基因重组方法=全
基因重组知识点总结

基因重组知识点总结一、基因重组的原理基因重组的原理是在DNA分子水平上,通过切割和重组DNA的不同片段,形成新的DNA 序列。
基因重组可以实现DNA片段的互换、合并、删除或插入操作,从而改变DNA的序列,并且产生新的基因组合。
基因重组的原理主要涉及到DNA的结构、酶的作用和DNA片段的互补配对等方面。
1. DNA的结构DNA是由四种碱基(腺嘌呤A、胞嘧啶T、鸟嘌呤G、胞嘧啶C)组成的双链分子,它的结构在空间上呈现出双螺旋的形态。
每一条DNA链都由磷酸和脱氧核糖组成,而这些单元组成了DNA的主干。
而碱基对(A-T、G-C)则连接了两条DNA链,形成了DNA的双链结构。
2. 酶的作用在基因重组的过程中,酶起着至关重要的作用。
例如,核酸酶能够切割DNA分子,使得DNA的特定区域被切割成不同的碱基序列;而连接酶则能够将不同的DNA片段连接起来,形成新的DNA序列。
此外,一些重组酶还可以通过其催化作用来促进DNA分子的重组。
这些酶的作用在基因重组的过程中起着关键的作用。
3. DNA片段的互补配对在DNA重组的过程中,DNA分子的互补配对起着非常重要的作用。
DNA的双链结构使得其具有互补配对的性质,即A会与T形成氢键,而G则会与C形成氢键。
这种互补配对性质使得DNA片段能够通过互补配对的方式进行连接或重组。
综上所述,基因重组的原理涉及到DNA的结构、酶的作用和DNA片段的互补配对等方面。
通过这些原理,我们可以实现DNA分子中某一段DNA片段的与同一DNA分子或不同DNA分子中的另一段DNA片段重新组合成新的DNA序列。
二、基因重组的方法基因重组的方法主要包括DNA重组、基因克隆、基因组编辑和CRISPR-Cas9等。
这些方法可以分别用于不同的应用领域,并且在现代生物技术中有着重要的价值。
1. DNA重组DNA重组是指通过DNA片段的切割和重组来形成新的DNA序列。
这一方法主要依赖于核酸酶的切割作用和连接酶的连接作用。
必修2-基因重组
酶Ⅱ
瓜氨酸
酶Ⅲ
精氨酸
⑴在一次实验中,经测定只发现鸟氨酸和瓜氨酸, 而没有精氨酸。产生此现象的可能原因是 ( 基因Ⅲ发生突变,酶Ⅲ缺乏 ) 致使( 不能形成精氨酸 )
⑵如果酶Ⅰ和酶Ⅲ正常,酶Ⅱ活性丧失则能否合成 鸟氨酸和精氨酸? ( 能合成鸟氨酸不能合成精氨酸 ) 原因是( 酶Ⅱ失活,鸟氨酸不能合成瓜氨酸,因 此精氨酸的合成缺乏原料。 )
发生 时间 及原 因 条件
意义
发生 可能
思维拓展 不同生物的可遗传变异来源: 病毒—— 基因突变 原核生物—— 基因突变
真核生物—— 基因突变、基因重组、 染色体变异
课堂练习
1. 下列现象中属于可遗传的变异的是( A、玉米由于水肥充足而长得穗大粒足 B、人由于晒太阳皮肤变黑 C、无籽番茄没有种子 D、人类的色盲病
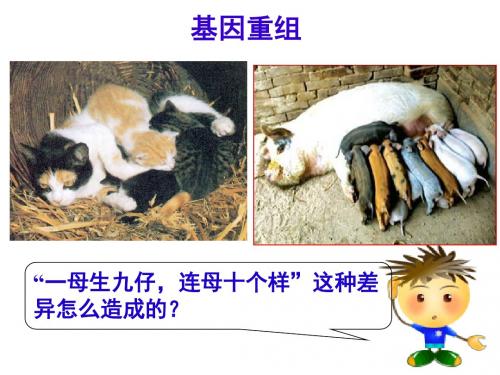
基Байду номын сангаас重组
“一母生九仔,连母十个样”这种差 异怎么造成的?
(一)基因重组的概念
在生物体进行有性生殖的过程中,控 制不同性状的非等位基因的重新组合。
(二)基因重组的方式 1、同源染色体的非姐妹染色单体之间的交 叉互换(交叉互换型重组) 2、非同源染色体上的非等位基因自由组合 (自由组合型重组) 3、人工基因重组(基因工程)
基因重组
不同基因的重新组合。
不产生新基因,而是产生新的基因型, 使不同性状重新组合。 减数第一次分裂前期的四分体时期 的交叉互换; 减数第一次分裂后期,非同源染色 体上的非等位基因自由组合。 有性生殖过程中进行减数分裂形成 生殖细胞。 是生物变异的重要来源,是形成生物 多样性的重要原因,对生物的进化也 具有重要的意义 有性生殖中非常普遍
(三)基因重组发生时期 减数第一次分裂前期(四分体时期)同源染 色体上的非姐妹染色单体之间的交叉互换
高中生物 基因重组(gene

