模拟增温对小麦田土壤碳氮循环过程的影响
青海高寒草甸马勃蘑菇圈增温作用的影响研究

第32卷 第3期V o l .32 No .3草 地 学 报A C T A A G R E S T I A S I N I C A2024年 3月M a r . 2024d o i :10.11733/j.i s s n .1007-0435.2024.03.017引用格式:曹 铭,王文颖,徐 进,等.青海高寒草甸马勃蘑菇圈增温作用的影响研究[J ].草地学报,2024,32(3):812-817C A O M i n g ,WA N G W e n -y i n g ,X UJ i n ,e t a l .T h e S t u d y a b o u t T h e r m a l e f f e c t o fQ i n g h a i L y c o p e r d a l e s F u n gu s F a i r -y R i n g [J ].A c t aA gr e s t i aS i n i c a ,2024,32(3):812-817青海高寒草甸马勃蘑菇圈增温作用的影响研究曹 铭1,王文颖2,3*,徐 进4,周华坤5,刘艳方1,德却拉姆2,杨玉青2(1.青海师范大学地理科学学院,青海西宁810008;2.青海师范大学生命科学学院,青海西宁810008;3.青海省青藏高原药用动植物资源重点实验室,青海西宁810016;4.山西农业大学园艺学院,山西太古030801;5.中国科学院西北高原生物研究所,青海西宁810008)收稿日期:2023-09-14;修回日期:2023-09-20基金项目:国家重点研发课题(2023Y F F 1304305)和青海省2021年度第一批中央引导地方科技发展专项资金(2021Z Y 002)资助作者简介:曹铭(1989-),男,回族,甘肃兰州人,博士研究生,主要从事蘑菇圈真菌相关研究,E -m a i l :791957817@q q.c o m ;*通信作者A u -t h o r f o r c o r r e s p o n d e n c e ,E -m a i l :w a n g w y0106@163.c o m 摘要:蘑菇圈是广泛存在于草地和森林的大型真菌子实体环状群落㊂有些蘑菇圈如马勃蘑菇圈会显著促进周围植物生长,其蘑菇圈真菌促进植物生长的机理一直是一个热点科学问题㊂本研究选取青海海北高寒草甸马勃蘑菇圈,从2023年6月15日至7月15日,对蘑菇圈土壤温度进行连续监测,使用红外热成像仪对蘑菇圈子实体进行拍照,同时测定了土壤理化性质与酶活性,检验蘑菇圈真菌生长对土壤温度和土壤酶活的影响及其相互关系㊂结果表明:蘑菇圈圈上土壤温度变化与圈内㊁圈外有明显差异,在当日23时后至次日凌晨5时,蘑菇圈上土壤温度高于圈外与圈内0.3ħ~1.6ħ,红外影像显示蘑菇圈真菌子实体温度显著高于环境温度4ħ㊂蘑菇圈圈上速效养分含量显著高于圈内圈外(P <0.05)㊂蘑菇圈土壤脲酶㊁酸性磷酸酶㊁蔗糖酶㊁过氧化氢酶活性显著高于圈内和圈外㊂因此马勃蘑菇圈真菌的生长增加了土壤环境温度㊁提高土壤酶活,通过增加土壤速效养分促进了蘑菇圈上植物的生长㊂关键词:蘑菇圈;高寒草甸;土壤增温;土壤酶活中图分类号:S 718.81 文献标识码:A 文章编号:1007-0435(2024)03-0812-06T h e S t u d y a b o u t T h e r m a l E f f e c t o f Q i n g h a i L y c o p e r d a l e s F u n g u s F a i r y R i n gC A O M i n g 1,WA N G W e n -y i n g 2,3*,X UJ i n 4,Z H O U H u a -k u n 5,L I U Y a n -f a n g 1,D E Q U EL a -m u 2,Y A N G Y u -q i n g2(1.C o l l e g e o fG e o g r a p h i c a l S c i e n c e ,Q i n g h a iN o r m a lU n i v e r s i t y ,X i n i n g ,Q i n g h a i P r o v i n c e ,810008,C h i n a ;2.C o l l e g e o fL i f eS c i e n c e ,Q i n g h a iN o r m a lU n i v e r s i t y ,X i n i n g ,Q i n g h a i P r o v i n c e 810008;3.Q i n g h a iK e y L a b o r a t o r y o fM e d i c i n a lA n i m a l a n dP l a n tR e s o u r c e s o f T i b e t a nP l a t e a u ,X i n i n g ,Q i n g h a i P r o v i n c e 810016,C h i n a ;4.C o l l e g e o fH o r t i c u l t u r e ,S h a n x iA g r i c u l t u r a lU n i v e r s i t y ,T a i g u ,Q i n gh a i P r o v i n c e 030801,C h i n a ;5.N o r t h w e s t I n s t i t u t e o f P l a t e a uB i o l o g y ,C h i n e s eA c a d e m y o f S c i e n c e s ,X i n i n g ,Q i n gh a i P r o v i n c e 810008,C h i n a )A b s t r a c t :F a i r y r i n g ,n a t u r a l l y o c c u r r i n g c i r c l e sm a d e u p o fm u s h r o o m s ,h a v e b e e n o b s e r v e d t o i n f l u e n c e t h e gr o w t h o f s u r r o u n d i n g p l a n t s ,p a r t i c u l a r l y p r o m o t i n g t h e g r o w t ho f p l a n t s i n t h eO Nz o n e .T h em e c h a n i s mo fm u s h r o o mr i n gf u ng i p r o m o t i n g p l a n t sh a s a l w a y s b e e n a c o n c e r n o f r e s e a r c h e r s .T hi s s t u d y f o c u s e s o n t h em e c h a n i s m s b y w h i c h f a i r yr i n g f u n g i ,s p e c i f i c a l l y H a i b e i L y c o p e r d a l e s ,e n h a n c e p l a n t g r o w t h .W em o n i t o r e d t h e s o i l t e m p e r a t u r eo f t h em u s h -r o o mc i r c l e s ,a n d t e s t e d t h e s o i l s p h y s i c a l a n d c h e m i c a l p r o p e r t i e s ,a sw e l l a s s o i l e n z ym e s .T h e r e s u l t s f o u n d t h a t t h e s o i l t e m p e r a t u r e c h a n g e s o n t h eO N z o n ew e r e s i g n i f i c a n t l y d i f f e r e n t f r o m t h o s e i n s i d e a n d o u t s i d e t h e f a i r y r i n g ,e s pe -c i a l l yf r o m11o c l o c k a t n igh t t o 5o 'c l o c ki n t h em o r n i n g .T h e a v e r a g e s o i l t e m p e r a t u r e o n t h e f a i r y r i