基因重组-精品PPT
合集下载
高中基因重组ppt课件

原因 这种重组通常是由于DNA链之间 的错误配对和修复机制引起的。
过程 非同源重组通常发生在两条DNA 链的特定区域,这些区域由不同 的DNA序列定义。在重组过程中 ,这些区域会相互交换,以产生 新的DNA组合。
转座因子引起的重组
定义
转座因子引起的重组是指由于转座因子的存在而 引起的DNA序列之间的交换。
过程
当转座因子在DNA链上移动时,它们可以引起 DNA序列的重新排列和复制,从而产生新的DNA 组合。
原因
转座因子是一种可以在DNA链上移动的基因,它 们可以引起DNA序列的重新排列和复制。
重要性
转座因子引起的重组在生物进化中起重要作用, 因为它们可以产生新的基因组合和功能。同时, 它们也是导致基因组不稳定和疾病发生的重要因 素之一。
Western印记杂交
总结词
一种用于检测特定蛋白质的技术。
详细描述
将蛋白质样本与特定的抗体进行杂交,然后通过曝光底片显示结果,从而判断是 否存在目标蛋白质。
06
基因重组的未来展望
基因重组技术的改进
技术优化
不断优化重组技术,提高重组效率、准确性和稳定性。
新的应用领域
拓展基因重组技术在生物医药、农业、环保等领域的应用。
基因重组技术的伦理和社会问题
安全性问题
重组技术可能产生不可预 测的后果,对人类健康和 生态环境造成潜在风险。
人类尊严问题
基因重组技术可能对人类 生命系统产生深远影响, 需要谨慎考虑其伦理和道 德问题。
社会接受度
公众对基因重组技术的接 受程度和态度需要关注和 引导。
THANKS。
解决伦理和社会问题
积极参与伦理和社会问题的讨论,推动合理、规范地应用基因重 组技术。基因重组技术的商业化前景源自010203
基因重组方法=全PPT课件

.
23
外源DNA被降解,转导失败。
(2)局限性转导(specialized transduction)
温和噬菌体感染
整合到细菌染色体的特定位点上
宿主细胞发生溶源化
溶源菌因诱导而发生裂解时, 在前噬菌体二侧的少数宿主 基因因偶尔发生的不正常切 割而连在噬菌体DNA上
部分缺陷的温和噬菌体
把供体菌的少数特定基因转移到受. 体菌中
的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
进行自然转化,需要二方面必要的条件:
建立了感受态的受体细胞
外源. 游离DNA分子
30
枯草芽孢杆菌的自然转化过程(革兰氏阳性菌的转化模型)
分泌感受态因子
与细胞表面受 体M相互作用
使细胞表面的 DNA结合蛋白 及核酸酶裸露出 来,使其具有与 DNA结合的活 性
b)决定因素也各有不同;
.
34
(2)人工转化
在自然转化的基础上发展和建立的一项细菌基因重组手段, 是基因工程的奠基石和基础技术。
不是由细菌自身的基因所控制;
用多种不同的技术处理受体细胞,使其人为地处于一 种可以摄取外源DNA的“人工感受态”。
用CaCl2处理细胞,电穿孔等是常用的人工转化手段。
质粒的转化效率高;
含有F因子的细胞:“雄性”菌株(F+),其细胞表面有性菌毛 不含F因子的细胞:“雌性”菌株. (F-),细胞表面没有性菌毛7
F因子为附加体质粒 既可以脱离染色体在细胞内独立存. 在,也可插入(整合)到染色8 体上
F因子的四种细胞形式
a)F-菌株, 不含F因子,没有性菌毛,但可以通过 接合作用接收 F因子而变成雄性菌株(F+);
.
基因突变和基因重组-ppt课件

探究一:基因突变
思考.讨论 3.癌细胞与正常细胞相比,具有哪些明显的特点?
正常的成纤维细胞
癌变的成纤维细胞
①能够无限增殖; ②形态结构发生显著变化; ③细胞膜上糖蛋白等物质减少,细胞间黏着性降低,易在体内分散和转移。
4.如何避免癌症的发生? 远离致癌因子,选择健康的生活方式
探究一:基因突变
5 基因突变的原因
➢ 遗传特性
基
发生在配子中
将遵循遗传规律传递给后代
因
突
人类体细胞中某些基因的突
变
发生在体细胞中 变可一能般发不展能遗为传癌细胞!!!
有些植物(无性繁殖的生物)的体细胞发生了
基因突变,可以通过无性生殖遗传。
探究一.基因突变
思考.讨论
2.健康人的细胞中存在原癌基因和抑癌基因吗?作用分别是什么呢?