能在选择性培养
基平板上形成微 小菌落就是流产 转导的特点。
2. 局限转导(specialized transduction)
定义:通过部分缺陷的温和噬菌体把供体菌的少数特定基 因携带到受体菌中,并与后者的基因组整合、重组,形成 转导子的现象。
特点:
只能转导供体菌的个别特定基因(一般为噬菌体 整合位点两侧的基因);
就同时获得复制的机会。所以在双重溶源菌中的正常λ 噬菌
体被称为助体(或辅助)噬菌体(helper phage)。 双重溶源菌的裂解物中含有等量的λ和λdgal粒子,称为
HFT(高频转导)裂解物。
高频转导和低频转导图解
E. coli K12 ()gal+(供体菌)
U.V.
(大量) + dgal+(10-5) E.coliK12Sgal-/ dgal+
结合在寄主染色体特定位置 上
紫外线诱导溶源菌 一般不稳定,呈缺陷溶原性 (对同源噬菌体具有免疫性, 但不表现出其它噬菌体的性 状)
转导子的区 别
3. 溶源转变
概念:当温和噬菌体感染宿主而使其发生溶源化时,因噬菌 体的基因整合到宿主的核基因组上,而使后者获得了除免疫 性以外的新性状的现象,称为溶源转变。 性质:表面上与转导相似,而本质上不同于转导。 区别:
低频转导
E. coli K12Sgal-(受体菌)
E.coliK12Sgal-/ dgal+/
(双重溶源转导子)
E. coli K12Sgal-/ dgal+/ (双重溶源菌供体)
U.V.
(50%) + dgal+(50%)
E. coli K12Sgal-(受体菌)
基因重组方式

基因重组方式
嘿,大家知道吗,基因可是个超级神奇的东西!今天咱就来聊聊基因重组方式。
基因重组就像是一场基因的大变身游戏。
有一种常见的方式叫减数分裂中的重组。
就好比搭积木,染色体在减数分裂时重新组合,产生新的基因组合。
这就像抽奖一样,每次都可能有不同的结果,是不是很有意思?
还有一种呢,是基因工程带来的重组。
这就像是我们人类当导演,把不同的基因片段拼接到一起,让生物拥有我们想要的特性。
比如说,我们可以把能产生某种有用蛋白质的基因放到细菌里,让细菌帮我们大量生产,这多厉害呀!
再说说同源重组,这就像是基因世界里的“交换生”活动。
相似的基因片段会相互交换位置,从而产生新的基因组合。
想象一下,两个差不多的拼图块换了一下位置,整个画面就不一样了,基因也是这样哦。
转座子重组呢,就像是基因里的“小调皮”。
这些转座子可以在基因里跳来跳去,改变基因的结构和功能。
这不就像一个调皮的孩子在房间里跑来跑去,把东西都弄乱了,但有时候也会带来一些意想不到的变化呀。
基因重组的意义可太大啦!它让生物有了更多的多样性,让世界变得丰富多彩。
没有基因重组,生物可能就一直是老样子,哪来这么多千奇百怪、有趣好玩的生物呢?
基因重组方式真的超级神奇,它让生命充满了无限可能。
我们对基因重组的研究和利用也会越来越深入,未来肯定会有更多让人惊叹的发现和应用!。
基因重组方法=全PPT课件

.
23
外源DNA被降解,转导失败。
(2)局限性转导(specialized transduction)
温和噬菌体感染
整合到细菌染色体的特定位点上
宿主细胞发生溶源化
溶源菌因诱导而发生裂解时, 在前噬菌体二侧的少数宿主 基因因偶尔发生的不正常切 割而连在噬菌体DNA上
部分缺陷的温和噬菌体
把供体菌的少数特定基因转移到受. 体菌中
的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
进行自然转化,需要二方面必要的条件:
建立了感受态的受体细胞
外源. 游离DNA分子
30
枯草芽孢杆菌的自然转化过程(革兰氏阳性菌的转化模型)
分泌感受态因子
与细胞表面受 体M相互作用
使细胞表面的 DNA结合蛋白 及核酸酶裸露出 来,使其具有与 DNA结合的活 性
b)决定因素也各有不同;
.
34
(2)人工转化
在自然转化的基础上发展和建立的一项细菌基因重组手段, 是基因工程的奠基石和基础技术。
不是由细菌自身的基因所控制;
用多种不同的技术处理受体细胞,使其人为地处于一 种可以摄取外源DNA的“人工感受态”。
用CaCl2处理细胞,电穿孔等是常用的人工转化手段。
质粒的转化效率高;
含有F因子的细胞:“雄性”菌株(F+),其细胞表面有性菌毛 不含F因子的细胞:“雌性”菌株. (F-),细胞表面没有性菌毛7
F因子为附加体质粒 既可以脱离染色体在细胞内独立存. 在,也可插入(整合)到染色8 体上
F因子的四种细胞形式
a)F-菌株, 不含F因子,没有性菌毛,但可以通过 接合作用接收 F因子而变成雄性菌株(F+);
.
《基因重组》 讲义

