第七章 内膜系统与蛋白质分选
细胞生物学 第七章_内膜系统与蛋白质的分选
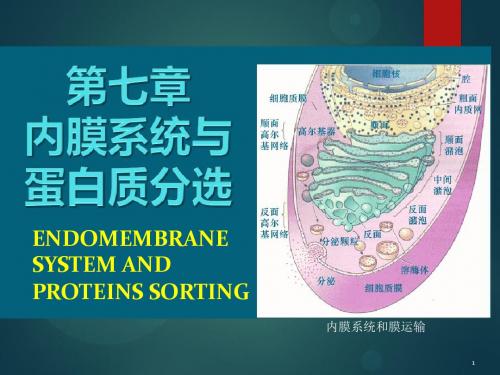
将细胞内部区分为不同的功能区域,保证各种生化反应所需的独
特的环境。
2
主要内容
内膜系统 内质网 高尔基体 溶酶体
蛋白质分选及其基本原理 膜泡运输
3
第一节 内膜系统 Endomembrane System
Endomembrane System : The structural and functional relationship organelles
混合功能氧化酶将底物羟基化 P354, Fig. 9-16
(1)被氧化的有毒底物(脂溶性)与细胞色素P-450结合; (2)细胞色素P-450中的铁原子被 NADPH还原; (3)氧与细胞色素P-450结合; (4)底物被一个氧原子氧化,另一个氧用于形成 水。被氧化的有毒底物带上羟基,由脂溶性转变为水溶性被排出体外。
sER合成磷脂酰胆碱 P352, Fig. 9-14
★ 内质网膜中脂肪酸与胞质溶胶中的磷酸甘油结合,然后脱磷生成甘油脂肪酸; ★ 在胆碱磷脂转移酶的作用下,将胞质中的CDP-胆碱与甘油脂肪酸结合形成磷脂酰胆碱; ★ 新合成的磷脂酰胆碱在内质网膜中磷脂转位酶的作用下翻转到内质网的腔面。
15
Detoxification of organiccompounds in liver cells. System of oxygenases ---cytochrome p450 family
O-linked: linked to the hydroxyl group serine or threonine via GalNac (in Golgi)
11
Quality control of newly synthesized proteins ---The role of N-linked glycosylationin ER protein folding
细胞生物学 翟中和版 总结笔记第七章

Cell biology细胞生物学第七章真核细胞内膜系统、蛋白质分选与膜泡运输细胞内被膜区分类:细胞质基质、细胞内膜系统、有膜包被的细胞器第一节细胞质基质的含义和功能一、细胞质基质的含义(1)含义:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质主要含有:(1)与代谢有关的许多酶(2)与维持细胞形态和物质运输有关的细胞质骨架结构细胞质基质是一个高度有序的体系,细胞质骨架纤维贯穿在粘稠的蛋白质胶体中,多数的蛋白质直接或间接地与骨架结合,或与生物膜结合,从而完成特定的功能。
细胞质基质主要是由微管、微丝和中间丝等相互联系形成的结构体系,蛋白质和其他分子以凝聚或暂时的凝聚状态存在,与周围溶液的分子处于动态平衡。
差速离心获得的胞质溶胶的组分和细胞质基质溶液成分很大不同。
胞质溶胶中的多数蛋白质可能通过弱键结合在基质的骨架纤维上。
二、细胞质基质的功能(1)蛋白质分选和转运N端有信号序列的蛋白质合成之后转移到内质网上,通过膜泡运输的方式再转运到高尔基体。
其他蛋白质的合成都在细胞质基质完成,并根据自身信号转运到线粒体、叶绿体、细胞核中,也有些蛋白驻留在细胞质基质中。
(2)锚定细胞质骨架(3)蛋白的修饰、选择性降解1 蛋白质的修饰辅基、辅酶与蛋白的结合磷酸化和去磷酸化糖基化N端甲基化(防止水解)酰基化2 控制蛋白质寿命N端第一个氨基酸残基决定寿命细胞质基质能够识别N端不稳定的氨基酸信号将其降解,依赖于泛素降解途径3 降解变性和错误折叠的蛋白质4 修复变性和错误折叠的蛋白热休克蛋白的作用第二节细胞内膜系统及其功能细胞内膜系统是指在结构、功能乃至发生上相互关联、由膜包被的细胞器或细胞结构。
研究方法:电镜技术免疫标记和放射自显影离心技术和遗传突变体分析一、内质网的形态结构和功能内质网是由封闭的管状或扁平囊状膜系统及其包被的腔形成的互相沟通的三维网络结构。
(一)内质网的两种基本类型糙面内质网和光面内质网。
糙面内质网:扁囊状整齐附着有大量核糖体功能:合成分泌性蛋白和膜蛋白光面内质网:分支管状,小功能:脂质合成,出芽位点部分细胞合成固醇类激素糙面内质网有20多种和光面内质网不同的蛋白,说明有特殊装置隔开两种内质网的组分。
细胞学第7章复习题

第七章真核细胞内膜系统、蛋白质分选与膜泡运输1一.名词解释1.细胞内膜系统:位于细胞质内,在结构、功能及发生上密切相关的内膜包绕形成的细胞器或细胞结构,是真核细胞特有的结构。