原癌基因
替 换AT GC
ACG
增 添 AT GC
ACG
缺 失 AT GC
ACG
A CGC GCG
A T AGC A T CG
A GC CG
只有使基因结 构发生改变才 是基因突变, 非基因区段的 碱基改变不是
基因突变。
2 基因突变的时期
通常发生在DNA 复制即分裂前的间期。
(DNA复制时要解旋为单链,单链DNA的稳定性会大大降低,极易受到影响而 发生碱基的改变。)
情境导入
【资料1】抗倒伏、抗条锈病水稻品种是利用抗倒伏、易感条锈病水稻品种 与易倒伏、抗条锈病水稻品种作为亲本,进行杂交和多年选育获得的。P13 【资料2】早在1987年,我国就将作物种子带入太空,利用太空中的特殊 环境诱导基因发生突变,然后在地面选择优良的品种进行培育。P80 【资料3】残翅果蝇幼虫在31℃环境中培养,将得到一些翅长接近正常的果 蝇成虫,但其再正常温度25℃下产生的后代仍然后残翅果蝇。P75
基因突变和基因重组ppt课件

思 考 基因重组能否产生新的基因?
基因重组是原有基因的重新组合,只产生新的基因型 和重组性状,不能产生新基因与新性状。
总结基因突变与基因重组
结果 类型 时间
基因突变
产生新基因 碱基的
替换、增添、缺失
细胞分裂间期(主要)
基因重组
产生新基因型 基因的交叉互换 基因的自由组合
MⅠ前期、MⅠ后期
意义
变异的根本来源 进化的原始材料
杂交水稻之父 ·袁隆平
从1964年起,袁隆平就开始研究杂交水稻, 到1975年,他研究出来的新品种就已经在全 国推广,并取得了非同凡响的成果。此后十年 内中国杂交水稻累计增产超亿吨,每年增产的 大米可以多养活6000万人。
概 念 在生物进行 有性生殖 的过程中,控制不同性
状的基因的 重新组合 。
思考
为什么这种变异性状不能遗传给子代?
分析:是环境因素引起的 ,自身的遗传物质没有 改变。
什么是生物变异? 亲代与子代、子代与子代个体之间的性状的差异性
表现型 = 基因型 + 环境
生物变异
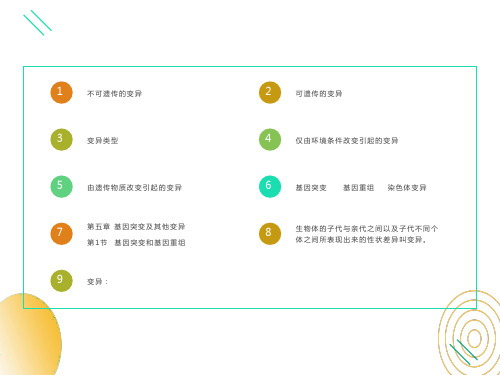
不可遗传的变异
(环境引起,不改变遗传物质)
基因突变 可遗传的变异 基因重组 (改变遗传物质) 染色体变异
▲注意:基因中碱基序列不发生改变,有时候也可通过表观遗传影响下一代。
① 在适宜条件下,能够无限增殖。 ② 形态结构发生显著变化。
癌细胞的扫描电镜照片
③ 细胞膜上的糖蛋白等物质减少,细 胞之间的黏着性显著降低,容易在机 体内分散和转移。
人和动物细胞中的 DNA 上本来就存在与癌变相关的基因:
原癌基因
抑癌基因
表达的蛋白质是细胞正常 的生长和增殖所必需的。
表达的蛋白质能抑制细胞的生长 和增殖,或者促进细胞凋亡。
第四节基因突变和基因重组ppt课件

开篇语
“一母生九子,连母十个样” 生物的变异是普遍存在的。有 的仅仅是由于环境因素影响造 成的;有的则是由于生殖细胞 内的遗传物质发生改变而引起 的。
圆饼状的红细胞
镰刀状的红细胞
事实
2、科学家进一步研究发现,镰 刀型细胞贫血症患者的血红蛋白的一 条多肽链上的氨基酸组成发生了变化。 DNA测序发现,决定血红蛋白的相关 基因也发生了变化。
资料一: 在北京培育的优质甘蓝品种,叶球最大
的有3.5KG,当引种到拉萨后,由于昼夜 温差大、日照时间长、光照强,叶球可重 达7KG左右。但再引回北京后,叶球又只 有3.5KG。
资料二: 太空椒(普通青椒种子遨游过太空后培
育而成)与普通青椒对比,果实明显增大, 将太空椒的种子种植下去,仍然是肥大果 实。
生物变异的类型
生物的变异
不可遗传的变异 基因突变
可遗传的变异 基因重组 (来源)
染色体变异
一、基因突变
➢基因突变的概念 ➢镰刀型细胞贫血症的分析 ➢基因突变的原因 ➢基因突变的特征 ➢基因突变的意义
基因突变:
是指DNA分子中碱基对的 增添、缺失或改变等变化。基 因突变可以产生新基因,是生 物变异的根本来源。
• 2、交换重组:同源染色体之间遗传物质的 交换。结果是导致染色体上基因的组成和 排列次序的改变,从而可能出现新的性状。
同源染色体之间交换示意图
基因重组的意义
✓是生物进化的源泉。 ✓是生物体多样性的重要 原因之一。Fra bibliotek积极思维
玫瑰鸡冠和豆鸡冠从何而来?
