乳糖操纵子和色氨酸操纵子
乳糖操纵子复习题

乳糖操纵子复习题
乳糖操纵子是分子生物学中一个重要的概念,它涉及到基因表达的调
控机制。
以下是关于乳糖操纵子的复习题内容:
1. 定义:请简述乳糖操纵子是什么,并解释其在细胞中的作用。
2. 组成:描述乳糖操纵子的基本组成部分,包括启动子、操纵基因、
结构基因等。
3. 调控机制:解释乳糖操纵子的正调控和负调控机制是如何工作的。
4. 诱导:阐述乳糖如何作为诱导剂激活乳糖操纵子的表达。
5. 抑制:描述在没有乳糖的情况下,乳糖操纵子是如何被抑制的。
6. cAMP-CAP复合物:解释cAMP-CAP复合物在乳糖操纵子调控中的作用。
7. 乳糖操纵子的发现:简述乳糖操纵子是如何被发现的,以及这一发
现对分子生物学的意义。
8. 应用:讨论乳糖操纵子在现代生物技术中的应用,特别是在基因工
程和基因治疗中的作用。
9. 比较:将乳糖操纵子与其他类型的操纵子(如色氨酸操纵子)进行
比较,指出它们的异同点。
10. 实验研究:列举一些实验方法,用于研究乳糖操纵子的调控机制。
11. 问题解决:提出一些可能在研究乳糖操纵子时遇到的问题,并给出可能的解决方案。
12. 未来方向:探讨乳糖操纵子研究的未来方向,以及这些研究可能对医学和生物技术带来的影响。
通过这些问题的复习,可以加深对乳糖操纵子及其调控机制的理解,为进一步的学习和研究打下坚实的基础。
乳糖操纵子的调控机理PPT课件

-
27
当有色氨酸时
无色氨酸时
-
28
色氨酸操纵子中的操纵基因和衰减子可以起双重负 调节作用。衰减子可能比操纵基因更灵敏, 只要 色氨酸一增多,即使不足以诱导阻遏蛋白结合操纵 基因,就足可以使大量的mRNA提前终止。反之, 当色氨酸减少时,即使失去了诱导阻遏蛋白的阻遏 作用,但只要还可以维持前导肽的合成,仍继续阻 止转录。这样可以保证细菌对色氨酸的充分利用。 防止堆积。
-
36
4、翻译产物对mRNA的翻译进行调控
有些mRNA编码的蛋白质,本身也可以对 相应的mRNA的翻译过程产生调节作用,这 是一种自身翻译调控作用。 ①核糖体蛋白翻译的自身调控 ② 翻译的RF2调节自身的翻译
UGAC
25
315
编码RF2蛋白m- RNA
37
5、小分子RNA抑制特定mRNA的翻译
❶小分子RNA调整基因表达产物的类型:
成
-
30
-
31
二、翻译水平的调控
1、SD序列对翻译的影响
SD序列(Shine-Dalgarno sequence): mRNA起 始密码前的一段富含嘌呤核苷酸的序列。(9-12bp)
❶SD序列的差异对翻译的影响 ❷ SD序列位置对翻译的影响
-
32
-
33
2、mRNA二级结构隐蔽SD序列
某些mRNA分子中,核糖体结合位点在一个二级 结构中(茎环)中,使核糖体无法结合,只有 打破茎环结构,核糖体才能结合。例如:带有 红霉素抗性的细菌
•基因表达调控的环节:
–基因活化、转录、转录后加工、翻译、 翻译后加工
-
6
第一节 原核生物基因表达调控
一、转录水平的调控是原核生物的主要调控环节
乳糖操纵子与色氨酸操纵子的区别
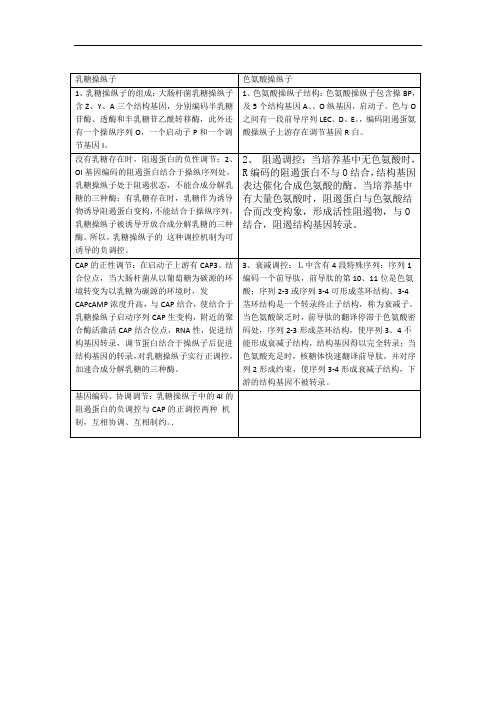
色氨酸操纵子
1、乳糖操纵子的组成:大肠杆菌乳糖操纵子含Z、Y、A三个结构基因,分别编码半乳糖苷酶、透酶和半乳糖苷乙酰转移酶,此外还有一个操纵序列O,一个启动子P和一个调节基因I。
1、色氨酸操纵子结构:色氨酸操纵子包含操BP,及5个结构基因A、、O纵基因,启动子。色与O之间有一段前导序列LEC、D、E。,编码阻遏蛋氨酸操纵子上游存在调节基因R白。
3是色氨酸;序列2-3或序列3-4可形成茎环结构。3-4茎环结构是一个转录终止子结构,称为衰减子。当色氨酸缺乏时,前导肽的翻译停滞于色氨酸密码处,序列2-3形成茎环结构,使序列3、4不能形成衰减子结构,结构基因得以完全转录;当色氨酸充足时,核糖体快速翻译前导肽,并对序列2形成约束,使序列3-4形成衰减子结构,下游的结构基因不被转录。
