中慢生型天山根瘤菌中自体诱导物分子结构的鉴定-土壤
微生物育种资料名词解释1.富集培养目的微生物含量较少时,根据

微生物育种资料名词解释1.富集培养:目的微生物含量较少时,根据微生物生理特点,设计一种选择性培养基,创造有利生长条件,是目的微生物在最适环境下迅速生长繁殖,数量增加,由劣种变为优势种,以利用分离所需要的菌种。
2.营养缺陷型:野生型菌株经过人工诱变或自然突变失去合成某种营养(氨基酸、维生素、核酸等)的能力,只有在基本培养基中补充所缺失的营养因子才能生长。
3.常规杂交育种:通过接合、转化、转导、溶源转化和转染等方式来获得重组体的杂交育种方法。
4.原生质体融合育种:通过酶解破除细胞壁后,制备微生物原生质体,然后诱导原生质体融合杂交,双亲本不受亲和力限制,甚至可以打破种属间遗传障碍。
获得远缘杂交重组体的特殊方式。
5.原生质体再生育种:微生物制备原生质体后直接再生,从再生菌落中分离筛选变异菌株,最终得到优良性状提高的正变菌株。
6.原生质体诱变育种:以微生物原生质体为育种材料,采用物理或化学诱变剂处理,然后分离到再生培养基中再生,并从再生菌落中筛选高产突变菌株。
解答1.工业生产的微生物菌种的特性①在遗传上必须是稳定的②易于产生许多营养细胞、包子或其他繁殖体②必须是纯种,不应带有其他杂菌及噬菌体④种子的生长必须旺盛、迅速⑤产生所需要的产物时间短⑥比较容易分离提纯⑦有自身保护机制,抵抗杂菌污染能力强⑧能保持较长的良好经济性能⑨菌株诱变处理较敏感,从而可以选育出高产菌株⑩在规定时间内,菌株必须产生与其数量的目的产物,并保持相对地稳定2.工业微生物的发展史(1)诱变育种。
以人工诱变手段诱发微生物基因突变,改变遗传结构和功能,通过筛选,从多种多样的变异体中筛选出产量高、性状优良的变异株,并找出发挥这个变株最佳培养基和培养条件,使其在最是环境条件下合成有效产物。
(2)杂交育种。
使双亲或多亲的遗传物质重新组合,以获得综合双亲优良性状的新品种的育种方法。
(3)代谢控制育种。
进行内因改变,通过定向选育某种特定的突变型,以达到大量积累由于产物的目的,定向选育包括改变代谢代谢通路;降低支路代谢终产物产生或切断支路代谢途径及提高细胞膜通透性。
2023年北京市朝阳区高考生物二模试卷+答案解析(附后)

2023年北京市朝阳区高考生物二模试卷1. 胞内体是动物细胞内的囊泡结构,能将细胞摄入的物质运往溶酶体降解。
下列推测不合理的是( )A. 胞内体可以将胞吞摄取的多肽运往溶酶体B. 胞内体与溶酶体融合体现膜的选择透过性C. 胞内体的膜由磷脂、蛋白质等分子构成D. 溶酶体水解产生的物质可被细胞再利用2. APC/C复合物促进有丝分裂进入后期,为研究其蛋白修饰对细胞周期调控的影响,利用药物对细胞进行同步化处理,测定洗去药物后不同时间的细胞周期时相,结果如图。
下列分析不正确的是( )A. G1、S、G2期为M期进行物质准备B. 药物使大部分细胞停滞在G1/S期C. APC/C缺失蛋白修饰会阻碍细胞周期顺利完成D. APC/C可能在后期将同源染色体拉向细胞两极3. ctDNA是肿瘤细胞死亡后释放到血液中的小片段核酸。
研究者采集18例肺癌患者手术前的ctDNA与手术切除的肿瘤组织DNA(tDNA)样本,检测到15人的ctDNA及18人的tDNA中存在肺癌相关基因。
以下说法错误的是( )A. 肿瘤细胞具有无限增殖、形态结构改变等特点B. 肺癌的发生与原癌基因,抑癌基因的突变有关C. 两类样本检测结果差异可能与tDNA易降解有关D. 研究结果提示ctDNA可作为肺癌筛查的参考指标4. 蜜蜂的蜂王(雌蜂)由受精卵发育而来,雄蜂由卵细胞发育而来,雄蜂精子的染色体数目与其体细胞相同。
控制体色和眼色的基因位于两对染色体上,杂交结果如表。
F1表型种类组合父本母本雌性雄性组合一褐体黑眼褐体黑眼1种4种组合二黑体黄眼褐体黑眼1种1种下列叙述正确的是( )A. 组合一中F1雌雄表型差异是伴性遗传的结果B. 组合一中F1雄性4种表型的比例接近1:1:1:1C. 组合二中的母本及F1的雄性均是杂合子D. 组合二中F1雌性与父本回交后代中黑体黄眼占5. 野生型马铃薯大多自交不亲和。
研究者培育DMP基因突变的马铃薯,开展如下杂交实验。
《基于转录组分析的华癸中慢生根瘤菌早期共生基因功能研究》

《基于转录组分析的华癸中慢生根瘤菌早期共生基因功能研究》一、引言华癸中慢生根瘤菌(Mesorhizobium huakui)是用于提高作物产量的重要生物菌剂之一。
它在植物体内形成的共生体系能有效固氮,提高作物对土壤中养分的吸收效率。
在根瘤菌与宿主植物相互作用的过程中,早期共生基因的表达和功能发挥着至关重要的作用。
近年来,随着转录组学技术的发展,越来越多的研究开始利用这一技术对根瘤菌的早期共生基因进行深入分析。
本文将通过转录组分析,探讨华癸中慢生根瘤菌早期共生基因的功能。
二、研究方法本研究采用转录组学技术,对华癸中慢生根瘤菌在共生过程中的早期阶段进行基因表达分析。
首先,我们收集了与宿主植物形成共生的华癸中慢生根瘤菌样品和未形成共生的样品。
随后,我们提取了这些样品的RNA,进行了转录组测序。
通过对测序数据的分析,我们可以得到不同条件下基因的表达情况,从而进一步分析早期共生基因的功能。
三、结果与分析1. 早期共生基因的筛选与鉴定通过对转录组数据的分析,我们筛选出了一批在共生过程中早期阶段表达差异显著的基因。
这些基因主要涉及细胞信号传导、代谢调控、细胞壁合成以及固氮酶基因等多个方面。
通过对这些基因的序列分析和功能预测,我们可以初步判断它们的生物功能和在共生过程中的作用。
2. 早期共生基因的表达模式分析通过对基因表达数据的进一步分析,我们得到了这些早期共生基因在共生过程中的表达模式。
这些模式主要包括在共生过程中的早期阶段显著上调或下调的基因、以及在不同环境条件下的表达变化等。
这些数据为我们进一步了解华癸中慢生根瘤菌的共生机制提供了重要的依据。
3. 关键早期共生基因的功能验证为了进一步验证我们筛选出的早期共生基因的功能,我们进行了基因敲除和过表达等实验。
通过这些实验,我们发现某些基因的敲除或过表达会影响根瘤菌与宿主植物的共生效果,从而验证了这些基因在共生过程中的重要性。
四、讨论与结论本研究通过转录组分析,深入探讨了华癸中慢生根瘤菌早期共生基因的功能。
【国家自然科学基金】_固氮基因_基金支持热词逐年推荐_【万方软件创新助手】_20140802
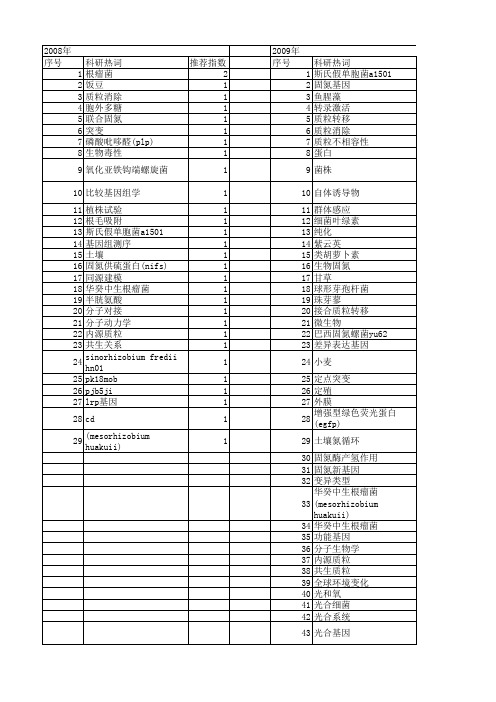
科研热词 推荐指数 鉴定 2 转bt水稻 2 联合固氮菌 2 筛选 2 甘蔗 2 水平基因转移 2 根际细菌 2 多样性 2 固氮细菌 2 阈值 1 近等位基因大豆 1 蛋白 1 结瘤 1 系统发育树 1 玉米 1 沙棘根际 1 氨单加氧酶基因(amoa) 1 氢氧化细菌 1 气候 1 模拟微重力 1 根瘤 1 放氢 1 微生物多样性 1 差异表达 1 小多肽 1 大豆 1 大肠杆菌 1 基因表达 1 基因克隆 1 基因 1 土壤类型 1 固氮 1 回转器 1 喀斯特 1 变形菌 1 发菜 1 原核表酸盐还原酶基因(nirs) 1 亚硝酸盐氧化还原酶基因(nxra) 1 wip 1 transgenic bt rice 1 sugarcane 1 screening 1 rt-pcr 1 rhizobacteria 1 pcr-dgge 1 nitrogen fixing bacteria 1
推荐指数 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
2011年 科研热词 推荐指数 发菜 2 双向电泳 2 原核表达 2 鱼腥藻pcc7120 1 重金属 1 过氧化物氧还蛋白 1 褐沙蒿 1 藻蓝蛋白 1 菌群结构 1 聚合酶链式反应-变性梯度凝胶电泳 1 耐盐植物 1 耐旱 1 系统发育分析 1 生物多样性 1 生物固氮 1 点形念珠藻atcc29133 1 检测技术 1 根际细菌 1 异形胞 1 差异表达 1 富营养化 1 实时荧光定量pcr 1 官厅水库 1 多样性 1 基因克降 1 固氮酶(nifh)基因 1 呼伦贝尔草原 1 厚壁孢子 1 内生细菌 1 克隆 1 促生 1 亚基基因 1 nifh基因 1 16s rdna 1
