京210023; 江苏省地理信息资源开发与利用协同创新中心,南京210023;南京师范大学地理科学学院,南京210023; 江苏省碳氮循环过程与污染控制重点实验室,南京210023; 江苏省地理信息资源开发与利用协同创新中心,南京210023【正文语种】中文水体富营养化是目前我国面临的重大水环境问题.沉积物是水生生态系统的重要组成部分,它既是营养物质的蓄积库又是水体中氮、磷等营养盐的重要来源,与水体富营养化密切相关[1],在沉积物中,微生物是极为敏感并易受环境影响的生物类群[2].它们不仅是系统中生物量的重要组成部分,而且也可以影响沉积物中营养物质的分布及转化,因此沉积物微生物群落结构是水体环境变化和演替的重要标志[3].沉水植物是水生生态系统中的重要组成部分,对维护健康的水生生态系统至关重要[4],它们不仅可以改善水质,同时也可以改变微生物的群落结构.苦草(Vallisneria natans)为多年生沉水植物,是我国常见沉水植物之一,由于其生态适应性广,吸污能力和耐污性强,常被作为富营养化水体植被恢复工程的主要选用品种[5].沉水植物对微生物群落的研究多集中于比较不同沉水植物的作用.俞振飞等[6]比较了黑藻和苦草两种沉水植物对沉积物中磷的去除效率,结果表明苦草对沉积物各形态磷含量降低的影响大于黑藻;李琳琳等[7]研究表明苦草和菹草不仅可以改善水质,同时也可以显著提高水体中细菌的多样性,改变细菌的群落结构;Zhao等[8]比较了太湖3种沉水植物金鱼藻、菹草、苦草对沉积物中微生物群落的影响,结果表明不同沉水植物作用下,沉积物中微生物群落结构不同.本文以沉水植物苦草为研究对象,应用磷脂脂肪酸(PLFAs)分析方法研究了在苦草的不同生长时期,表层沉积物中的微生物群落结构变化特征,以深入了解沉水植物生长过程对沉积物中微生物的影响,以期为浅水湖泊沉积物内源释放的微生物作用机理提供依据.1.1 实验材料构建室内模拟生态系统:选取高密度聚乙烯桶(顶直径×底直径×高=55cm×45cm×75cm),内放置聚乙烯小桶(顶直径×底直径×高=10cm×10cm×20cm).苦草取自南京采月湖,在温室内驯化15 d.沉积物分别过100目筛去除其中的植物残体、石块等杂质后,平铺于聚乙烯小桶.然后选取生长状况基本一致、长势良好驯化后的苦草进行移栽,每小桶8株苦草.实验组设置6个平行样,没有种植苦草的对照组设置3个平行样.实验在江苏省环境演变与生态建设重点实验室的水环境生态修复中试平台玻璃房内进行,实验时间为2013年6-12月,分别在生长初期(第20 d)、生长旺盛期(第80 d)、衰亡期(第160 d)采集样品,每次采集6个实验组平行样和3个对照组平行样,6个实验组平行样中3个用来分析苦草根系生长状况,另外3个用来研究苦草对沉积物微生物群落结构的影响.由于前3cm的表层沉积物中苦草根系的生物量分配质量分数达到50%,因此选取前3cm作为研究对象.将采集的新鲜样品分成2份.一份立即处理测定微生物活性,测定前除去沉积物样品中可见植物残体.另一份经冷冻干燥后在-70℃保存,一周内完成PLFAs及磷形态测定.1.2 分析项目与测定方法含水率通过测定105℃烘干至恒重时的失重获得(GB 7172-1987);烧失率通过950℃灼烧1 h 达恒定质量后,测定失重获得(GB 7876-1987);有机质含量通过烧失率获得[9-10].沉积物中总磷的分析采用Ruban等[11-12]在欧洲标准测试委员会框架下发展的SMT分离方法.底泥中微生物活性采用FDA(荧光素双醋酸酯)法[13],称取2 g 左右湿沉积物于50 ml的反应管中,加入15 ml 60 mmol 磷酸盐缓冲液(pH=7.6)后再加入0.2 ml FDA溶液,启动反应.不添加FDA底物作为空白对比.混合均匀后在30℃下100转/min摇床培养20 min.培养结束后,立即加入体积比为2 ∶1的氯仿/甲醇15 ml,以终止反应并摇匀.2000转/min离心3 min,提取上清液并过滤,490nm下测吸光度.转换成单位质量干泥的活性强度.PLFAs由Agilent GC7890A GC 相色谱仪及MIDI Sherlock®全自动脂肪酸微生物鉴定系统检测:色谱柱:P/N 19091J-413;HP-5毛细柱:30 m,320μm×0.25μm;GC条件:进样口温度250℃,柱温300℃,载气为高纯氮气(20 ml/min),不分流进样,进样量1μl.PLFAs标样选用Supelco公司的Bacterial Acid Methyl Esters Mix和478852-U内含37种FAMEs(fatty acid methyl esters)脂肪酸甲酯混合标样,可涵盖沉积物中常见微生物的特征PLFAs.脂肪酸的命名:以“X ∶YωZ(c/t)”表示,其中“X”代表脂肪酸分子的碳原子总数,“Y”代表不饱和烯键的数目,“Z”为烯键或环丙烷链的位置,“ω”代表烯键距离羧基的位置.前缀i(iso)表示顺式支链,a(antieso)表示反式支链,“cy”代表环丙基支链,后缀“c”和“t”分别代表顺式和反式双键.特征脂肪酸是不同微生物类群的生物标志物,根据现有的研究成果总结,细菌源脂肪酸为14 ∶0、15 ∶0、16 ∶0、17 ∶0、i15 ∶0、a15 ∶0、il6 ∶0、16 ∶1ω9、il7 ∶0、cyl7 ∶0;真菌源脂肪酸以18 ∶1ω9c+t、18 ∶2ω6c+t、18 ∶3ω3表示;革兰氏阴性菌(G-)以cy17 ∶0表示;革兰氏阳性菌(G+)以il5 ∶0、al5 ∶0、il6 ∶0和il7 ∶0表示[14].