n g w a s h i gh e r t h a n t h a t o u t s i d e t h e f a i r y r i n g a n d i n s i d e t h e f a i r y r i n g .W i t h i n o n em o n t h o f d e t e c t i o n ,t h e t e m pe r a t u r e o n t h e c i r c l e a t 5a mw a s h i g h e r b y 0.3t o 1.6d e g r e e sC e l s i u s t h a n t h a t o u t s i d e t h e c i r c l e a n d i n s i d e t h e c i r c l e .U s i n g an i n f r a r e d t e m p e r a t u r e c a m e r a t o c o m p a r e t h e p h y s i c a l a n d c h e m i c a l p r o pe r t i e s of t h e s o i l i n t h em u s h r o o mc i r c l e ,i tw a s f o u n d t h a t t h e t e m p e r a t u r e o f t h e f u ng u s f r u i t i n g b o d y o f th em u s h r o o mci r c l ew a s s i g n i f i c a n t l y h i g h e r t h a n t h e s u r r o u n d i n ge n v i r o n m e n t .Af t e r a n a l y z i ng th e p h y si c a l a n d c h e m i c a l p r o pe r t i e s of t h e s o i l ,i t w a s f o u n d t h a t t h e c o n t e n t o f a v a i l a b l e n u t r i e n t s i n t h em u s h r o o mc i r c l ew a s s ig n i f i c a n t l yhi g h e r t h a n t h a t o u t s i d e t h e i n n e r c i r c l e ,s i g n i f i c a n t l y h i gh e r t h a n I N z o n e a n d o u t z o n e a n dO U Tz o n e .A c i d p h o s p h a t a s e e n z y m e (S -A C P ),s u c r a s e e n z y m e s (S -S C ),c a t a l a s e e n z ym e s (S -C A T )a n d u r e a s e e n z y m e s (S -U E )w e r e d e t e c t e d f r o md i f f e r e n t s i t e s f r o m t h r e e f a i r y r i n g.T h e c a t a l a s e c o n c e n t r a t i o n第3期曹铭等:青海高寒草甸马勃蘑菇圈增温作用的影响研究o n t h e c i r c l ew a s s i g n i f i c a n t l y h i g h e r t h a n t h a t o u t s i d e t h e c i r c l e a n d i n s i d e t h e c i r c l e.T h e s o i l e n z y m e s a c t i v i t y i n I N z o n ew e r e h i g h e r t h a n i n t h eO U T z o n e,i n d i c a t i n g a l o n g-t e r me f f e c t o f m u s h r o o mc i r c l e f u n g i o n s o i l e n z y m e a c t i v i t y.I n c o n c l u s i o n,o u r e x p e r i m e n t a l r e s u l t s s u g g e s t t h a tm u s h r o o mr i n g f u n g i c a n a f f e c t s o i l a v a i l a b l e n u t r i e n t c o n c e n t r a-t i o n,e n z y m e a c t i v i t y a n d s o i l t e m p e r a t u r e.C h a n g e s i n t h e s e f a c t o r sm a y b e i m p o r t a n t f a c t o r s a f f e c t i n g p l a n t g r o w t h. K e y w o r d s:F a i r y r i n g;A l p i n em e a d o w;S o i lw a r m i n g;S o i l e n z y m e s蘑菇圈是大型真菌辐射生长形成的子实体环状带,蘑菇圈会影响周围草地植物的生长,即靠近蘑菇圈的植物生长茂盛,形成 绿草圈 [1]㊂关于蘑菇圈真菌如何影响植物生长,研究者做了相关研究,主流的研究结论是蘑菇圈真菌通过分解土壤有机质,将有机质中养分以可溶性形式释放出来,促进植物吸收矿物质,刺激植物生长[2-3]㊂青藏高原地处世界第三极,草甸植物的生长长期受到低温的限制㊂温度是制约该地区植物生长的重要因素㊂目前高寒草甸蘑菇圈真菌产热对植物生长影响的研究尚未见报道㊂蘑菇圈真菌作为一种草甸上常见的土壤真菌,分解吸收土壤有机质,在这个过程必然会产生热量,前人的检测结果表明蘑菇圈真菌会显著消耗圈上有机碳,导致圈上有机碳与植物地下部分显著低于圈外[5]㊂这说明圈上碳分解的过程要比圈外剧烈㊂而这个过程产生的热量还未能引起相关研究者的重视㊂研究蘑菇圈真菌产生热量对周围微环境的影响对全面了解蘑菇圈微生态互作有重要的意义㊂土壤酶活是衡量土壤环境和肥力变化的重要手段㊂对于探讨蘑菇圈真菌如何影响草地生态系统结构有很重要的研究意义㊂土壤酶是生态循环中重要的 催化剂,没有土壤酶,土壤有机物的转化㊁腐殖质的生成都不会发生㊂土壤酶在复杂的土壤物质转换起到关键的作用,土壤酶活性与土壤养分循环速率紧密相关㊂而土壤酶活对多种环境因子变化十分敏感,如土壤C㊁N㊁P含量㊁土壤温度均会显著影响土壤酶活性[6]㊂土壤微生物碳㊁氮与脲酶㊁蔗糖酶等酶活性显著正相关[7],刘琳等[8]发现模拟增温条件下土壤酶活性有显著增强,增温对土壤纤维素酶㊁过氧化物酶㊁脲酶和磷酸酶活性显著相关㊂因此,本研究以马勃蘑菇圈为研究对象,探寻蘑菇圈真菌生长对土壤温度和酶活的影响,为深入理解蘑菇圈植物㊁真菌㊁土壤之间的关系提供科学依据㊂1材料与方法1.1研究区自然地理概况本研究在海北高寒草地生态系统野外科学观测研究站开展㊂该站地理位置37.48ʎ~37.75ʎN, 101.20ʎ~101.38ʎE,海拔为3200m[17]㊂气候条件表现为只有冷暖季,暖季潮湿多雨,7月份温度达到峰值,最高气温为27.6ħ㊂冷季寒冷干燥,1月份极端最低气温为-37.1ħ㊂年均降水量为580m m,集中在5-8月份,占全年降水总量的80%㊂全年日照时间为2462.7h[9]㊂高寒草甸主要优势种为矮嵩草(K o b r e s i a h u m i l i s)㊁藏嵩草(K o b r e s i a t i b e t i c a)等㊂植被生长期为5-8月,5月份为植被返青期,6-8月为植被生长期,9月份进入枯黄期㊂土壤类型为高寒草甸土,土壤为草毡寒冻雏形土,土层厚度60c m左右[9]㊂1.2样方设置与取样马勃蘑菇圈(L y c o p e r d a l e s f u n g u s f a i r y r i n g)是高寒草甸常见的生态景观,也是研究蘑菇圈大型真菌影响草甸生态学的理想地点㊂海北站蘑菇圈通常半径几米到几十米不等,圈上植物比圈内和圈外生长旺盛,且蘑菇圈上禾本科植物多度显著高于圈外㊂本研究选择三个完整的马勃蘑菇圈(L y c o p e r d a l e s f u n g u s f a i r y r i n g),分别在三个蘑菇圈圈上(O Nz o n e)㊁圈内(I Nz o n e)和圈外(O