没有乳糖存在时,阻遏蛋白的负性调节:2、OI基因编码的阻遏蛋白结合于操纵序列处,乳糖操纵子处于阻遏状态,不能合成分解乳糖的三种酶;有乳糖存在时,乳糖作为诱导物诱导阻遏蛋白变构,不能结合于操纵序列,乳糖操纵子被诱导开放合成分解乳糖的三种酶。所以,乳糖操纵子的这种调控机制为可诱导的负调控。
2、阻遏调控:当培养基中无色氨酸时,R编码的阻遏蛋白不与O结合,结构基因表达催化合成色氨酸的酶。当培养基中有大量色氨酸时,阻遏蛋白与色氨酸结合而改变构象,形成活性阻遏物,与O结合,阻遏结构基因转录。
基因编码、协调调节:乳糖操纵子中的4I的阻遏蛋白的负调控与CAP的正调控两种机制,互相协调、互相制约。.
CAP的正性调节:在启动子上游有CAP3、结合位点,当大肠杆菌从以葡萄糖为碳源的环境转变为以乳糖为碳源的环境时,发CAPcAMP浓度升高,与CAP结合,使结合于乳糖操纵子启动序列CAP生变构,附近的聚合酶活激活CAP结合位点,RNA性,促进结构基因转录,调节蛋白结合于操纵子后促进结构基因的转录,对乳糖操纵子实行正调控,加速合成分解乳糖的三种酶。
乳糖操纵子的表达调控

阻遏调控机制
阻遏蛋白有活性 阻遏蛋白无活性
三.色氨酸操纵子的弱化调控机制
实验观察表明:当色氨酸达到一定浓度、
但还没有高到能够活化阻遏蛋白使其起阻 遏作用的程度时,产生色氨酸合成酶类的 量已经明显降低,而且产生的酶量与色氨 酸浓度呈负相关。仔细研究发现这种调控 现象与色氨酸操纵子弱化调控机制有关。
前导序列
研究还发现,当mRNA 合成起始以后,除非培养
基中完全没有色氨酸,否则转录总在这个区域停 止,这就是123-150序列缺失提高色氨酸基因表达 的原因。因为转录发生在这个区域并且这种终止 能被调节,因此这个区域被称为弱化子或衰减子。 该区域序列的mRNA可通过自我配对形成茎-环结构, 具有典型的终止子的结构特点。
。茎-环结构
mRNA前导区序列分析
trp前导区的碱基序列已经全部测定,发现完整 的前导序列可分为1、2、3、4区域,这四个区 域的片段能以两种不同的方式进行碱基配对, 有时以1-2和3-4配对,有时只以2-3方式互补配 对。而3-4配对区正好位于终止密码子识别区, 当这个区域发生破坏自我碱基突变,有利于转 录的继续进行。
乳糖操纵子的正调控机制如图三
正调控机制
图三
正调控意义
由于Plac是弱启动子,单纯因乳糖的存在发生去阻遏 使lac操纵子转录开放,还不能使细菌很好利用乳糖, 必需同时有CAP来加强转录活性,细菌才能合成足 够的酶来利用乳糖。lac操纵子的强诱导既需要有乳 糖的存在又需要没有葡萄糖可供利用。通过这机制, 细菌是优先利用环境中的葡萄糖,只有无葡萄糖而 又有乳糖时,细菌才去充分利用乳糖。
转录不终止
RNA聚合酶继续转录
弱化作用的意义
细菌通过弱化作用弥补阻遏作用的不足, 因为阻遏作用只能使转录不起始,对于已 经起始的转录,只能通过弱化作用使之中 途停下来。阻遏作用的信号是细胞内色氨 酸的多少;弱化作用的信号则是细胞内载 有色氨酸的tRNA的多少。它通过前导肽的 翻译来控制转录的进行,在细菌细胞内这 两种作用相辅相成,体现着生物体内周密 的调控作用。
乳糖操纵子

(3) 操纵区
操纵区(operator)是指能被调控蛋白 特异性结合的一段DNA序列,常与启动子邻近 或与启动子序列重叠,当调控蛋白结合在操 纵子序列上,会影响其下游基因转录的强弱。
16 07.04.2021
以 前 将 操 纵 区 称 为 操 纵 基 因 ( operator gene)。但现在基因定义是为蛋白质或RNA编 码的核酸序列,而操纵序列并不是编码蛋白 质的基因,却是起着调控基因表达强弱的作 用,正如启动序列不叫启动基因而称为启动 子一样,操纵序列就可称为操纵区。operon 译为操纵子,即基因表达操纵的单元之意。
5 07.04.2021
这种典型的诱导现象,是研究基因表达调控极 好的模型。针对大肠杆菌利用乳糖的适应现象, 法国的Jocob和Monod等人做了一系列遗传学和 生 化 学 研 究 实 验 , 于 1961 年 提 出 乳 糖 操 纵 子