《基因重组》讲义一、什么是基因重组基因重组是指在生物体进行有性生殖的过程中,控制不同性状的基因重新组合。
它是生物遗传变异的一种重要来源,为生物的进化提供了丰富的材料。
简单来说,基因重组就像是一场基因的“重新洗牌”。
在这个过程中,原本存在于不同个体中的基因组合在一起,产生了新的基因组合,从而导致后代的性状可能与亲代有所不同。
基因重组主要发生在减数分裂过程中。
在减数第一次分裂前期,同源染色体的非姐妹染色单体之间会发生交叉互换;在减数第一次分裂后期,非同源染色体自由组合,这两种情况都会导致基因重组。
二、基因重组的发生机制1、减数分裂前期的交叉互换在减数第一次分裂前期,同源染色体两两配对形成四分体。
此时,同源染色体的非姐妹染色单体之间可能会发生片段的交换。
这就好比两条绳子在某些地方相互缠绕,然后交换了一小段。
这种交换使得原本在不同染色体上的基因有了重新组合的机会。
例如,假设一条染色体上有控制眼睛颜色的基因 A 和控制头发颜色的基因 B,另一条同源染色体上相应位置的基因是 a 和 b。
经过交叉互换,可能会产生新的组合,如 A 和 b 或者 a 和 B 在同一条染色体上。
2、减数分裂后期的非同源染色体自由组合在减数第一次分裂后期,同源染色体分离的同时,非同源染色体自由组合。
这意味着来自父方和母方的染色体可以以各种不同的方式组合在一起。
比如说,一个生物体细胞中有两对染色体,一对是 A 和 a,另一对是 B 和 b。
在减数分裂时,A 可能与 B 组合进入一个配子,也可能与 b 组合;同样,a 也可能与 B 或者 b 组合。
这样就产生了多种不同的基因组合。
三、基因重组的意义1、增加生物的多样性基因重组使得生物能够产生丰富多样的基因组合,从而增加了生物的遗传多样性。
这种多样性为生物适应不断变化的环境提供了更多的可能性。
例如,在一个多变的环境中,某些基因组合可能使个体更能适应寒冷的气候,而另一些组合可能更适合炎热的环境。
微生物基因重组的方式

微生物基因重组的方式
一、转化
嘿,同学们!咱们先来说说转化这一微生物基因重组的方式。
转化就像是微生物界的“知识传递”。
一些细菌能从周围环境中摄取游离的 DNA 片段,然后把这些外来的基因整合到自己的基因组里,实现基因重组。
这就好像是一个“学霸”把自己的学习心得分享给“学渣”,“学渣”吸收了之后成绩突飞猛进一样。
比如说肺炎双球菌的转化实验,大家还记得不?是不是很神奇呀!
二、转导
聊聊转导。
转导就像是微生物世界里的“快递小哥”。
病毒在感染细菌时,能把细菌的部分基因包裹在自己的蛋白质外壳里,然后当它去感染另一个细菌时,就把这些基因“送”了过去。
这就好比你给朋友寄了个包裹,里面装的是珍贵的“宝贝”——基因。
这是不是很有趣呢?
三、接合
最后说说接合。
这个方式就像是微生物之间的“牵手合作”。
通过细胞与细胞的直接接触,遗传物质就能从一个细胞转移到另一个细胞啦。
比如说大肠杆菌,它们就经常通过这种方式来交换基因。
感觉就像是小伙伴之间互相分享自己的小秘密,然后大家都变得更厉害了!
微生物基因重组的方式真是太奇妙啦,让微生物们有了更多的可能性去适应环境,不断进化。
咱们得好好研究,说不定能发现更多好玩的东西呢!。
基因重组技术的研究方法

基因重组技术的研究方法基因重组技术是当前生物学研究领域中的一个热门话题,其涉及到基因的组合、修饰、重构,以及基因对生物外在表现的调控作用等方面,是探究生物学各个层次的目标之一。
在这篇文章中,我们将探讨基因重组技术的研究方法,以及其在生物学研究中的应用。
一、PCR 技术PCR 技术是从一个 DNA 分子扩增出大量同一序列的方法。
通过PCR 技术,可以提取出生物细胞中任何一个基因序列,或是大量同源序列的 DNA 片段,然后创造无数相同的序列。
PCR 技术的应用已经成为基因重组的基础方法之一,对于寻找正确的基因片段和重组序列来说非常重要。
二、限制酶切割技术限制酶切割技术是一个用于分割 DNA 片段的方法,因为当限制酶与 DNA 结合后,酶会消耗 DNA 片段中特定的碱基序列,并将剩余的 DNA 片段切成一定长度的片段。
对于基因重组,这个技术的应用非常必要。
这个技术的使用使科学家能够使用这些片段创建复杂的彩虹 DNA,将各种来源的 DNA 成分摆放在一起。
三、酶联免疫吸附技术 (ELISA)酶联免疫吸附技术识别在基因重组技术中的特定抗原。
ELISA 技术的优点是能够快速、容易地测量特定抗原结构的存在和分布,对于诊断和治疗方面有很重要的应用。
在基因重组方面,这种技术可以寻找与特定基因相关的蛋白。
此外,ELISA 还可以用于研究如何使重组的基因表达出来,并在此基础上管理重组基因的活动。
四、过滤技术通过过滤技术,可以通过分离液体和固体来提取同源 DNA 片段。
随着过滤技术的不断改进,现在已经可以通过过滤技术来排除可能导致假阳性结果的多个过程。
在基因重组方面,这个技术使得若干个来源的DNA 片段可以很容易地组合在一起,形成一份新的 DNA 序列。
五、电泳技术电泳技术是一种控制 DNA 片段移动的方法。
将 DNA 片段放置在凝胶状的物质中,然后在电场中提供能量,这样 DNA 片段就会根据大小和电荷移动。
电泳技术是将 DNA 片段进行分离和鉴定的必不可少的工具之一。
人工基因重组育种的方法