U Tz o n e)放置Y Y W-S7I N1多参数温度传感器㊂传感器由江苏云与雾科技有限公司生产,测量精度为ʃ0.2ħ,共9个传感器㊂将传感器探针刺入土层深10c m处,进行温度测量,每十五分钟自动记录土温与气温,监测时间从6月15日持续到7月15日;在2023年7月15日使用艾睿天眼T2热成像仪对蘑菇圈子实体进行拍照,艾睿天眼T2热成像仪由北京普利斯特科技有限公司生产,测量精度为ʃ0.3ħ;在三个蘑菇圈圈上㊁圈内㊁圈外随机选择3个50c mˑ50c m的样方(共27个样方),进行样品采集㊂每个样方测定植物群落盖度和物种分盖度,然后将各植株从基部剪下,烘干后测定每种植物的地上生物量㊂完成地上生物量取样后,用土壤钻在样方内随机采集土壤3钻(5c m直径,10c m深)㊂分为两份,一份用于理化性质检测,一份用于土壤酶活测定㊂1.3土壤理化特性的分析土壤p H使用电位计法测量(德国s a r t o r i u s P B-10),土壤总氮(T N%)㊁总碳(T C%)含量应用燃烧法在元素分析仪(V a r i o M a xC N;E l e m e n t a r,G e r m a n y)上测定㊂土壤铵态氮㊁硝态氮含量应用连续流动注射318草 地 学 报第32卷分析系统(C o n t i n u o u s F l o w A n a l y s i s S ys t e m ,C F A )测定㊂方法同陈立红等[10]的相关研究文献㊂1.4 土壤酶活性的测定本研究测定了土壤脲酶活性㊁过氧化氢酶活性㊁蔗糖酶活性㊁酸性磷酸酶活性㊂脲酶活性采用苯酚钠-次氯酸钠比色法测定;蔗糖酶酶活使用3,5-二硝基水杨酸比色法测定;过氧化氢酶采用分光光度法测量;磷酸酶活性采用磷酸苯二钠比色法测定,测定方法见关松荫所著文献[11]㊂2 结果与分析2.1 蘑菇圈生长对植物群落组成和地上生物量的影响蘑菇圈上㊁圈内和圈外植物群落种类组成和地上生物量结果见表1㊂蘑菇圈上生物量最大,达到353.66g ㊃m -2,显著高于圈内(215.73g ㊃m -2)和圈外(255.31g ㊃m -2)㊂圈上㊁圈内和圈外禾本科牧草地上生物量分别为248.98,115.31和142.62g㊃m -2,尤其是垂穗披碱草圈上显著高于圈外,圈外显著高于圈内㊂莎草科牧草地上生物量在圈上㊁圈内和圈外差异不显著㊂圈上㊁圈内和圈外阔叶杂类草生物量分别为98.85,100.42和111.69g㊃m -2,圈上㊁圈外和圈内无显著差异㊂从表1可以看出,蘑菇圈的生长显著提升了圈上地上生物量特别是禾草科植物的生物量㊂但是也注意到,有一些矮小的杂类草在蘑菇圈上是缺失的,比如唐松草(T h a l i c t r u ma l pi n u m ),肉果草(L a n c e a t i b e t i c a ),假龙胆(G e n t i a n e l l am o e n c h ),矮生忍冬(L o n i c e r a m i n u t a ),线叶龙胆(G e n t i a n a f a r r e r i )㊂表1 蘑菇圈上㊁圈内㊁圈外植物群落种类组成及地上生物量T a b l e 1 C o m p o s i t i o no f s p e c i e s a n d t h e a b o v e g r o u n db i o m a s s o n f a i r y r i n g a n d i n s i d e ,o u t s i d e o f f a i r y r i n g单位:g㊃m -2植物种类P l a n t s pe c i e s 圈上生物量B i o m a s s i nO Nz o n e圈内生物量B i o m a s s i n I Nz o n e圈外生物量B i o m a s s i nO U Tz o n e禾草科P o a c e a e 248.98ʃ51.34a115.31ʃ26.03b142.62ʃ34.02b针茅S t i p a c a p i l l a t a 1.43ʃ0.34b3.20ʃ0.89a 2.52ʃ1.45a冷地早熟禾P o a a n n u a 102.21ʃ16.86a 76.46ʃ18.79b 94.45ʃ23.95a b 垂穗披碱草E l ym u s n u t a n s 145.34ʃ26.35a 35.65ʃ5.99b 46.65ʃ9.28b 莎草科C y p e r a c e a e 4.49ʃ0.94a6.19ʃ1.29a 5.02ʃ1.45a 矮嵩草K o b r e s i a h u m i l i s4.49ʃ0.94a 6.19ʃ1.29a 5.02ʃ1.45a 阔叶杂草103.46ʃ19.02a87.27ʃ6.46b101.20ʃ19.76a钝苞雪莲S a u s s u r e a n i g r e s c e n s 33.93ʃ5.02a --青海苜蓿M e d i c a g o a r c h i d u c i s -n i c o l a i 4.48ʃ2.35a2.14ʃ0.42b1.99ʃ0.46b披针叶黄华T h e r m o ps i s l a n c e o l a t a 0.17ʃ0.15b2.64ʃ0.46a 2.67ʃ0.45a 高山豆T ib e t i a h i m a l a ic a 0.59ʃ0.08b 10.03ʃ0.37a10.52ʃ1.90a 麻花艽G e n t i a n a s t r a m i n e a 7.30ʃ0.59b 4.67ʃ0.93c 13.77ʃ1.02a 细叶亚菊A j a n i a t e n u i f o l i a 0.41ʃ0.14b 1.21ʃ0.48a 1.26ʃ0.39a 蓬子菜G a l i u mv e r u m 2.71ʃ0.86a --湿地繁缕S t e l l a r i a u d a 8.71ʃ2.35a1.21ʃ0.64b1.33ʃ0.02b 野决明T h e r m o p s i s l u p i n o i d e s 3.21ʃ2.10a-1.12ʃ0.12b 圆萼刺参M o r i n a c h i n e n s i s 24.54ʃ3.91a 9.21ʃ0.66b18.67ʃ4.09a b 棉毛茛R a n u n c u l u sm e m b r a n a c e u s 0.65ʃ0.06b -1.27ʃ0.66a 钉柱委陵菜P o t e n t i l l a a n s e r i n a 0.43ʃ0.39b 1.23ʃ0.32a 1.51ʃ0.10a 甘肃马先蒿P e d i c u l a r i s k a n s u e n s i s 1.43ʃ0.28b 3.67ʃ0.68a 3.91ʃ0.89a 疏齿银莲花A n e m o n e o b t u s i l o b a 1.26ʃ0.23b 2.77ʃ0.25a 2.61ʃ0.89a 硬毛拉拉藤G a l i u mb o r e a l e v a r .c i l i a t u m-1.33ʃ0.22a 1.65ʃ0.27a 唐松草T h a l i c t r u ma l p i n u m -10.11ʃ0.48a 5.67ʃ0.33b 美丽风毛菊S a u s s u r e a s u pe r b a -4.45ʃ0.93b 6.32ʃ2.79a 肉果草L a n c e a t i b e t i c a -15.71ʃ0.59a 11.34ʃ3.14a 假龙胆G e n t i a n e l l am o e n c h -5.58ʃ0.43a 4.79ʃ1.18a 矮生忍冬L o n i c e r am i n u t a -1.42ʃ0.40a 0.91ʃ0.21b 线叶龙胆G e n t i a n af a r r e r i -8.34ʃ0.42b 10.34ʃ0.85a狭苞紫菀A s t e r f a r r e r i13.74ʃ0.50a2.32ʃ0.28b -总地上生物量A b o v e -gr o u n db i o m a s s 353.66ʃ63.42a215.73ʃ46.78b255.31ʃ54.84b 注:同行不同小写字母代表圈上㊁圈内和圈外生物量差异显著(P <0.05)N o t e :D i f f e r e n t l o w e r c a s e l e t t e r sw i t h i n t h e s a m e r o wi n d i c a t e s i g n i f i c a n t d i f f e r e n c e s o f d i f f e r e n t s pe c i e s b i o m a s s a t t h e 0.05l e v e l 418第3期曹 铭等:青海高寒草甸马勃蘑菇圈增温作用的影响研究2.2 