主要包括内质网、高尔基体和溶酶体等。
2.初级溶酶体:是指刚从高尔基体的边缘膨大分离出来,还未同消化物融合的潜伏状态,无活性。
3.次级溶酶体:是酶在进行或完全消化作用的溶酶体,内含水解酶和相应的底物。
根据自身消化物质得来源不同,分为自噬溶酶体和异噬溶酶体4. 细胞质基质:真核细胞的细胞质中除去可分辨的细胞器以外的胶状物质,主要含有与中间代谢有关的酶和维持细胞形态、胞内物质运输有关的细胞质骨架结构等。
二.填空题1.在内质网上合成的蛋白主要包括分泌蛋白、膜整合蛋白、细胞器驻留蛋白等。
2.蛋白质的糖基化修饰主要分为N-连接的糖基化修饰,指的是蛋白质上的天冬酰胺残基与N-乙酰葡萄糖胺直接连接,和O-连接的糖基化修饰,指的是蛋白质上的丝氨酸或苏氨酸残基与N-乙酰半乳糖胺直接连接。
3.肌细胞中的内质网异常发达,被称为肌质网。
4.原核细胞中核糖体一般结合在细胞质膜上,而真核细胞中则结合在粗面内质网上。
5.真核细胞中,光面内质网是合成脂类分子的细胞器。
6.内质网的标志酶是葡萄糖-6-磷酸酶。
7.细胞质中合成的蛋白质如果存在信号肽,将转移到内质网上继续合成。
如果该蛋白质上还存在停止转移序列,则该蛋白被定位到内质网膜上。
8.高尔基体的标志酶是糖基转移酶。
9.具有将蛋白进行修饰、分选并分泌到细胞外的细胞器是高尔基体。
10.被称为细胞内大分子运输交通枢纽的细胞器是高尔基体。
11.蛋白质的糖基化修饰中,N-连接的糖基化反应一般发生在内质网中,而O-连接的糖基化反应则发生在和高尔基体中。
12.蛋白质的水解加工过程一般发生在高尔基体中。
13.从结构上高尔基体主要由顺面膜囊、中间膜囊和反面膜囊和囊泡组成。
14.植物细胞中与溶酶体功能类似的结构是圆球体、中央液泡和糊粉粒。
细胞生物学 第七章 细胞内膜系统及蛋白质分选与泡膜运输

周围大小不等的囊泡——物质运输
高尔基体与细胞骨架关系密切; 高尔基的膜囊上存在微管的马达蛋白(
kinesin)和微丝的马达蛋白(myosin)。最近 还发现特异的血影蛋白(spectrin)网架 。它 们在维持高尔基体动态的空间结构以及复杂的 膜泡运输中起重要的作用。
❖ 残余小体(residual body),次级溶酶体未被消化 的残渣,又称后溶酶体。
用电镜细胞化学技术显示其中含有的酸性磷酸酶, M:线粒体,L:溶酶体(朴英杰)
动物细胞溶酶体系统示意图
溶酶体膜的特征: 嵌有质子泵,形成和维持溶酶体中酸性的内环境; 具有多种载体蛋白用于水解的产物向外转运; 膜蛋白高度糖基化,可能有利于防止自身膜蛋白的
❖ 二、内膜系统的结构与功能 ❖ (二)高尔基体( Golgi complex ) ❖ 2、功能
❖ (3)蛋白酶的水解和其他加工过程
蛋白质在高尔基体中酶解加工的几种类型
无生物活性的蛋白原(proprotein)高尔基体切除 N-端或两端的序列成熟的多肽。如胰岛素、胰高血糖 素及血清白蛋白等
蛋白质前体高尔基体水解同种有活性的多肽,如 神经肽等
一种分选途径。
❖ 二、内膜系统的结构与功能
❖ (二)高尔基体( Golgi complex ) ❖ 2、功能
(1)高尔基体与细胞的分泌活动 ❖ 蛋白质的分选及其转运 ❖ 溶酶体酶的分选 (2)蛋白质糖基化及其修饰
(3)蛋白酶的水解和其他加工过程
蛋白质糖基化类型
特征
N-连接
O-连接
1. 合成部位
2. 合成方式
第七章 真核细胞内膜系统蛋白质分选与膜泡运输优秀课件

(一)内质网的两种基本类型
SER由分支小管或圆形小泡构成,表面光滑,无核糖体。是脂 质合成的重要场所。作为出芽的位点,将内质网上合成的蛋白 质或脂质转移到高尔基体内。 多分布脂类合成旺盛细胞(肝、肾上腺皮质细胞)。
细胞物质运输、能量交换和信号转导通路 与细胞质骨架相关的功能 维持细胞形态、细胞运动、大分
子定位等。 蛋白质的分选转运、修饰和蛋白质选择性降解
蛋白质的修饰; 控制蛋白质寿命(泛素依赖的降解途径); 降解变性和错误折叠的蛋白质(泛素依赖的降解途径); 帮助变性或错误折叠的蛋白质重新折叠(热休克蛋白)。
真核细胞的3大基本结构体系
生物膜系统
遗传信息表达结构系统
细胞骨架系统
细胞内被膜: 1. 细胞质基质; 2. 细胞内膜系统; 3. 其他由膜包被的 各类细胞器(如线 粒体、叶绿体)
细胞内区室化是真核细胞结构和功能的基本特征之一。
第一节 细胞质基质的涵义与功能
一、细胞质基质的涵义