(lac operon)学说。
6 07.04.2021
2. 操纵子的基本组成
13 07.04.2021
14 07.04.2021
(2) 启动子
启动子(promoter, P)是指能被RNA聚合酶
识别、结合并启动基因转录的一段DNA序列。操 纵子至少有一个启动子,一般在第一个结构基 因5'侧上游,控制整个结构基因群的转录。
用RNA聚合酶与分离的一段DNA双链混合, 再加入外切核酸酶去水解DNA,结果只有被RNA 聚合酶识别结合而被保护的那段DNA不被水解, 由此可以测出启动子的范围及其序列。
18 07.04.2021
阻遏蛋白与操纵区结合,就妨碍了RNA聚合 酶与启动子的结合及其后ß -半乳糖苷酶等基 因的转录起始,从而阻遏了这群基因的表达。
分子生物学简答题

3)原核转录起始不需要转录因子,RNA聚合酶直接结合启动子,由σ因子决定基因表达的特异性;真核转录起始需要基础、特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用,调控转录激活。
4)原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白质的协调表达机制更为复杂。
(6)转录后调控包括对mRNA的加工修饰、转运、细胞质定位以及稳定性等多方面的调控。翻译调控点主要在起始阶段和延长阶段,翻译起始因子的磷酸化可调节蛋白质翻译。另外,小分子RNA通过干扰翻译过程抑制基因表达。
真核基因表达调控特点:1)既有瞬时调控,又有发育调控2)调控环节更多3)染色质结构变化影响转录效率4)转录调控以正调控为主5)调控元件复杂并且可以远离转录区6)转录因子种类多,调控机制更复杂
(1)具有自主复制起点,使载体在宿主细胞中进行自主复制,并能使克隆的外源DNA得到同步扩增;(2)至少有一个筛选标志;(3)有适宜的限制性核酸内切酶单一酶切位点,可供外源基因插入时选择。
11.简述分子生物学实验中的α互补和蓝白斑筛选的原理。
β-半乳糖苷酶(β-gal)的α片段与受体菌编码的ω片段(lacZ-ω)可以互补结合发挥β-gal的活性,称作α互补。一些质粒上带有β-半乳糖苷酶α片段的编码序列LacZ’,转化进入受体菌,可形成α互补,即可催化底物X-gal产生蓝色产物,使菌落变蓝。由于这些质粒的多克隆位点位于LacZ’内部,插入外源DNA片段后,使LacZ’不能编码产生有功能的β-gal的α片段,不能发生α互补,在X-gal存在下受体菌落呈白色。因此,蓝色菌落代表载体中LacZ’基因活性完好无损,没有插入外源DNA片段,白色菌落则表明着所含质粒带有外源DNA片段,为重组质粒。用蓝白斑筛选可以来区分转化进入受体菌的是空载体还是重组质粒。
医学生物化学(第十五章)

(2) 锌指 (zinc finger) 约30氨基酸残基,4个氨基酸残基(两个cys,两个 his, 或4个cys)以配位键与Zn2+相互作用
(3)亮氨酸拉链 (leucine zippers) 一段肽链中每隔7个氨基酸即有一个亮氨酸,该肽段所 形成的螺旋可出现疏水及亲水二个面,疏水面即亮氨 酸拉链。
3.反应元件
概念: 特点:协同作用。
4. 沉默子(Silencer)
概念: 特点:负性调节元件。
2 反式作用因子
两个必需结构域: 与顺式元件结合的结构域 与反式元件或RNA聚合酶结合的激活结构域
3 反式作用因子的结构模式
(1) α螺旋—β转角—α螺旋 (helix- turn- helix)
其中一个为识别螺旋,含有较多能与DNA相互 作用的AA残基
一、具有转录活性的染色质结构的变化
—便于RNA聚合酶及转录因子附着
1.DNase I 超敏感位点 ( DNase I hypersensitive site) 一般100-200 bp,转录基因5‘端 1000bp 内,一般不存 在核小体结构 2.DNA拓扑结构变化; 3.组蛋白变化:H1蛋白减少;其他组蛋白发生乙酰化、 甲基化等修饰 4.DNA甲基化修饰发生变化:去甲基化 m5CpG→CpG