蘑菇圈圈上㊁圈内㊁圈外土壤碳氮磷含量特征蘑菇圈圈内㊁圈上和圈外土壤碳氮磷含量见表2㊂从表中可以看出,圈上土壤总碳含量(2.81g ㊃k g -1)显著低于圈内(3.57m g ㊃k g -1)与圈外(4.00m g ㊃k g -1)㊂土壤总氮和总磷含量在圈内㊁圈上和圈外之间无显著差异性㊂但是圈上土壤速效养分(铵态氮:27.58m g ㊃k g -1;硝态氮20.61m g ㊃k g -1;可溶性磷84.66m g ㊃k g -1)显著高于圈内(铵态氮:13.38m g ㊃k g -1;硝态氮8.40m g ㊃k g -1;可溶性磷65.44m g ㊃k g -1)与圈外(铵态氮:14.81m g k g -1;硝态氮8.78m g ㊃k g -1;可溶性磷72.81m g ㊃k g -1)㊂表2 蘑菇圈圈内㊁圈上㊁圈外土壤碳氮磷含量特征T a b l e 2 s o i l C ,N ,Pc o n t e n t p r o p e r t i e s o f f a i r y r i n g采样位置S t u d y si t e 土壤总碳S o i lT C/g ㊃k g-1土壤总氮S o i lT N/g ㊃k g-1土壤总磷S o i lT P/g k g-1土壤铵态氮S o i lN H +4/m g ㊃k g-1土壤硝态氮S o i lN O -3/m g ㊃k g-1速效磷S o i lA P /m g ㊃k g-1C /N圈内I Nz o n e3.57ʃ0.66a0.32ʃ0.06a1.87ʃ0.18a13.38ʃ4.34b8.40ʃ0.89b65.44ʃ12.04b11.09ʃ1.15a圈上O Nz o n e2.81ʃ0.25b0.28ʃ0.03a 1.86ʃ0.30a 27.58ʃ5.08a 20.61ʃ1.71a84.66ʃ6.39a 10.79ʃ0.72a 圈外O U Tz o n e4.00ʃ1.01a 0.25ʃ0.01a 1.62ʃ0.16a 14.81ʃ3.34b 8.78ʃ1.97b 72.81ʃ3.96b 11.82ʃ0.39a 注:同列不同小写字母代表圈上㊁圈内和圈外土壤碳氮指数差异显著(P <0.05)N o t e :D i f f e r e n t l o w e r c a s e l e t t e r sw i t h i n t h e s a m e c o l u m n i n d i c a t e s i gn i f i c a n t d i f f e r e n c e s a t t h e 0.05l e v e l 2.3 蘑菇圈土壤酶活性特征蘑菇圈土壤酶活特征见图1,蘑菇圈圈内和圈上土壤脲酶活性显著高于圈外㊂土壤酸性磷酸酶㊁蔗糖酶㊁过氧化氢酶活性均表现为圈上显著高于圈外和圈内㊂2.4 蘑菇圈微环境气温和土壤温度特征应用远红外摄像机拍摄的蘑菇圈子实体及周围环境温度照片见图2㊂可以看出:蘑菇圈子实体温度显著高于周围环境温度㊂子实体温度最高为22.3ħ,比周围环境温度平均可高4ħ㊂图1 三个蘑菇圈土壤脲酶㊁酸性磷酸酶㊁过氧化氢酶㊁蔗糖酶活性特征F i g .1 S -U E ,S -A C P ,S -S C ,S -C A Te n z y m e a c t i v i t yi n I Nz o n e ,O Nz o n e a n dO U Tz o n e 注:同行不同小写字母代表圈上㊁圈内和圈外酶活性差异显著(P <0.05)N o t e :D i f f e r e n t l o w e r c a s e l e t t e r s i n d i c a t e s i gn i f i c a n t d i f f e r e n c e s o f d i f f e r e n t t r e a t m e n t s a t t h e 0.05l e v e l 518草 地 学 报第32卷图2 蘑菇圈马勃子实体红外热成像图F i g .2 T h e r m a l i m a g e o fm u s h r o o mo nF a i r y r i n g以2023年7月2日全天土壤温度监测结果为例发现:凌晨到日出前(7:30分前),蘑菇圈土壤平均温度高于圈外与圈内,而7:30分以后蘑菇圈外和圈内土壤温度迅速回升,到下午两点达到峰值,而这一阶段蘑菇圈上土壤温度增温速度没有圈内与圈外快㊂随后太阳逐渐落下,圈上土壤温度下降速度较圈外和圈内缓慢,直至夜晚10:00圈上土壤平均温度明显高于圈内和圈外㊂查看全月每日温度数据,除一些暂时气候异常变化(阵雨,强降温,强风)外,每日蘑菇圈土壤温度变化按照图3所示规律循环变化㊂考虑到清晨日光升温因素的干扰,我们选取30天凌晨5点蘑菇圈圈内㊁圈上㊁圈外气温和土壤温度变化进行研究,发现从6月15日到7月14日凌晨5点气温逐渐上升,蘑菇圈圈上㊁圈内和圈外土壤温度也波动上升(图4),6月15日 7月15日凌晨五点的气温在-4ħ~6ħ,土壤温度在8ħ~13ħ之间波动,但圈上土壤温度始终高于圈外㊁圈内0.3ħ至1.5ħ㊂蘑菇圈圈上土壤温度在夜晚高于圈内与圈外不是偶然事件,而是稳定存在的㊂图3 蘑菇圈圈上㊁圈内㊁圈外土壤温度24小时变化规律F i g .3 S o i l t e m p e r a t u r e o f a l l -d a y va r i a t i o no f I Nz o n e ,O Nz o n e ,O U Tz o n e i n J u l y2图4 蘑菇圈圈内㊁圈上和圈外凌晨5时气温和土壤温度变化特征F i g .4 T h e 5a mt e m pe r a t u r e v a r i a t i o nf r o mJ u n e 15t o J u l y 15i n f a i r y r i n g3 讨论本研究发现蘑菇圈圈上土壤总碳含量显著低于圈内与圈外,推断大量的有机质在蘑菇圈真菌生长过程中被蘑菇圈真菌吸收消耗了,蘑菇圈真菌分解有机质㊁新陈代谢的过程不可避免会产生热量,远红外热成像仪子实体的成像侧面证明蘑菇圈真菌会增加环境温度㊂增温效应对高寒草地植物的影响一直是生态工作者关注的热点之一㊂有相关研究发现地温变化1ħ就会引起植物生长发育和矿物吸收的显著变化[12]㊂M a v s t r o m 等[13]对极地不同地区的植物进行研究发现,在气温低的区域温度是植物生长的主要限制因子,温度较高的地区矿物营养是主要限制因子,增温会显著增加植物的高度与生物量㊂赵艳超等[14]在增温实验中发现青藏高原4个功能群(豆科㊁莎草㊁禾草㊁杂草)地上生物量要显著高于控制组㊂微生物在进行呼吸作用的时候会产生热量,这种热量与微生物的代谢强度和活性息息相关㊂H o l l e s e n 等[15]在格陵兰岛冻土层的6个地点采集了21个天然冻土并发现气候变化和微生物产生的热量促使冻土层融化并加剧了C O 2释放,他们的模型模拟结果显示,土壤温度和碳分解之间存在一种反馈,土壤微生物产生热量的研究在全球增温的大背景下尤为重要㊂温度传感器对蘑菇圈土壤温度24小时监测结果显示:日出后气温逐渐上升,蘑菇圈外和圈内土壤温度上升速率高于圈上,甚至在全天气温最高点,圈上平均土壤温度低于圈外与圈内㊂然而日落后随气温迅速降低,圈外和圈内土壤温度下降速度也比圈上快,到日出前圈上土壤温度高于圈外与圈内㊂造成这618第3期曹铭等:青海高寒草甸马勃蘑菇圈增温作用的影响研究种结果的原因,一方面是由于蘑菇圈圈上植物比圈内圈外茂盛,植物构成保温层,减少土壤热量向空气流失,另一方面,蘑菇圈真菌分解有机质,进行新陈代谢呼吸过程会释放一部分热量,这一部分热量在白天由于相较日光过于微弱,但是在无光照的夜晚对土温的维持是不可忽视的㊂对蘑菇圈30天的土壤温度监测结果看:凌晨5点圈上土壤平均温度要比圈外㊁圈内土壤温度高0.3ħ~1.5ħ,在青藏高原高寒地区,这种夜晚的增温作用不可忽视㊂科学家已经证实了土壤微生物呼吸作用产生的热量对周围环境的增温作用,格陵兰岛冰原上发现冰层下土壤微生物的呼吸作用会显著影响地上冰层与冻土的消融,苔藓植物的生长,微弱的气温升高会 激活 极地土壤微生物结束休眠,对环境造成 正反馈 效用[15]㊂土壤酶活是一种重要的衡量土壤肥力的指标㊂现已在土壤中检测出60多种土壤酶[16],土壤酶活性对土壤中物质循环有重要作用㊂大部分土壤酶来源于土壤微生物,植物和动物也贡献了一部分土壤酶[18]㊂土壤酶能水解大分子有机物质如纤维素㊁蛋白质㊁腐殖质[19]㊂脲酶能促进有机分子中肽键的水解,专门参与土壤含N有机化合物的转化㊂磷酸酶可以促进有机磷化合物的分解,能增加土壤中可溶性磷㊂蔗糖酶促进蔗糖分解为葡萄糖和果糖㊂过氧化氢酶反映土壤有机质氧化程度㊂过氧化氢酶促进过氧化氢的分解,一般与土壤腐殖化强度和有机质积累强度有关㊂本研究结果表明蘑菇圈圈上脲酶㊁蔗糖酶㊁磷酸酶㊁过氧化氢酶活性平均值均高于圈外㊂土壤酶活对植物生长有显著的影响㊂脲酶可以将含氮有机物分解为植物可以利用的速效氮,过氧化氢酶可以促进腐殖质合成,减轻过氧化物对植物的毒害作用,磷酸酶可以将有机磷分解成可利用的可溶性磷酸,这些酶活性增强在一定程度上会促进植物生长㊂4结论高寒草甸马勃蘑菇圈真菌生长提高土壤温度,显著提升土壤脲酶㊁蔗糖酶㊁氧化氢酶㊁磷酸酶的活性与速效养分的含量,促进蘑菇圈圈上植物生长㊂本研究对蘑菇圈环境温度-土壤-植物间互作关系有了新的发现,对高寒草地蘑菇圈绿草环形成机制有了进一步的理解㊂参考文献[1]M