定义:在真核细胞的细胞质中,除去可分辨的细胞器以外 的胶状物质,称细胞质基质。(扩散速率仅为水溶液1/5)
甘油磷脂 磷脂酸盐
甘油磷脂
二磷酸甘油酯
磷脂酰胆碱
酰基转移酶
胆碱磷酸酯转移酶
磷脂转位因子
合成的磷脂由内质网向其它膜转运主要有两种方式: ⑴ 以出芽的方式转运到高尔基体、溶酶体和细胞膜上; ⑵ 凭借水溶性的载体蛋白-磷脂转运蛋白(phospholipid exchange
细胞质基质是细胞的重要组分,其体积约占细胞质的一半。 主要成分:参与中间代谢的数千种酶、细胞质骨架结构、
细胞生物学 第7章 真核细胞内膜系统蛋白质分选与膜(共103张PPT)
一,其意义在于:大大增加了细胞内膜的表面积;为多种酶特
别是多酶体系提供了大面积的结合位点;酶系统的隔离与连接;
蛋白质、糖、脂肪的合成、加工和包装;运输分泌物;扩散屏
障及膜电位建立;离子梯度的维持等。
二、内质网的形态结构与功能
(一)、内质网的形态结构及化学组成
②赋予蛋白质传导信号的功能;
③某些蛋白只有在糖基化之后才能正确折叠。
糖基一般连接在4种氨基酸上,分为2种:
(O-linked glycosylation):与
Ser、Thr和Hyp的OH连接,连接的糖为半乳糖
或N-乙酰半乳糖胺,在高尔基体上进行。
(N-linked glycosylation):与
丝氨酸、苏氨酸、
羟赖氨酸、羟脯氨酸
一般1~4个糖残基,
但ABO血型抗原较长
N—乙酰半乳糖胺等
内质网上进行N-连接的糖基化。
糖分子首先被糖基转移酶转移到膜上的磷酸多萜醇
(dolichol phosphate)分子上,装配成寡糖链。
再被寡糖转移酶转到新合成肽链特定序列(Asn-X-Ser
或Asn-X-Thr)的天冬酰胺残基上。
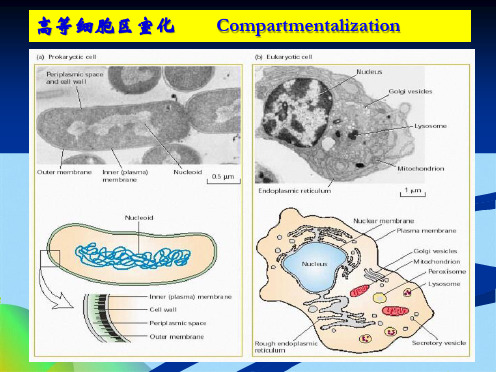
高等细胞区室化
Compartmentalization
质膜
内膜
生物膜
原核细胞 – 质膜构成的单一区室
真核细胞 – 内膜 – 各种细胞器
内膜系统出现的意义:
各自独立,各司其职
相互依存,协调同一
大大提高了细胞代谢反应的效率
细胞核
细
胞
细胞质
过氧化物酶体
线粒体
细胞骨架
真核细胞内膜系统、蛋白质分选与膜泡运输

1.清除无用的生物大分子、衰老的细胞器及衰老损伤和死亡的细胞 2.防御功能 3.其它重要的生理功能
⑴ 作为细胞内的消化“器官”为细胞提供营养; ⑵ 在分泌腺细胞中,溶酶体常常含有摄入的分泌颗粒,可能参与分泌过程的调节; ⑶ 某些特定细胞编程性死亡及周围活细胞对其清除; ⑷ 精子的顶体(acrosome)相当于特化的溶酶体,在受精过程中的能溶解卵细胞膜。
二、高尔基体的形态结构与功能
高尔基体结构成分的标志细胞化学反应嗜锇反应:cis面膜囊被特异地染色焦磷酸硫胺素酶(TPP酶):trans面的膜囊胞嘧啶单核苷酸酶(CMP酶)或核甘酸二磷酸酶: trans面1~2层囊膜扁平囊膜烟酰胺腺嘌呤二核苷酸磷酸酶(NADP酶)或甘露糖酶:中间几层
嗜锇反应
甘露糖酶
动物细胞
植物细胞
1
数量:平均为每细胞20个。在低等真核细胞中, 有时只有1~2个,有的可达一万多个。在分泌功能旺盛的细胞中, 高尔基复合体都很多。
2
分布:只存在于真核细胞中, 原核细胞中则无。
3
主要的酶有糖基转移酶、磺基-糖基转移酶、氧化还原酶、磷酸酶、蛋白激酶、甘露糖苷酶、转移酶和磷脂酶等。
4
标志酶为糖基转移酶。
溶酶体几乎存在于所有的动物细胞中。溶酶体是单层膜围绕、内含多种酸性水解酶类的囊泡状细胞器。其主要功能是进行细胞内的消化作用。
溶酶体的形态结构与类型
溶酶体的功能
溶酶体的发生
溶酶体与过氧化物酶体
三、溶酶体的形态结构与功能
No.1
溶酶体(lysosome)是单层膜围绕、内含多种酸性水解酶类的囊状细胞器,其主要功能是进行细胞内的消化作用。
M6P是溶酶体水解酶分选的重要识别信号。所有糖蛋白离开ER都具有N-连接的寡糖链,只有溶酶体水解酶的寡糖链被磷酸化形成M6P,在每个水解酶上有其特异的信号斑(signal patch),再通过两种酶的相继催化作用而形成。
第七章 真核细胞内膜系统、蛋白质分选与膜泡运输

二、细胞质基质的功能