事实:研究发现,鸡冠的形态是 由两对基因决定的。若玫瑰鸡冠和豆 鸡冠交配,则子一代全部都是胡桃鸡 冠。如果子一代胡桃鸡冠相互交配, 子二代出现4种不同的性状。
4.2 基因重组使子代出现变异 课件(共18张PPT)浙科版必修二

优点:实现种间遗传物质的交换,克服远 缘杂交不亲和的障碍
转基因生物:由于外源基因的导入而引起 原有遗传物质组成发生改变的生物。
植物转基因技术过程:用人工方法将人类 所需要的目的基因导入受体细胞内,使其 整合到受体的染色体上。
植物转基因常用方法: 农杆菌转化法
基础
训练
P57
B
A
基因重组
概念:具有不同遗传性状的雌、雄个体进行有性生殖时,控制不同 性状的基因重新组合,导致后代出现不同于亲本类型的现象或过程。
F6抗病、红果肉个体中纯合子占多少?31/33
抗病、红果肉 aaRR
基础 训练 P57
D
基础 训练 P57
选择 纯合化
②
3/16
1/3
连续自交
性状分离
基因重组可应用于杂交育种
杂交育种:利用基因重组原理,可以有目的地将两个或多个品种的 优良性状组合在一起,培有出更优良的新品种。
过程:一般可以通过杂交、选择、纯合化等手段培育出新品种
F1
易感病、红果肉 SsRr
×
稳定遗传的纯合子占 1/3 不稳定遗传的杂合子占 2/3
F2 易感病、红果肉 易感病、黄果肉 抗病、红果肉
9/16 S_R_
3/16 S_rr
3/16 ssR_
能获得抗病、红果肉番茄的关键是什么?
……×
F1产生了基因型为sR的配子→自由组合(基因重组)
×
抗病、黄果肉 1/16 ssrr
条件:非等位基因!!! 方式
非同源染色体间的自由组合导致基因重组 同源染色体的非姐妹染色单体之间交叉互换导致基因重组 物种间:转基因技术(基因工程)
应用:杂交育种、基因工程育种
抗病、黄果肉 1/16 ssrr
转基因生物:由于外源基因的导入而引起 原有遗传物质组成发生改变的生物。
植物转基因技术过程:用人工方法将人类 所需要的目的基因导入受体细胞内,使其 整合到受体的染色体上。
植物转基因常用方法: 农杆菌转化法
基础
训练
P57
B
A
基因重组
概念:具有不同遗传性状的雌、雄个体进行有性生殖时,控制不同 性状的基因重新组合,导致后代出现不同于亲本类型的现象或过程。
F6抗病、红果肉个体中纯合子占多少?31/33
抗病、红果肉 aaRR
基础 训练 P57
D
基础 训练 P57
选择 纯合化
②
3/16
1/3
连续自交
性状分离
基因重组可应用于杂交育种
杂交育种:利用基因重组原理,可以有目的地将两个或多个品种的 优良性状组合在一起,培有出更优良的新品种。
过程:一般可以通过杂交、选择、纯合化等手段培育出新品种
F1
易感病、红果肉 SsRr
×
稳定遗传的纯合子占 1/3 不稳定遗传的杂合子占 2/3
F2 易感病、红果肉 易感病、黄果肉 抗病、红果肉
9/16 S_R_
3/16 S_rr
3/16 ssR_
能获得抗病、红果肉番茄的关键是什么?