I L L E RSL,G O N G L O F F A.F a i r y r i n g s,a s s o c i a t e df u n g i,a n d a s s e s s m e n t o f t h e i r d i s t r ib u t i o n ac r o s s e n v i r o n m e n t a l v a r i-a b l e s u s i n g G I S[J].F u n g a l E c o l o g y,2021,50,101040[2] S HA N T Z H L,P I E M E I S E LR L.F u n g u s f a i r y r i n g s i ne a s t-e r nC o l o r a d oa n dt h e i ref f e c to nv eg e t a t i o n[J].A g r i c u l t u r a lR e s e a r c h,1917,11:191-246[3] Z O T T IM,D EF I L I P P I SF,C E S A R A N O G,e t a l.O n e r i n g t or u l e t h e ma l l:A ne c o s y s t e me n g i n e e r f u n g u s f o s t e r s p l a n t a n d m i c r o b i a ld i v e r s i t y i n a M e d i t e r r a n e a n g r a s s l a n d[J].N e w P h y t o l o g i s t,2020,227(3):884-898[4] C A E S A R-T O N T H A T TC,E S P E L A N DE,C A E S A R AJ,e t a l.E f f e c t s o fA g a r i c u s l i l a c e p s f a i r y r i n g s o ns o i l a g g r e g a t i o na n dm i-c r o b i a l c o m m u n i t y s t r u c t u r e i nr e l a t i o nt o g r o w t hs t i m u l a t i o no fw e s t e r n w h e a t g r a s s(P a s c o p y r u m s m i t h i i)i n E a s t e r n M o n t a n a r a n g e l a n d[J].M i c r o b i a l E c o l o g y,2013,66:120-131[5] F I D A N Z A M A,C I S A RJL,K O S T K ASJ,e t a l.P r e l i m i n a r yi n v e s t i g a t i o no f s o i l c h e m i c a l a n d p h y s i c a l p r o p e r t i e s a s s o c i a t e dw i t h t y p e-I f a i r y r i n g s y m p t o m s i n t u r f g r a s s[J].H y d r o l o g i c a l P r o c e s s e s,2007,21(17):2285-2290[6]裴海昆.不同施肥量对天然草地土壤酶活性的影响[J].青海畜牧兽医杂志,2001,31(2):15-16[7]蔡晓布,钱成,张永清.退化高寒草原土壤生物学性质的变化[J].应用生态学报,2007,18(8):1733-1738[8]刘琳,朱霞,孙庚,等.模拟增温与施肥对高寒草甸土壤酶活性的影响[J].草业科学,2011,28(8):1405-1410[9]中国科学院西北高原生物研究所.中国科学院海北高寒草甸生态系统定位站.中国科学院院刊,2018,33(10):1 [10]陈立红,阎伟,刘健.草原蘑菇圈对牧草长势影响的分析[J].西北植物学报,2002,22(6):1421-1425[11]关松荫.土壤酶及其研究法[M].北京:农业出版社,1986:274-339[12]WA L K E RJ M.O n ed e g r e e i n c r e a s m e n t i ns o i l t e m p e r a t u r ea f f e c t sm a i z e s e e d i n gb e h a v i o r[J].S o i lSc i e n c eS o c i e t y o fA-m e r i c a J o u r n a l,1969,33(5):729-736[13]H A V S T R OM M,C A L L A G H A N T V,J O N A S S O NS.D i f f e r-e n t i a l g r o w t h r e s p o n s e so fC a s s i o p e t e t r a g o n a,a na r c t i cd w a r f-s h r u b,t o e n v i r o n m e n t a l p e r t u r b a t i o n s a m o n g t h r e e c o n t r a s t i n gh i g ha n d s u b a r c t i c s i t e s[J].O i k o s,1993:389-402[14]C H O I JH,O HN I S H IT,Y AMA K AWA Y,e t a l.T h es o u r c eo f F a i r y r i n g s :2-a z a h y p o x a n t h i n e a n d i t sm e t a b o l i t e f o u n d i n an o v e l p u r i n e m e t a b o l i c p a t h w a y i n p l a n t s[J].A n g e w a n d t eC h e m i e I n t e r n a t i o n a l E d i t i o n,2014,53(6):1552-1555[15]H O L L E S E NJ,MA T T H I E S E N H,MO L L E RA,e t a l.P e r m a-f r o s t t h a w i ng i n o r g a n i cA r c t i c s o i l s a c c e l e r a t e d b y g r o u n dh e a tp r o d u c t i o n[J].N a t u r eC l i m a t eC h a n g e,2015,5(6):574-578[16]B U R N SR G,D I C K RP.E n z y m e s i n t h eE n v i r o n m e n t:E c o l o-g y,A c t i v i t y a n dA p p l i c a t i o n s[J].S o i lB i o l o g y a n dB i o c h e m i s-t r y,2004,36(10):1525-1698[17]G R AM S SG,V O I G T K D,B E R GMA N N H.F a c t o r s i n f l u e n-c i n g w a t e rs o l u b i l i t y a nd p l a n ta v a i l a b i l i t y o f m i ne r a lc o m-p o u n d si n t h et r i p a r t i t ef a i r y r i n g s o f M a r a s m i u s o r e a d e s(B o l t.:F r.)F R[J].J o u r n a l o fB a s i c M i c r o b i o l o g y:A nI n t e r-n a t i o n a lJ o u r n a lo nB i o c h e m i s t r y,P h y s i o l o g y,G e n e t i c s,M o r-p h o l o g y,a n dE c o l o g y o fM i c r o o r g a n i s m s,2005,45(1):41-54[18]周礼恺.