蛋白质的分选和定向运输
第8章蛋白质分选和膜泡运输

蛋白牢固地结合在膜上。 • 当小泡从膜上释放后,衣被很快就解体。
COPII Coated vesicle
三、膜泡运输的定向机制
• 衣被小泡沿着细胞内的微管被运输到靶细胞器,马达蛋白 水解ATP提供运输的动力。
• 各类运输小泡之所以能够被准确地和靶膜融合,是因为运 输小泡表面的标志蛋白能被靶膜上的受体识别,其中涉及 识别过程的两类关键性的蛋白质是SNAREs(soluble NSF attachment protein receptor)和Rabs(targeting GTPase)。 其中SNARE介导运输小泡特异性停泊和融合,Rab的作用 是使运输小泡靠近靶膜。
COP II Sar 1 Sec23/Sec24复合体,Sec 内质网→高尔基体 13/31复合体,Sec 16
(一)笼形蛋白衣被小泡
• 运输途径:质膜→内体;高尔基体→内体;高尔 基体→溶酶体、植物液泡。
• 衣被结构:3重链、3轻链,形如triskelion。clathrin 的曲臂交织在一起,形成5边形网孔的笼子。
• 当衣被小泡形成时,可溶性蛋白dynamin聚集成一 圈围绕在芽的柄部,使柄部的膜尽可能地拉近 (小于1.5nm),导致膜融合,pinch off衣被小泡。
(二)COP I衣被小泡
• 功 能 : 回 收 、 转 运 内 质 网 逃 逸 蛋 白 ( escaped proteins)返回内质网;也可介导高尔基体不同区 域间的蛋白质运输。
谢谢大家!
• Rabs促进和调节运输小泡的停泊和融合。 • Rabs还有许多效应因子,帮助运输小泡聚集和靠
近靶膜,触发SNAREs释放它的效应因子。许多运 输小泡只有在包含了特定的Rabs和SNAREs之后才 能形成。
细胞内蛋白质的分选和转运机制

细胞内蛋白质的分选和转运机制作者:陈建坤来源:《中学生物学》2018年第03期蛋白质作为生命活动的执行者和体现者,与生物的遗传、疾病等都有着重要关联。
在细胞内,有些蛋白质是先合成再进行分选转运,如线粒体、叶绿体、细胞核等结构中的蛋白质;而有些是边合成边分选转运,如分泌蛋白、膜蛋白等。
细胞根据蛋白质是否携有分选信号(信号序列)以及分选信号的性质,有选择地将蛋白质运送到细胞的不同部位。
1分选信号的种类分选信号有两类:①信号肽:蛋白质多肽链上一段连续的特定氨基酸序列,一般位于新肽链的N端,属于一级结构。
完成分选任务后常被切除。
②信号斑:位于多肽链不同部位的几个特定氨基酸序列经折叠后形成的斑块区,是一种三维结构。
完成分选任务后,仍然存在。
2原核细胞中蛋白质分选转运途径原核细胞(如细菌)没有复杂的生物膜系统。
但是为了维持生命,原核细胞需要合成一些蛋白质分泌到细胞质或者转运到细胞外发挥作用。
原核生物中蛋白质的转运分泌途径主要包括3种:①一般分泌途径,即SEC途径;②双精氨酸移位酶途径,即TAT途径;③信号颗粒识别途径,即SRP途径。
除此之外,还有V型分泌途径、TPS分泌途径和分子伴侣引导分泌途径等。
①一般分泌途径,即SEC途径:SEC分泌途径是原核生物中蛋白质主要的跨膜运输机制,主要由SEC移位酶作为介导。
SEC途径可以转运多种蛋白质,包括毒性因子、菌毛、黏附素和蛋白酶等。
SEC途径主要功能是把尚未折叠完成的蛋白质转运到质膜外,在质膜外折叠成有活性的蛋白质。
SEC途径大致可分为3个过程:信号序列的识别与定位、跨膜转运和多肽的释放。
②双精氨酸移位酶途径,即TAT途径:TAT途径识别的肽链N端信号序列通常含有两个连续的精氨酸残基。
TAT途径主要转运已经折叠完成的蛋白质,而尚未折叠完成的蛋白质通常不能通过该系统分泌,从而避免未完成折叠蛋白在胞外被降解的命运,保证了分泌产物的结构和功能的准确性。
此外,TAT途径还可以将少数蛋白质整合到质膜中。
细胞内蛋白质分选转运与细胞结构体系装配要求

共转移(信号肽)