• 1、细胞质担负着一系列重要的功能: • ①许多中间代谢过程都在细胞质基质中进行,如糖酵 解过程、磷酸戊糖途径、糖醛酸途径、糖原的合成与 部分分解过程等等。②蛋白质的合成与脂肪酸的合成 也在细胞质基质中进行(P172,L2,WR9) 。 • 2、空间定位:通过与支架蛋白分子间的选择性结合, 使生物大分子锚定在细胞骨架三维空间的特定区域 (P173,L14,W19) 。 • 3其他还有①蛋白质的修饰(P173,L17,W1) ;②控制 蛋白质的寿命(P173,LR3,W1) ;③降解变性和错误 折叠的蛋白质(P174,LR15,W1) ;④帮助变性或错 误折叠的蛋白质重新折叠形成正确的分子构象 (P174,LR7,W1) 。
一、内质网的形态结构与功能
• 概述 • (一)内质网的两种基本类型 • (二)内质网的功能
概述
• 1、内质网(Endoplasmic reticulum, ER)是由封闭 的管状或扁平囊状膜系统及其包被的腔形成互相沟通 的三维网络结构(P175,LR5,W1) 。 • 2、功能:①大大增加了细胞内膜的表面面积 (P176,L1,W5) 。②通过分隔,更有利于它们的加工 和运输(P176,L3,WR12) 。③是蛋白质、脂质和糖 类合成的基地(P176,L4,W1) 。 • 3、发现、命名:1945年K. R. Porter发现 (P176,L7,WR3) 。微粒体包含内质网膜与核糖体两 种基本组分(P176,L13,WR4) 。
第二节 细胞内膜系统及其功能
• • • • 概述 一、内质网的形态结构与功能 二、高尔基体的形态结构与功能 三、溶酶体的形态结构与功能
பைடு நூலகம்
概述
• 细胞内膜系统(Endomembrane system)是在结构、功能乃至发生上相 关的,由膜围绕的细胞器或细胞结构。 主要包括①内质网、②高尔基体、③溶 酶体、④胞内体和⑤分泌泡等 (P175,L4,W1)。
翟中和细胞生物学第七章总结2(名词解释)

第七章真核细胞内膜系统、蛋白质分选与膜泡运输1.细胞质基质:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质,也称胞质溶胶,内含水、无机离子、酶以及可溶性大分子和代谢产物。
21、许多中间代谢过程在细胞质基质中进行。
包括糖酵解过程、磷酸戊糖途径、糖醛酸途径、糖原合成与分解以及蛋白质与脂肪酸的合成等。
2、细胞质骨架是细胞质基质的主要结构成分,与维持细胞形态、细胞运动、物质运输及能量传递有关,而且也是细胞质基质结构体系的组织者,为细胞质基质中其他成分和细胞器提供锚定位点。
3、与蛋白质的修饰及选择性降解有关。
①蛋白质的修饰,在细胞质中发生的蛋白质修饰的类型主要有:辅酶或辅基与酶的共价结合;磷酸化与去磷酸化,用以调节很多蛋白质的生物活性;糖基化作用;对某些蛋白质的N端进行甲基化修饰;酰基化。
②控制蛋白质的寿命。
③降解变性和错误折叠的蛋白质。
④帮助变性或错误折叠的蛋白质重新折叠,形成正确的分子构象。
这一功能主要靠热休克蛋白来完成。
3①辅酶或辅基与酶的共价结合。
②磷酸化与去磷酸化,用以调节很多蛋白质的生物活性。
③糖基化作用:糖基化主要发生在内质网和高尔基体中,在细胞质基质中发现的糖基化是指在哺乳动物的细胞中把N-乙酰葡糖胺分子加到蛋白质的丝氨酸残基的羟基上。
④对某些蛋白质的N端进行甲基化修饰:这种修饰的蛋白质,如很多细胞支架蛋白和组蛋白等,不易被细胞内的蛋白质水解酶水解,从而使蛋白质在细胞中维持较长的寿命。
⑤酰基化:最常见的一类酰基化修饰是内质网上合成的跨膜蛋白在通过内质网和高尔基体的转运过程中发生的,它由不同的酶来催化,把软脂酸链共价地连接在某些跨膜蛋白的暴露在细胞质基质中的结构域;另一类酰基化修饰发生在诸如src基因和ras基因这类癌基因的产物上,催化这一反应的酶可识别蛋白中的信号序列,将脂肪酸链共价地结合到蛋白质特定的位点上。
如src基因编码的酪氨酸蛋白激酶与豆蔻酸的共价结合。
酰基化与否并不影响酪氨酸蛋白激酶的活性,但只有酰基化的激酶才能转移并靠豆蔻酸链结合到细胞质膜上。
第七章真核细胞内膜系统、蛋白质分选与膜泡运输

根据内质网上是否附有核糖体,分为两类:糙面内 质网(RER)和光面内质网(SER)
RER:呈扁囊状,主要功能是合成分泌性的蛋白和 多种膜蛋白,其膜上有一种称为“易位子”的蛋白 复合体,可能与新合成的多肽进入内质网有关
SER:呈分支管状,是脂质合成的重要场所,往往 也作为出芽的位点,将内质网上合成的蛋白质或脂 质转移到高尔基体内
5、其他功能
解毒功能:肝细胞SER丰富,含有一些酶类,如细胞色素 P450家族酶系,能清除脂溶性的废物和代谢产生的有害物 质
钙储存功能:肌质网,与肌肉收缩有关;另对运输小泡的 形成也有重要的调节作用