人工基因重组育种的方法一、同源重组育种方法同源重组育种方法是一种基于同源序列的基因重组技术。
在同源重组过程中,两个DNA片段具有同源序列,通过交换和重组,产生新的DNA结构。
这种方法被广泛应用于人工基因重组育种。
1.1 同源重组原理同源重组的基本原理是利用两个DNA片段之间的同源序列,通过交换和重组,实现基因的重新组合。
在同源重组过程中,DNA片段之间的同源序列形成联合,并交换对应位置的基因,最终产生新的DNA结构。
1.2 同源重组技术应用同源重组技术在基因功能研究和遗传工程领域得到广泛应用。
通过同源重组技术,可以构建基因敲除、基因敲入、基因敲减等基因编辑载体,实现特定基因的调控和表达。
此外,同源重组技术还可用于构建转基因植物和动物,提高作物产量、抗病性和抗虫性等。
1.3 同源重组优缺点同源重组技术的优点包括高效性、准确性、特异性等。
同源重组技术可以准确地定位到目标基因并进行编辑,减少了基因突变和突变的自发率。
然而,同源重组技术的缺点是操作复杂、成本高昂,且在某些情况下可能受到细胞内同源重组酶活性的限制。
二、非同源末端连接育种方法非同源末端连接育种方法是一种基于非同源末端序列的基因重组技术。
在非同源末端连接过程中,两个DNA片段的非同源末端序列通过连接酶的作用实现连接和重组。
2.1 非同源末端连接原理非同源末端连接的基本原理是利用两个DNA片段的非同源末端序列,通过连接酶的作用形成新的DNA结构。
在非同源末端连接过程中,连接酶识别并催化两个DNA片段的非同源末端序列之间的连接反应,形成新的DNA双链结构。
2.2 非同源末端连接技术应用非同源末端连接技术在基因敲除、基因敲入和基因敲减等领域得到广泛应用。
通过非同源末端连接技术,可以构建各种类型的基因编辑载体,实现特定基因的调控和表达。
此外,非同源末端连接技术还可用于构建转基因植物和动物,提高作物产量、抗病性和抗虫性等。
2.3 非同源末端连接优缺点非同源末端连接技术的优点包括高效性、广谱性和可操作性等。
原核生物基因重组的四种方式

原核生物基因重组的四种方式
原核生物基因重组的四种方式包括转化、转导、接合和原生质体融合。
转化是指受体菌直接吸收来自供体菌的DNA片段,通过交换整合到自己的基因组中,经复制使自己变成一个转化子。
转导是以完全缺陷或部分缺陷的噬菌体为媒介,把供体细胞的DNA片段携带到受体细胞中,通过交换与整合使后者获得前者部分遗传性状的现象。
接合是指两个细菌通过直接接触形成基因转移的桥梁,通过交换与整合使受体菌获得供体菌部分遗传性状的现象。
原生质体融合是指将两种细菌的原生质体融合在一起,通过交换与整合使融合后的细胞获得两种细菌的遗传性状的现象。
基因重组技术制备重组蛋白

基因重组技术制备重组蛋白在生物学领域中,基因重组技术是一项非常重要的技术,它可以帮助科学家们将不同的基因组合起来制造出人工合成的蛋白质,这种制造方法叫做基因重组蛋白制造法。
基因重组蛋白的制备方法可以应用在医学领域中,例如生产药品、生产诊断试剂、制造疫苗等。
一、基因重组技术的基本原理基因重组技术就是将不同的基因从不同的实体中提取出来,并将这些基因重新组合,生成一个新的DNA序列,将这个新的DNA序列植入到另一个细胞中。
其中的难点就在于如何将新的DNA序列精确插入到另一个细胞中。
基因重组制备重组蛋白的过程,分为三个主要的步骤。
第一步是提取目标基因。
科学家们可以通过转录RNA和反转录DNA的技术,从DNA中提取出目标基因,这是制备重组蛋白的第一步。
第二步是将目标基因植入到表达载体中。
表达载体是一个能够承载新的DNA序列并将其表达出来的细胞。
将目标基因植入到表达载体中,就是为了让这个基因被表达出来,并在表达的基础上生产出重组蛋白。
在最后一步,科学家们将表达载体与宿主细胞并列在一起,让载体表达新的基因,这样新的蛋白质就会被制造出来。
二、制备重组蛋白的应用制备基因重组蛋白的应用非常广泛。
其中最常见的就是用基因重组技术来生产药品。
重组的蛋白质能够用于制造多种种类的药物,例如一些生长因子,以及用于治疗某些癌症的药物等。
除此之外,重组的蛋白质还能够用于生产诊断试剂。
目前,基于重组蛋白制备出的诊断试剂使用非常广泛,可以用于诊断多种疾病,并且能够改善疾病的治疗效果。
还有一种重要的应用就是基于重组蛋白制造出有效的疫苗。
这种制备方式被广泛应用于制造疫苗的过程中,已经成为疾病预防的重要工具。
例如,新冠病毒疫苗和流感疫苗是基于基因重组技术制备出来的一种疫苗。
这些疫苗制造方式不仅安全可靠,同时也能够快速发展,用于抗击突然出现的疫情。
三、基因重组蛋白的优缺点通过基因重组技术制备蛋白对人类的医学予以了不可估量的贡献。
由于这种方法限制较少,且制备的蛋白质有较高的生物活性,并且往往具有高效的药理学效应,因此能够更准确,也更安全的治疗人体内的疾病。
基因工程中的基因重组方法及实验步骤探讨