主要是在粗面内质网 上合成的蛋白;
特点:蛋白质边合成 边转移
内质网腔
后转移(post translocation) 1 分子伴侣结合
线粒体,叶绿体等的蛋 胞质中多肽
白;
2 导肽结合线粒体
基本特征:
上的受体
蛋白完全在细胞质基质 中合成,然后再转移到 某些细胞器中。
3 当多肽转移时, 伴随ATP水解,胞 质中的分子伴侣 脱离
②膜泡运输(vesicular transport):蛋白由不同类 型转运小泡从其ER合成部位转运至高尔基体进 而分选运至细胞的不同部位。
③孔门运输(gated transport):细胞质基质中合成 的蛋白质通过核孔复合体到核内或相反。
④细胞质基质中蛋白质的转运:和细胞骨架相关。
二 膜泡运输
普遍存在于真核细胞中,是蛋白运输的一种 特有方式。在转运过程中涉及蛋白本身的修 饰、加工和组装,及不同膜泡定向运输和复 杂的调控过程。 >10种运输小泡参与完成胞内的膜泡运输, 其上有特殊标志,可以保证转运物质到达特 定部位。
蛋白质跨膜转移需
4 线粒体中的分子
ATP使多肽去折叠,及一 伴侣结合转运中的
些蛋白的帮助(如Hsp70) 多肽
使其正确折叠。
5随ATP水解
而释放,多肽在
线粒体中折叠
2.蛋白质分选的四种基本类型:
①跨膜转运(transmembrane transport): 在细胞 质基质中合成的蛋白质转运到内质网、线粒体、 质体(包括叶绿体)和过氧化物酶体等细胞器。 蛋白以非折叠态跨膜。
组成: COPI包被含有8种蛋白亚基, 其依赖ARF(GTP 酶)调节包被的装配与去装配;
功能: ① 负 责 回 收 、 转 运 内 质 网 逃 逸 蛋 白 ( escaped proteins)返回内质网(Golgi →ER,逆行转运); ②行使从Golgi →ER-Golgi中间组分→ ER的物质 运输。 ③在组成性分泌过程中,其在非选择性的批量运 输中行使功能。
细胞生物学题库答案 - 湖北大学

《细胞生物学》题库参考答案第八章 蛋白质的分选一、名词解释1.信号肽--是引导新合成肽链转移到内质网上的一段多肽,位于新合成肽链的N端,一般16~30个氨基酸残基,包括疏水核心区、信号肽的C端和N端等三部分,称信号肽(signal peptide)或信号序列(signal sequence)。
2.共转运--肽链边合成边转移至内质网腔中的方式。
3.后转运--蛋白质在细胞基质中合成以后再转移到某些细胞器中的转移方式。
4.开始转移序列--引导肽链穿过内质网腔的信号肽,称为开始转移序列(start transfer sequence)。
5.停止转移序列--肽链上的一段特殊序列,与内质网膜的亲合力很高,能阻止肽链继续进入内质网腔,使其成为跨膜蛋白质,称为停止转移序列(stop transfer sequence)。
6.分子伴侣--通称为结合蛋白,在细胞中这类蛋白能识别正在合成的多肽或部分折叠的多肽,并与多肽的一定部位相结合,帮助这些多肽的转运、正确折叠或组装,但其本身并不参与最终的形成。
7.导肽--又称转运肽(transit peptide)或导向序列(targeting sequence),它是游离核糖体上合成的蛋白质的N-端信号。
8.NLS序列--亲核蛋白上带有的一段特殊的氨基酸序列,这些特殊短肽保证了亲核蛋白质能通过核孔复合体被转运到细胞核内,这段具有"定向"、"定位"作同的序列被称为核定位信号(nuclear localization signal,NLS)。
9.内体--酸性的不含溶酶体的小囊泡。
10.信号假说--即分泌性蛋白N端序列作为信号肽,指导分泌性蛋白导内质网膜上合成,在蛋白合成结束之前信号肽被切除。
11.蛋白质跨膜运输--主要是指在细胞质基质中合成的蛋白质转运到内质网、线粒体、质体和过氧化物酶体等细胞器,但进入内质网与进入线粒体、叶绿体和过氧化物酶体等细胞器的机制又有不同。
细胞生物学模拟试题答案