数据用统计软件SPSS 17.0进行单因素方差分析(One way ANOVA),以比较实验组与对照组间差异的显著性;用PRIMER 5对PLFA数据进行MDS和SIMPER分析.2.1 苦草的不同生长时期对沉积物有机质及磷的影响从苦草生长初期到旺盛期,沉积物的含水率发生显著变化(P<0.05),在生长旺盛期达到57.93%.含水率的大小直接影响到磷在沉积物与上覆水之间的重新分配[15].苦草对沉积物中的磷有一定的吸收能力,从生长初期到旺盛期,沉积物中的总磷、有机磷、无机磷的含量都显著下降(P<0.05).有机质含量呈先下降再上升的趋势(表1).生长初期到旺盛期有机质含量下降,可能是由于有机质腐解过程中,微生物优先利用易降解的有机物和简单的有机物(可溶性糖、淀粉、有机酸等)进行生命活动,有机碳的分解速率加快;而在后期随着沉水植物的衰亡有机质含量快速增加,是苦草在衰亡期有一定的残体沉积引起的.植物残体等的沉积是沉积物有机质的主要来源,占有机质来源的90%以上[16].沉积物中的有机质与沉积物-水界面磷的生物地球化学循环密切相关[17],有机质的矿化过程能导致沉积物中氧化还原电位和pH值的改变, 从而影响沉积物中磷化合物的吸附解析和相互转化过程[18].从苦草生长前期到旺盛期总磷含量快速下降,可能是因为沉水植物在生长过程中可以通过茎、叶和根系吸收上覆水和沉积物中的磷,以合成体内的蛋白质、核酸和酶等物质来满足自身生长的需求[19].苦草是一种根系发达的沉水植物,对营养盐的吸收能力较强,因此,在前期沉积物中总磷的含量快速下降.在后期随着苦草的生长速度缓慢直至衰亡,对沉积物中营养盐的吸收量减少,因此后期总磷含量呈缓慢下降趋势.2.2 苦草的生长对沉积物微生物活性的影响从图1可以看出,从苦草生长初期到旺盛期,沉积物微生物活性显著上升(P<0.05),实验组微生物活性由生长初期的238.73 μg/(g·h)上升到旺盛期的299.42 μg/(g·h).在生长初期和旺盛期,实验组的沉积物微生物活性大于对照组,而在衰亡期空白组的沉积物微生物活性略高于实验组.这可能是由于植物根系分泌物含有较高浓度的糖类、氨基酸和维生素等物质,为根际微生物的生存和繁殖提供所需的营养和能源物质,从而影响根际微生物的数量和活性[6].对照组的沉积物微生物活性随时间的变化逐渐上升,从最初的210.43 μg/(g·h)上升到结束时的299.46 μg/(g·h),这可能与温度和微生物种群有关,对照组没有苦草等其他外界的干扰,微生物群落结构向着有利方向改变,适合生长的微生物种群能够快速生长[20].孙芳等[21]提出底泥中的微生物活性(FDA)和碱性磷酸酶活性(APA)能够影响底泥中氮、磷营养盐形态转换,碱性磷酸酶的分泌主要受微生物活性的影响,即APA与FDA 活性之间相关性极显著,而APA与磷含量变化趋势呈显著正相关,因此可以推断微生物活性与磷含量也呈正相关.而本实验结果却得出实验组微生物的活性与总磷呈极显著负相关(P<0.01).高光等[22]也认为APA与磷浓度呈负相关, 这是由于当水体中磷浓度较低时, APA得以激发.2.3 苦草的生长对沉积物微生物群落结构的影响应用PLFAs技术分析了苦草不同生长时期表层沉积物微生物群落结构的变化.整个生长周期共检测出C10~C20共60种脂肪酸,其中旺盛期检测出47种脂肪酸,而衰亡期检测出36种脂肪酸,检测出的脂肪酸种类随苦草的生长而减少,可能是由于沉水植物的生长吸收了大量的营养盐,导致微生物种类降低[4].对指示不同微生物类群的特征脂肪酸进行归纳和总结,可以将为生物群落分为细菌、真菌、放线菌3大类,其中细菌还可以分为G+、G-、好氧菌等几类.从图2可以看出,在整个苦草生长周期沉积物中细菌占主要成分,且在细菌中G+占主要优势,而G-则随苦草的生长呈下降趋势.一般认为G-大多对养分变化比其它类型的微生物敏感[23],G+/G-比值与沉积物的有机质含量呈负相关,沉积物的有机质含量越高,则G+/G-比值越小[24],这与本实验在衰亡期有机质含量增加而G+/G-增加不相符,可能是因为在衰亡期温度下降和苦草残体的分解影响了沉积物微环境.在3个不同时期G+/G-比值依次增加,这可能是由于苦草在生长过程中对沉积物中有机质的吸收使可利用碳源减少,这与裴雪霞及李越蜀等的研究相一致[23,25].通过沉积物微生物特征与理化性质的相关性分析(表2),表明G-与有机磷、无机磷都呈显著正相关,而G+与总磷和无机磷呈显著负相关,可能由于底泥微生物群落是推动氮、磷循环转化的主要因素,微生物群落活性及多样性的升高有助于氮、磷元素的固定以及沉积物自净能力的提高[3],这与董黎明等[26]的研究结果相一致,说明磷可能是影响沉积物中微生物群落结构及丰富度的限制性营养因素.在苦草3个生长时期检测到PLFAs包括饱和脂肪酸、支链脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸等,其中,饱和脂肪酸包括12 ∶0、13 ∶0、14 ∶0、16 ∶0、17 ∶0、18 ∶0,这6种是沉积物中含量最丰富的脂肪酸种类,其相对含量为19.1%~49.9%.对苦草不同生长时期PLFAs数据进行MDS(多维标度法)分析可以看出,在苦草不同生长时期,沉积物微生物群落结构发生了明显变化,微生物的组成结构差异显著(R=0.678,P<0.01,图3).进一步对PLFAs数据进行SIMPER分析得出:3个不同时期共同的脂肪酸主要有16 ∶0、15 ∶0 iso、15 ∶0 anteiso、16 ∶1 w7c、18 ∶1 w7c等,从差异度来讲,生长初期与旺盛期差异度为25.89%,与衰亡期的差异度为44.42%,而生长旺盛期与衰亡期的差异度为44.76%,由此可以看出,在苦草生长的不同时期沉积物的微生物群落结构发生了明显的变化.