为细胞质基质中的很多蛋白提供附着位点
内质网的扁囊和管道可能还有储存与运输物质的功能,在 能量与信号传递、细胞的支持和运动方面可能具有一定的 作用
1、辅基或辅酶与酶的共价结合 2、磷酸化与去磷酸化,用以调节很多蛋白质的生物活性。 3、糖基化。多数为O-连接的糖基化。 在哺乳动物细胞中把N-乙酰葡糖胺分子加到蛋白质的丝 氨酸的羟基上 。 4、蛋白质N端的甲基化修饰,使蛋白质维持较长的寿命。 例如:组蛋白 中间纤维 5、酰基化。 蛋白质与脂肪酸形成脂蛋白。 跨膜蛋白在内质网、高尔基体的转运过程中,暴露在细胞 质基质中的结构域被酰基化。 酶与癌基因的产物特异性的识别,将脂肪酸链共价结合在 特异的蛋白质上。例如:Src、Ras中的脂肪酸链。
降解变性与错误折叠的蛋白和短寿命蛋白质
泛素:是一个由76个氨基酸残基构成的小分子蛋白质。
泛素的功wzf能:依:赖泛素化降解蛋白质途径,在19世纪70年 帮助清理代细~8胞0年中代短由寿美命蛋国白、和以错色误列折三叠位的科蛋学白家质提;出, 参与细胞20周04期年调这控三;位科学家获得诺贝尔化学奖。 DNA修复;细胞的程序性死亡。
第七章真核细胞内膜系统、蛋白质分选与膜泡运输

第七章真核细胞内膜系统、蛋白质分选与膜泡运输第七章真核细胞内膜系统、蛋白质分选和膜泡运输名词解释1、胞内体endosome动物细胞内由膜包围的细胞器,其作用是转运由胞吞作用新摄取的物质到溶酶体被降解。
胞内体被认为是胞吞物质的主要分选站。
2、胞吐作用exocytosis携带有内容物的膜泡与质膜融合,将内容物释放到胞外的过程。
3、胞吞作用endocytosis通过质膜内陷形成膜泡,将细胞外或细胞质膜表面的物质包裹到膜泡内并转运到细胞内(胞饮和吞噬作用)4、糙面内质网rough endoplasmic reticulum,RER附着有核糖体的内质网。
糙面内质网由许多扁平膜囊组成,主要功能包括合成分泌性蛋白质、溶酶体蛋白、膜整合蛋白以及膜脂分子。
5、蛋白质分选protein sorting依靠蛋白质自身信号序列,从蛋白质起始合成部位转运至其功能发挥部位的过程。
蛋白质分选不仅保证了蛋白质的正确定位,也保证了蛋白质的生物学活性。
6、反面高尔基体管网状结构trans Golgi network,TGN处于高尔基体反面的管网状结构,主要功能是负责对蛋白质进行分选,并定向将蛋白质转运到胞内或胞外的最终位置。
7、分泌途径secretory pathway经内质网、高尔基体到细胞表面、溶酶体或植物细胞液泡等的物质分泌路径。
其中涉及分泌物的合成、修饰和分泌物外排等过程。
该过程包括组成型分泌和可调控型分泌。
8、分子伴侣chaperone/molecular chaperone一种与其他多肽或蛋白质结合的蛋白质,以防止蛋白质错误折叠、变性或聚集沉淀,对蛋白质的正确折叠、组装以及跨膜转运有意义。
9、甘露糖-6-磷酸mannose-6-phosphate,M6P溶酶体酶的寡糖链甘露糖残基被磷酸化形成的一个分选标记。
10、高尔基复合体Golgi complex一种由管网结构和多个膜囊组成的极性细胞器,主要功能是对ER 转运来的脂分子及蛋白质进行加工、修饰以及分选。
第七章真核细胞内膜系统、蛋白质分选与膜泡运输

◆扁平囊泡(sacuules)
◆小囊泡(vesicle) ◆大囊泡(vacuoles)
The Golgi Apparatus
培养的上皮细胞中高尔基体的分布 (高尔基体为红色,核为绿色)
高尔基体的形态结构
高尔基体的极性
◆高尔基内侧网络 (cis-Golgi network, CGN) 顺面、形成面 ◆中间潴泡 (medial cisternae) ◆高尔基外侧网络 (trans Golji network,TGN) 外侧面、成熟面
磷脂,磷脂酰胆碱含量较高,鞘磷脂含量较少,没有或很 少含胆固醇。
ER约有30多种膜结合蛋白,另有30多种位于内质网腔, 这些蛋白的分布具有异质性,如:葡糖-6-磷酸酶,普遍存
在于内质网,被认为是标志酶,核糖体结合糖蛋白
(ribophorin)只分布在RER,P450酶系只分布在SER。
内质网的模式图
解毒作用
2+ Ca 离子浓度的调节作用
磷脂的合成
磷脂交换蛋白的作用
解毒作用
◆光面内质网含有丰富的氧化酶系统(如细 胞色素P450、NADH细胞色素C还原酶等)能 使许多有害物质解毒 ,转化为易于排出的 物质。
粗面内质网的功能 --信号肽与蛋白质运输
核糖体循环
?