……×
F1产生了基因型为sR的配子→自由组合(基因重组)
×
抗病、黄果肉 1/16 ssrr
条件:非等位基因!!! 方式
非同源染色体间的自由组合导致基因重组 同源染色体的非姐妹染色单体之间交叉互换导致基因重组 物种间:转基因技术(基因工程)
应用:杂交育种、基因工程育种
抗病、黄果肉 1/16 ssrr
基因突变和基因重组ppt课件

一、基因突变的实例
思考·讨论
正常人
镰状细胞贫血患者
碱基对 T
根本原因:
DNA
替换
A
基因层面
转录
mRNA
A
U
翻译
氨基酸 蛋白质
谷氨酸 正常
缬氨酸 异常
直接原因: 蛋白质层面
一、基因突变的实例
思考·讨论
3.如果这个基因发生碱基的增添或缺失,氨基酸序列是否也会改变?所 对应的性状呢?
【答案】如果这个基因发生碱基的增添或缺失,氨基酸序列也会发 生改变,所对应的性状肯定会改变。
本质
发生 时间 原因
基因突变
基因结构改变,产生 新基因、可能会产生新性状
分裂间期
自然突变、诱发突变
基因重组
不同基因的重新组合,产生新 的基因型,使不同性状重新组合
减I(前期、后期)
自由组合;互换
特点 意义
普遍性、随机性、低频性、 不定向性。
变异的根本来源,为进化 提供原材料
有性生殖生物中普遍存在
变异的来源之一,对进化有 重要意义
取得了极大的经济效益。
讨论
1.航天育种的生物学原理是什么? 基因突变
2.如何看待基因突变造成的结果?
基因突变的本质是基因的碱基序列发生改变,这种改变可以直 接表现在性状上,改变的性状对生物的生存可能有害,可能有利, 也可能既无害也无益。
一、基因突变的实例
镰状细胞贫血
镰状细胞贫血(也叫镰刀型细胞贫血症)是一种遗传病。正常 人的红细胞是中央微凹的圆饼状,而镰状细胞贫血患者的红细胞 却是弯曲的镰刀状这样的红细胞容易破裂,使人患溶血性贫血, 严重时会导致死亡。
一、基因突变的实例
镰状细胞贫血
基因突变和基因重组课件
叁
结 果:
基因工程又叫做基因拼接技术或DNA重组技术。通俗的说,就是按 照人们的意愿,把一种生物的某种基因提取出来,加以修饰改造,然 后放到另一种生物的细胞里,定向地改造生物的性状。
1. 提取目的基因 2. 目的基因与运载体结合 3. 将目的基因导入受体细胞 4. 目的基因的检测和表达
基因工程的基本操作步骤:
三.基因重组发生的时期
○ 在生物体进行有性生殖的过程中,控制不同性状的基因的重新 组合。 一.在减数第一次分裂前期:同源染色体中非姐妹染色单体 上的等位基因间的交叉互换 二.在减数第一次分裂后期:非同源染色体上的非等位基因 的自由组合
C
基因重组能够发生在下列哪种生物中() 原核生物 病毒 熊猫
单击此处添加正文, 文字是您思想的提炼, 为了演示发布的良好 效果,请言简意赅地 阐述您的观点。
1. 基因突变的原因
2. 基因突变的特点
3. 基因突变的意义
○ 外界因素: ○ 内在因素:
● 普遍性、随机性、不定向性、低频性和多害少利性。 1. 是新基因产生的途径; 2. 是生物变异的根本来源; 3. 是生物进化的原材料。 • 物理因素、化学因素和生物因素 • 偶尔会出现复制出错
三.下列哪种情况能产生新的基
C
因( )
A
B
C
D
01 B
02 果蝇约有104对基因,假定 每个基因的突变率都是105,一个大约有109个果蝇 的果蝇群,每一代出现的 基因突变数是( )
03 A 2×109
B
04 C 2×107
D
2×108
108
2×104×10-5×109=
2×108个。
8. 诱变育种的应用 原理: 方法: 实例: 利 用 优物 点理 :因 素 或 化 学 因 素 处 理 生 物 缺点:
基因重组___上课(好)ppt课件

14
6、基因重组的应用
在生产上的应用
1.杂交育种
简述用纯种黄色圆粒豌豆和绿色皱粒豌豆培育纯种绿色圆粒豌豆的过程。 要求写出遗传图解并加以适当的文字解释。
2.基因工程
;.