在苦草不同生长时期对照组的微生物群落结构也发生了一定的改变(图3),可能由于随时间的变化,温度、光照等外界环境的改变促使微生物群落结构向着有利方向改变[27],对PLFAs数据进行SIMPER分析得出:不同时期共同的脂肪酸主要有16 ∶0、16 ∶1 w7c、15 ∶0 iso、15 ∶0 anteiso等,从差异度来讲,生长初期与旺盛期差异度为26.41%,与衰亡期的差异度为34.54%,而生长旺盛期与衰亡期的差异度为31.33%,与实验组不同时期的差异度相比明显降低,这可能是由于苦草根系的分泌物为微生物的生长提供碳源,根系对水分的吸收改变底泥通风状态,从而影响气体和溶解态营养物质的扩散,且苦草能够从大气中输送不同的气体(氧气、气态氮) 到底泥中去[28],同时,植物通过根系分泌物向沉积物中输入大量酶和生长因子,可以改善微生物群落结构,促进其良性发展[29].而微生物的生长繁殖及活性也受到温度变化的显著影响,植物在不同季节的生长状况和代谢活动不同,对有机质的吸收利用存在差异,导致植物根际微生物群落的变化[30],同时由于根系的分泌物和脱落物对微生物具有选择和富集的作用,必然导致植物根际微生物群落多样性的变化[31].1) 苦草可以有效地减少沉积物中有机质及磷含量,衰亡期磷含量显著低于生长初期(P<0.05),生长初期到衰亡期总磷、有机磷、无机磷分别下降8.97%、7.81%、10.28%.2) 从苦草生长初期到旺盛期微生物活性显著上升(P<0.05),实验组的微生物活性大于对照组,说明沉水植物对沉积物微生物活性有一定影响;在衰亡期,对照组的微生物活性略高于实验组,可能是由于根际分泌物的影响.3) 沉水植物的生长可以增加沉积物中革兰氏阳性菌的百分含量,同时降低革兰氏阴性菌的百分含量,改变其微生物群落组成及生态学功能.相关性分析表明革兰氏阳性菌与总磷、无机磷呈显著负相关,而革兰氏阴性菌与有机磷、无机磷呈显著正相关,磷可能是影响沉积物中微生物活性及群落结构的限制性营养因素.致谢:南京师范大学地理科学学院韩城博士、范婤博士、张丁予硕士等在采样和分析中给予帮助, 在此表示感谢!【相关文献】[1] 金相灿. 沉积物污染化学.北京:中国环境科学出版社, 1992.[2] Cotner JB, Biddanda BA. Small players, large role: microbial influence on biogeochemical processes in pelagic aquatic ecosystems. Ecosystems, 2002, 5(2): 105-121.[3] 朱亮,孙凌宇,储如花等. 城市纳污河流沉积物微生物群落结构特征. 水科学进展, 2013, (1): 132-137.[4] Hilt S, Gross EM. Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Basic and Applied Ecology, 2008, 9(4): 422-432.[5] 宋玉芝,杨美玖,秦伯强等. 苦草对富营养化水体中氮磷营养盐的生理响应. 环境科学,2011, 32(9): 2569-2575.[6] 陈晓娟,吴小红,刘守龙等.不同耕地利用方式下土壤微生物活性及群落结构特性分析:基于PLFA和MicrorespTM方法.环境科学,2013,34(6):2375-2382.[7] 李琳琳,汤祥明,高光等. 沉水植物生态修复对西湖细菌多样性及群落结构的影响. 湖泊科学, 2013, 25(2): 188-198.[8] Zhao D, Liu P, Fang C et al. Submerged macrophytes modify bacterial community composition in sediments in a large, shallow, freshwater lake. Canadian Journal of Microbiology, 2013, 59(4): 237-244.[9] John B. A comparison of two methods for estimating the organic matter content of sediments. Journal of Paleolimnology, 2004, 31(1): 125-127.[10] 朱广伟. 水动力作用与浅水湖泊磷的内源负荷研究. 中国科学院南京地理与湖泊研究所(博士后研究工作报告), 2003.[11] Ruban V, Brigault S, Demare D et al. An investigation of the origin and mobility ofphosphorus in freshwater sediments from Bort-Les-Orgues Reservoir, France. Journal of Environmental Monitoring, 1999, 1(4): 403-407.[12] Ruban V, Lopez-Sanchez JF, Pardo P et al. Harmonized protocol and certified reference material for the determination of extractable contents of phosphorus in freshwater sediments-a synthesis of recent works. Fresenius’ Journal of Analytical Chemistry, 2001, 370(2/3): 224-228.