二、ER的功能(RER)
第二节
细胞内膜系统及其功能
定义:指在结构、功能乃至发生上相互关联、由膜包被的 细胞器或细胞结构, 主要包括内质网、高尔基体、溶酶体、
胞内体和分泌泡等。
功能:区隔化;增加内表面积,提高代谢和调节能力。 从系统发生来看内膜系统起源于质膜的内陷和内共生。 从个体发生来看新细胞的内膜系统来源于原有内膜系统的 分裂,具有核外遗传的特性。
第7章 内膜系统与蛋白质分选和膜运输(2)

1 运输小泡的类型与分选信号
被膜小泡(coated vesicles)类型 ◆披网格蛋白小泡(clathrin-coated vesicle) ◆COPII被膜小泡(COPII coated vesicles) ◆COPI被膜小泡(COPI coated vesicles)概述 ◆概念 ◆细胞分泌活动的过程 ●内质网: ●高尔基体: ●细胞质膜:
分泌活动的类型
◆组成型分泌途径 (Constitutive secretory pathway) ◆调节型分泌途径 (Regulated secretory pathway)
受体介导的内吞作用
◆受体 ◆配体(Ligand) Ⅰ.营养物、 Ⅱ.有害物质 Ⅲ.免疫物质、Ⅳ.信号物质 内吞过程 小窝→内吞泡的形成→初级内体 →次级内体→溶酶体。
内吞过程中受体与配体的命运
◆受体再循环、配体被降解 ◆受体与配体一起再循环 ◆受体与配体都被降解 ◆转胞吞作用
第七节 小泡运输机制
Constitutive and regulated secretory pathway
极性细胞的分泌活动
分泌过程中的蛋白质加工 ◆内质网∶信号肽切除; ◆高尔基体∶ ◆分泌泡: 高尔基体蛋白的驻留
胞吐作用(exocytosis)
◆概念 ◆胞吐作用与分泌活动 ◆胞吐作用的意义 ◆胞吐作用与膜融合
被膜小泡运输途径
◆COPII小泡介导从ER到高尔基体的运输. ◆COPI介导的运输方向是从高尔基体到内质网的回 流. ◆披网格蛋白 ●披网格蛋白介导物质从TGN 到内体,溶酶体,植 物液泡. ●披网格蛋白介导物质从质膜到胞质细胞器.
真核细胞内膜系统蛋白质分选和

真核细胞内膜系统蛋白质分选和
真核细胞内膜系统蛋白质分选和
分泌性蛋白的合成与转运
➢游离核糖体上信号肽的合成
➢SRP识别信号肽,信号识别颗粒(SRP)-核糖体复合体形成,翻译暂停
程主 要 过
➢核糖体与内质网膜结合,形成SRP-SRP受体-核糖体复合体 ➢SRP脱离并参加再循环,核糖体翻译继续进行
➢信号肽被切除,成熟的蛋白质落入内质网腔
动物和真菌细胞内。
真核细胞内膜系统蛋白质分选和
第三节 高尔基复合体 Golgi complex
一、高尔基复合体的形态结构 二、高尔基复合体的化学组成 三、高尔基复合体的功能 四、高尔基复合体的来源
真核细胞内膜系统蛋白质分选和
一、高尔基复合体的形态结构
真核细胞内膜系统蛋白质分选和
一、高尔基复合体的形态结构
主要特点: 细胞质基质是一个高度有序的体系;
通过弱键而相互作用处于动态平衡的结构体系。
真核细胞内膜系统蛋白质分选和
细胞质基质的功能
➢ 完成各种中间代谢过程 ➢ 蛋白质的分选与运输 ➢ 与细胞质骨架相关的功能 ➢ 蛋白质的修饰
真核细胞内膜系统蛋白质分选和
细胞质基质的功能
蛋白质的修饰
(1)辅基或辅基与酶的共价结合 (2)磷酸化与去磷酸化,调节很多蛋白质的生物活性 (3)糖基化。糖基化主要发生在内质网和高尔基体中 (4)对某些蛋白质的N端进行甲基化修饰 (5)酰基化
二、高尔基复合体的化学组成
蛋白质 60% ;脂类 40%
集中在高尔基复合体中的酶,有硫氨素焦磷酸酶和若 干糖基转移酶,也含有酸性磷酸酶和其它溶酶体的酶。 其中糖基转移酶是高尔基复合体最具特征的酶,他能把 低聚糖转移到蛋白质上形成糖蛋白。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第七章内膜系统与蛋白质分选名词:膜结合细胞器:指细胞质中所有具有膜结构的细胞器。
包括细胞核、内质网、高尔基体、溶酶体、分泌泡、线粒体、叶绿体和过氧化物酶体等。
由于它们都是封闭的膜结构,内部都有一定的空间,所以又称为膜结合区室。
通过形成膜结合细胞器,使细胞的功能定位在一定的细胞结构并组成相互协作的系统。
内膜系统: 内膜系统是指内质网、高尔基体、溶酶体和液泡(包括内体和分泌泡)等四类膜结合细胞器, 因为它们的膜是相互流动的,处于动态平衡,在功能上也是相互协同的。
广义上的内膜系统概念也包括线粒体、叶绿体、过氧化物酶体、细胞核等细胞内所有膜结合的细胞器。
小泡运输(膜泡运输):细胞内部内膜系统各个部分之间的物质传递常常通过膜泡运输方式进行。
膜泡运输是一种高度有组织的定向运输,各类运输泡之所能够被准确地运到靶细胞器,主要是因为细胞器的胞质面具有特殊的膜标志蛋白。