环境工程微生物学第三版周群英课后题

环境工程微生物学第三版周群英课后题答:(1)鞭毛纲:在污水生物处理中系统中,活性污泥培养初期或在处理成效差时鞭毛虫大量显现,可作污水处理的指示生物。
(2)肉足纲:变形虫喜α-中污带或β-中污带的自然水体中生活。
在污水生物处理中系统中,活性污泥培养中期有显现。
(3)纤毛纲:纤毛纲中的游泳型纤毛虫多数是在α-中污带或β-中污带,少数在寡污带中生活。
在污水生物处理中系统中,活性污泥培养中期或在处理成效差时纤毛虫会显现。
2. 如何判定某水样是否被粪便污染?答:假如水样中检测出有大肠杆菌群,则认为该水样被粪便污染。
3. 微生物呼吸作用的本质是什么?可分为哪几种类型?各类型有什么特点?答:微生物呼吸作用的本质是氧化与还原的统一过程,这过程中有能量的产生和转移。
微生物呼吸作用的可分为发酵、好氧呼吸和无氧呼吸。
第五章微生物的生长繁育与生存因子1.什么叫灭菌?灭菌方法有哪几种?试述其优缺点。
答:灭菌是通过超高温或其他的物理、化学因素将所有的微生物的营养细胞和所有的芽孢或孢子全部杀死。
灭菌的方法有干热灭菌法和湿热灭菌法。
湿热灭菌法比干热灭菌法优越,因为湿热的穿透力和热传导都比干热的强,湿热时微生物吸取高温水分,菌体蛋白易凝固变性,因此灭菌成效好。
2.什么叫消毒?加热消毒的方法有哪几种?答:消毒是用物理、化学因素杀死致病菌,或是杀死所有微生物的营养细胞或一部分芽孢。
方法有巴斯德消毒法和煮沸消毒法两种。
3.细菌、放线菌、酵母菌、霉菌、藻类和原生动物等的正常生长繁育分别要求什么样的pH?答:大多数细菌、藻类和原生动物的最适宜pH为6.5~7.5,它们的pH适应范畴在4~10之间。
放线菌为7.5~8.0。
酵母菌和霉菌在3~6。
第六章微生物的遗传和变异1. 什么是微生物的遗传性和变异性?遗传和变异的物质基础是什么?如何得以证明?答:微生物将其生长发育所需要的营养类型和环境条件,以及对这些营养和外界条件产生的一定反应,或显现的一定性状传给后代,并相对稳固的一代一代传下去。
慢生型大豆根瘤菌(Bradyrhizobium

慢生型大豆根瘤菌(Bradyrhizobium应用与环境生物2003,9(3):307~312ChinJApplEnvironBiol=ISSN1006-687X2oo3.-06-25慢生型大豆根瘤菌(Bradyrhizobiumjaponicum)聚羟丁酸合成酶基因(phbC)突变体的构建及其特性戴美学武波柏学亮张成冈0马庆生CHARLESTrev0rC(山东师范大学生命科学学院济南250014)(广西大学农业分子遗传农业部重点实验室南宁530005)(中国科学院沈阳应用生态研究所沈阳110015)(滑铁卢大学生物系加拿大滑铁卢N2L3G1)摘要通过转座子Tn5诱变和同源重组,构建了BradyrhizobiumjaponicumUSDA110聚羟丁酸合成酶基因(phbC)突变体.序列测定确定了转座子插入的精确位置,所获得的4个转座子诱变的质粒其Tn5插在6c基因内两个相距仅9bp的位点.被Southern和PCR证实的突变体菌株仍能产生相当于野生型菌株12.97%~25.10%的PHB,并且在突变体和野生型菌株总DNA杂交图上都呈现出一条约5kb的阳性带,推测在B.japonicum基因组中存在不止一个聚羟丁酸合成酶基因.图3表4参17关键词慢生型大豆根瘤菌;聚羟丁酸合成酶基因;聚羟丁酸;转座子诱变;Tn5CLCQ78:Q939.114 CoNSTRUCTIoNANDCHARACTERISTICSoFPoL Y.3. HYDRoXYBUTYRA TElSYNTHASEGENEMUTANTS1lBRADYRHIZoBIUMJAPoNICUMsDAQDAIMeixue,WUBo,BAIXueliang,ZHANGChenggang,MAQingsheng& amp;CHARLESTrevorC(1ifeScienceCollege,ShandongNormalUnive~i@,Jinan250014,China) (KeyLab.ofAgriculturalMolecularGeneticsofChineseMin~tryofAgricult ure,GuangxiUniversity,Nanning530005,China); (ShenyangInstituteofAppliedEcology,ChineseAcademyofSciences,Shen yang110015,China)(BiologyDepartment,Unive~i@ofWaterloo,WaterlooN2L3G1,Canada) AbstractPoly一3一hydroxybutyratesynthasegene(phbC)mutantswereconstructedinBradyrhiz obiumjaponicumUSDA110throu【shtransposonTn5mutagenesisandhomologousrecombination.Theprecise locationsofTn5insertionsinphbCgenewere determinedbysequencingandtheresultsindicatedthatthe4Tn5insertionsac quiredwerelocalizedattwositeswithanintervalofonly9basepair.ThephbCmutantswhichhadbeenconfirmedbySouthernblotandPCRcouldstillproduce10.7%~27.3%PHBoftheamountproducedbywildstrain,andapositivebandabout5k bshoweduponthegenomicDNASouthern blotofbothwildstrainandphbCmutantstrains,inferringthatthereweremoret hanonepoly一3一hydroxybutyratesynthasegenes inBradyrhizobiumjaponwumgenome.Fig3,Tab4,Ref17 KeywordsBradyrhizobiumjaponwum;poly-3一hydroxybutyratesynthasegene;poly一3一hydroxybutyrate(PHB);transposonmutagenesis;Tn5CLCQ78:Q939.114在非碳素营养受限而碳源过剩时,很多细菌可在细胞内积累聚羟烷酸(poly---3hydroxyalkanoates,PHAs),当碳源不足时.这种内部储存的聚合物又可用作细菌的碳源和能源¨].PHAs因其具有热塑性和生物可降解性而倍受关注¨l3].聚羟丁酸(poly-3一hydroxybutyrate,PHB)因其物理特性适于开发广泛用途的生物可降解塑料而成为PHAs中研究最为广泛的一种.收稿日期:2002-04-04修回日期:2oo2.o8-2l通讯作者Correspondingauthor(E-mail:**********************;Tel*************)Bradyrhizobiumjaponicum作为慢生根瘤菌的典型代表,是研究共生固氮体系的模式菌种之一.B.japonicum在自生阶段和共生阶段均可积累PHB,尤其是在共生阶段的类菌体中可积累高达细胞干重50%的PHB;如此高含量的PHB的存在,应该具有其生理意义.有资料认为,积累于类菌体中的PHB可能对于当植物光合作用降低时为固氮过程提供能量, 或当根瘤衰老后固氮作用的延续是重要的],但迄今并无直接证据.