细胞生物学模拟试题一、填空题1、细胞周期的调控主要依赖两类蛋白,分别为细胞周期蛋白和细胞周期蛋白依赖的蛋白激酶。
2、染色质 DNA 的三种功能元件自主复制DNA序列、着丝粒DNA序列、端粒DNA序列3、核孔复合体是一种特殊的跨膜运输蛋白复合体,对物质的运输具有双功能性和双向性的特性。
4、生物体内的化学信号分子一般可以分为两类,一是_亲脂性的信号分子 _,一是_亲水性的信号分子_。
5、具有跨膜信号传递功能的受体可以分为__离子通道偶联的受体、 G蛋白偶联的受体和与酶偶联的受体(催化性受体)__。
6、一般将细胞外的信号分子称为__第一信使_,将细胞内最早产生的信号分子称为_第二信使_。
7、受体一般至少包括两个结构域_结构结构域(与配体结合的区域)和催化结构域(产生效应的区域)_。
8、由 G 蛋白介导的信号通路主要包括:_cAMP信号通路和磷脂酰肌醇信号通路。
9、线粒体在超微结构上可分为__内膜、外膜、膜间隙、基质__。
10、线粒体中,氧化和磷酸化密切偶联在一起,但却由两个不同的系统实现的,氧化过程主要由_电子传递链(呼吸链)_ 实现,磷酸化主要由_ ATP合成酶_完成。
11、细胞内膜上的呼吸链主要可以分为两类,__NADH呼吸链和FADH2__呼吸链。
12、植物细胞中具有特异的质体细胞器,主要分为叶绿体、有色体、白色体。
13、叶绿体在显微结构上主要分为_叶绿体膜、基质、类囊体_。
14、在自然界中含量最丰富,并且在光合作用中起重要作用的酶_核酮糖-1 ,5-二磷酸羧化酶_。
15、光合作用的过程主要可分为三步:原初反应、电子传递和光合磷酸化和碳同化。
16、光合作用根据是否需要光可分为__光反应和暗反应__。
17、真核细胞中由双层膜包裹形成的细胞器是线粒体和叶绿体。
18、含有核外 DNA 的细胞器有线粒体和叶绿体。
19、引导蛋白到线粒体中去的具有定向信息的特异氨基酸序列被称为_导肽_。
20、叶绿体中每 3 个 H+穿过叶绿体 ATP 合成酶,生成1个 ATP 分子,线粒体中每2 个 H+ 穿过 ATP 合成酶,生成_1_个 ATP 分子。
蛋白质分选与膜泡运输

第一节 细胞内的蛋白质分选
一、信号假说与蛋白质分选信号
信号序列的发现和证实
1972年Milstein等发现骨髓瘤细胞中 提取的免疫球蛋白分子的N端要比分 泌到细胞外的免疫球蛋白分子N端的 氨基酸序列多出一截。
途径1 表示核基因编码的mRNA在 细胞质基质游离核糖体上完成多 肽链的合成。
途径2 表示合成的蛋白质不含信号 序列,并留在细胞质基质中。
途径3.4.5 分别表示通过跨膜转运 方式转运至线粒体、叶绿体和过 氧化物酶体。
途径6 .表示通过门控转运方式转 运至细胞核。
途径7 表示核基因编码的mRNA 在细胞质基质游离核糖体上起始 合成,然后在信号肽引导下与内 质网膜结合并完成蛋白质合成 (途径8)。
1. COPⅡ有被小泡的组装与运输
功能 – 解折叠后与暴露疏水面结合,防止互作凝聚、错误折叠或被细胞内蛋白酶
水解,对跨膜运送和复合物组装起重要作用。无专一性,需水解ATP获能。
分子伴侣
– 转运发动机 Simon 布朗棘轮模型主要观点: 棘轮原理:把无规则运动过滤成单向动力。 蛋白在转运孔道内,多肽链做布朗运动摇摆不定,一旦前体蛋白进入线粒体 腔,立即有mHsp70(内膜热休克蛋白)高能构象结合导肽,和内膜Tim44 锚定,防止肽链退回细胞基质中。随着肽链的进一步延伸,有更多的 mHsp70结合。 mHsp70转变为松弛低能构象促使导肽进入内膜。
强的亲合力从而使之结合在脂双层中,这段序列不再转入 内质网腔中。 开始转移序列和终止转移序列的数目决定多肽跨膜次数。
三、线粒体和叶绿体蛋白质的转运
(一)线粒体蛋白质的转运因子
蛋白质的分选
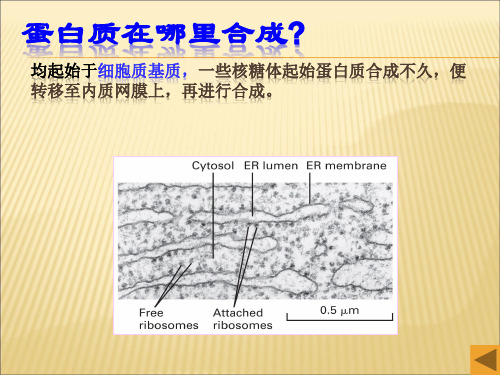
4.细胞质基质起始和粗内质网合成的蛋白质主要包括:
(1)胞外分泌蛋白 常以分泌泡的形式通过细胞的外排作用输送到细胞外。 (2)膜整合蛋白 细胞质膜、内质网、高尔基体和溶酶体膜上的膜蛋白等都具有 方向性,其方向性在内质网上合成时就已确定。 (3)构成细胞器的可溶性驻留蛋白 有些驻留蛋白需要与其它细胞组分严格隔离,如溶酶体与植物 液泡中的酸性水解酶类;内质网、高尔基体和胞内体 (endosome)中固有的蛋白以及其它有重要生物活性的蛋白。
共翻译转运(Cotranslational translocation) 蛋白质类型:
(1)胞外分泌蛋白:分泌泡形式胞吐到胞外。 (2)膜整合蛋白:细胞质膜、内质网、高尔基体和溶酶 体的膜蛋白等,具有方向性。 (3)内膜系统细胞器内的可溶性驻留蛋白:需要与其它 细胞组分严格隔离,如溶酶体与植物液泡中酸性水解酶等。
1.蛋白质分选的两条基本途径:
共翻译转运(Cotranslational translocation),在细 胞质基质,蛋白质合成起始后,转移至粗面内质网,至高 尔基体,溶酶体,分泌泡,细胞膜乃至胞外。
翻译后转运(post-translational translocation), 在细胞质基质中完成多肽链的合成,然后转运至细胞质基 质特定部位或膜围绕的细胞器。包括线粒体、叶绿体、过 氧化物酶体、内质网及细胞核,细胞质基质驻留蛋白、骨 架蛋白等。
膜泡运输为细胞内膜系统进行物质转运的主要方式, 定向运输取决于膜和膜泡(至少十数种)的表面识别特征。
转运膜泡类型(按功能分类): 1.COPII有被小泡 2.COPI有被小泡 3.网格蛋白有被小泡 内膜系统细胞器保持其特异性蛋白组成的两种机制 有被小泡与靶膜的融合:选择性融合 内膜系统各结构保持形态与膜成分相对稳定的方法
细胞学第八章答案

《细胞生物学》题库参考答案第八章蛋白质的分选一、名词解释1.信号肽——是引导新合成肽链转移到内质网上的一段多肽,位于新合成肽链的N端,一般16~30个氨基酸残基,包括疏水核心区、信号肽的C端和N端等三部分,称信号肽(signal peptide)或信号序列(signal sequence)。
2.共转运——肽链边合成边转移至内质网腔中的方式。
3.后转运——蛋白质在细胞基质中合成以后再转移到某些细胞器中的转移方式。
4.开始转移序列——引导肽链穿过内质网腔的信号肽,称为开始转移序列(start transfer sequence)。
5.停止转移序列——肽链上的一段特殊序列,与内质网膜的亲合力很高,能阻止肽链继续进入内质网腔,使其成为跨膜蛋白质,称为停止转移序列(stop transfer sequence)。
6.分子伴侣——通称为结合蛋白,在细胞中这类蛋白能识别正在合成的多肽或部分折叠的多肽,并与多肽的一定部位相结合,帮助这些多肽的转运、正确折叠或组装,但其本身并不参与最终的形成。
7.导肽——又称转运肽(transit peptide)或导向序列(targeting sequence),它是游离核糖体上合成的蛋白质的N-端信号。
8.N LS序列——亲核蛋白上带有的一段特殊的氨基酸序列,这些特殊短肽保证了亲核蛋白质能通过核孔复合体被转运到细胞核内,这段具有“定向”、“定位”作同的序列被称为核定位信号(nuclear localization signal,NLS)。
9.内体——酸性的不含溶酶体的小囊泡。
10.信号假说——即分泌性蛋白N端序列作为信号肽,指导分泌性蛋白导内质网膜上合成,在蛋白合成结束之前信号肽被切除。
11.蛋白质跨膜运输——主要是指在细胞质基质中合成的蛋白质转运到内质网、线粒体、质体和过氧化物酶体等细胞器,但进入内质网与进入线粒体、叶绿体和过氧化物酶体等细胞器的机制又有不同。
12.膜泡运输——蛋白质通过不同类型的转运小泡从其糙面内质网合成部位转运至高尔基体进而分选运至细胞的不同部位,其中涉及各种不同的运输小泡的定向转运,以及膜泡出芽与融合的过程。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