基因工程中的基因重组方法及实验步骤探讨基因工程是一项重要的生物技术,它涉及到对生物体的基因进行操作和编辑,以改变其遗传特征。
基因重组是基因工程中常用的一种方法,它通过重新组合DNA分子,创造出具有特定功能的基因。
基因重组的方法有多种,包括限制性内切酶法、聚合酶链式反应法、DNA连接酶法等。
下面我们将对这些方法进行探讨,并介绍相应的实验步骤。
限制性内切酶法是基因重组中最常用的方法之一。
限制性内切酶是一类能够识别DNA序列并切割DNA链的酶。
在基因重组中,首先需要选择合适的限制性内切酶,根据目标基因或DNA片段的序列,确定限制性内切酶的切割位点。
然后,将目标基因和载体DNA分别用相同的限制性内切酶切割,生成切割的DNA片段。
接下来,将目标基因和载体DNA的切割片段进行混合,然后使用DNA连接酶将它们连接在一起。
最后,将连接好的DNA转化到宿主细胞中,使其表达目标基因。
聚合酶链式反应(PCR)法是另一种常用的基因重组方法。
PCR是一种在体外扩增DNA片段的技术。
在基因重组中,首先需要设计引物,引物是一对短的DNA序列,它们能够与目标DNA片段的两个末端互补结合。
引物结合到目标DNA片段后,通过PCR反应体系中的DNA聚合酶,通过不断的酶切和合成反应,使得目标DNA片段得以扩增。
PCR反应通常具有循环反应的性质,每个循环包括DNA的解性、引物结合、DNA合成和DNA的再解性。
通过多个循环的反复操作,可以在较短的时间内扩增出大量目标DNA片段。
DNA连接酶法是基因重组中用于连接两个DNA片段的方法。
DNA连接酶是一种酶类,它能够识别DNA的末端序列,并在双链DNA末端之间形成一个磷酸二酯键。
在基因重组中,首先需要将待连接的DNA片段分别用限制性内切酶切割,并生成具有互补末端序列的DNA片段。
然后,将这些片段与DNA连接酶一起反应,使它们连接成一个完整的DNA分子。
连接反应通常在适当的温度和时间下进行。
最后,通过转化技术将连接好的DNA分子导入宿主细胞中,使其表达目标基因。
基因重组的方式

基因重组的方式随着基因科学的不断发展,基因重组技术取得了巨大的进步,现在它已成为研究基因的最重要的手段之一。
在生物学上,基因重组是指通过将不同的基因片段以某种方式组合在一起,在原有基因序列和构建新基因序列之间建立关系,来改变某个基因组的基因组成。
它可以帮助研究者探索基因组织结构、功能和进化过程。
基因重组技术可以采用多种方式,包括光子化学剪接、质粒子联合和质粒克隆等。
光子化学剪接可以通过使用光子来分割特定基因,并将其与另一个基因片段结合在一起,来获得新的基因结构。
质粒子联合是一种低等离子体聚合的方式,可以将两个不同的基因片段结合在一起,从而生成一个新的基因。
而质粒克隆技术则是一种从基因组中分离出特定基因片段的技术,允许研究者有效地利用质粒片段来分析基因表达。
基因重组技术在许多研究领域内都发挥着重要作用,例如,它可以帮助研究者更好地了解基因组的构成,进而帮助确定基因的功能,探究基因的结构和进化,以及修饰基因的功能。
此外,基因重组技术还能用于研究非编码RNA的表达、改变植物和动物基因特性以及进行药物研发等。
基因重组技术也开放了各种新的技术,例如,基因工程、遗传改良和生物技术等,这些技术可以克服生物学上的一些问题,并对传统的动物和植物育种技术有很大的推动作用。
此外,基因重组技术也有助于推进分子治疗的发展,同时可以提高人类的健康水平。
基因重组技术是一项强大的技术,为科学家们研究基因提供了全新的可能性,并且它可以用于各种不同的用途,如:生物学、育种和分子治疗等。
但不可否认的是,基因重组技术也存在一定的风险,因此,我们通过加强监管和监督的方式,来确保基因重组技术的安全性和可控性,以保护人类和环境。
总之,基因重组技术实现了从基因的组成到基因的表达的重大的跨越,对于研究基因的进化、功能和表达起着至关重要的作用。
开发和应用基因重组技术可以帮助我们探索基因组织结构,了解基因表达的调控机制,进而改善物种的质量和效率,提高植物和动物的育种水平,推进药物的发展,促进分子治疗的发展,以及促进人类健康水平的提高。
基因重组方法=全PPT文档共47页