许多膜标志蛋白存在于不止一种细胞器,可见不同的膜标志蛋白组合,决定膜的表面识别特征。
胞内膜泡运输沿微管或微丝运行,动力来自马达蛋白内质网:内质网是细胞内的一个精细的膜系统。
是交织分布于细胞质中的膜的管道系统。
两膜间是扁平的腔、囊或池。
内质网分两类,一类是膜上附着核糖体颗粒的叫粗糙型内质网,另一类是膜上光滑的,没有核糖体附在上面,叫光滑型内质网。
粗糙型内质网的功能是合成蛋白质大分子,并把它从细胞输送出去或在细胞内转运到其他部位。
光滑型内质网的功能与糖类和脂类的合成、解毒、同化作用有关,并且还具有运输蛋白质的功能。
溶酶体:溶酶体(lysosomes)真核细胞中的一种细胞器;为单层膜包被的囊状结构,直径约0.025~0.8微米;内含多种水解酶,专司分解各种外源和内源的大分子物质。
高尔基体:是真核细胞中内膜系统的组成之一,它由扁平膜囊(saccules)、大囊泡(vacuoles)、小囊泡(vesicles)三个基本成分组成。
信号斑:信号斑是由几段信号肽形成的一个三维结构的表面, 这几段信号肽聚集在一起形成一个斑点被磷酸转移酶识别。
信号斑是溶酶体酶的特征性信号。
信号识别颗粒:在真核生物细胞质中一种小分子RNA和六种蛋白的复合体,此复合体能识别核糖体上新生肽末端的信号,顺序并与之结合,使肽合成停止,同时它又可和ER膜上的停泊蛋白识别和结合,从而将mRNA上的核糖体,带到膜上。
SRP上有三个结合位点:信号肽识别结合位点,SRP受体蛋白结合位点,翻译暂停结构域。
细胞分泌:动物细胞和植物细胞将在粗面内质网上合成而又非内质网组成部分的蛋白和脂通过小泡运输的方式经过高尔基体的进一步加工和分选运送到细胞内相应结构、细胞质膜以及细胞外的过程称为细胞的分泌。
调节型分泌途径:调节型分泌(regulated secretory pathway)小泡形成的方式可能与溶酶体相似, 分泌蛋白在高尔基体反面网络中通过分选信号与相应的受体结合,使其分选到分泌泡中。
分泌泡比运输溶酶体的运输小泡大, 所含的蛋白质远远多于膜受体的量, 因此有人认为这种分选可能更象细胞表面的受体介导的内吞过程, 有网格蛋白参与。
组成型分泌途径:在这种分泌途径中, 运输小泡持续不断地从高尔基体运送到细胞质膜,并立即进行膜的融合,将分泌小泡中的蛋白质释放到细胞外, 此过程不需要任何信号的触发, 它存在于所有类型的细胞中。
在大多数细胞中, 组成型分泌途径的物质运输不需要分选信号, 从内质网经高尔基体到细胞表面的物质运输是自动地进行的。
组成型分泌途径除了给细胞外提供酶、生长因子和细胞外基质成分外,也为细胞质膜提供膜整合蛋白和膜脂。
异质性细胞器:是指在不同生物及不同发育阶段,该细胞器的形态、大小, 甚至所含有酶的种类都有很大的不同,如溶酶体、过氧化物酶体。
自体吞噬:自体吞噬指细胞将自己细胞质的一部分(如线粒体和内质网)包围起来形成液泡(自体吞噬泡),再依靠初级溶酶体供应的水解酶将其消化。
自体吞噬可由于饥饿和激素的作用等诱导产生。
异体吞噬:细胞吞噬感染的病毒、细菌或其它一些颗粒等称为异体吞噬自溶作用:自溶作用是细胞的自我毁灭, 即溶酶体将酶释放出来将自身细胞降解起始转移信号:蛋白质氨基末端的信号序列除了作为信号被SRP识别外, 还具有起始穿膜转移的作用。
在蛋白质共翻译转运过程中,信号序列的N-端始终朝向内质网的外侧,插入蛋白质转运通道后与通道内的信号序列结合位点(受体)结合,其后的肽序列是以袢环的形式通过运输通道。
不过N-端的起始转移序列是可切除的序列,它的旁边有信号肽酶的作用位点,以N-端信号序列作为起始转移信号的一般都是分泌蛋白。
内部信号序列:内部信号序列又称内部信号肽,它不为于N端,但具有信号序列的作用,故称内部信号序列。
它可以作为蛋白质共翻译转移的信号被SPR识别,同时它也是起始转移信号,可以插入蛋白质转运通道,并与通道中的受体结合,引导其后的多肽序列转运。
终止转移序列:存在于新生肽中使肽链终止转移的一段信号序列,可导致蛋白质锚定在膜的双脂层中。
因终止转移序列的作用而形成单次跨膜的蛋白质,那么该蛋白质在结构上只有一个终止转移序列没有内部转移信号,但在N端有一个信号序列作为起始转移信号。
矽肺/硅肺矽肺:是由于长期吸入石英粉尘所致的以肺部弥漫性纤维化为主的全身性疾病蛋白质分选:主要是指膜结合核糖体上合成的蛋白质,通过信号肽,在翻译的同时进入内质网,然后经过各种加工和修饰,使不同去向的蛋白质带上不同的标记,最后经过高尔基体反面网络进行分选,包装到不同类型的小泡,并运送到目的地,包括内质网、高尔基体、溶酶体、细胞质膜、细胞外和核膜等。
广义的蛋白质分选也包括在游离核糖体上合成的蛋白质的定位。
衔接蛋白:参与披网格蛋白小泡组装的一种蛋白质, 分子量为100kDa, 在披网格蛋白小泡组装中与受体的细胞质结构域相互作用, 起衔接作用。