克隆与PHB代谢相关的基因,构建各基因突变体, 有助于从分子生物学水平上研究和揭示PHB在丑japonicum应用与环境生物ChinJApplEnvironBiol9卷与大豆共生过程中的作用.PHB的代谢涉及多个基因,聚羟丁酸合成酶基因(phbC)编码聚羟丁酸合成酶,催化PHB合成代谢的最后一步,即由单体的3-羟丁酸聚合成聚羟丁酸.我们从鼠japonicum 基因文库中克隆了phbC基因,并完成了该基因全序列的测定(序列在GenBank的登记号为A Y077580).在此基础上,通过转座子诱变和同源重组构建了且japonicumphbC基因突变体, 并研究了该基因突变对菌株共生结瘤,固氮能力的影响.本文报道这些研究结果.1材料与方法1.1菌株,质粒和转座子本文所用菌株,质粒和转座子见表1.表1菌株,质粒及转座子Tab1Strains,plasmidsandtransposon3期戴美学等:慢生型大豆根瘤菌聚羟丁酸合成酶基因(6c)突变体的构建及其特性1.2培养基和培养条件所用培养基有LB,TY,YMB(gL~:K2HPO40.5;MgS04.70,0.2;NaC1,0.1;Mannitol,10;yeastextract,0.5),M9基本培养基,M9一AA[M9基本培养基中加入终浓度为15mmol/L的乙酰乙酸(Acetoacetate,AA)作为唯一碳源],MOPS(gL~:MOPS10.5,Mannitol10,谷氨酸钠3.2,pH7.4;灭菌后加入0.3ms/mEBiotin1.0mE,1mol/LMgSO41.0mE,0.5mol/L0.5mE).根瘤菌培养于30℃,大肠杆菌培养于30℃或37℃.如需要使用抗生素,除文中特别说明外,使用浓度为:Ampicillin (sodiumsalt),100mL;Tetracyclinehydrochloride,20g/mL;Kanamycinsulfate,20~g/mL;Spectinomycindihydrochlo—ride,50~g/mL;Chloramphenicol,10b~g/mL;Gentamicinsul—fate,10txec’mL;Streptomycin,100I.Lg/mL;Neomycinsulfate,100p,g/mL.1.3酶和试剂’限制性内切酶,连接酶为Promega或MBI产品;抗生素购自Sigma或BoehringerMannheim.1.4DNA制备质粒DNA的制备按Maniatis等人的方法进行.根瘤菌总DNA的制备按文献[16]方法进行.1.5杂交试验Southernblot杂交根据Roche推荐的方法进行;DNA探针采用DIGHi【ghPrimeKit进行DIG标记;杂交膜使用CDP—Star (RocheDiagnosticsGmBH),在nuorchem8000(Advanced Fluorescence,ChemiluminescenceandVisibleLightImaging;Mo—lecularDynamics)中进行检测.1.6DNA序列测定将要测序的片段亚克隆到载体pUC19,采用Amersham pharmaciabiotech的ThermoSequenaseTMCyTM5/Cy5.5末端标记的测序试剂盒在VisibleGeneticsOpenGeneLong—ReadTow—e测序仪上进行.所用引物M13Forward:5gtaaaacgacggc—cagt3(17mer);M13Reverse:5caggaaacagctatgac3(17mer);以及根据需要设计的引物(见文中).数据分析采用Macvector version6.0.1(OxfordMolecularGroup).1.7PHB测定被试菌株接种于YMB液体培养基,30℃,100r/rain培养3d(快生型根瘤菌)或5d(慢生型根瘤菌),离心收集菌体.灭菌0.85%NaC1,ddH2O各洗一遍,然后按Law&Slepecky描述的方法Ⅲ进行PHB测定.1.8植株试验植株试验所用容器是根据Magenta缸的原理自行设计组装的.由上下两容器组成,上面的容器为上口直径15cm的塑料盆,内装栽培基质,中心部位穿一条粗棉絮条以使上下两容器相连;下面为1000mL的玻璃烧杯,内盛营养液.栽培基质为50%细沙和50%蛭石.每盆加入400mLJensen氏无氮营养液,锡箔覆盖,121oC灭菌1h,冷却后移去锡箔,植入3日龄大豆芽苗(供试大豆品种为”Bayfield”),每处理3盆,每盆3株? 植苗后第二天,接入10mL浓度为10/mL的菌液稀释液(YMB30oC.200r/min培养45d);空白对照组接入同样体积的灭菌ddHO.供试植株培养于自动调温,调湿,调光的中型培养室内;16h,25℃,光照;8h,20oC,黑暗;光照强度420txEmSm一.根据需要向培养容器中补加灭菌的ddH:O,植株试验周期为6wk.2结果2.1phbC基因转座子Tn5诱变pDC2是一个从BradyrhizobiumjaponicumUSDA110基因文库中克隆到的能与oc突变体菌株Rml1144功能互补的重组粘粒,载体pLAFR1所携带的16kb外源片段含有B.japoni? clzm的完整的phbC基因.首先,DH5a/pDC2与MT614(染色体上带有转座子Tn5)在MT616(MT607/pRK600)辅助下进行接合,接合后菌液经适当稀释后涂布于含Km和Tc的LB平板上进行筛选.如此将带有6c基因的重组粘粒pDC2引入染色体上带有转座子Tn5的MT614菌株(MT614/pDC2).其次,MT614/pDC2,MT609,MT616进行第二步三亲接合,在含Km,Tc和sp的LB平板上筛选.如此将KmTc的质粒(即pDC2上插有Tn5的质粒)导人具有Sp抗性的MT609菌株.共获得600多个菌落.然后,进行第三次接合以筛选Tn5插入6c基因内部的克隆.随机挑取100个LB+KmTcSp平板上的菌落,分别与苜蓿根瘤菌Rml021菌株的phaC突变体Rml1144菌株在辅助菌株MT616的作用下进行接合;涂布于TY+Sm.00Nm.00平板上30℃培养3d.结果所有100个接合组平板上都获得相应的菌落.从各接合组平板上各挑少数几个菌落在TY十Sm.. Nm..平板上划线纯化两次.最后,对这100个纯化的接合子进行一系列的测定,以筛选Tn5插在phbC阅读框架内的克隆.将此100个接合子分别接种于TY(3d),M9一AA(5d),MOPS(4d)平板上30℃培养,并于同平板上接种野生型菌株Rml021和phaC突变体菌株Rml1144作为对照.结果见表2,所有被试菌株均可在TY上生长;96个菌株在M9一AA上培养5d可以形成明显菌落,类似于野生型菌株Rml021;而4个菌株于同样条件同样时间内不能形成明显的菌落,情况类似于phaC突变体菌株Rml1144.这4个菌株在MOPS平板上也均不能形成粘液型菌落.最后,4 菌株分别用YMB液体培养基培养(30oC,100r/min,3d),收集菌体,测定PHB;结果(表2)4菌株培养物PHB均为阴性.经过上述一系列测定,我们确认这4个菌株所携带的粘粒,均于6c基因阅读框架内插入了Tn5,分别命名为pDC11 (pDC2phbCl~l::Tn5),pDC12(pDC2phbCFt2::Tn5),pDC13 (pDC2phbCl~3::Tn5)和pDC14(pDC2phbCDA::Tn5).3lO应用与环境生物ChinJApplEnvironBiol9卷表2生长性状及PHB测定Tab2GrowthcharacteristicsandPHBassay菌株1’YM95.AStrain3ddAM40PsdPHB5通过三亲接合分别将pDC11,pDC12,pDC13,pDC14从菌株Rml1144转入DH5ct;提取质粒DNA,试用不同限制性内切酶酶切并与pDC2进行比较,发现经SacI酶切后所有4个质粒都没有pDC2所具有的4.5kbSacI片段,代之出现一约10.3 kbSacI片段,其大小恰好相当于4.5kbSacI片段加上5.8kbTn5片段(图1).