二、参与基因调控的顺式作用元件和反式 作用因子
1 顺式作用元件
和被转录的结构基因在距离上比较接近的DNA序列 1. 启动子和启动子上游近侧序列 TATA box CAAT box GC box CpG岛 (MeCP1、 MeCP2)
2. 增强子(enhancer)
概念: 特点:不受与启动子距离、序列及方向的制约; 有组织特异性 。
第一节 原核生物基因表达的调控
色氨酸操纵子

色氨酸操纵子色氨酸是构成蛋白质的组分,一般的环境难以给细菌提供足够的色氨酸,细菌要生存繁殖通常需要自己经过许多步骤合成色氨酸,但是一旦环境能够提供色氨酸时,细菌就会充分利用外界的色氨酸、减少或停止合成色氨酸,以减轻自己的负担。
细菌这种对色氨酸利用的调节是通过色氨酸操纵子(trp operon)来实现的。
一、色氨酸操纵子的结构与阻遏蛋白的负性调控色氨酸操纵子的结构与乳糖操纵子相似,结构基因由合成色氨酸所需要酶类的基因E、D、C、B、A等头尾相接串连排列组成,结构基因上游为启动子P trp 和操纵序列O,不过其调控基因trpR的位置远离P-O-结构基因群,在其自身的启动子作用下,以组成性方式低水平表达其编码分子量为47KD的调控蛋白R。
点击后看大图色氨酸操纵子是属于一种负性调控的、可阻遏的操纵子。
以组成性方式低水平表达的阻遏蛋白R并不具有与O结合的活性,只有当环境能提供足够浓度的色氨酸时,R与色氨酸结合后构象变化,才能够与操纵序列O特异性亲和结合,阻遏结构基因的转录。
因此这类操纵子通常是开放转录的,有效应物(色氨酸为阻遏剂)作用时则关闭转录。
细菌不少生物合成系统的操纵子都属于这种类型,其调控可使细菌处在生存繁殖最经济最节省的状态。
二、衰减子及其作用实验观察表明:当色氨酸达到一定浓度、但还没有高到能够活化R使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,而且产生的酶量与色氨酸浓度呈负相关。
仔细研究发现这种调控现象受转录衰减(attenuation)机制的调节。
在色氨酸操纵子P trp-O与第一个结构基因trpE之间有一段162bp的前导序列构成衰减子区域(attenuator region),研究证明当色氨酸有一定浓度时,RNA 聚合酶的转录会终止在这里。
这段序列能够编码14个氨基酸的短肽,其中有2个色氨酸相连,在此编码区前有核糖体识别结合位点(RBS)序列,提示这段短序列在转录后是能被翻译的。
乳糖操纵子与色氨酸操纵子的区别
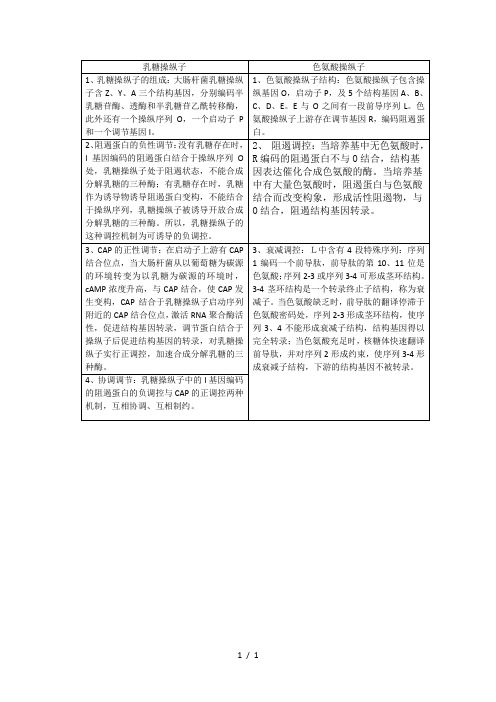
色氨酸操纵子
1、乳糖操纵子的组成:大肠杆菌乳糖操纵子含Z、Y、A三个结构基因,分别编码半乳糖苷酶、透酶和半乳糖苷乙酰转移酶,此外还有一个操纵序列O,一个启动子P和一个调节基因I。
1、色氨酸操纵子结构:色氨酸操纵子包含操纵基因O,启动子P,及5个结构基因A、B、C、D、E。E与O之间有一段前导序列L。色氨酸操纵子上游存在调节基因R,编码阻遏蛋白。
4、协调调节:乳糖操纵子中的I基因编码的阻遏蛋白的负调控与CAP的正调控两种机制,互相协调、互相制约。
3、衰减调控:L中含有4段特殊序列:序列1编码一个前导肽,前导肽的第10、11位是色氨酸;序列2-3或序列3-4可形成茎环结构。3-4茎环结构是一个转录终止子结构,称为衰减子。当色氨酸缺乏时,前导肽的翻译停滞于色氨酸密码处,序列2-3形成茎环结构,使序列3、4不能形成衰减子结构,结构基因得以完全转录;当色氨酸充足时,核糖体快速翻译前导肽,并对序列2形成约束,使序列3-4形成衰减子结构,下游的结构基因不被转录。
3、CAP的正性调节:在启动子上游有CAP结合位点,当大肠杆菌从以葡萄糖为碳源的环境转变为以乳糖为碳源的环境时,cAMP浓度升高,与CAP结合,使CAP发生变构,CAP结合于乳糖操纵子启动序列附近的CAP结合位点,激活RNA聚合酶活性,促进结构基因转录,调节蛋白结合于操纵子后促进结构基因的转录,对乳糖操纵子实行正调控,加速合成分解乳糖的三种酶。