[13] Schnürer J, Rosswall T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Applied and Environmental Microbiology, 1982, 43(6): 1256-1261.[14] 孙芳,郑忠明,陆开宏等. 铜锈环棱螺(Bellamya aeruginosa)生物扰动对藻华水体沉积物微生物群落结构的影响. 海洋与湖沼, 2012, (2): 357-362.[15] 朱广伟,高光,秦伯强等. 浅水湖泊沉积物中磷的地球化学特征. 水科学进展, 2003, 14(6): 714-719.[16] 朱广伟,陈英旭. 沉积物中有机质的环境行为研究进展. 湖泊科学, 2001, 13(3): 272-279.[17] 易文利. 有机质对磷素在沉积物-水-沉水植物间迁移转化的影响[学位论文]. 杨凌: 西北农林科技大学, 2008.[18] Wang S, Jin X, Zhao H et al. Effect of organic matter on the sorption of dissolved organic and inorganic phosphorus in lake sediments. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 2007, 297(1): 154-162.[19] 王立志. 沉水植物对湖泊沉积物-上覆水中磷迁移转化的影响[学位论文]. 南京: 南京师范大学, 2011.[20] 杨磊,林逢凯,胥峥等. 底泥修复中温度对微生物活性和污染物释放的影. 环境污染与防治, 2007, 29(1): 22-25.[21] 孙芳,郑忠明,陆开宏等. 底泥微生物活性对蓝藻水华水柱及沉积物间隙水氮磷分布的影响. 生态科学, 2011, 30(3): 217-222.[22] 高光,秦伯强,朱广伟等. 太湖梅梁湾中碱性磷酸酶的活性及其与藻类生长的关系. 湖泊科学,2004, 16(3):245-251.[23] 裴雪霞,周卫,梁国庆等. 长期施肥对黄棕壤性水稻土生物学特性的影响. 中国农业科学, 2010, 43(20): 4198-4206.[24] Peacock AG, Mullen MD,Ringelberg DB et al. Soil microbial community responses to dairy manure or ammonium nitrate applications. Soil Biology and Biochemistry, 2001,33(7): 1011-1019.[25] 李越蜀,郑忠明,翟海佳等. 不同模式凡纳滨对虾 (Litopenaeus vannamei) 养殖池塘沉积物酶活性及其微生物群落结构分析. 海洋与湖沼, 2012, 43(6): 1154-1260.[26] 董黎明. 利用磷脂脂肪酸表征白洋淀沉积物微生物特征. 中国环境科学, 2011, 31(11): 1875-1880.[27] 白震,何红波,解宏图等. 施肥与季节更替对黑土微生物群落的影响. 环境科学, 2008, 29(11):3230-3239.[28] 邢鹏,孔繁翔,陈开宁等. 生态修复水生植物根际氨氧化细菌的研究. 环境科学, 2008, 29(8): 2154-2159.[29] 陆松柳,张辰,徐俊伟. 植物根系分泌物分析及对湿地微生物群落的影响研究. 生态环境学报, 2011, 20(4): 676-680.[30] 梁威,吴振斌,詹发萃等. 人工湿地植物根区微生物与净化效果的季节变化. 湖泊科学, 2004, 16(4): 312-317.[31] Wasaki J, Rothe A, Kania A et al. Root exudation, phosphorus acquisition, and microbial diversity in the rhizosphere of white lupine as affected by phosphorus supply and atmospheric carbon dioxide concentration. Journal of Environmental Quality, 2005, 34(6): 2157-2166.。
南京师范大学-环境生态学(王国祥)-期末考试-复习资料第二章

第二章生物与环境生态因子:指环境中对生物的生长、发育、生殖、行为和分布有着直接或间接影响的环境要素,如温度、湿度、食物、氧气、二氧化碳和其他相关生物等。
生态因子也可认为是环境因子中对生物起作用的因子。
生态类群:多种不同的植物,对某一生态因子(如生理、形态、结构以及物候等),具有相似需求性和耐受力,表现出比较一致的适应性特征.生态型(ecotype): 分布在不同地域的同一种群,适应于分布区内各个部分的局部条件,而出现某些生理、形态上的差异,称为生态型,也称生态差型(ecocline),以表示种内生态属性的连续性变化。
光补偿点(light compensation point):光合作用合成与呼吸所消耗的碳水化合物达到平衡时的光照强度,即光合作用所固定的CO2与呼吸释放的CO2相等时的光照强度。