16
杂交育种:
原理:
基因重组
方法: 优点:
杂交——自交——筛选 操作简单、能根据人的预见把位于两个生物体上的优良性状集于一身。
缺点:
“一母生九仔,连母十个样”,这种个体的差异,主要是什么原因产生的? 基因重组
1
基因重组: 是指生物体在进行有性生殖的过程中,控制不同性状的基因重新组合
的过程。
2
第2节 基因重组 一、细菌的基因重组 二、高等生物的基因重组 三、基因的人工重组—基因工程
4
二、高等生物的基因重组 1.类型:
①基因的自由组合: (减I后期)
减数第一次分裂前期的四分体时期的交叉互 换; 减数第一次分裂后期,非同源染色体上的非 等位基因自由组合。
条件 意义
外界环境条件的变化和内部 因素的相互作用。
新基因产生的途径,是生物变异的 根本来源,是生物进化的原始材料。
有性生殖过程中进行减数分裂形成生殖细 胞。
是生物变异的重要来源,是形成生物多样性 的重要原因,对生物的进化也具有重要的意 义
22
1.基因突变:
基发因生时___期__:内___部____结_改_构_变__,__它__________(能__或__否__)_产_ 生新的能基因
特点:①普遍性、 ②随细机胞性分、裂③间_期__(__D_N__A__复_、制时) ④少利多害性、⑤不定向性。
2.基因重组:
低频性
控制不同性状的_____________,_______新基因,可形成新的________。
6、基因重组的应用
在生产上的应用
1.杂交育种
简述用纯种黄色圆粒豌豆和绿色皱粒豌豆培育纯种绿色圆粒豌豆的过程。 要求写出遗传图解并加以适当的文字解释。
2.基因工程
;.
16
杂交育种:
原理:
基因重组
方法: 优点:
杂交——自交——筛选 操作简单、能根据人的预见把位于两个生物体上的优良性状集于一身。
缺点:
“一母生九仔,连母十个样”,这种个体的差异,主要是什么原因产生的? 基因重组
1
基因重组: 是指生物体在进行有性生殖的过程中,控制不同性状的基因重新组合
的过程。
2
第2节 基因重组 一、细菌的基因重组 二、高等生物的基因重组 三、基因的人工重组—基因工程
4
二、高等生物的基因重组 1.类型:
①基因的自由组合: (减I后期)
减数第一次分裂前期的四分体时期的交叉互 换; 减数第一次分裂后期,非同源染色体上的非 等位基因自由组合。
条件 意义
外界环境条件的变化和内部 因素的相互作用。
新基因产生的途径,是生物变异的 根本来源,是生物进化的原始材料。
有性生殖过程中进行减数分裂形成生殖细 胞。
是生物变异的重要来源,是形成生物多样性 的重要原因,对生物的进化也具有重要的意 义
22
1.基因突变:
基发因生时___期__:内___部____结_改_构_变__,__它__________(能__或__否__)_产_ 生新的能基因
特点:①普遍性、 ②随细机胞性分、裂③间_期__(__D_N__A__复_、制时) ④少利多害性、⑤不定向性。
2.基因重组:
低频性
控制不同性状的_____________,_______新基因,可形成新的________。
基因突变与基因重组(原创优质课)精品PPT课件

原句: The cat sat on the mat.
①The hat sat on the mat. ②The cat sat on then mat.
问题3③:The cat on the mat. DNA在复制的时候会不会出 现同样的错误呢?
正常红细胞
镰刀型红细胞
镰刀型红细胞与正常细胞相比:容易破裂 使人患溶血性贫血,严重时会导致死亡
蓝 紫 色 花 后 代 仍 是 蓝 紫
色
生物变异的类型
不可遗传的变异:仅仅由环境不同引起,遗
变
传物质没有改变,不能进
异
一步遗传给后代。
的
类
基因突变
型 可遗传的变异: 基因重组
(第一节)
(由于遗传物质
发生了改变而引 染色体变异(第二节内容)
起的变异)
第五章 第一节 基因突变与基因重组
抄写英语句子:
看我们哥仨长得一样不?
同种生物个体之间,在性状上表现出来的差异现象, 就叫做:
问题1:通过美容手术,纹成弯弯的柳叶眉,这 种柳叶眉能遗传吗?为什么?
不可遗传
问题2:在种植花卉时,发现红花的后代出现了蓝
紫花,蓝紫色花的后代仍是蓝紫色。蓝紫色花可
以遗传吗?为什么?