2、阻遏蛋白的负性调节:没有乳糖存在时,I基因编码的阻遏蛋白结合于操纵序列O处,乳糖操纵子处于阻遏状态,不能合成分解乳糖的三种酶;有乳糖存在时,乳糖作为诱导物诱导阻遏蛋白变构,不能结合于操纵序列,乳糖操纵子被诱导开放合成分解乳糖的三种酶。所以,乳糖操纵子的这种调控机制为可诱导的负调控。
乳糖操纵子和色氨酸操纵子

大肠杆菌乳糖操纵子包括4类基因:①结构基因,能通过转录、翻译使细胞产生一定的酶系统和结构蛋白,这是与生物性状的发育和表型直接相关的基因。
乳糖操纵子包含3个结构基因:lacZ、lacY、lacA。
LacZ合成β—半乳糖苷酶,lacY合成透过酶,lacA合成乙酰基转移酶。
②操纵基因O,控制结构基因的转录速度,位于结构基因的附近,本身不能转录成m RNA。
③启动基因P,位于操纵基因的附近,它的作用是发出信号,mRNA合成开始,该基因也不能转录成m RNA。
④调节基因i:可调节操纵基因的活动,调节基因能转录出mRNA,并合成一种蛋白,称阻遏蛋白。
操纵基因、启动基因和结构基因共同组成一个单位——操纵子(ope ron)。
调节乳糖催化酶产生的操纵子就称为乳糖操纵子。
其调控机制简述如下:抑制作用:调节基因转录出mRNA,合成阻遏蛋白,因缺少乳糖,阻遏蛋白因其构象能够识别操纵基因并结合到操纵基因上,因此RNA聚合酶就不能与启动基因结合,结构基因也被抑制,结果结构基因不能转录出m RNA,不能翻译酶蛋白。
诱导作用:乳糖的存在情况下,乳糖代谢产生别乳糖(allo Lacto se),别乳糖能和调节基因产生的阻遏蛋白结合,使阻遏蛋白改变构象,不能在和操纵基因结合,失去阻遏作用,结果RN A聚合酶便与启动基因结合,并使结构基因活化,转录出mRNA,翻译出酶蛋白。
负反馈:细胞质中有了β—半乳糖苷酶后,便催化分解乳糖为半乳糖和葡萄糖。
乳糖被分解后,又造成了阻遏蛋白与操纵基因结合,使结构基因关闭。
3、详述大肠杆菌色氨酸操纵子的调控机理。
(12分)答:大肠杆菌色氨酸操纵子的转录受阻遏和衰减两种机制的控制,前者通过阻遏蛋白和操纵基因的作用控制转录的起始,后者通过前导序列形成特殊的空间结构控制转录起始后是否进行下去。
基础生物化学—赵武玲

第三节原核转录调控P371页细胞的基因表达是指由DNA转录成RNA再翻译成蛋白质的过程,是受到严格的调控的。
细胞响应调节信号,使基因表达产物的水平升高或降低的过程,就称之为基因表达调控(regulated gene expression)。
基因的表达调控可以在多种水平上进行,如DNA结构的调控、转录水平的调控及翻译水平的调控。
原核生物基因表达调控以转录起始水平的调控为主。
下面以乳糖操纵子和色氨酸操纵子为例介绍原核生物转录起始阶段的调控。
一、乳糖操纵子操纵子(operon)模型很好地说明了原核生物基因表达的调节机制,在原核生物基因调控中具有普遍性。
操纵子是原核生物染色体上控制蛋白质合成的功能单位,包括结构基因区(structural gene region)和调控区。
有些操纵子中还具有其他的位点。
结构基因由功能上彼此相关的几个基因组成,编码具有酶功能或结构功能的蛋白质。
一个操纵子中若干个结构基因排列在一起,它们的表达作为一个整体受到调控区的调节,通过转录形成的是一条多顺反子mRNA。
调控区由启动子(promoter,P)、操纵序列(operator,O)所组成。
调控区可接受调节基因(regulatory gene)产物的调节。
调节基因不在操纵子内,它编码调节蛋白,结合在DNA的特殊位点上调节基因表达。
如果调节基因编码的蛋白质与操纵序列结合后激活了基因的表达,这样的调控被称为正调控(positive control),相应的调节蛋白就称为激活蛋白(activator)。