56、死去何所道,托体同山阿。 57、春秋多佳日,登高赋新诗。 58、种豆南山下,草盛豆苗稀。晨兴 理荒秽 ,带月 荷锄归 。道狭 草木长 ,夕露 沾我衣 。衣沾 不足惜 ,但使 愿无违 。 59、相见无杂言,但道桑麻长。 60、迢迢新秋夕,亭亭月将圆。
6、最大的骄傲于最大的自卑都表示心灵的最软弱无力。——斯宾诺莎 7、自知之明是最难得的知识。——西班牙 8、勇气通往天堂,怯懦通往地狱。——塞内加 9、有时候读书是一种巧妙地避开思考的方法。——赫尔普斯 10、阅读一切好书如同和过去最杰出的人谈话。——
基因重组的方式

基因重组的方式
基因重组技术是现代生物学家重要的工具,它可以用来改变和改良生活体的性状。
本文将探讨基因重组的基本原理,以及不同的基因重组方法。
基因重组技术的核心是,从一个生物体中提取一段DNA,将其接入另一个生物体的基因组中,或者改变基因组中的一段DNA的结构。
通过这种方式,DNA的构成和表达状态得以修改,而生物体的性状也随之改变。
基因重组技术可以提供独特的功能,比如抗菌性,耐药性,抗病毒性等。
基因重组可以通过不同的方式实现,比如转基因(转录),基因突变,随机插入和正交分离。
转基因技术是将基因从一种物种转移到另一种物种,从而改变物种的性状。
转录技术则是将特定基因的信息从一种物种转移到另一种物种,从而获得新的表达特性。
基因突变技术是通过改变DNA序列来改变性状,它可以在一种物种中进行。
随机插入技术是将一段DNA植入基因组中,从而引起基因表达的改变。
正交分离技术是把多个基因混合在一起,然后通过统计学方法分解出每个基因的表达频率,从而获得更完整的基因信息。
基因重组技术在生物学上有着重要的应用。
它可以修改物种的某些性状,比如抗病毒性,抗菌性,耐药性等,检测和诊断遗传疾病,生成疫苗,开发药物疗法,研制抗农药作物等。
此外,基因重组技术也可以用于家畜和家禽的培育,从而获得更高的产量。
基因重组技术的重要性不言而喻,它是当今生物学发展的重要支
柱。
基因重组技术的大规模应用,将为人类的健康,社会的发展和经济的可持续发展提供强大的支撑。
因此,我们有必要加强对基因重组技术的研究,确保其安全可靠,从而促进和发展生物技术,并为人类社会提供更多的可能性。
基因重组技术

基因重组技术一、什么是基因重组基因重组技术是以遗传基因工程方法将植物、动物或微生物等生物体改良其特性,而达成的一种新生命体。
基因可以透过复制的过程,将遗传信息传递给下一代,从而控制生物的个体性状表现,除此之外,基因还可以制造出一些结构蛋白,直接或间接地影响生物的生理表现。
而目前的基因工程技术,其发展乃建立在过去四十多年分子生物学的成果上,可说是分子生物学的延伸应用。
其基本原理是利用能切割特定DNA序列的限制酵素,将来自不同生物的DNA作切割,再以连接酵素连接带有相同切口的DNA片段,如此,在应用上便可将一些特殊DNA片段与载体进行接合,形成重组DNA,如果再将这重组DNA,经转形的作用送入宿主细胞中,经由宿主细胞的不断分裂,便可持续的复制这DNA片段。
整个基因工程技术实际上就是这些不同工具间的组合搭配,以期达到最好的基因表现或基因转殖的目的。
二、基因重组技术的方法基因改造生物主要是透过以下三种基因重组技术方式来制造:技术方式增加法为了改变动物或植物的表现性状,而从某一物种抽取个别基因,将其殖入另一动物或植物的基因组内,例如将人类黄体素的基因殖入酵母菌中以大量生产避孕药所需的主成份;又如把抗除草剂的基因殖入大豆,令大豆能够抗除草剂。
今天,科学家已可以将细菌、病毒、昆虫、动物,甚至人类的基因,引入植物内,制造基因改造植物。
减少法使特定动植物基因发生缺失,令动植物丧失某些原有性质与功能,如减少蕃茄内催熟基因的数量,将减缓其组织成熟软化,以延迟蕃茄的成熟期。
调节法去除或增加某固定基因的控制因子,可以改变生物特性的表现程度,甚至是功能,造成调整生物生命特性。
就像紫外线可以促成癌细胞的发生一般。
又例如透过刺激头发的基因,改变头发的色素;又如调节菜籽油内控制饱和脂肪酸的基因,使其减少制造饱和脂肪酸,使菜籽油的饱和脂肪酸含量降低。
1、如何起作用?每个有机体(无论是细菌还是人)的细胞都含有一个或多个DNA分子,被喻为遗传资讯库。
基因重组的4种类型