红 花 的 后
可遗传 代 变 了 蓝 紫 色
能力提升
原核生物某一基因起始端插入了 1 个碱基对。在 插入位点的附近,再发生下列哪种情况有可能对 其编码的蛋白质结构影响最小( )
A.置换单个碱基对 B.增加 4 个碱基对
C.缺失 3 个碱基对 D.缺失 4 个碱基对
课件下载后可自由编辑,如有不理解
之处可根据本节内容进行提问
Thank you for coming and listening,you can ask questions according to this section and this courseware can be downloaded and edited freely
①The hat sat on the mat. ②The cat sat on then mat.
问题3③:The cat on the mat. DNA在复制的时候会不会出 现同样的错误呢?
正常红细胞
镰刀型红细胞
镰刀型红细胞与正常细胞相比:容易破裂 使人患溶血性贫血,严重时会导致死亡
蓝 紫 色 花 后 代 仍 是 蓝 紫
色
生物变异的类型
不可遗传的变异:仅仅由环境不同引起,遗
变
传物质没有改变,不能进
异
一步遗传给后代。
的
类
基因突变
型 可遗传的变异: 基因重组
(第一节)
(由于遗传物质
发生了改变而引 染色体变异(第二节内容)
起的变异)
第五章 第一节 基因突变与基因重组
抄写英语句子:
看我们哥仨长得一样不?
同种生物个体之间,在性状上表现出来的差异现象, 就叫做:
问题1:通过美容手术,纹成弯弯的柳叶眉,这 种柳叶眉能遗传吗?为什么?
不可遗传
问题2:在种植花卉时,发现红花的后代出现了蓝
紫花,蓝紫色花的后代仍是蓝紫色。蓝紫色花可
以遗传吗?为什么?
红 花 的 后
可遗传 代 变 了 蓝 紫 色
能力提升
原核生物某一基因起始端插入了 1 个碱基对。在 插入位点的附近,再发生下列哪种情况有可能对 其编码的蛋白质结构影响最小( )
A.置换单个碱基对 B.增加 4 个碱基对
C.缺失 3 个碱基对 D.缺失 4 个碱基对
课件下载后可自由编辑,如有不理解
之处可根据本节内容进行提问
Thank you for coming and listening,you can ask questions according to this section and this courseware can be downloaded and edited freely
基因突变和基因重组ppt课件-(2)

细胞结构 细胞代谢
调节基因
间接控制作用
性状
正常型红细胞(左)与镰刀型红细胞(右 )
正常人的红细 胞是圆饼状的,镰刀型细胞贫血症患者的 红细胞却是弯曲的镰刀状的。这样的红细胞容易破裂, 使人患溶血性贫血,严重时会导致死亡。
血红蛋白是由四条多肽链组成.研究证明,在整个 多肽链的574个氨基酸中,病者与正常人的573个氨基 酸是相同的。所不同的是一个谷氨酸被缬氨酸替代, 因而引起血红蛋白结构异常。
但一般不能传给后代(产生性原细胞时可以)
B.减数第一次分裂间期 产生生殖细胞可以发生基因突变 可以通过受精作用直接传给后代
基因突变的结果:
基因突变是染色上某一个位点上基因的改变,使
一个基因变成它的等位基因。
基因A
例如 1
(DNA分子复制时)
外界因 素诱导
碱基序列改变 2
遗传信息改变
变突 基因a1或a2… 3 小麦从高秆变成矮秆
由遗传物质发生了 可遗传的变异
诱
改变,其后代将继
基因重组 因
承这种改变。
三种来源: 基因突变
染色体变异
讨论:是不是环境因素引起的变异就一定不能遗传呢?
基因与性状之间的关系
DNA
DNA复制
基因
转录 逆转录
翻译
RNA
RNA复制
基因型
决定 表现型
直接控制作用
蛋白质 性状
结构基因 选择信息
表达 结构蛋白 信息 酶或激素
基因重组
1、概念: 是指在生物体进行有性生殖的过程中, 控制不同性状来自基因的自由组合.2、类型:
①基因的自由组合:
非同源染色体上的非 等位基因的自由组合
②基因的互换: 同源染色体上的非 姐妹染色体之间发 生局部互换.