这表明转座子Tn5恰好插入到4.5kbSacI片段E.MarkerpDC2pDCllpDC12pDC13pDC1410.3kb图lpDC2及其Tn5插入质粒SacI酶切结果的比较FiglComparisonoftheSacIdigestionresultsfrompDC2anditsTn5insertions2.2转座子插入点的定位我们已测定了phbC基因的全序列(该序列在GenBank的登记号为A Y077580).为对Tn5插入点进行精确定位,首先用BamH1分别酶切pDC11,pDC12,pDC13,pDC14,将含有部分phbC基因序列和部分Tn5序列(只有一个IS50)的片段克隆到载体pUC19上,分别得pMX110,pMX112,pMX109,pMX113. 以根据Tn5IS50序列自行设计并合成的引物TrLSR(5 GCCCAGTCGGCCGCACGA TGAAGAGCA327met:匹配TI15 的129—102&5690—5716bp区域)对pMX110,pMX112,pMX109,pMX113分别测序.所得序列采用软件DNAStar同p c基因序列进行分析比较.分析结果表明,pDC11和pDC13的Tn5插在同一位点,即phbC基因的654—655碱基之间;pDC12,pDC14的Tn5位于同一位点,即6C基因的645—646碱基之间;两插入位点仅相距9个碱基(图2). TTGTGACGGAACCAAGAAAA TI~GTCGCAGTGCAACACA TCIT CGAGAGTGTI~AACGTC—primerMX7AAACA TCrCGAGCTTCGCGGTGACGAA TACGrGAA TGAA TCGTF ACTGTFCTGCTCA TGAGCACGACCGACACGCCCAAACCCGACACGAAGTrCAA TGCG GAAGCCITCCrCGA TG61AA TGTCCrCCrCGGGCGA TGGAGAGCGGCGGCAAGGC(T0GC CCrCCT]’CCTCAAGCCGCGC21GAGAGCGC~GAGGTGCAGGACCGTCCGCCGGCCGAGCTrG CCGAAGTCA TCAAGGCCTrC481G1CCAGCA001℃ACC从C(GCTGTC~C’I~CGAACITCGTGCTGACCAA TCCGGAAGTG.∞∞.∞..∞.∞.∞∞∞....601GAGGACA TCGCGCrCCGGCAAGGGCA TGCTGAAGA TCCCrC CAGTC~ACCCC,~AIIA TCTC645—646654-655——66lG1℃GTCGGcGTCAACAr构建是在完成phbC序列测定前进行的.因为所得的4个破坏了phbC功能的Tn5诱变的质粒pDC11,pDC12,pDC13,pDC14经酶切比较发现,rn5都位于pDC2的4.5kbSacI片段,从而形成10.3kbSacI片段.因此我们便克隆这个10.3kbSacI片段用于突变体的构建.又因pDC11和pDC13上Tn5插入点完全相同,pDC12和pDC14上rn5插入点完全相同,所以这里只描述以pDC13和pDC14进行突变体构建的过程.pJQ200(SK)是QuandtJ和HynesMF14j构建的一种自杀质粒,大小约5kb,具Gm抗性,带有编码蔗糖多聚酶的sacB基因,当携带该质粒的细胞培养于含有蔗糖的培养基上时,蔗糖多聚酶可将蔗糖聚合为多聚蔗糖,从而导致细胞死亡.利用该质粒自杀特性构建突变体,为突变体的筛选提供了方便.分别将pDC13,pDC14的10.3kbSacI片段克隆到pJQ2o0(sK)的Sacl位点上分别得pDC39和pDCA1.通过转化将pDC39,pDC41引入菌株S17—1(具有tra基因,接合时不需使用辅助菌株),然后分别与野生菌株.一pon&umUSDA110进行接合,接合后的菌液涂于YMA+Cm. Km平板上30℃培养6d.在各接合组平板上共获得608—822个菌落.将上述接合组平板上所获得的菌落点接于TY+5%蔗糖+Cm.Km20的平板上测定生长情况;结果4.8%一10.2%可于含5%蔗糖的培养基上生长.再将上述可在5%蔗糖培养基上生长的菌落划线接种于成对的rY和TY+Gm,.平板上,测定对Gm的敏感性.结果约69%~81%的被测菌对Gm敏感.这样最终获得的具CmKin抗性,可在含有蔗糖的培养基上生长,而又对Gm敏感的菌株被认为是6c突变体菌株,分别命名为Bj30062,Bj30063.在ITY平板上各突变体菌株生长速度明显低于野生型菌株(同样培养时间内形成较小菌落),而在YMA平板上突变体3期戴美学等:慢生型大豆根瘤菌聚羟丁酸合成酶基因(6c)突变体的构建及其特性3ll与野生菌株的生长速度没见明显差异.2.4突变体的证实2.4.1Southern杂交分别提取Bj30062,Bj30063及Bjus—DA110总DNA,经EcoRI酶切,电泳;转膜后,用DIG标记的pMX5的2.3kbnI—SacI片段作探针进行杂交,该片段包含BradyrhizobiumiaponicumUSDA110phbC基因的自起始密码子至1253bp片段及其上游约1.1kb片段,该片段内无oⅪ酶切位点.采用严谨方式洗膜.杂交结果见图3,野生菌株Bjus—DA110在预计的7.5Id)处呈阳性;而4个被试突变体菌株没有7.5kb的阳性带,而是出现一条13kb左右的阳性带.这可解释为13kb阳性带来自7.5kbEcoRI片段插入了Tn5.另外,值得注意的是,除了预料中的7.5kb和13kb阳性带外,在野生菌株和突变体菌株的杂交图谱中均出现一条大约5kb的阳性带.Mll0300623006323.1kb_一9.4kb_一6.5kb_一——●}.一擘一4.3k一’at-柚▲一图3Southern杂交图Fig3Southernblot用EcoRI酶切总DNA;以DIG标记的pMX5的2.3kbnI—SacI片段作探针,M,110,30062,30063分别代表Marker(k/H/ndn1),BjUS.DA110,Bj3oo62和Bj30063GenomicDNAweredigestedwithEcoRI. TheDIG—labeled2.3kbnI—SacIfragmentofpMX5wasusedas probe.M,110,30062and30063representsMarker(k/HindⅢ),Bjus-DA110,Bi30062andBj30063,respectively2.4.2PCR因所构建的突变体菌株经PHB测定,仍能产生一定量的PHB(后述),为进一步证实突变体,我们根据已完成的6c序列采用DNAStar设计了一对引物:MX7(5CG. GTGACGAA TACGGAA TGAA TCG324mer).MX8(5 AAGTCCACCTGTGCCGCGAAGAAC324mer).这对引物所匹配的序列见图2,其匹配区间的大小为1.16kb.使用这对引物,分别以野生型菌株BjUSDAllO和突变体菌株的总DNA为模板进行PCR.PCR条件:95℃2min;95oC30S,55oC30S,65℃10rain;30cycles;4℃hold.结果(图片未显示),以野生型菌株SjUSDA110总DNA为模板扩增出预想中的1.16kb片段;而以突变体菌株总DNA为模版均未扩增到此片段,但代之扩增出约7kb片段;这说明1.16kb片段上带有Tn5(5.818kb).PCR结果进一步证实了所构建的突变体.2.5突变体互补菌株的获得通过接合将重组粘粒pDC2分别引入突变体菌株,获得互补菌株Bj30062/pDC2和Bj30063/pDC2.2.6PHB测定测定结果见表3.突变体菌株经YMB培养,仍可测得占细胞干重3.06%一5.92%的PHB,相当于野生菌株所产PHB量的12.97%~25.10%.说明phbC基因的突变影响了PHB合成的数量,但并未使菌株彻底丧失合成PHB的能力.将携带phbC基因的重组粘粒导入突变体细胞后,虽然使突变体产生PHB的能力有明显提高(提高至野生型菌株PHB 量的67.06%一67.95%),但并未使突变体完全恢复至野生型的水平.