光饱和点(light saturation point):在一定的光强范围内,植物的光合强度随光照度的上升而增加,当光照度上升到某一数值之后,光合强度不再继续提高时的光照度值。
例如10℃为某作物的生物学零度,其中某三天12。
1℃、9.8℃、13。
5℃,则这三天的活动积温为25.6℃,其中9.8℃低于生物学零度,不统计在内.例如,某作物的生物学零度为10℃,某日温度12。
1℃,则该日有效温度为12。
1-10.0=2。
1℃,蒸腾作用(transpiration):水分以气体状态通过植物表面(主要是叶子)蒸发散溢到体外的现象。
蒸腾作用可以促进植物对水分的吸收和运转,降低植物的体温,促进营养盐的运转和分布。
土壤供水适宜、通气好、光照强、温度高、风速大,都能促进蒸腾作用。
生态对策:是生物进化过程中所形成的各种特有的生活史特征,是生物适应于特定的环境所具有的一系列生物学特性的设计。
也称为生活史对策(life history strategy)。
一、生态因子主要包括哪些类型?1、以主客体分:①生物因子(动物、植物、微生物等)②非生物因子(条件因子:温度、湿度、pH;资源因子:营养物、水、光照等。
太湖西岸沉积物中典型挥发性硫化物的分布特征研究

太湖西岸沉积物中典型挥发性硫化物的分布特征研究黄鹤勇;刘宪圣;许晓光;赵艳萍;王国祥【期刊名称】《生态科学》【年(卷),期】2018(037)004【摘要】挥发性硫化物是微囊藻死亡分解产生的主要藻源性嗅味污染物,伴随微囊藻水华的暴发而大量产生,不仅严重影响了水体的水质状况,也给沿湖居民以及水生生物带来危害.本研究分析了夏季太湖西岸近岸带上覆水以及沉积物中三种典型挥发性硫化物二甲硫醚(DMS)、二甲二硫醚(DMDS)、二甲三硫醚(DMTS)的分布特征及其与营养盐之间的关系.结果表明,挥发性硫化物(VSCs)在沉积物表层含量最高,其中DMDS浓度最高达262.25 ng·g-1,DMTS浓度最高达50.90 ng·g-1.空间分布上:在近岸带,表层沉积物(0 cm-4 cm)自然芦苇带内挥发性硫化物的含量低于人工挖掘的漕沟内;近岸带嗅味物质明显高于湖区内.研究认为,藻类大量聚集死亡沉积至湖泊底部,降低沉积物的氧化还原电位,促进了致嗅物质的产生,为湖泛的暴发留下隐患.建议在近岸带蓝藻聚集区适时打捞,底泥疏浚等生态治理措施,降低湖泛风险.【总页数】8页(P16-23)【作者】黄鹤勇;刘宪圣;许晓光;赵艳萍;王国祥【作者单位】南京师范大学地理科学学院, 南京 210023;南京师范大学分析测试中心, 南京 210023;南京师范大学地理科学学院, 南京 210023;南京师范大学环境学院, 南京 210023;南京师范大学环境学院, 南京 210023;南京师范大学环境学院, 南京 210023【正文语种】中文【中图分类】X131.2【相关文献】1.太湖西岸沉积物中两种致嗅物质的分布特征 [J], 刘宪圣;许晓光;黄鹤勇;赵艳萍;陈翀;赵月;王国祥2.太湖五里湖区表层沉积物中酸挥发性硫化物和同步提取金属 [J], 李金城;宋进喜;王晓蓉3.东湖沉积物中酸挥发性硫化物的季节、深度分布特征研究 [J], 方涛;张晓华;徐小清4.太湖西岸典型区域沉积物的硫铁分布特征及环境意义 [J], 吴松峻;汪旋;季秋忆;王明玥;赵艳萍;王国祥5.太湖梅梁湾沉积物中酸挥发性硫化物垂直变化特征研究 [J], 宋进喜;李金成;王晓蓉;刘康;马俊杰因版权原因,仅展示原文概要,查看原文内容请购买。
蓝藻的休眠期去除方法及其设备[发明专利]
![蓝藻的休眠期去除方法及其设备[发明专利]](https://img.taocdn.com/s3/m/572679e2cc175527062208c5.png)
专利名称:蓝藻的休眠期去除方法及其设备专利类型:发明专利
发明人:杨浩,王国祥
申请号:CN200810244179.X
申请日:20081218
公开号:CN101532284A
公开日:
20090916
专利内容由知识产权出版社提供
摘要:蓝藻的休眠期去除方法及其设备。
方法的步骤如下,(1)在蓝藻的休眠期,采用泵筒通过吸嘴将处于水-泥界面处的休眠期蓝藻藻体、死亡藻体、部分粘土矿物和水一起吸取到船上;(2)将吸取到的休眠期蓝藻藻体、死亡藻体、部分粘土矿物和水充分分离;(3)将分离出的粘土矿物和水返回湖体。
该方法所使用的蓝藻的休眠期去除设备,其特征在于,结构如下:在污泥泵的入口上连接有软管和吸嘴;该污泥泵的出口与过滤器连接;过滤器的出口接排污口。
本发明的效果是:由于藻类暴发有一定的时间性和阶段性,在藻类非暴发阶段,藻体往往富集在沉积物的表面,在这个阶段对藻体进行杀灭和拦阻,可以非常有效的防止藻类的大量暴发。
申请人:南京师范大学
地址:210097 江苏省南京市宁海路122号
国籍:CN
代理机构:南京知识律师事务所
代理人:栗仲平
更多信息请下载全文后查看。
洪泽湖面临的生态环境问题及其成因

洪泽湖面临的生态环境问题及其成因
葛绪广;王国祥
【期刊名称】《人民长江》
【年(卷),期】2008(039)001
【摘要】受淮河及湖泊周围地区环境污染和人类不合理的开发活动的影响,我国第4大淡水湖泊洪泽湖水生生态环境呈现退化趋势.分析了洪泽湖湖面萎缩、湿地锐减、水质下降、灾害频发、生物多样性减少等问题;论述了洪泽湖生态环境问题的成因.围湖造田、围网养殖、泥沙淤积是湖面萎缩的主要因素,入湖污染物的总量超过洪泽湖水体自净能力造成水质下降,自然因素和人为因素造成灾害频发,生物资源的不合理开发、生态环境不稳定、外来物种的入侵使生物多样性减少.提出了保护和恢复洪泽湖和谐生态环境的有效措施.