《基因重组》课件

生物学意义
转座重组有助于基因组的 多样性和进化,但也可能 导致基因表达的异常和疾 病的发生。
03 基因重组的机制
重组酶的作用
识别DNA序列
重组酶能够识别DNA上的 特定位点,为重组过程提 供起始点。
催化DNA链断裂
重组酶能够催化DNA链断 裂,形成单链缺口,为重 组提供必要的结构基础。
催化碱基配对
同源重组是生物进化的重要机制之一 ,有助于维持基因组的稳定性,同时 也可以修复DNA损伤。
机制
同源重组依赖于RecA蛋白等分子伴侣 的参与,通过寻找同源序列,形成异 源二聚体,再通过DNA链的断裂、交 换和重连完成重组。
非同源重组
定义
非同源重组是指两条没有同源 序列的DNA分子之间发生的
重组过程。
些罕见病。
基因克隆
03
通过基因重组技术将目的基因克隆到载体中,实现基因的高效
表达和纯化。
克隆技术
动物克隆
利用基因重组技术复制动物个体,实现快速繁殖和拯救濒危物种 。
植物克隆
通过基因重组技术繁殖植物,实现植物的快速繁殖和品种改良。
细胞克隆
通过基因重组技术培养特定细胞系,用于药物筛选、疾病研究等 。
疾病治疗与预防
伦理考量
在编辑人类基因时,需要权衡潜在的 健康益处与潜在的风险和副作用,以 及涉及的伦理问题,如潜在的基因歧 视和人类进化干预。
基因重组与生物多样性
生物多样性影响
基因重组可能导致生物多样性减少,因为重组可能导致物种的基因库减少,降低 物种适应环境变化的能力。
伦理考量
需要关注基因重组对生物多样性的影响,并采取措施确保生物多样性的保护和维 持。
疫苗研发
利用基因重组技术生产疫苗,预防和控制传染病。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
轻链的基因片段:
L
V
J
C
重链的基因片段:
L
V
D
J
C
CACAGTG(12/23)ACAAAAACC
GTGTCCAC
TGTTTTTGG
基因片段
重组信号序列
重链(IgH)基因的V-D-J重排和轻链(IgL)基 因的V-J重排均发生在特异位点上。在V片段的 下游,J片段的上游以及D片段的两侧均存在保 守 的 重 组 信 号 序 列 (recombination signal sequence, RSS) 。 此 重 排 的 重 组 酶 基 因 rag (recombination activating gene)共有两个, 分别产生蛋白质RAG1和RAG2。
免
间插DNA
疫 球
V片段
RSS
RSS J片段
蛋
单链切开 RAG1 RAG2
白
OH
基
OH
因
分子内转酯反应
重
排
过
单链切开
程
转移核苷酸 修复、连接
V
J
目录
四、转座重组
由插入序列和转座子介导的基因移位或重 排称为转座(transposition)。
由 McClintock ( 1956 ) 在 玉 米 上 首 先 发 现 遗传学发展史上的重要里程碑之一。
IR Transposase Gene IR
发生形式: 保守性转座(conservative transposition) 复制性转座(duplicative transposition)
插 入 序 列 的 复 制 性 转 座
目录
(二)转座子转座
转座子(transposons) ——可从一个染色体位 点转移到另一位点的分散重复序列。
转座子可在染色体组内移动,从一个位点 切除,插入到一个新的位点,引起基因的突变或 染色体重组。
(一)插入序列转座
插入序列(insertion sequences, IS)组成: 二个分离的反向重复(inverted repeats, IR)序列 特有的正向重复序列 一个转座酶(transposase)编码基因
ag A g+
ag 5 :3
异常 a g+ 分离 A g+
Ag
ag
图 23-7 基 因 转 变 和 减 数 后 分 离
6 :2
正常分离
4:4 异常分离
Holliday 模型
以E.coli的同源重组为例,了解同源重组 机制的Holliday模型。
Holliday模型中,同源重组主要4个关键步骤
①两个同源染色体DNA排列整齐 ②一个DNA的一条链断裂、并与另一个DNA对应的
IS-两端有IR,只编码转座酶
原 类转座因子-结构同IS,但不能独立存在,仅作为复合转座子的两端组件
核 复合转座子-两端由IS或类IS构成,可编码抗抗菌素物质
TnA转座子家族-两端为IR,可编码转座酶、解离酶和抗性物质
转 AC-Ds-植物(玉米)中的激活-解离因子
转 座 因 子
座 子
P 因子-果蝇中父本因子,在M♀×P♂中导致杂种不育
断裂重接模型和模板选择复制模型的否定
断裂重接模型不能解释基因转变和极化子现象 模板选择复制模型存在的问题:
(1) 违背了半保留复制; (2) DNA复制应在S期,重组应在偶线期,
不应同时发生。 (3)不能解释3线和4线交换。
基因转变----与遗传重组有关