基因重组的4种类型
基因重组是指在生物体内,通过人为的调整基因的结构,使其形成新的基因序列,从而改变该生物体的性状。
目前,已经有四种不同的基因重组技术,它们是:物理基因重组、基因交换、遗传工程和基因敲除。
一、物理基因重组
物理基因重组技术是由美国科学家 Paul Berg 于1972 年提出的。
它是一种将两个不同的 DNA 分子混合在一起,并将其中一个DNA 分子的基因复制到另一个DNA 分子上的技术。
这种技术可以用来制作“融合”基因,它们包含来自两个不同DNA 分子的基因序列。
物理基因重组技术的主要应用是在生物制造中。
二、基因交换
基因交换是指在不同的生物体之间,使用一种类似“入侵”的方法,将基因序列从一个生物体转移到另一个生物体中。
这种技术常用于改变植物和动物的性状。
它也被用于开发新的药物,如疫苗和抗生素。
三、遗传工程
遗传工程是指将一种基因插入另一种基因组中,从而改变该细胞的性状。
这项技术可以被用来开发新的品种,
增强植物或动物的抗病能力,改变食物的营养价值,以及增强和改变植物和动物的性状。
四、基因敲除
基因敲除是一种技术,它可以用来破坏特定的基因,从而改变植物或动物的性状。
基因敲除技术可以用来研究特定基因对生物体的影响,以及基因突变对性状的影响。
基因敲除技术也可以用来研究不同生物体之间基因的互作关系。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子, 特称为F′因子。
F′×F-与F+×F-的不同:给体的
部分染色体基因随F′一起转入受体细胞
a)与染色体发生重组;
b)继续存在于F′因子上, 形成一种部分二倍体;
3.1.2 细菌的转导(transduction)
缺陷噬菌体在宿主细胞内能够象正常的λDNA分子一样进行复制、
包装,提供所需要的裂解功能,形成转导颗粒。
但没有正常噬菌体的溶源性和增殖能力,感染受体细胞后,通过 DNA整合进宿主染色体而形成稳定的转导子。
局限性转导与普遍性转导的主要区别:
a)被转导的基因共价地与噬菌体DNA连接,与噬菌体DNA一起 进行复制、包装以及被导入受体细胞中。而普遍性转导包装的
1) 意外的发现
的其它菌种是否也存在接合作用,用二株具不同的多重营养缺陷型 的鼠伤寒沙门氏菌进行类似的实验:
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外
用“U”型管进行同样的实验时,在给体和受体细胞
不接触的情况下,同样出现原养型细菌!
沙门氏菌LT22A是携带P22噬菌体的溶源性细菌 另一株是非溶源性细菌
3.1 原核生物的基因重组
细菌的三种水平基因转移形式
接合
转导 自然转化
接合 (conjugation): 细胞与细胞的直接接触(由F因子介导) 转导(transduction):
由 genetic transformation):
游离DNA分子 + 感受态细胞
证实接合过程需要细胞间的直接接触的 “U”型管实验( Bernard Davis,1950 )
(2) 机制
(大肠杆菌的接合机制)
接合作用是由一种被称为F因子的质粒介导
F因子的分子量通常为5×107,上面有编码细菌产生性菌毛 (sex pili)及控制接合过程进行的20多个基因。
含有F因子的细胞:“雄性”菌株(F+),其细胞表面有性菌毛 不含F因子的细胞:“雌性”菌株(F-),细胞表面没有性菌毛
可能全部是宿主菌的基因。
b)局限性转导颗粒携带特定的染色体片段并将固定的个别基因 导入受体,故称为局限性转导。而普遍性转导携带的宿主基 因具有随机性。
溶源转变(lysogenic conversion):
一个与转导相似又不同的现象 温和噬菌体感染细胞后使之发生溶源化,因噬菌体的基因 整合到宿主染色体上,而使后者获得了新性状的现象。
特点:在选择培养基平板上形成微小菌落
转导DNA不能进行重组和复制,但其 携带的基因可经过转录而得到表达。
流产转导(abortive transduction) 进入受体的外源DNA通过与细胞染色体 的重组交换而形成稳定的转导子
外源DNA被降解,转导失败。
(2)局限性转导(specialized transduction)
摄取DNA的能力,或人为地将DNA导入细胞内。
(该过程与细菌自身的遗传控制无关!)
(1)自然遗传转化(简称自然转化)
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae) 的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
进行自然转化,需要二方面必要的条件:
因为染色体上的pac与P22 DNA的pac序列不完全相同, 利用效率较低,这种“错装”机率一般仅约10-6-10-8
普遍性转导的基本要求:
形成转导颗粒的噬菌体可以是温和的也可以是烈性的,但必须 具有能偶尔识别宿主DNA的包装机制并在宿主基因组完全降解 以前进行包装。
DNA 不能复制,因此群体中仅一个细胞含有DNA, 普遍性转导的三种后果: 而其它细胞只能得到其基因产物,形成微小菌落。
d)F′菌株,Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子,特称为F′因 子。 细胞表面同样有性菌毛。
1) F+×F-杂交
F+菌株的F因子向F-细胞转移,但含F因子的宿主细胞 的染色体DNA一般不被转移。