土壤酶学[M].北京:科学出版社,1989:11-34[19]D I C KRP,B R E A K W E L LDP,T U R C ORF.S o i l e n z y m e a c t i v i t i e sa n db i o d i v e r s i t y m e a s u r e m e n t s a s i n t e g r a t i v em ic r o b i o l o g i c a l i nd i c a-t o r s[J].M e t h o d s f o rA s s e s s i n g S o i lQ u a l i t y,1997,49:247-271[20]D I C KRP.S o i l e n z y m e a c t i v i t i e s a s i n d i c a t o r s o f s o i l q u a l i t y[J].D e-f i n i ng S o i lQ u a l i t y f o r a S u s t a i n a b l eE n v i r o n m e n t,1994,35:107-124(责任编辑彭露茜)718。
增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响

第40卷第13期2020年7月生态学报ACTAECOLOGICASINICAVol.40,No.13Jul.,2020基金项目:国家自然科学基金(41871090,41671105,41571089);中国科学院前沿科学重点研究项目(QYZDJ⁃SSW⁃DQC013)资助收稿日期:2019⁃11⁃12;㊀㊀修订日期:2020⁃05⁃08∗通讯作者Correspondingauthor.E⁃mail:songyanyu@iga.ac.cn,DOI:10.5846/stxb201911122396高思齐,宋艳宇,宋长春,马秀艳,蒋磊.增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响.生态学报,2020,40(13):4617⁃4627.GaoSQ,SongYY,SongCC,MaXY,JiangL.Effectsofwarmingandexogenouscarboninputontheabundanceofkeymicrobialfunctionalgenesofcarbon⁃nitrogencycleinpeatlandsoil.ActaEcologicaSinica,2020,40(13):4617⁃4627.增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响高思齐1,2,宋艳宇1,∗,宋长春1,马秀艳1,蒋㊀磊1,21中国科学院东北地理与农业生态研究所,湿地生态与环境重点实验室,长春㊀1301022中国科学院大学,北京㊀100049摘要:为探讨温度升高和外源碳输入对泥炭地土壤碳氮循环关键微生物的影响,于2017年7月采集多年冻土区泥炭地表层(0 10cm和10 20cm)土壤样品,在10㊁15ħ两个温度下开展为期42d的增温模拟试验,同时设置葡萄糖添加处理,利用荧光定量PCR技术分析泥炭地土壤碳氮循环关键微生物丰度变化,同时分析增温和外源碳输入对泥炭地土壤活性碳组分和无机氮含量的影响㊂结果表明:温度升高可导致北方泥炭地表层土壤微生物丰度以及群落结构变化,0 10cm土壤微生物比10 20cm土壤微生物更加敏感㊂增温条件下微生物首先快速分解活性有机碳,同时温度升高加快土壤氮周转速率,增加有效氮含量㊂外源碳输入整体提高了深层土壤微生物丰度,使得10 20cm土壤细菌㊁产甲烷菌㊁甲烷氧化菌㊁氨氧化细菌以及反硝化细菌丰度显著增加,说明外源碳输入可能会促进10 20cm土壤甲烷氧化过程㊁氨氧化过程和反硝化过程㊂温度和葡萄糖的交互作用对泥炭地表层土壤碳氮循环关键微生物丰度均有显著影响㊂在增温和外源碳输入条件下,北方泥炭地表层土壤微生物丰度受土壤碳氮活性基质的影响㊂关键词:增温;泥炭地;土壤微生物;可利用有机碳;外源碳输入Effectsofwarmingandexogenouscarboninputontheabundanceofkeymicrobialfunctionalgenesofcarbon⁃nitrogencycleinpeatlandsoilGAOSiqi1,2,SONGYanyu1,∗,SONGChangchun1,MAXiuyan1,JIANGLei1,21KeyLaboratoryofWetlandEcologyandEnvironment,NortheastInstituteofGeographyandAgroecology,ChineseAcademyofSences,Changchun130102,China2UniversityofChineseAcademySciences,Beijing100049,ChinaAbstract:Inordertoinvestigatetheeffectsoftemperatureriseandexogenouscarboninputonkeymicroorganismsofcarbonandnitrogencyclinginpeatlandsoil,surfacesoilsamples(0 10cmand10 20cm)ofpeatlandinpermafrostregionwerecollectedinJuly2017.Thesimulationexperimentoftemperatureincreasingwasconductedfor42daysunder10ħand15ħ,andglucoseadditiontreatmentwassetup.FluorescencequantitativePCRtechnologywasusedtoanalyzethechangesofkeymicrobialabundancesinsoilcarbonandnitrogencyclesinpeatlands.Simultaneouslyanalyzetheeffectsofwarmingandexogenouscarboninputonsoilactivatedcarboncomponentsandinorganicnitrogencontentinpeatlands.Theresultsshowedthattheincreasingoftemperaturecouldchangetheabundanceandcommunitystructureofsoilmicroorganismsinnorthernpeatlandssoil.Soilmicroorganismsin0 10cmsoilsweremoresensitivethanthosein10 20cm.Undertheconditionofincreasingtemperature,microorganismsrapidlydecomposedactiveorganiccarbon.Meanwhile,theincreasingoftemperatureacceleratedtheturnoverrateofsoilnitrogenandincreasedtheavailablenitrogencontent.8164㊀生㊀态㊀学㊀报㊀㊀㊀40卷㊀Exogenouscarboninputoverallimprovedthemicrobialabundanceofdeepsoil,whichsignificantlyincreasedtheabundanceofbacteria,methanogens,methanotrophores,ammoxidesanddenitrifiersin10 20cmsoil,andhencepossiblypromotedtheprocessofmethaneoxidation,ammoniaoxidationanddenitrificationin10 