蛋白质分选的基本途径与类型
• 蛋白质的分选大体可分为两条途径:
1、在细胞质中完成多肽链的合成,然后转运 至膜围绕的细胞器,如线粒体、过氧化物 酶体、细胞核及细胞质基质的特定部位, 最近发现有些还可转运至内质网中。
2、蛋白质合成起始后转移至糙面内质网,新 生肽边合成边转入糙面内质网腔中,随后 经高尔基体运至溶酶体、细胞膜或分泌到 细胞外,内质网与高尔基体本身的蛋白成 分的分选也是通过这一途径完成的。
蛋白质的分选和运输
• 除线粒体和植物叶绿体中能合成少量蛋白
质外,绝大多数的蛋白质均在细胞质基质 中的核糖体上开始合成然后运至细胞的特 定部位。这已过程称蛋白质的定向转运或 分选。
信号假说与蛋白质分选信号
• 1975年,G.Blobel和D.Sabatini等提出了信
号假说,即分泌蛋白N端序列作为信号肽, 知道分泌蛋白到内质网上合成,在蛋白合 成结束之前信号肽被切除。现已确认,知 道分泌性蛋白在糙面内质网上合成的 决定 因素是蛋白质N端的信号肽。
• 有证据表明,COP I有被小泡行使高尔基体
到内质网逆行转运外,也可行使从内质网 到高尔基体的顺行转运,以至通过高尔基 体的不同分区。
在细胞的膜泡运输中,糙面内质网相当于 重要的物质供应站,而高尔基体是重要集 散中心。由于内质网的驻留蛋白具有回收 信号,即使有的蛋白发生逃逸,也会保留 或回收回来,所以有人将内质网比喻成 “开放的监狱” 。
• 3、选择性的门控转运
指在细胞质基质中合成的蛋白质通过核孔 复合体选择性地完成核输入或从细胞核返 回细胞质。
• 4、细胞质基质中的蛋白质的转运
上述几种分选类型均涉及蛋白质在细胞基 质中的转运,这一过程显然与细胞骨架系 统密切相关。当由于细胞质基质的结构尚 不清楚,因此蛋白质转运特别是信号转导 途径中的蛋白分子的转运及信号传递方式 了解很少。
膜泡运输
• 膜泡运输是蛋白运输的一种特有的方式,
普遍存在于真核细胞中。在转运过程中不 仅涉及蛋白本身的修饰、加工和装配,还 涉及导多种不同膜泡定向运输及其复杂的 调控过程。
• 完成细胞内的膜泡运输至少需要10种以上
的运输小泡,目前发现三种不同类型的有 被小泡具有运输不同的物质运输作用。
• 1、网络蛋白有被小泡
• 如果从蛋白质分选的类型或机制的角度看,
可分为四种基本类型:
1、蛋白质的跨膜转运
主要是指在细胞质基质中合成的蛋白质转 运到内质网、线粒体、质体和过氧化物酶 体等细胞器,当进入内质网与进入线粒体、 叶绿体和过氧化物酶体等细胞器的机制又 有不同。
• 2、膜泡运输
蛋白质通过不同类型的转运小泡从其糙面内 质网合成部位转运至高尔基体进而分选运 至细胞的不同部位,其中涉及各种不同的 运输小泡的定向转运,以及膜泡出芽与融 合的过程。
负责蛋白质从高尔基体TNT向质膜、胞内体 或溶酶体和植物液泡运输。另外,在受体 介导的细胞内吞吐途径中也负责将物质从 质膜运往细胞质,以及从胞内体到溶酶体 的运输。
• 2、COP II有被小泡
负责从内质网到高尔基体的物质运输。
COP II包被蛋白由5种蛋白亚基组成,这些 亚基首先在酵母细胞突变体中被鉴定,这 些突变体不能完成从内质网到高尔基体的 物质转运。COP II有被小泡具有选择性的确定,一是因为COP II蛋白能识别
并结合跨膜内质网蛋白胞质面一端的信号 序列,二是跨膜内质网蛋白的腔面一端作 为受体ER腔中的可溶性蛋白结合。
• 3、 COP I有被小泡
负责收回、转运内质网逃逸蛋白返回内质网。
早期研究表明,在糙面内质网中合成的蛋白质, 除了某些有特殊标志的蛋白驻留在内质网或高尔 基体或选择性地进入溶酶体和调节性分泌泡外, 其余的蛋白主要是通过一种称为default pathway 方式来完成蛋白质的转运过程。在这种组成型分 泌过程中,COP I有被小泡在非选择性的批量运输 中行使功能。