杂交的结果:给体细胞和受体细胞均成为F+细胞
理化因子的处理可将F因子消除而使F+菌株变成F-菌株
温和噬菌体感染
整合到细菌染色体的特定位点上
宿主细胞发生溶源化
溶源菌因诱导而发生裂解时, 在前噬菌体二侧的少数宿主 基因因偶尔发生的不正常切 割而连在噬菌体DNA上 部分缺陷的温和噬菌体
把供体菌的少数特定基因转移到受体菌中
温和噬菌体λ裂解时的
不正常切割:包含gal或bio基因
(几率一般仅有10-6)
建立了感受态的受体细胞 外源游离DNA分子
枯草芽孢杆菌的自然转化过程(革兰氏阳性菌的转化模型)
分泌感受态因子 与细胞表面受 体M相互作用
使细胞表面的 DNA结合蛋白及 核酸酶裸露出来 ,使其具有与 DNA结合的活性
诱导特异蛋白质如自溶素表达
双链DNA与感受态细胞表面的特定位 点吸附,DNA的一条单链被降解,另 一条单链进入受体细胞,并与受体 细胞染色体DNA的同源部分配对,接 着受体染色体上相应单链片段被切 除,并被外来的单链DNA交换、整合 和取代,形成杂种DNA,
Hfr 菌株仍然保持着 F+细胞的特征,具有F性菌毛,并象F+一样与F2) Hfr ×F-杂交 细胞进行接合。所不同的是,当OriT序列被缺刻螺旋酶识别而产生 Hfr菌株的F因子插入到染色体DNA上,因此只要发生接合转移转移 缺口后,F因子的先导区(leading region)结合着染色体 DNA向受体细 过程,就可以把部分甚至全部细菌染色体传递给F 细胞并发生重组 胞转移,F因子除先导区以外,其余绝大部分是处于转移染色体的末 ,由此而得名为 。 端,由于转移过程常被中断,因此 F因子不易转入受体细胞中,故 Hfr×F-杂交后的受体细胞(或接合子)大多数仍然是F-。
LA-22端出现了原养型 的个体(his+,try+)
溶源性菌株LA-22 中少数细胞在培养 过程中自发释放出温和型噬菌体P22
通过滤板感染另一端的敏感菌株LA-2 LA-2裂解后,产生大量的“可滤过因子 ”,其中极少数在成熟过程中包裹了 LA-2的DNA片段(含try+基因)
通过滤板再度感染LA-22
3.1.1细菌的接合作用(conjugation) (参见 P 215)
通过细胞与细胞的直接接触而产生的 遗传信息的转移和重组过程
(1).
1946年,Joshua Lederberg 和Edward L.Taturm
细菌的多重营养缺陷型杂交实验
中间平板上长出的原养型菌落 是两菌株之间发生了遗传交换 和重组所致!
b)决定因素也各有不同;
(2)人工转化
在自然转化的基础上发展和建立的一项细菌基因重组手段, 是基因工程的奠基石和基础技术。 不是由细菌自身的基因所控制; 用多种不同的技术处理受体细胞,使其人为地处于一 种可以摄取外源DNA的“人工感受态”。 用CaCl2处理细胞,电穿孔等是常用的人工转化手段。
质粒的转化效率高;
3、基因重组
基因重组(gene recombination)是两个独立基因组内 的遗传基因,通过交换与重新组合形成新的稳定基因组 的过程。
原核微生物基因组 通常只是部分遗传物质的转移和重
组,并且通过转化、接合和转导等形式进行;
真核微生物的基因重组 发生在有性繁殖过程中,通过
减数分裂后整套染色体发生高频率交换(基因重组) 在真核微生物中的部分真菌存在不通过减数分裂而在有 丝分裂过程产生低频率基因重组的准性生殖方式
3.2 真核生物的基因重组
在自然环境中,真核微生物就存在有性生殖和准 性生殖等基因重组的形式。
酵母菌有性杂交 霉菌的准性杂交
3.2.1 酵母菌的有性杂交
酵母菌的生活史既包括二倍体、单倍 体世代,又包括有性和无性世代。 减数分裂形成子囊孢子
二倍体
单倍体
每个子囊中含 有四个单倍体 的子囊孢子
酿酒酵母的双倍体和单倍体细胞比较
溶源转变与转导的不同?
a)不携带任何供体菌的基因; b)这种噬菌体是完整的,而不是缺陷的;
3.1.3细菌的遗传转化(genetic transformation)
定义:同源或异源的游离DNA分子(质粒和染色体DNA)被自然
或人工感受态细胞摄取,并得到表达的水平方向的基因转移过程
感受态细胞:具有摄取外源DNA能力的细胞
F因子为附加体质粒 既可以脱离染色体在细胞内独立存在,也可插入(整合)到染色体上
F因子的四种细胞形式
a)F-菌株, 不含F因子,没有性菌毛,但可以通过 接合作用接收 F因子而变成雄性菌株(F+);
b)F+菌株, F因子独立存在,细胞表面有性菌毛。
c)Hfr菌株,F因子插入到染色体DNA上,细胞表面有性菌毛。
细胞与细胞的直接接触(由F因子介导)
转导(transduction):
由噬菌体介导 游离DNA分子 + 感受态细胞
自然遗传转化(natural genetic transformation):
“接合” “转导” 及“自然转化”这三种在自然界 存在的细菌遗传重组过程各自的特点:
a)外源DNA的来源及进入途径有差异;
酿酒酵母
两种接合型α和a (单倍体)
融合
二倍体细胞(α/a)
酿酒酵母接合型是由自身的遗传特性决定的
受MAT活性区调控,由MAT启动子控制
α和a基因是不表达的沉默基因,只在发生接合型 (单倍体)转变时用作α或a基因插入的来源
(competent cell)
自然遗传转化(natural genetic transformation) 人工转化(artificial transformation)