【总页数】3页(P28-30)
【作者】葛绪广;王国祥
【作者单位】南京师范大学,地理科学学院,江苏,南京,210046;南京师范大学,地理科学学院,江苏,南京,210046
【正文语种】中文
【中图分类】X52
【相关文献】
1.洪泽湖流域生态环境问题及治理对策 [J], 张利民;刘伟京;尤本胜;夏明芳;张磊;王春
2.全球性生态环境问题的制图表示——以《星球中外地图集》中“当今世界面临的全球性生态环境问题”图组为例 [J], 陈毓芬;殷畅
3.洪泽湖湿地退化成因及生态修复实践——以江苏泗洪洪泽湖湿地国家级自然保护区为例 [J], 李成之
4.生态主义对生态环境问题成因及解决方案的分析 [J], 黄晓云
5.保护生态环境刻不容缓——谈当前人类面临的生态环境问题 [J], 任平
因版权原因,仅展示原文概要,查看原文内容请购买。
利用水生植物净化水体中的悬浮泥沙

利用水生植物净化水体中的悬浮泥沙
郭长城;王国祥;喻国华
【期刊名称】《环境工程》
【年(卷),期】2006(024)006
【摘要】针对特定的滨江城市水环境问题,利用湿地植物风车草(Cyperus alternifolius)、菖蒲(Gladiolus hybridus)、菱(Vallisneria spiralis)、金鱼藻(Ceratophyllum demersum)、轮叶黑藻(Hydrilla verticillata)和亚洲苦草(Vallisneria spiralis)构建的湿地植物生态物理模型进行了含沙水体泥沙净化的实验研究.结果表明,湿地植物系统对不同悬浮泥沙含量的水体具有不同的净化效果,净化效果最好的湿地植物分别是金鱼藻、黑藻、菱、菖蒲和风车草.随水体含沙量增加净化效果有所增强.不同粒径的悬浮泥沙在湿地植物群落中的沉降、吸附和过滤的效果不同.
【总页数】3页(P31-33)
【作者】郭长城;王国祥;喻国华
【作者单位】南京水利科学研究院,江苏,210029;南京师范大学地理科学院,江苏,210097;南京水利科学研究院,江苏,210029
【正文语种】中文
【中图分类】X7
【相关文献】
1.利用水生植物净化水体中的悬浮泥沙 [J], 郭长城;王国祥
2.水生植物净化水体中氮磷含量的研究进展 [J], 张扬;杨友才;李燕子
3.利用水生植物净化富营养化水体的研究进展 [J], 朱斌;陈飞星;陈增奇
4.水生植物净化静动态水体的试验研究 [J], 武雅;马凡凡;席粉鹊;李倩
5.水生植物净化水体作用的研究进展 [J], 陈罡
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第二章生物与环境生态因子:指环境中对生物的生长、发育、生殖、行为和分布有着直接或间接影响的环境要素,如温度、湿度、食物、氧气、二氧化碳和其他相关生物等。
生态因子也可认为是环境因子中对生物起作用的因子。
生态类群:多种不同的植物,对某一生态因子(如生理、形态、结构以及物候等),具有相似需求性和耐受力,表现出比较一致的适应性特征.生态型(ecotype):分布在不同地域的同一种群,适应于分布区内各个部分的局部条件,而出现某些生理、形态上的差异,称为生态型,也称生态差型(ecocline),以表示种内生态属性的连续性变化。
光合作用合成与呼吸所消耗的碳水化合物达到平衡时的光照强度,即光合作用所固定的CO2与呼吸释放的CO2相等时的光照强度。
光饱和点(lightsaturation point):在一定的光强范围内,植物的光合强度随光照度的上升而增加,当光照度上升到某一数值之后,光合强度不再继续提高时的光照度值。
例如10℃为某作物的生物学零度,其中某三天12.1℃、9.8℃、13.5℃,则这三天的活动积温为25.6℃,其中9.8℃低于生物学零度,不统计在内。
例如,某作物的生物学零度为10℃,某日温度12.1℃,则该日有效温度为12.1-10.0=2.1℃,蒸腾作用(transpiration):水分以气体状态通过植物表面(主要是叶子)蒸发散溢到体外的现象。
蒸腾作用可以促进植物对水分的吸收和运转,降低植物的体温,促进营养盐的运转和分布。
土壤供水适宜、通气好、光照强、温度高、风速大,都能促进蒸腾作用。
生态对策:是生物进化过程中所形成的各种特有的生活史特征,是生物适应于特定的环境所具有的一系列生物学特性的设计。