如果调节基因编码的蛋白质与操纵序列结合后阻遏了基因的表达,这样的调控就称为负调控(negative control),相应的调节蛋白就称为阻遏蛋白(repressor)。
大肠杆菌的乳糖操纵子模型(ιac operon )是第1个被阐明的基因表达系统,由Francois Jacob和Jacques Monod于1962年提出的。
大肠杆菌乳糖操纵子有3个结构基因Z,Y,A,分别编码3种参与乳糖分解代谢的酶,即β-半乳糖昔酶(β—galactosidase)、β-半乳糖昔透过酶(permease)和硫代半乳糖昔转乙酞基酶(thiogalactoside transacetylase)。
第7章原核生物基因表达的调控

Z编码β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖。
Y编码β-半乳糖苷透过酶:使外界的β-半乳糖苷(如乳糖)能透过大肠杆
菌细胞壁和原生质膜进入细胞内。
A编码β-半乳糖苷乙酰基转移酶:乙酰辅酶A上的乙酰基转到β-半乳糖苷
上,形成乙酰半乳糖。
gene
正调控
调控蛋白
负调控
结构基因表达
▪ 负调控:抑制基因表达的调控方式 ▪ 正调控:促进基因表达的调控方式
B、特殊代谢物的调控
诱导(induction)
阻遏(repression)
inducer
gene
repressor
gene
特殊代谢物
诱导 阻遏
结构基因表达
诱导物、可诱导基因 阻遏物、可阻遏基因
无葡萄糖、 有乳糖-----cAMP水平高 (2)cAMP与CRP结合形成有活性的
CRP- cAMP 复合物 (3)CRP-cAMP 与Plac结合 (4)增强了RNA聚合酶与启动子的结合
(5)lacZ, lacY 、 lacA高表达
105
40
105
41
乳糖、G存在与否及与操纵子正、负控因素、 基因开放与关闭情况如下:
CRP
Binding
RNA
Promoter
Operator
CRP
Pol. Repressor
cAMP
LacZ
LacY
LacA
Repressor mRNA
STOP
Right there
CRP
Polymerase
cAMP
Repressor
cAMP
CRP
第11章 原核生物基因表达的调控

Ø 葡萄糖代谢导致cAMP浓度下降; Ø cAMP可以活化乳糖操纵子的激活蛋白:
CRP: cAMP receptor protein(cAMP受体蛋白) CAP: catabolite gene activator protein
(代谢降解物活化蛋白)
Ø cAMP-CRP/CAP
乳糖操纵子的正调控
Ø 每个阻遏蛋白四聚体与两个 operator 结合; Ø 阻遏蛋白与Operator结合导 致DNA弯折,干扰mRNA的 合成。
p.286 图11-7
乳糖操纵子的正调控
当细菌在含有葡萄糖和乳 糖的培养基中生长时,通常 总是优先利用葡萄糖,而不 利用乳糖;只有当葡萄糖耗 尽后,细菌经过一段停滞期, 才能在乳糖的诱导下,合成 β-半乳糖苷酶等分解利用 乳糖的酶类,细菌才能利用 乳糖。
ttrrppRR
OOPPtrptrEpE trptDrpDtrpCtrpCtrpBtrpBtrpAtrpA
ttrrppRR
OOPPtrptrEpE trptDrpDtrpCtrpCtrpBtrpBtrpAtrpA
色氨酸操纵子的衰减作用
trpR
OP trpL trpE trpD trpC trpB trpA
5’
(1) 新合成的正链 RNA可以翻译A蛋白;
3’ (-) A
5’(+)
5’
但是很快形成二级结构,阻止A蛋白 的继续合成;
所以 A蛋白与C蛋白的量为1:180
Ø Rep的合成依赖于C蛋白的表达, 证据:C基因的codon6发生无义突 变:核糖体停留在该处,导致rep基 因RBS附近的二级结构无法打开, 则rep基因无法表达。
AraC既是阻遏蛋白, 又是激活蛋白;
原核表达调控

在Trp操纵子Ptrp-O与trpE间有一段162bp的先导序列(leading sequence, L)。在L内有一段123~150bp的序列,它在转录起始后可调控转录过程的进行。