1930年Winkler把真菌中不规则分离现象解释为基因转变 (conversion)
复
连接
制
IR (34bp) 和 Tn3 解离 倒位
无
相似
复
制
倒位
无
复
制
倒位
无
复
制
DR, IR
DR → 切 无
离和缺 复
失
制
IR → 倒
位
靶
整 合 酶 在 整合
修
LTR 的 3′
复
端 产 生 2b
性
缺口在靶上
复
交错切割连
制
接填补缺口
同源特 异重组
转座 重组
模板 转换
特殊 重组
酵母交配 型转变
复制型
非复制型
5´
3´
5´ 3´
5´3´
5´ 片 段
重
组
体
3´ 5´
3´ 内切酶
5´
(ruvC)
3´ 5´
5´ 3´
5´ 3´
拼
5´
接 3´
重
组 体 53´´
3´ 5´
DNA
连接酶
3´ 5´
3´ 5´
目录
(1) 5’ A 3’ 3’ 5’ a
(2) 5’ A 3’ 3’ 5’ a
B 3’
5’
5’
b 3’ 联会
B 3’
λ噬菌体的整合酶识别噬菌体和宿主染色体的 特异靶位点发生选择性整合;
反转录病毒整合酶可特异地识别、整合反转录 病毒cDNA的长末端重复序列(long terminal repeat, LTR)。
λ噬菌体 P O P’
包装
感染宿主
B O P’
P O P’ ×
B O B’
Int
Int
IHF
IHF
Xis P O B’
链连接,形成Holliday中间体 ③通过分支移动产生异源双链DNA ④Holliday中间体切开并修复,形成两个双链重
组体DNA,分别为:
片段重组体(patch recombinant)
拼接重组体(splice recombinant)
5´
3´
5´
3´
5´ 内切酶 3´
3´ 5´
5´
3´
(recBCD)
现了异常分离。认为这种异常分离是有丝分裂的产物, 称为减数后分离(postmeiotic segreation)
A g+
A g+ ag
全部校正
4:4
正常分离
A g+ a g+ a g+
ag
A g+
A g+
a g 部分
校正
a g+
A g+
A
g
a
g
ag
A g+
a g+
a g+
A g+
Ag ag
ag
未校正
保守型
I 类内含子 归巢
反转录转 座子 Ⅱ类内含 子归巢 免疫球蛋 白 的 VJC 连接
完全 同源 无 无 无 无
区域 同源
Rid 52, HO
文档,文
基因重组
三级安全培训资料
2006年7月
1
本章要求 第一节 DNA的重组
DNA重组
同源重组 (homologous recombination) 接合作用 (conjugation) 转化作用 (transformation) 转导作用 (transduction) 位点特异的重组(site-specific recombination) 转 座 (transposition)
Lindegren,C.C(1949)报道了酵母有规律的异离常分离 现象:交配型A×a的杂交中,有些子囊所含的孢子为 (3A+1a),也称为基因转变。
50年代中期Mitchell,M. B. 异常分离不是由于整个染色体 的异常行为,而是由于特定位点的“基因转变”所致, 即属于基因内重组。
Olive, E1-Ani和Kitani等在粪壳菌(Sodaria fimicola)中也发
反转录病毒、RNA →DNA→整合宿主靶DNA
真 核
反 转 录
病毒超家族
Ty Copia LINSL1
(1)有长末端重复序列 (2)编码反转录酶或整合酶 (3)可含内含子
转
(1)无重复序列
座 子
非病毒超家族
SINSB1/ Alu
(2)不编码转座子产物 (3)无内含子
假基因
λ整合
区域同源 Int, IHF
3´ 5´
5´
3´
5´
3´
5´ 分支迁移 3´
3´
5´ (recA) 3´
5´
3´
5´
内切酶
(recBCD)
3´ 5´ 5´
3´ 5´ 3´
DNA侵扰 (recA)
3´
5´
5´ 3´
5´
3´
DNA 5´
3´
3´
5´ 连接酶 3´
5´
3´
5´
3´
5´
3´
3´
5´
3´
Holiday中间体
目录
片段重组体 (见模型图左边产物): 切开的链与原来断裂的是同一条链,重组
(7)
A
B 3’
5’
5
b
3’
分支点移动
B
(11) A a
A a
b
B
b a
B b
B b
a
A B
b a
A
b
a
B
A
b
a
B
a
b
图 23-8 Holiday 重组模型。
Holiday模型说明了遗传重组过程,也说 明了基因转变的现象
二、细菌的基因转移与重组
(一)接合作用
当细胞与细胞、或细菌通过菌毛相互接 触时,质粒DNA从一个细胞(细菌)转移至 另一细胞(细菌)的DNA转移称为接合作用 (conjugation)。
质粒 —— 细菌染色体外的小型环状双链DNA分子 可接合质粒如 F 因子(F factor)