表3PHB测定结果Tab3ResultsofPHBassay2.7植株实验植株实验结果见表4.被测突变体菌株Bj30062,Bj30063平均每株瘤数和平均单株瘤重均高于野生型菌株;但平均单瘤重及植株平均干重同野生型菌株相比,未显示明显差异.3讨论通过转座子Tn5诱变获得了4个破坏了粘粒pDC2互补Rml1144(phaC::Tn5-233)能力的诱变质粒.序列测定结果显示phbCfH::Tn5和phbC~3::Tn5转座子插在同一位点,即phbC基因的654—655碱基间;phbCO2::Tn5和phbC~4::Tn5 转座子也插在同一位点,即645—646碱基之间;且两插入位点非常靠近,仅相距9个碱基.这是否意味着该区域的序列具有特殊性,有待进一步分析.要从分子遗传学角度研究积累于慢生大豆根瘤菌自由生长细胞及类菌体中的PHB在慢生大豆根瘤菌与大豆共生过程中的可能作用及其机理,首先必须构建PHB代谢相关基因系列突变体.本文成功地构建了慢生大豆根瘤菌USDA110菌株聚羟丁酸合成酶基因突变体,为开展这方面的研究奠定了初步基础.Southern杂交和PCR结果证实了所构建的B.japonicum6c突变体,但PHB测定结果却显示突变体菌株仍能产生一定数量的PHB,这说明B.japonicum中可能存在不止一个聚羟丁酸合成酶基因.另外,值得提及的是,在总DNASouthern杂交图谱中,突变体菌株和野生型菌株都呈现出一条预料之外的约5kb的阳性带,这进一步印证了在B.japonicum中存在一个以上phbC基因的推断.通过搜索GenBank,已发现有两种细菌中存在两个phbC基因,即Pseudomonaspseudoalcaligene$ 和Pseudomonasaureofaciens.筛选克隆可能存在的另一(几) phbC基因,构建6c基因完全突变体,对于研究,揭示PHB在B.]aponicum共生过程中的作用具有重要意义.此外,植株3l2应用与环境生物ChinJApplEnvironBiol9卷表4植株实验结果试验发现,phbC突变体菌株平均单株瘤数和平均单株瘤重均高于野生型菌株,对这一现象的研究正在进行中.致谢本文部分工作于加拿大UniversityofWaterloo生物系TrevorC.Charles教授的实验室完成,衷心感谢Charles教授和该室王春霞博士的大力帮助.ReferenceslAndersonAJ,DawesEA.Occurrence,metabolism,metabolicrole,andindustrialufo,sofbacterialpolyhydroxyMkanoates.MicrobiolRev,l99o.54:450~4722LeeSY.Bacterialpolyhydruxyalkanoates.BiotechnolBioeng,1996,49: ll43DawesEA.SeniorPJ.Theroleandregulationofenergyreservepoly—mer$inmicroorganisms.AdvMicrobiolPhysiol,1973,10:135~2664WongPP,EvansHJ.Poly-B—hydroxybutyrateutilizationbysoybean nodulesandassessmentofitsrolesinmaintenanceofnitrogenaseactivi—ty.Plantphysml,1971,47:750~7555Be~ersenFJ,PeopleMB,TurnerGL.Aroleforpoly-B—hydroxybu—tyrateinbacteriodsofsoybeanrootnodules.ProcRSocSer.1991.B245:59~646GersonT,PatelJJ,WongMN.Theeffectsofage,darknessandnitrate onpoly一hydroxybutyratelevelsandnitrogen—fixingabilityofRhizobiuminLup/nusangust~olius.PhyswlPlant,1978,42:420~4247CharlesTC,CaiGQ,AnejaP.Megaplasmidandchromosomallocifor thePHBdegradationpathwayinRhizobium(Sinorhizobium)meliloti. Genetics,l997,146:121l~12208SimonR,PricerU,PuhlerA.Abroadhostrangemobilizationsystemf0rinvivogeneticengineering:transposonmutagenesisinGram—nega—tivebacteria.B/o/Technology,1983,1:784~7919FinanTM,KunkelB,DevosGF,SignerER.Secondsymbioticmega—plasmidinRhizobiummeliloticarryingexopolysaccharideandthiamine synthesisgenes.JBacteriol,1986,167:66~7210MeadeHM,LongSR,RuvkunGB,BrownSE,AusubelFM.Physi—calandgeneticcharacterizationofRhizobiummelilotiinducedbytrans—posonmutagenesis.JBacteriol,1982,149:114~122l1BergCM,esoftransposableelementsandmapsofknown insenions.CellularandMolecularBiologyWashingtonDC.American SocietyforMicrobiology,1987,1071~110912FriedmanAM,LongSR,BrownSE,BuikemaWJ,AusubelFM. Constructionofabroadhostrangecloningvectoranditsuseinthege—neticanalysisofRhizobiummutants.Gene,1982,18:289~29613Y anisch—PerronC.VieiraJ.MessingJ.ImprovedM13phagecloning vectorsandhoststrains:nucleotidesequencesoftheM13mpl8andpUC19vectors.Gene,1985,33:103~11914QuandtJ,HynesMF.V ersatilesuicidevectorswhichallowdirectse—lectionforgenereplacementinGram—negativebacteria.Gene,1993, 127:l5~2l15ManiatisT,FritechEF,SambrookT.MolecularCloning:ALaborato—ryManua1.ColdSpringHarborN.Y:ColdSpringHarborLaboratory,.198216AusubelFM,BrentR,KingstonRE,MooreDD,SeidmanJG,Smith JA,StruhlK.CurrentProtocolsinMolecularBiology.NewY ork: JohnWiley&Sons.199717LawJH,SlepeckyRH.Assayofpoly一hydroxybutyricacid.JBacte—riol,1961,82:33~36。
2021年河南师范大学微生物学专业《微生物学》期末试卷A(有答案)

2021年河南师范大学微生物学专业《微生物学》期末试卷A(有答案)一、填空题1、细菌表面常有毛状附属物,其中用于运动的称为______,帮助细菌附着到物质表面的称为______,在不同菌株间传递遗传物质的称为______。