葡萄糖存在时
若无Gal,Gal R可结合在gal OE和OI上,并相互作用形成环,从S1、S2的转录都被阻遏。 当gal存在时,Gal R的阻遏解除,但由于葡萄糖的存在,P1不能启动而只有P2低水平启动转录。 无葡萄糖存在;若存在gal,gal的阻遏解除,依赖于CRP的P1高水平表达产生利用Gal的酶类,以Gal为能源和碳源。
弱化子的共同特点是某些外部因素控制着弱化子的发夹形成。即形成终止子结构。弱化子为结构基因的转录设了一道关卡。
前导肽
分析前导序列发现,它包括起始密码子AUG和终止密码子UGA,能产生一个含有14个氨基酸的多肽,这个假设的多肽被称为前导肽。
在前导序列的第10和第11位上有相邻的两个色氨酸密码子。组氨酸操纵子含有7个相邻的组氨酸密码子,苯丙氨酸操纵子也有7个苯丙氨酸密码子,这些密码子参与了操纵子中的转录弱化机制。
在trp操纵子中,阻遏蛋白的负调控起到粗调的作用,而衰减子起到细调的作用。 细菌通过弱化作用弥补阻遏作用的不足,因为阻遏作用只能使转录不起始,而对于已经起始的转录,只能通过弱化作用使之中途停顿下来。阻遏作用的信号是细胞内色氨酸的多少,弱化作用的信号则是细胞内载有色氨酸的tRNA的多少,通过前导肽的翻译来控制转录的进行。 细菌细胞内这两种作用相辅相成,体现着生物体内周密的调控作用。
操纵子

操纵子(operon):指启动基因、操纵基因和一系列紧密连锁的结构基因的总称。
转录的功能单位。
很多功能上相关的基因前后相连成串,由一个共同的控制区进行转录的控制,包括结构基因以及调节基因的整个DNA序列。
主要见于原核生物的转录调控,如乳糖操纵子、阿拉伯糖操纵子、组氨酸操纵子、色氨酸操纵子等1961 年,法国巴斯德研究所的Monord 和Jacob 提出了乳糖操纵子概念,后来人们在大肠杆菌中又陆续发现了色氨酸操纵子、组氨酸操纵子、半乳糖操纵子、阿拉伯糖操纵子等多种操纵子,从而不断的充实和完善了被誉为生命第三原理的基因调控理论,在这个理论中提出的操纵子概念也被人们普遍接受和证实。
操纵子学说是关于原核基因结构及其表达调控的学说,由法国巴斯德研究所著名科学家的Monod和Jacob在1961年首先提出[1]。
他们以对乳糖操纵子的研究,通过大量的试验及分析建立了现在已经被人们广泛接受的乳糖操纵子控制模型[2]。
后来人们在大肠杆菌中又陆续发现了色氨酸操纵子、组氨酸操纵子、半乳糖操纵子、阿伯糖操纵子等多种操纵子,从而不断的充实和完善了被誉为生命第三原理的基因调控理论,在这个理论中提出的操纵子概念也被人们普遍接受和证实。
操纵子学说:1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文,提出操纵子学说,开创了基因调控的研究。
四年后的1965年,莫诺与雅可布即荣获诺贝尔生理学与医学奖。
莫诺与雅可布最初发现的是大肠杆菌的乳糖操纵子。
这是一个十分巧妙的自动控制系统,这个自动控制系统负责调控大肠杆菌的乳糖代谢。
乳糖可作为培养大肠杆菌的能源。
大肠杆菌能产生一种酶(叫做“半乳糖苷酶”),能够催化乳糖分解为半乳糖和葡萄糖,以便作进一步的代谢利用。
编码半乳糖苷酶的基因(简称z)是一个结构基因(structural gene)。
乳糖操纵子概述
半 乳 糖 苷 酶 表 达 量
β-
操纵子的发现
1961年, Francois Jacob and Jacques Monod 提出细菌基因 表达调控的操纵子学说。
1965年,获得诺贝尔生理学与 医学奖。
Jacob, Monod & Lwoff
©
操纵子(operon):很多功能上相关的
乳糖分解所需酶(+)
1940年,Monod发现大肠杆菌在含有葡萄糖和乳糖的培养基 上生长时,细菌先利用葡萄糖,葡萄糖用完以后,才利用乳糖。
通透酶(lactose permease) 促使乳糖进入细菌
细 菌 生 长 曲 线
葡萄糖耗尽 乳糖存在