也称为生活史对策(life history strategy).一、生态因子主要包括哪些类型?1、以主客体分:①生物因子(动物、植物、微生物等)②非生物因子(条件因子:温度、湿度、pH;资源因子:营养物、水、光照等。
)2、以因子性质分类:①气候因子:如温度、湿度、光、降水、风、气压和雷电等。
②土壤因子:土壤是在岩石风化后在生物参与下所形成的生命与非生命的复合体,土壤因子包括土壤结构、土壤有机和无机成分的理化性质及土壤生物等。
③地形因子:如地面的起伏,山脉的坡度和阴坡阳坡等,这些因子对植物的生长和分布有明显影响。
④生物因子:包括生物、生物之间的各种相互关系,如捕食、寄生、竞争和互惠共生等。
⑤人为因子:人为因子是生物因子的一种类型,指人及人类活动产生的二、生态因子作用的一般规律。
1、综合作用:多因子对生物的联合作用、因子之间的相互作用2、主导因子作用:在诸多因子中,对生物起决定作用的生态因子称为主导因子,主导因子的作用就是对生物群体生长、发育、演替等生态过程有决定意义的作用。
3、直接和间接作用:一些生态因子直接对生物及其群落产生影响,而一些生态因子通过对其他因子的影响发生作用,这称为间接作用。
如地形因子中起伏程度、坡向、坡度、海拔高度及经纬度等对生物的作用不是直接的。
4、阶段性作用:在生物及其群落生长发育的不同阶段,各种因子的作用特性差异明显。
5、因子的不可替代性和补偿作用:各生态因子对生物的作用虽然不尽相同,但都不可缺少,一个因子的缺失不能由另一个因子来替代。
但某一因子的数量不足,有时可以靠另一因子的加强而得到调剂和补偿。
例如如果光照不足,可以增加二氧化碳浓度来补偿。
三、限制因子与Liebig最小因子定律及其应用。
1、限制因子规律(不同于木桶原理):任何一种生态因子只要接近或超过生物的耐受范围,它就成为这种生物的限制因子。
限制因子(Limitingfactors):生物的生存和繁殖依赖于各种生态因子的综合作用,其中限制生物生存和繁殖的关键性因子称为限制因子。
限制因子的特点:xx、狭域性、敏感性限制因子作用的补充和完善:①限制因子定律只有在严格稳定状态下适用;②因子之间的相互作用,因子的补偿作用;③限制因子是对物种、种群而言的,不同生物乃至同一生物种群的不同发育阶段,限制因子可能会发生变化;④限制因子强调的是因子的不足,实际上因子过高也会对生物生长发育产生抑制作用。
2、Liebig(利比希)最小因子规律:植物的生长取决于处在最小量状况的食物的量。
应用:1、园林绿化的植物品种引种栽培与管理;2、农业生产中,农作物品种的引种驯化;3、农业生产中,合理科学施肥;4、渔业生产中,天然饵料生物的增殖;5、有害生物防治与控制;6、生态修复重建中,生境修复与关键生物引种。
四、耐受限定律、生态幅的概念以及应用。
Shelford(xx)耐性定律:任何一个生态因子在数量和质量上的不足或者过多,即当其接近或达到某种生物的耐受限度时,就会使该生物衰退或不能生存。
生态幅(耐受力、耐受幅、生态价):每一个种对环境因子适应范围的大小即生态幅,亦称生态价。
五、举例说明指示生物及意义。
指示生物:一些生物在与环境相互作用、协同进化过程中,留下了明显的形态、生理等特征,可以反映和指示环境的某些特征。
意义:1、气候指示:热带地区:椰树;寒带:针叶植物;亚热带:xx栎。
2、水分指示:水生植物在水湿环境中生长,使它具有柔嫩、硕大的叶子,但是根系并不发达。
莲、芦苇的生长则反应了水湿环境。
旱生植物在干旱环境中生长,使它的叶子已经退化成细小的刺,以减少水分的蒸腾;但是根系很发达,能从很深很广的地下吸取水分。
如骆驼刺的生长反应了干旱环境。
3、土壤指示:铁芒萁是我国长江流域一带红土壤等酸性土壤的指示植物;碱蓬的生长则指示了盐碱性的土壤环境;弹尾类和蜱螨类土壤动物指示土壤重金属污染。
4、地质指示:海州香薷(“铜草”)生长茂盛的地方就有铜矿;鸡脚蘑、凤眼兰等植物生长旺盛的地方,往往是藏金之地。
5、季节(时间)指示:雁子南飞,预示冬季的到来;报春花开花、柳树吐絮,则标志着春天的到来;蛇床花凌晨3点开放;牵牛花凌晨4点开放等。
6、环境污染指示:苔藓对重金属(砷、镉、铬、铜、铁、铅、镍、矾、锌)污染的指示意义;菠菜可以探测重金属;菖兰可探测HF、氯化氢;苜蓿和黑麦草可探测铁、硫;荨麻可探测过氧乙酰硝酸;玉米可探测HF、SO2、重金属;矮牵牛可探测乙烯、烃、甲醛;烟草可探测臭氧、NO等等。
7、环境演变指示:地层年龄;古地理气候变迁;冰川消长;太阳黑子活动等。