2、病毒在寄主体外,很易因外界环境如高温,射线等作用而______;带封套的病毒容易被______溶剂破坏,因而可用消毒剂如______来消毒。
3、光能自养微生物有产氧与不产氧两大类,前者如______等,后者如______等。
4、从化合物水平来看,微生物的氮源主要有______、______、______、______、______、______和______等。
5、真核生物的细胞核由______、______、______和______等构成。
6、在微生物学历史上,固体培养基的发明人是______,用于固体培养基的优良凝固剂琼脂的发明人是______。
7、微生物的细胞分裂有两种模式:______和______。
8、植物根际微生物对植物有益的方面有______、______、______和 ______等。
9、DNA分子中一种嘧啶被另一种嘌呤取代称为______。
10、免疫T细胞分为许多亚群,能协助特异性免疫反应的辅助T细胞是______,TS细胞叫______细胞,TM细胞又称______细胞。
二、判断题11、大肠杆菌和枯草芽孢杆菌属于单细胞生物,唾液链球菌和金黄色葡萄球菌属于多细胞生物。
()12、所有碳源物质既可以为微生物生长提供碳素来源,也可以提供能源。
()13、细菌产生酒精,只有通过ED途径才能达到。
()14、噬菌体核酸既有单链DNA、双链DNA,又有单链RNA、双链RNA。
()15、霉菌、酵母菌均是没有分类学意义的普通名称。
()16、在微生物的形态特征十分丰富的条件下,菌种鉴定就可完全依据这些特征来进行,例如真菌、放线菌和酵母菌等。
()17、应用大型发酵罐进行深层液体通气搅拌培养,是现代好氧菌发酵工业最典型的培养技术。
根瘤菌群体感应系统在共生结瘤过程中的功能

根瘤菌群体感应系统在共生结瘤过程中的功能丑敏霞;魏新元【摘要】根瘤菌能够合成群体感应(quorum sensing,QS)信号分子——酰基高丝氨酸内酯(N-acyl homoserine lactone,AHL)类化合物,其在与豆科植物共生结瘤的过程中,以群体密度依赖的方式通过AHLs诱导相关基因的表达.目前,根瘤菌QS 系统的研究结果主要来自根瘤菌属和中华根瘤菌属.不同根瘤菌中的QS系统明显不同,它们对共生过程的影响有很大区别.同时,寄主植物也能通过分泌AHLs类似物等方式介导根瘤菌QS系统的表达.本文在简要介绍革兰氏阴性菌群体感应系统的基础上,综述了不同根瘤菌群体感应系统的类型及其对共生结瘤的影响,以及豆科植物对群体感应系统的响应等方面的研究进展,并对当前根瘤菌与豆科植物共生互作早期QS系统功能研究中存在的问题及今后的研究方向进行了分析与展望.【期刊名称】《西北农林科技大学学报(自然科学版)》【年(卷),期】2010(038)010【总页数】7页(P171-177)【关键词】根瘤菌;群体感应;共生互作;基因表达;豆科植物【作者】丑敏霞;魏新元【作者单位】西北农林科技大学生命科学学院,陕西,杨凌,712100;西北农林科技大学食品科学与工程学院,陕西,杨凌,712100【正文语种】中文【中图分类】Q935细菌之间通过感知周围环境中同类细胞密度变化而调节基因表达的现象,称为群体感应(quorum sensing,QS)。
QS是目前已知细菌间通讯的主要方式之一。
菌群在生长过程中,不断产生被称为自身诱导物(autoinducer,AI)的信号分子,并将其分泌到周围环境中,然后通过感应AI分子的浓度,监测周围细菌的数量,再根据菌群密度的波动调节基因表达。
革兰氏阴性细菌的信号分子大多是酰基高丝氨酸内酯(acyl-homoserine lactone,AHL)类衍生物,其能够自由通过细胞膜,并且在胞外环境中积累。
土壤微生物多样性实验研究方法概述_1

土 壤 (Soils), 2004, 36 (4): 346~350土壤微生物多样性实验研究方法概述 章家恩 蔡燕飞 高爱霞 朱丽霞 (华南农业大学热带亚热带生态研究所广州 510642)摘 要对有关土壤微生物多样性,包括微生物类群多样性、群落结构多样性、功能多样性以及基因多样性等的描述与表征方法进行了探讨,同时,对当前国内外土壤微生物多样性的一些实验研究方法,包括土壤微生物分离培养方法、Biolog微平板方法、FAME分析方法和分子生物学方法等进行了介绍和评述。
并指出在土壤微生物多样性研究中,如果可能的话,需要将各种方法结合起来使用,方可得到有关土壤生物多样性的较为全面的信息和理解。
关键词土壤微生物;生物多样性;FAME;Biolog;PCR中图分类号 Q938.1;S154.36生物多样性是当今国际上共同关注的问题,它也因此日益成为学术界的热点研究领域之一。
目前在地上部植物和动物的多样性方面开展了大量的研究,但对土壤生物多样性,特别是在土壤微生物多样性方面的研究仍较薄弱。
其中一个重要的原因就是在土壤微生物多样性的研究方法方面还存在着较大的困难和缺陷,很多方法还不能全面地、准确地研究土壤微生物多样性状况。
为此,本文对当前国内外土壤微生物多样性研究方法进行概述,旨在为相关研究提供一些参考。
1 土壤微生物多样性的表示方法 生态学意义上的“生物多样性”是指生物与环境形成的生态复合体以及与此相关的各种生态过程的总和,一般包括遗传多样性、物种多样性和生态系统多样性3个层次[1]。
生物多样性通常用“多样性指数”来表示。
生物(物种)多样性指数的计算一般需要知道群落中物种的种数以及各物种的个体数等相关的信息。
对于土壤微生物,由于方法上的限制和微生物自身的变异性,目前还不可能将土壤中的全部微生物培养出来,而且对于土壤微生物种属的鉴定分类也是一件不易的工作。
因此,在实际研究中,通常从某一侧面或某一角度来近似或间接地描述土壤微生物的多样性状况。
基因水平转移在根瘤菌进化中的研究进展

基因水平转移在根瘤菌进化中的研究进展陈雪莲; 江高飞; 钟增涛【期刊名称】《《生物技术通报》》【年(卷),期】2019(035)010【总页数】7页(P18-24)【关键词】基因水平转移; 根瘤菌; 共生固氮; 适应性进化【作者】陈雪莲; 江高飞; 钟增涛【作者单位】南京农业大学生命科学学院南京 210095【正文语种】中文根瘤菌是一类革兰氏阴性土壤细菌,能够与大多数豆科植物共生固氮。
根瘤菌与植物共生形成根瘤或茎瘤,在根瘤内分化成类菌体后能够将空气中的氮气通过生物固氮作用转化为铵。
植物进一步将铵转化为氨基酸等氮源物质,满足其生长繁殖需要[1]。
豆科植物是植物界最大的一个科,约有650属19 700种,大多数豆科植物可以与根瘤菌建立共生固氮体系[2]。
由于根瘤菌的宿主专一性较强,如何扩大根瘤菌在豆科植株上共生固氮的宿主范围,最终实现在非豆科植物上的共生固氮是根瘤菌研究的最终目标[3],也是生物固氮研究的目的之一。
1 基因水平转移(HGT)原核生物具有从环境或其他生物体基因组获取部分DNA的独特能力,并在称为基因水平转移(Horizontal gene transfer,HGT)的过程中将其整合到自身基因组中[4]。
基因水平转移是相对垂直进化(通过亲代传给子代)而言的,可以让细菌快速获得新的遗传性状从而促进细菌的进化。
基因水平转移在微生物进化过程中发挥着重要的作用,它打破了遗传依赖亲缘关系的界限,使遗传物质能够在不同的物种之间进行交换,原核生物通过基因水平转移的方式可提高对环境的适应性,并具备遗传的多样性[5-6]。
通过这种方式,微生物可以获得新的基因型和表型,例如毒力、抗生素抗性、代谢和共生等[6]。
基因水平转移的主要类型分别是:(1)转化:从环境中吸收外源DNA的能力;(2)转导:通过噬菌体介导将DNA从一个细胞转移到另一个细胞;(3)接合:通过质粒或者接合转移元件将DNA从供体细胞向受体细胞进行接触型转移。
中慢生根瘤菌Z1-4在制备细菌胞外多糖中的应用[发明专利]
![中慢生根瘤菌Z1-4在制备细菌胞外多糖中的应用[发明专利]](https://img.taocdn.com/s3/m/e42056ca192e45361066f5f5.png)