20cmsoil.Therewasasignificantinteractiveeffectoftemperatureincreasingandglucoseadditionontheabundanceofkeymicroorganismsinsoilcarbonandnitrogencycleinpeatlandsurfacesoil.Undertheconditionsofincreasingtemperatureandexogenouscarboninput,theabundanceofmicroorganisminnorthernpeatlandssurfacesoilwasinfluencedbytheavailabilityofactivecarbonandnitrogensubstrates.KeyWords:warming;peat;soilmicroorganisms;availableorganiccarbon;exogenouscarboninput土壤微生物是植物⁃土壤物质循环过程中的重要组分[1],对其周围的环境变化具有较强的敏感性,能通过调节其数量㊁功能和种群结构来适应环境变化,对湿地生态系统的健康状况具有指示作用[2]㊂温度和外源碳均是影响微生物代谢的重要因素㊂Song[3]等研究表明,温度和活性基质能通过调节土壤微生物丰度和酶活性对增温的响应,从而影响碳氮耦合关系㊂在气候变暖背景下,温度升高会通过改变北方泥炭地土壤微生物群落结构和代谢途径,进而改变土壤有机碳分解速率[4]㊂而外源碳输入能够通过提高土壤细菌活性,进而促进土壤碳的周转和释放[5]㊂有研究表明,外源碳输入有利于土壤微生物量和呼吸速率的增加,但碳源组成及浓度的不同会影响温室气体排放[6⁃7]㊂葡萄糖是植物根系分泌的最常见的代谢物,也是微生物重要的可溶性有机碳源,进入土壤可以被微生物分解利用[8]㊂可溶性碳的添加有利于土壤微生物呼吸速率的提高以及土壤养分的有效化过程[9]㊂同时,地上植物与地下微生物的竞争可能导致不同土壤深度的微生物丰度不同㊂已有研究表明0 20cm土壤微生物生物量㊁活性和多样性较深层土壤更高[10]㊂温度升高和外源碳输入可能会改变参与土壤碳氮循环相关微生物基因丰度[11],进而影响土壤呼吸,从而导致土壤碳氮循环和温室气体排放的变化[12]㊂土壤细菌㊁产甲烷菌㊁甲烷氧化菌㊁氨氧化细菌以及反硝化细菌是参与土壤碳氮循环的主要功能微生物㊂土壤细菌㊁产甲烷菌和甲烷氧化菌群落在温室气体排放过程中起重要作用[13⁃14],其中编码甲基辅酶M还原酶(mcrA)和微粒甲烷单氧合酶(pmoA)的基因是参与产甲烷过程和甲烷氧化过程的重要基因㊂氨氧化细菌和反硝化细菌是土壤氮循环的重要组成部分,编码氨单加氧酶活性位点多肽(amoA),参与硝化过程[15],亚硝酸还原酶基因nirK和nirS作为反硝化微生物最重要的功能基因参与反硝化过程[16]㊂已有研究表明,微生物丰度的变化对土壤有机碳的分解有重要影响[3]㊂而微生物丰度的变化影响土壤活性有机碳和有效氮含量,土壤活性有机碳和有效氮的变化影响土壤有机质的微生物降解能力,从而影响土壤碳氮循环过程㊂因此,在全球变暖背景下,研究土壤碳氮循环关键微生物功能基因丰度的变化对进一步探讨温度升高影响下土壤碳氮循环的微生物机制具有重要意义㊂泥炭地作为一个重要的碳汇,对陆地生态系统有重要作用,能调控全球气候变化,但随着全球温度的持续升高,北方冻土泥炭地冻融过程发生变化,从而导致泥炭地释放出大量CO2,可能使泥炭地改变其碳汇功能,显著影响该地区碳氮生物地球化学循环过程[17⁃18]㊂因此,本研究通过模拟增温和外源碳输入模拟实验,分析大兴安岭冻土区泥炭地表层土壤在增温和外源碳输入条件下土壤碳氮循环关键微生物功能基因丰度变化及其与土壤碳氮组分的关系,探讨温度升高影响下泥炭地土壤碳㊁氮转化的微生物驱动机制,以期为准确预测北方高纬度泥炭地碳氮平衡对气候变暖的响应提供重要的理论依据㊂1㊀材料与方法1.1㊀研究区概况研究区位于大兴安岭图强林业局奋斗林场(52.94ʎN,122.86ʎE),该区为多年冻土区,属于寒温带季风性气候,海拔约467m,年平均温度(1991 2010年)为-3.9ħ,年平均降雨量为450mm(7 8月占全年降水量的45%)㊂该区主要植被类型为常绿灌木㊁落叶灌木,草本植物[19],以及地被植物,主要植物类型为细叶杜香(Ledumpalustre)㊁笃斯越橘(Vacciniumuliginosum)㊁羊胡子草(Eriophorumvaginatum)㊁泥炭藓(Sphagnumpalustre),属于寒温带针叶林区,土壤类型为泥炭土㊂0 10cm和10 20cm土壤初始总碳含量分别为366.33㊁384.45mg/g,总氮含量分别为15.65㊁18.68mg/g,总磷含量分别为2.74㊁3.43mg/g,溶解性有机碳(DOC)含量分别为620.25㊁338.42μg/g㊂1.2㊀样品采集与培养2017年7月,选取大兴安岭冻土区典型泥炭地,随机设置4块采样地,采样地之间的距离至少为10m㊂在每块采样地随机选取5个采样点,利用土钻在每个采样点采集0 10cm和10 20cm的土壤样品,将样品混合均匀后立即送往实验室㊂人工挑出植物根系㊁新鲜凋落物和石块等杂物,过4mm筛㊂取一部分土样,于阴凉处风干,研磨后,过0.25mm筛,测定土样的初始总碳㊁全氮和全磷,剩余土壤于4ħ下冷藏保存测定溶解有机碳含量和用于培养实验㊂将新鲜土样(相当于10g干重)置于500mL广口瓶中,预培养3天,尽量减少温度突变对微生物活性的影响㊂用保鲜膜密封广口瓶,并扎4个孔,以保持里面的有氧环境,同时减缓水的蒸发损失[20]㊂在10㊁15ħ两个温度下开展为期42d的增温模拟试验㊂同时设置葡萄糖添加处理(添加浓度为1000μgGlu/g干土)和对照两组处理,每一种处理都由来自两层的土壤样品组成,每个处理重复四次㊂实验过程中,每周使用蒸馏水补充挥发水分㊂42d培养结束后,分析测定土壤微生物功能基因丰度和土壤溶解性有机碳㊁微生物量碳㊁铵态氮㊁硝态氮含量㊂1.3㊀样品测定方法按照土壤DNA快速提取试剂盒(FastDNASPIN(MPbio,USA))说明提取0.3g土壤DNA样品[21]㊂提取后用0.5%低熔点琼脂糖凝胶纯化DNA提取物,然后用苯酚⁃氯仿⁃丁醇萃取㊂为了获得更具代表性的DNA样本,对每个样本重复3次处理[22]㊂通过使用荧光定量PCR仪(7500,ABI,美国)定量测定总细菌16SrRNA,mcrA,pmoA,amoA,nirK和nirS基因丰度[3]㊂扩增所用引物见表1,实时荧光定量PCR的反应体系为25μL,包括:12.5μL1ˑSYBR缓冲液(TaKaRa,Japan),0.4μL引物(10μM),0.5μLROXΙΙ(TaKaRa),0.875μL3%BSA,0.625μL二甲亚砜(DMSO),10ngDNA模板㊂表1㊀土壤微生物功能基因引物及扩增程序Table1㊀Primersandamplificationproceduresofsoilmicrobialfunctionalgene目标基因Targetgene引物Primer序列Sequence(5ᶄ⁃3ᶄ)扩增程序Amplificationprocedure参考文献References细菌BacteriaBacteria⁃338FCCTACGGGAGGCAGCAG95ħ2min,35cycles,95ħ30s,60ħ30s,72ħ30s,80ħ15s[22]Bacteria⁃518RATTACCGCGGCTGCTGG产甲烷菌mcrAmlasGGTGGTGTMGGDTTCACMCARTA95ħ3min30s,36cycles,95ħ15s,55ħ30s,72ħ30s,83ħ30s[23]mcrA⁃revCGTTCATBGCGTTVGGRTAGT甲烷氧化菌pmoAA189fGGNGACTGGGACTTCTGG95ħ10min,40cycles,95ħ30s,60ħ30s,72ħ30s,85ħ15s[24]mb661r_ndCCGGMGCAACGTCYTTACC氨氧化细菌amoAamoA1FGGGGTTTCTACTGGTGGT95ħ5min,95ħ30s,55ħ30s,72ħ30s,83ħ15s,40cycles[25]amoA2RCCTCKGSAAAGCCTTCTTC反硝化细菌nirKF1aCuATCATGGTSCTGCCGCG95ħ10min,6touchdowncycles:95ħ15s,63ħ30s(-1ħ),72ħ30s;95ħ15s,58ħ30s,72ħ30s,80ħ30s,35cycles[26]R3CuGCCTCGATCAGRTTGTGGTT反硝化细菌nirScd3aFGTSAACGTSAAGGARACSGG95ħ10min,94ħ1min,57ħ1min,72ħ1min,83ħ30s,40cycles[27]R3cdGASTTCGGRTGSGTCTTGA9164㊀13期㊀㊀㊀高思齐㊀等:增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响㊀0264㊀生㊀态㊀学㊀报㊀㊀㊀40卷㊀㊀㊀利用MultiN/C2100TOC仪(AnalytikJena,德国)采用高温燃烧法测定土壤全碳含量;土壤样品经硫酸湿法消解后,采用AA3连续流动分析仪(SealAnalytical,德国)测定总氮㊁总磷含量[28]㊂采用氯仿熏蒸浸提法测定土壤微生物量碳含量[29],将土壤样品用CHCl3熏蒸24h,用K2SO4溶液(0.5mol/L)振荡30min,分别对熏蒸和非熏蒸