六、简述光的生态作用以及生物的适应性。
1、光强的生态作用与生物的适应①光强对光合作用的影响光补偿点:光合作用合成与呼吸所消耗的碳水化合物达到平衡时的光照强度,即光合作用所固定的CO2与呼吸释放的CO2相等时的光照强度。
光饱和点:在一定的光强范围内,植物的光合强度随光照度的上升而增加,当光照度上升到某一数值之后,光合强度不再继续提高时的光照度值。
xx现象:植物因受光不足,不能形成叶绿素而呈现黄色、机械组织不发达、茎细软、伸长较快、叶子不舒展的现象。
某些植物的黄化后具有较高的经济价值,如豆芽、韭黄、石刁柏等。
②影响光强的环境因素a、纬度、天气、地形(山地的阴坡、阳坡);b、植被(高大植物、森林植物顶冠层的遮光);c、水体(光在水体中的衰减、水体消光系数、光补偿深度等);d、大气中的颗粒物组成及含量。
③植物对光照强度的适应:据需光度,植物的生态型分为阳生植物(heliophyte,喜阳植物heliophyllus)适应性特点:叶一般小而厚,枝叶稀疏、树冠透光、枝下高较高。
光补偿点相当于全光照的3%~5%。
如:xx、xx、柳、桦、松、杉和栓皮栎。
阴地植物(sciophyte,喜阴植物、适阴植物sciophiles)适应性特点:xx一般大而薄,枝xx浓密、枝下高较矮。
光补偿点相当于全光照的0.5%~1%。
如:人参、三七、半夏、细辛、铁杉、观音座莲、山酢浆草、连钱草、紫果云杉、红豆杉。
2、光质的生态作用与生物的适应①光质的生态效应:生理有效辐射、生理无效辐射。
光谱成分红、橙、黄、绿、青、蓝、紫,其随空间发生变化的一般规律为:短波光随纬度增加而减少,随海拔升高而增加。
在时间变化上,冬季长波光增多,夏季短波光增多;一天之内中午短波光较多,早晚长波光较多.生理有效辐射:植物光合作用利用的光谱范围在可见光区(380~760 nm)大部分波段的光,这部分辐射称为生理有效辐射。
红、橙光主要被叶绿素吸收,对叶绿素合成有促进作用,糖的合成;蓝紫光也能被叶绿素和类胡萝卜素吸收,促进蛋白质合成;生理无效辐射:绿光等很少被植物吸收利用的辐射。
②影响光质的主要因素:海拔高度、空气湿度、大气中的尘埃、臭氧层、植物顶冠层。
③有色薄膜的生态应用:有色薄膜可改变光质影响作物生长,达到增产,改善品质的目的。
实验研究表明:浅蓝色薄膜育秧与无色薄膜相比,浅蓝色薄膜秧苗根系较粗壮,插后成活快,生长健壮,叶色浓绿,鲜重和干重都有增加,测定的淀粉、蛋白质含量较高。
主要是因为太阳光通过有色薄膜时,被选择透过和吸收,这样薄膜内的光质因薄膜颜色不同而发生变化。
如浅蓝色薄膜可以大量透过光合作用所需的380-490纳米的光(透过率60%以上),因而有利于植物的光合和代谢过程。
3、光周期的生态作用及生物的适应①光周期:昼夜节律、季节节律光周期现象:受光照和黑夜时间长短变化所制约,植物的各种生理活动依次有规xx出现的现象。
②影响光周期的因素:由于地球的自转和公转规律恒定,因此,光周期是地球上最严格和最稳定的周期性变化现象。
影响光周期的主要因素有季节变化和纬度变化。
③植物对光周期的适应长日照植物:通常是在日照时间超过一定数值时,植物才能够开花,否则只能进行营养生长,不能形成xx。
如:凤仙花、冬小麦、大麦、油菜、菠菜、甜菜、甘蓝和萝卜等。
短日照植物:通常是在日照时间短于一定数值植物才能够开花,否则只能进行营养生长而不开花。
如:牵牛、菊类、水稻、玉米、大豆、烟草、麻和棉等,通常在早春或深秋开花。
中日性植物:中日性植物只有在某一定中等长度的日照条件下才能开花,而在较长或较短日照下均保持营养生长状态的植物。
例如甘蔗,最适日照长度是12.5小时,再长和再短的日长下都不开花。
中间性植物:在任何日照条件下都可以开花,如番茄、黄瓜等。
动物的光周期:xx:xx的迁徙由日照xx的变化所引起;鸟类的开始生殖时间也受日照长度的周期性变化决定;鱼类的生殖和迁移也有光周期现象;昆虫的光周期现象;哺乳动物的生殖和换毛:长日照兽类:随着春天日照长度的逐渐增加而开始生殖,如雪貂、野兔和刺猬等,尤其是高纬度的种类;短日照兽类:随着秋天日照逐渐缩短,开始交配繁殖,到春天条件较好时产出幼子。
七、温度对生物作用的“三基点”和积温的生态学意义。
①三基点:温度是生物生命活动不可缺少的因素,任何生物都生活在具有一定温度的外界环境中并受着温度变化的影响。
生物在长期的演化过程中,各自选择了自己最合适的温度,通常分为最低温度、最适温度和最高温度。