专利名称:中慢生根瘤菌Z1-4在制备细菌胞外多糖中的应用专利类型:发明专利
发明人:杨希,杨桥,李家洲,吕晓慧,张厂,邓俊劲
申请号:CN202110077157.4
申请日:20210120
公开号:CN112646751A
公开日:
20210413
专利内容由知识产权出版社提供
摘要:本发明属于微生物多糖制备技术领域,具体涉及中慢生根瘤菌Z1‑4在制备细菌胞外多糖中的应用,为发掘新的EPS高效合成菌,本发明从海洋藻际资源中,筛选得到一株中慢生根瘤菌属(Mesorhizobium sp.)的菌株,定名为Mesorhizobium alexandrii Z1‑4,实验证明,该菌株具有合成胞外多糖(EPS)的能力,有望应用于制备细菌胞外多糖,具有作为EPS工业生产菌株的潜在价值。
申请人:广东省农业科学院动物科学研究所
地址:510000 广东省广州市天河区五山
国籍:CN
代理机构:深圳市创富知识产权代理有限公司
代理人:叶灿才
更多信息请下载全文后查看。
中慢生型天山根瘤菌MsiR组成型蛋白突变株的筛选

中慢生型天山根瘤菌MsiR组成型蛋白突变株的筛选赵亚琪;马腾;蔡韬【摘要】MsiR蛋白来源于中慢生型天山根瘤菌(Mesorhizobium tianshanense),是LysR转录调控蛋白家族中的一员,它可以响应宿主植物释放的抗代谢物-刀豆氨酸,激活外运蛋白msiA编码基因的转录表达.通过构建MsiR突变文库,筛选得到了7个MsiR组成型突变蛋白,L166P、A147V、P83L、A278T组成型突变蛋白丧失了对刀豆氨酸的响应,在没有刀豆氨酸时的荧光值为800左右;A147T、E59G组成型突变蛋白仍然可以响应刀豆氨酸的诱导信号,添加刀豆氨酸时荧光值是未添加时的1.7倍.通过蛋白同源建模分析了组成型突变的氨基酸残基在MsiR蛋白上的空间分布及其对MsiR蛋白调控功能可能的影响.组成型突变蛋白的研究对进一步揭示LysR家族蛋白转录调控的机理有重要意义.【期刊名称】《中国酿造》【年(卷),期】2016(035)010【总页数】5页(P130-134)【关键词】LysR家族转录调控蛋白;MsiR蛋白;组成型突变;mCherry报告基因【作者】赵亚琪;马腾;蔡韬【作者单位】天津科技大学生物工程学院,天津300222;中国科学院天津工业生物技术研究所,天津300308;天津科技大学生物工程学院,天津300222;中国科学院天津工业生物技术研究所,天津300308;中国科学院天津工业生物技术研究所,天津300308【正文语种】中文【中图分类】Q819中慢生型天山根瘤菌(Mesorhizobium tianshanense CCBAU3306)是1995年定义命名的新菌种,分离于新疆土壤,可在刺果甘草(Glycyrrlazeuralensis)的根部形成共生根瘤并进行固氮[1];刀豆氨酸最早在1929年由日本科学家发现,是精氨酸的结构类似物,因此会被生物体误掺入正在合成的多肽链中,形成没有活性的蛋白质,最终导致生物体死亡。
中慢生型天山根瘤菌中自体诱导物分子结构的鉴定

中慢生型天山根瘤菌中自体诱导物分子结构的鉴定赵家星;杨梦华;曹慧娟;周蕾;杨瑞馥;钟增涛;朱军【期刊名称】《土壤》【年(卷),期】2010(042)001【摘要】通过用乙酸乙脂分别对天山根瘤菌 CCBAU3306 培养物上清液中自体诱导物分子进行抽提、浓缩,并采用液质联用对自体诱导物分子进行结构分析,最终获得该根瘤菌所产生酰基高丝氨酸内酯类信号分子的结构,并采用纯品进行活性的比较,发现调控蛋白与纯品结合后具有生物活性,能与目的 DNA 片断结合,证明了该结构的准确性.【总页数】4页(P148-151)【作者】赵家星;杨梦华;曹慧娟;周蕾;杨瑞馥;钟增涛;朱军【作者单位】南京农业大学生命科学学院,南京,210095;南京农业大学生命科学学院,南京,210095;南京农业大学生命科学学院,南京,210095;中国军事医学科学院微生物与流行病所,北京,100071;中国军事医学科学院微生物与流行病所,北京,100071;南京农业大学生命科学学院,南京,210095;南京农业大学生命科学学院,南京,210095【正文语种】中文【中图分类】Q933【相关文献】1.中慢生型天山根瘤菌CCBAU060A自体诱导物合酶基因的筛选及其在大肠杆菌中的表达 [J], 曹慧娟;郑会明;杨梦华;钟增涛;朱军2.中慢生型华癸根瘤菌Mesorhizobium huakuii AS9自体诱导物合成酶基因的筛选及其功能分析 [J], 袁全;杨梦华;郑会明;钟增涛;朱军3.中慢生型天山根瘤菌胞外多糖相关基因exo5及其在根毛吸附和生物膜形成中作用的研究 [J], 周晶;王鹏;钟增涛;朱军4.中慢生型天山根瘤菌中MrtR蛋白的融合表达研究及鉴定 [J], 戴俊;郑会明;杨梦华;钟增涛;朱军5.利用高效检测菌株对中慢生根瘤菌及红壤中自体诱导物的检测 [J], 钟增涛;周晶;李路;高轶静;朱军因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
土 壤(Soils), 2010, 42 (1): 148~151中慢生型天山根瘤菌中自体诱导物分子结构的鉴定①赵家星1, 杨梦华1, 曹慧娟1, 周 蕾2, 杨瑞馥2, 钟增涛1*, 朱 军1 (1南京农业大学生命科学学院,南京 210095;2中国军事医学科学院微生物与流行病所,北京 100071)Identification of Acyl-Homoserine Lactone Autoinducer Molecules in Mesorhizobium TianshanenseZHAO Jia-xing1, YANG Meng-hua1, CAO Hui-juan1, ZHOU Lei2, YANG Rui-fu2, ZHONG Zeng-tao1, ZHU Jun1(1 College of Life Science, Nanjing Agricultural University, Nanjing 210095, China;2 Institute of Microbiology and Epidemiology, Academy of Military Medical Sciences, Beijing 100071, China)摘 要:通过用乙酸乙脂分别对天山根瘤菌 CCBAU3306 培养物上清液中自体诱导物分子进行抽提、浓缩,并采用液质联用对自体诱导物分子进行结构分析,最终获得该根瘤菌所产生酰基高丝氨酸内酯类信号分子的结构,并采用纯品进行活性的比较,发现调控蛋白与纯品结合后具有生物活性,能与目的DNA片断结合,证明了该结构的准确性。
关键词:群体感应;中慢生型天山根瘤菌;液质联用;结构测定中图分类号: Q933许多与植物共生的细菌都是通过一种依赖于细胞密度的方式来调控特定基因的表达,这种调控方式被称之为群体感应(quorum sensing)[1]。
酰基高丝氨酸内酯类化合物(acylated homoserine lactone, AHL)作为革兰氏阴性细菌群体感应调控中普遍存在的信号分子[2],其在细胞中的浓度随着细胞密度的增加而增加,当积累到阈值浓度时就能够诱导特定基因的表达[3]。
目前已经鉴定出的AHL合成酶有3种类型,分别是LuxI,LuxM以及HtdS[4]。
AHLs由一个可变的乙酰基链尾部与一个稳定的高丝氨酸内酯头部相连,乙酰基链长度为4~18个碳,其第三个碳上的氢常被羟氧基或氧基取代,另一个改变是在乙酰链上存在双键。
目前发现最长的乙酰基链是由苜蓿中华根瘤菌合成的带18个碳的AHLs[5]。
氮是植物生长所需的重要营养物质,而根瘤菌能在豆科植物根部形成根瘤,或者定殖于部分非豆科植物根部,从而固定空气中的分子氮,与植物建立互惠的共生关系[6-7]。
根瘤的形成是一个复杂的信号交换过程,群体感应参与调节共生固氮相关基因的表达[8],因而发挥着重要的作用,但是目前对于中慢生型根瘤菌群体感应的研究报导却很少。