装配反应因子ARF:一类与被膜小泡装配相关的蛋白因子。
在COP被膜小泡装配中, ARF被认为是外被体外被的装配和去装配的信号。
装配反应因子是一种单体GTPase,由一条多肽链组成,同时含有一个脂肪酸的尾部,属单体GTP结合蛋白。
SNARE 假说:每一种运输小泡都有一个特殊的V-SNARE标志,能够同适当的靶膜上的T-SNARE标志相互作用。
一种运输小泡在没有找到合适的靶位点之前有可能同几种不同的膜位点进行暂时性的接触,这种接触是不稳定的,只有找到真正的靶位点才会形成稳定的机构。
存在于小泡膜上的V-SNAREs是在外被体外被形成时共包装到转运小泡上的,它同靶位点膜上的T-SNAREs蛋白的结合决定了转运小泡的选择性地停靠。
思考题:1.内质网的形态、组成及功能。
内质网膜约占细胞总膜面积的一半,是真核细胞中最多的膜。
内质网是内膜构成的封闭的网状管道系统。
具有高度的多型性。
粗面内质网(RER)呈扁平囊状,排列整齐,膜围成的空间称为ER腔,膜外有核糖体附着。
SER呈分支管状或小泡状,无核糖体附着。
肌肉细胞中的内质网是一种特化的滑面内质网(SER),称为肌质网,可贮存Ca2+,引起肌肉收缩。
SER功能:1、糖原分解释放游离的葡萄糖2、ca2+库3、类固醇激素的合成4、脱毒5、膜脂的合成与转运RER功能:1、蛋白质的合成转运2、合成蛋白的质量监控3、蛋白质的修饰加工处理2.高尔基体的形态、组成及功能。
高尔基体是由数个扁平囊泡堆在一起形成的高度有极性的细胞器。
常分布于内质网与细胞膜之间,呈弓形或半球形,凸出的一面对着内质网称为形成面或顺面。
凹进的一面对着质膜称为成熟面或反面。
顺面和反面都有一些或大或小的运输小泡,在具有极性的细胞中,高尔基体常大量分布于分泌端的细胞质中。
功能:高尔基体的主要功能将内质网合成的蛋白质进行加工、分类、与包装,然后分门别类地送到细胞特定的部位或分泌到细胞外。
3.为什么说高尔基体是一种极性细胞器?结构上的极性:高尔基体可分为几个不同的功能区室。
①靠近内质网的一面是由一些管状囊泡形成的网络结构,通常将这一面称为顺面, 或称形成面。
由于顺面是网络结构,所以又称顺面高尔基网络。
从功能上看,CGM被认为是初级分选站,负责对从ER转运来的蛋白质进行鉴别,决定哪些需要退回,哪些可以进入下一站。
②高尔基体中间膜囊由扁平囊和管道组成,形成不同的区室, 但功能上是连续的、完整的膜体系。
多数糖基修饰、糖脂的形成、以及与高尔基体有关的多糖的合成都发生在中间膜囊中。
③反面高尔基网络, 是高尔基复合体最外面一侧的管状和小泡状物质组成的网络结构,它是高尔基复合体的组成部分,并且是最后的区室。
蛋白质的运输信号在此被特异的受体接受,进行分拣,集中,形成不同的分泌小泡,被运送到不同的地点。
因此, 它的主要功能是参与蛋白质的分类与包装,并输出高尔基体。
某些“晚期”蛋白质的分类与包装也发生在TGN中。
功能上的极性:高尔基体虽然是由膜囊构成的复合体,但是不同的膜囊有不同的功能,执行功能时又是“流水式”操作,上一道工序完成了,才能进行下一道工序,这就是高尔基体的极性。
4.溶酶体的特征及酶蛋白M6P标记是怎样形成的? 如何经M6P分选途径进行分选?①溶酶体真核细胞中的一种细胞器;为单层膜包被的囊状结构,直径约0.025~0.8微米;内含多种水解酶,专司分解各种外源和内源的大分子物质。
②酶蛋白M6P标记是怎样形成:溶酶体的酶在膜旁核糖体上合成,通过信号肽的引导进入粗面内质网,在粗面内质网进行N-连接糖基化。
在此过程中,溶酶体酶蛋白先带上3个葡萄糖、9个甘露糖和2个N-乙酰葡萄糖胺,切除三分子葡萄糖和一分子甘露糖后转运到高尔基体;在高尔基体顺面网络对N连接的糖链进行磷酸化修饰,带上6-磷酸甘露糖的标记③M6P分选途径:具有M6P标记的溶酶体酶在反面高尔基体网络与受体结合后,在网格蛋白帮助下形成具有网格蛋白外被的溶酶体酶分泌小泡, 网格蛋白解聚后的溶酶体酶分泌小泡与一种具有分选作用的细胞器:次级内体融合, 由于次级内体内部的pH≈5.5, 融合后的内体中的pH低于6, 所以与M6P受体结合的溶酶体酶与受体脱离, 释放到内体中;接着,由次级内体中的磷酸酶使溶酶体酶脱磷酸,防止溶酶体酶与M6P受体重新结合。
融合后的次级内体可以通过出芽形成两种类型的小泡, 一种含有溶酶体酶蛋白但不含M6P受体,这种小泡即是成熟的溶酶体。
另一种小泡只含有M6P受体,不含有酶, 它们主要是同反面高尔基体膜融合,偶尔这种小泡也会同质膜融合完成M6P 的再循环。
5.举例说明溶酶体与某些疾病的关系。
①硅肺:二氧化硅尘粒吸入肺泡后被巨噬细内吞噬,含有矽尘的吞噬小体与溶酶体合并成为次级溶酶体。
二氧化硅的羟基与溶酶体膜的磷脂或蛋白形成氢键,导致吞噬细胞溶酶体崩解,细胞本身也被破坏,矽尘释出,后又被其他巨噬细胞吞噬,如此反复进行。