β -半乳糖苷酶
(β -galactosidase)
催化乳糖分解
乳糖代谢所需酶(+)
下次课的学习内容
乳糖诱导的负调控
CAP介导的正调控 协调调控基因前后相成串,由一个共同的控制区进行转
录调控。包括结构基因及调节基因的整个DNA序 列,共同组成一个转录单位。
主要见于原核生物的转录调控,如乳糖操纵子、 色氨酸操纵子等。
操纵子(operon)
调节基因
结构基因 Structural genes
启动序列 Promoter(P)
操纵基因 Operator(O)
promoter
二. 乳糖操纵子(lac operon)
调控基因 结构基因
操纵序列
通透酶 启动序列
乙酰转移酶
β -半乳糖苷酶
阻遏蛋白基因LacI
阻遏蛋白
启动序列 RNApol
阻遏蛋白 操纵序列
编码序列
三. 乳糖操纵子调节机制
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
负反馈:细胞质中有了β—半乳糖苷酶后,便催化分解乳糖为半乳糖和葡萄糖。乳糖被分解后,又造成了阻遏蛋白与操纵控机理。(12分) 答:大肠杆菌色氨酸操纵子的转录受阻遏和衰减两种机制的控制,前者通过阻遏蛋白和操纵基因的作用控制转录的起始,后者通过前导序列形成特殊的空间结构控制转录起始后是否进行下去。 1)色氨酸操纵子的可阻遏系统: 在阻遏系统中,起负调控的调节基因的产物是一个无活性的阻遏蛋白,色氨酸是辅阻遏物;当色氨酸不足时,阻遏蛋白无活性,不能和操纵基因结合,色氨酸操纵子能够转录;当色氨酸充足时,阻遏蛋白和它结合而被激活,从而结合到操纵基因上,而色氨酸操纵子的操纵基因位于启动基因内,因此,活性阻遏物的结合排斥了RNA聚合酶的结合,从而抑制了结构基因的表达。 2)色氨酸操纵子的衰减调控 在色氨酸操纵子的操纵基因和第一个结构基因之间有一段前导序列L,在前导序列上游部分有一个核糖体结合位点,后面是以起始密码AUG开头的14个氨基酸的编码区,编码区有两个紧密相连的色氨酸密码子,后面是一个终止密码子UGA,在开放阅读框下游有一个不依赖ρ因子的终止子,是一段富含G/C的回文序列,可以形成发夹结构,因此可以在此处终止转录。另外前导序列包含4个能进行碱基互补配对的片断1区、2区、3区和4区。它们能以1、2和3、4或2、3的方式进行配对,从而使前导序列形成二级结构的变化。在细菌中,翻译与转录偶连,一旦RNA聚合酶转录出trp mRNA中的前导肽编码区,核糖体便立即结合上去翻译这一序列。当细胞中缺乏色氨酸时,Trp-tRNATrp的浓度很低,核糖体翻译前导肽至两个连续的色氨酸密码子处就陷入停顿,这时核糖体只占据1区,由RNA聚合酶转录的2区和3区便可配对,4区游离在外,这样就不能形成终止子结构,RNA聚合酶就可以一直转录下去,最后完成trp全部结构基因的转录,得到完整的mRNA分子。当细胞中存在色氨酸时,就有一定浓度的Trp-tRNATrp,核糖体便能顺利通过两个连续的色氨酸密码子而翻译出整个前导肽,直到前导肽序列后面的终止密码子UGA处停止。此时,核糖体占据了1区和2区,结果3区和4区配对,形成转录终止子结构,使RNA聚合酶终止转录。实现衰减调控的关键在于时间和空间上的 巧妙安排。在空间上,两个色氨酸密码子的位置很重要,不可随意更改;在时间上,核糖体停顿于两个色氨酸密码子上时,序列4应当还未转录出来
调节乳糖催化酶产生的操纵子就称为乳糖操纵子。其调控机制简述如下:
抑制作用:调节基因转录出mRNA,合成阻遏蛋白,因缺少乳糖,阻遏蛋白因其构象能够识别操纵基因并结合到操纵基因上,因此RNA聚合酶就不能与启动基因结合,结构基因也被抑制,结果结构基因不能转录出mRNA,不能翻译酶蛋白。
诱导作用:乳糖的存在情况下,乳糖代谢产生别乳糖(alloLactose),别乳糖能和调节基因产生的阻遏蛋白结合,使阻遏蛋白改变构象,不能在和操纵基因结合,失去阻遏作用,结果RNA聚合酶便与启动基因结合,并使结构基因活化,转录出mRNA,翻译出酶蛋白。