本实验室前期实验对不同中慢生型天山根瘤菌群体感应中自体诱导物活性和种类进行研究,发现差异较大,为进一步阐明同属不同种根瘤菌间群体感应系统的多样性,本研究通过采用液质联用的方法,以酰基高丝氨酸内酯环的特征碎片峰m/z102对高效液相分离出组份中的酰基高丝氨酸内酯类信号分子进行二维质谱分析,根据不同类型酰基高丝氨酸内酯类化合物的结构特点及[M+H]+推算出各种信号分子的结构,并利用纯品通过凝胶阻滞分析证明了该结构的生物学功能,研究为进一步阐明自体诱导物合成酶多样性与信号分子多样性间相互关系及不同菌株间信号系统间分子对话具有重要意义。
1 材料与方法1.1 材料1.1.1 菌株、质粒及生长条件表1为本试验所使用表 1 供试菌株菌株特性来源天山根瘤菌 CCBAU3306野生型中国农业大学JZA1 自体诱导物检测菌株本实验室保存R10(pCF218) 阳性对照菌株本实验室保存①基金项目:国家自然科学基金项目(30870065, 30800013)资助。
*通讯作者(ztzhong@)作者简介:赵家星(1984—),男,河南郑州人,硕士研究生,主要从事根瘤菌群体感应研究。
E-mail: 2006116084@第1期 赵家星等:中慢生型天山根瘤菌中自体诱导物分子结构的鉴定 149的菌株特性。
根瘤菌在 TY 培养基中 28℃ 培养,JZA1、R10 分别在 LB 和 A T 培养基中 28℃ 培养。
1.1.2 试剂和仪器 邻硝基苯-β-D-半乳吡喃糖苷(O-Nitrophenyln-β-D- Galactopyranoside ,ONPG ),异丙基-β-硫代半乳糖苷(Isopropyl-β-Dthiogalactosid ,IPTG ),5-溴-4-氯-3-吲哚-β-D-半乳糖苷(5-bromo-4-chloro- 3-indolyl-β-D-galactoside ,X-Gal )购于 Sigma 公司;C 18 反相薄层层析板购于 Merck 公司。
高效液质联用仪为 Waters micromass 公司生产,型号为 quattro 。
1.2 方法1.2.1 自体诱导物活性检测(β-半乳糖苷酶法)按参考文献[9-10]的方法进行。
1.2.2 C 18 反相薄层层析(thin-layer chromatography ,TLC ) 按参考文献[11]的方法使用 Whatman 的反向层析进行。
1.2.3 自体诱导物缺失突变株的筛选 按参考文献[11]的方法进行。
1.2.4 AHL 结构的质谱分析 将待测菌株的培养上清液用二氯甲烷萃取,真空旋转蒸发后溶于适量二氯甲烷中,用于电喷雾电离质谱(electrospray ionization mass spectra ,ESI MS/MS )分析。
1.2.5 DNA 凝胶阻滞分析(gel retardation assay ) 通过 PCR 的方法扩增得到需要分析的 DNA 片段(含有或不含有MrtR box ),并在扩增过程中加入带有α-P 32同位素标记的dATP ,可以得到带有同位素标记的 DNA 片段。
把带有标记的 DNA 片段、纯化的MrtR 蛋白以及 AI 分子以适当比例混合后,加入 1/10 体积的结合缓冲液,用超纯水把反应体系补充到适当体积,于 30℃ 作用 30 min 后,反应体系中加入甘油至终浓度为5%,然后用低浓度(6%)聚丙烯酰胺凝胶电泳检测。
2 结果与分析2.1 中慢生天山根瘤菌自体诱导物信号分子结构的测定为进一步揭示不同菌株间信号分子的差异,通过液质联用的方法对天山根瘤菌模式菌株 CCBAU3306 产生的信号分子结构进行了快速测定。
因为 AHLs 类信号分子具有固定的高丝氨酸内酯环,因此在质谱中会形成特征的 m/z102 的碎片峰[12](图 1a ),以此为母离子进行电喷雾电离质谱,能对样品中的 AHLs 分子进行质谱分析。
图 1 AHLs 分子裂解的方式 (a) 和分子结构示意图 (b, c)因为 AHLs 分子的可变端为烃链,而酰基高丝氨酸内酯基团是稳定的,根据这一现象根据分子离子峰的数值可以得出以下公式计算 AHLs 分子的分子式:氢取代类:/117128m z n --=, n 为自然数;羟基取代类:/118728m z n --=,n 为自然数;不饱和羰基类:/118528m z n --=,n 为自然数。
以氢取代为例,m /z -1 为检测的信号分子的分子量,R 为烷基取代,其基本的氨基高丝氨酸内酯基团的分子量是 100(如图1b ),RCO-基团来源于脂肪酸合成途径,该酰基链为偶数个碳,最简单的是C 4 -HSL ,分子量是171,其他的分子与它的差别在于(-CH 2-CH 2- )n 基团的数量,因此用分子量减去C 4 - HSL 的基本分子量 171,氢取代类的差别在于(-CH 2-CH 2-)n 基团的数量,该基团的分子量为 28,按公式得出的 n 值的两倍即为比 C 4-HSL 多的碳原子数,加 4 后即为该分子的碳链长度,也就可知该类信号分子的分子式。
同理在另两类信号分子中(如图1c )用分子量分别减去最基本信号分子的分氢取代类(b)AHLs 分子裂解的方式 (a)不饱和基取代类(c)m/z 102[M+H]*ion[M+H-101]*ion150 土 壤 第42卷子量,再按(-CH 2-CH 2-)n 基团的数量的差别进行计算可以分别得出各自的分子式。
根据高效液质联用的结果(图2)可以得出天山根瘤菌 CCBAU3306所产生自体诱导物分子式为3OH-C 12-HSL 和 3OH-C 14-HSL (图3a )。
21.15102ES+100母离子信号峰丰度(%)14.3735.0010.0015.0020.0025.00时间 (min)图 2 天山根瘤菌CCBAU3306自体诱导物色质联用检测结果Fig.2 Detections of AHLs produced by Mesorhizobium tianshanense CCBAU3306 with HPLC–mass spectrometer图 3 天山根瘤菌CCBAU3306自体诱导物结构及功能验证2.2 中慢生天山根瘤菌自体诱导物信号分子活性的检测为进一步验证测定结果的准确性,采用标样进行生物活性检测。
因为群体感应系统的调控蛋白能与其自体诱导物合成酶合成的信号分子结合,形成二聚体后具有生物活性,并与目的基因片断发生结合,所以用合成的 3OH-C 12-HSL 与该根瘤菌群体感应系统的调控蛋白 MrtR 作用,通过凝胶阻滞分析发现在加入标准品后 MrtR 能与目前片断结合(图3b ),因此证明该信号分子能与调控蛋白作用,该结构是其合成酶合成的一种 AHLs 类信号分子。
3 讨论本研究建立了一种快速的检测方法确定细菌产生AHLs 类信号分子分子式的方法,这种群体感应系统信号分子的结构、合成酶基因和调控基因存在多样性,3OH-C 12-HSL3OH-C 14-HSLBind DNAFree DNA3OH-C 12-HSLR10 HMZ0(a)(b)+-100160180200 220240260280300 320340360380400 420 440 460 480 500140160180200220240260280300320 340 360 380 400 420 440 460 480500520102ES+母离子信号峰326.2102ES+母离子信号峰300.2100 丰度(%)(保留 时间21.340 m i n )丰度(%)(保留时间14.408 m i n )m/zm/z第1期赵家星等:中慢生型天山根瘤菌中自体诱导物分子结构的鉴定 151而且合成酶的专一性不高,往往一种酶能合成几种不同结构的AHL分子[13],在不同的细菌中调控的生理功能各不相同,具有相对的独立性,但在特定的情况下例如当土壤中两种不同细菌产生相同的AHL分子时也能产生信号间分子对话,因此对AI-1型群体感应系统的研究除了对相关基因进行分析,对其产生的自体诱导物分子结构进行分析也十分重要,这对我们了解AI-1型信号途径的多样性具有重要的意义。