土壤样品进行抽提,过滤后采用MultiN/C2100TOC仪(AnalytikJena,德国)测定提取液中碳浓度㊂土壤微生物量碳含量的计算公式:[MBC]=Ec/0.45㊂其中Ec为熏蒸土壤提取的有机碳与未熏蒸土壤提取的有机碳之差[30]㊂按照Ghani等[31]的方法测定土壤溶解性有机碳含量,将土壤样品用去离子水在常温下振荡提取30min,然后以8000r/min的速度高速离心20min,上清液通过0.45μm滤膜过滤后,用MultiN/C2100TOC仪(AnalytikJena,德国)测定溶液中总碳和无机碳浓度,土壤溶解性有机碳含量即为土壤浸提液中总碳与无机碳浓度的差值㊂用2mol/LKCl溶液从土壤样品中提取无机氮,过滤后用AA3连续流动分析仪(SealAnalytical,德国),测定滤液中铵态氮和硝态氮含量㊂1.4㊀数据处理与分析运用SPSS19.0软件,采用双因素方差分析(two⁃wayANOVA)和显著性差异(LSD)以及Pearson系数等方法分别对不同温度和不同葡萄糖添加情况以及不同土壤深度下泥炭地土壤碳氮循环关键微生物基因丰度㊁活性碳组分含量以及无机氮含量进行差异显著分析,检验温度和外源碳输入对土壤碳氮循环相关微生物数量和土壤碳氮含量的影响,并运用Excel2007及minitab进行绘图㊂2㊀结果2.1㊀泥炭地土壤碳氮循环关键微生物丰度变化由荧光定量PCR测得的土壤细菌丰度表明:10 20cm土壤细菌丰度小于0 10cm土壤细菌丰度(图1)㊂温度和外源碳输入对10 20cm土壤细菌丰度均有显著影响(表2),在无外源碳输入的情况下,温度升高使得10 20cm土壤细菌丰度增加了32.77%(图1)㊂在10ħ和15ħ条件下,外源碳输入使得10 20cm土壤细菌丰度分别增加了15.97%和49.79%㊂温度和葡萄糖的交互作用对两种土壤深度的细菌丰度均有显著影响(表2),在增温和外源碳输入条件下,0 10cm和10 20cm土壤细菌丰度分别显著增加了36.28%和71.50%(图1)㊂图1结果表明:10 20cm土壤产甲烷菌(mcrA)丰度大于0 10cm土壤产甲烷菌(mcrA)丰度㊂温度对两种土壤深度的甲烷氧化菌丰度均有显著影响(表2),温度升高使得0 10cm土壤甲烷氧化菌丰度降低了45.22%,10 20cm土壤甲烷氧化菌丰度增加了58.39%㊂葡萄糖添加对10 20cm土壤甲烷氧化菌和产甲烷菌均有显著影响(表2),在10ħ和15ħ条件下,葡萄糖添加使得10 20cm土壤产甲烷菌丰度分别增加了45.02%和22.89%,10 20cm土壤甲烷氧化菌丰度分别增加了39.05%和24.47%,温度和葡萄糖的交互作用对0 10cm土壤产甲烷菌和甲烷氧化菌丰度均有显著影响(表2),在葡萄糖添加条件下,温度增加5ħ导致0 10cm土壤产甲烷菌和甲烷氧化菌丰度分别增加了47.66%和41.79%㊂对于硝化和反硝化细菌,在无增温和外源碳输入条件下,10 20cm土壤硝化和反硝化细菌丰度均小于0 10cm土壤硝化和反硝化细菌丰度㊂温度升高使得0 10cm土壤氨氧化细菌(amoA)丰度显著降低了96.11%,10 20cm土壤氨氧化细菌丰度显著增加㊂葡萄糖添加以及温度和葡萄糖的交互作用对10 20cm土壤氨氧化细菌丰度均有显著影响(表2)㊂在有外源碳的条件下,10ħ时10 20cm土壤氨氧化细菌丰度降低了39.22%,15ħ时10 20cm土壤氨氧化细菌丰度增加了150.16%㊂温度升高导致0 10cm和10 20cm土壤反硝化细菌丰度增加,其中0 10cm土壤nirK和nirS基因丰度分别增加了33.97%和28.75%㊂温度对10 20cm土壤nirS基因丰度有显著影响(表2),温度升高使得10 20cm土壤nirS基因丰度增加了16.43%㊂温度和葡萄糖的交互作用对10 20cm土壤nirS基因丰度有显著影响(表2),在10ħ和15ħ条件下,葡萄糖添加使得10 20cm土壤nirS基因丰度分别增加了22.07%和74.40%㊂在添加葡萄糖的条件下,温度升高使得0 10cm土壤nirS基因丰度降低3.31%,10 20cm土壤nirS基因丰度增加66.35%,葡萄糖添加对10 20cm土壤nirS基因丰度有显著影响(表2)㊂图1㊀温度升高和葡萄糖添加对土壤碳氮循环关键微生物功能基因丰度的影响Fig.1㊀Effectsoftemperatureriseandglucoseadditionongeneabundanceofkeymicrobesinsoilcarbonandnitrogencycling2.2㊀土壤活性有机碳和有效氮含量变化及其与碳氮循环关键微生物丰度的关系在增温和外源碳输入条件下,0 10cm和10 20cm土壤溶解性有机碳含量分别为756.6 847.5μg/g和702.5 862.5μg/g,0 10cm和10 20cm土壤微生物量碳含量分别为4298 4779μg/g和3064 4770μg/g(表3)㊂在无外源碳输入的情况下,10 20cm土壤溶解性有机碳与微生物量碳含量均小于0 10cm土壤溶解性有机碳与微生物量碳含量㊂温度对10 20cm土壤溶解性有机碳和微生物量碳含量有显著影响(表2),温度升高使得10 20cm土壤微生物量碳含量降低了25.16%㊂葡萄糖添加对10 20cm土壤溶解性有机碳和微生物量碳含量有显著影响,在10ħ和15ħ两种温度条件下,葡萄糖添加使得10 20cm土壤溶解性有机碳含量分别呈现下降和上升的趋势,10 20cm土壤微生物量碳含量分别提高了16.49%和46.47%㊂温度和葡萄糖的交互作用对10 20cm土壤溶解性有机碳含量有显著影响,在有外源碳输入的条件下,温度升高5ħ使得10 20cm土壤溶解性有机碳含量增加了22.31%㊂由person相关性分析表明,土壤细菌(Bacteria1264㊀13期㊀㊀㊀高思齐㊀等:增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响㊀16SrRNA)㊁甲烷氧化菌(pmoA)以及反硝化细菌(nirK㊁nirS)丰度均与溶解性有机碳含量呈正相关(P<0.01,P<0.05,表4)㊂表2㊀温度升高和葡萄糖添加对土壤微生物功能基因丰度及碳氮含量影响的方差分析结果Table2㊀Resultsofvarianceanalysisoftheeffectsoftemperatureriseandglucoseadditiononsoilmicrobialfunctionalgeneabundanceandcarbonandnitrogencontent土壤深度Soildepth/cm处理Treatment细菌Bacteria产甲烷菌mcrA甲烷氧化菌pmoA氨氧化细菌amoA反硝化细菌nirK反硝化细菌nirS溶解性有机碳Dissolvedorganiccarbon微生物量碳Microbialbiomasscarbon铵态氮Ammoniumnitrogen硝态氮Nitratenitrogen0 10温度0.0020.0724.928∗81.531∗∗2.7630.5733.8350.811179.169∗∗28.708∗∗葡萄糖4.1986.897∗0.0050.7770.0034.2822.7710.99682.961∗∗13.566∗∗温度ˑ葡萄糖6.956∗8.043∗48.691∗∗0.3750.081.1470.2832.2874.0052.7810 20温度10.875∗∗0.05539.719∗∗132.469∗∗2.89311.818∗∗6.673∗10.168∗∗52.793∗∗45.084∗∗葡萄糖5.473∗5.137∗17.718∗∗28.291∗∗3.68414.71∗∗7.148∗26.053∗∗6.158∗2.846温度ˑ葡萄糖2.043∗0.373040.064∗∗2.5155.189∗7.34∗3.31620.289∗∗7.864∗表3㊀温度升高和葡萄糖添加对土壤碳氮含量影响Table3㊀Effectsoftemperatureriseandglucoseadditiononsoilcarbonandnitrogencontent深度Depth/cm温度Temperature/ħ葡萄糖Glucose溶解性有机碳Dissolvedorganiccarbon/(μg/g)微生物量碳Microbialbiomasscarbon/(μg/g)铵态氮Ammoniumnitrogen/(μg/g)硝态氮Nitratenitrogen/(μg/g)0 1010未加756.56ʃ12.28a4779.17ʃ35.12c397.61ʃ24.35b2.39ʃ0.33a添加785.00ʃ9.03ab4298.47ʃ161.60bc231.61ʃ24.08a1.03ʃ0.38a15未加792.38ʃ43.56ab4317.08ʃ329.67bc756.99ʃ12.31d7.13ʃ1.07a添加847.50ʃ19.75b4415.56ʃ103.22bc497.51ʃ29.29c3.52ʃ0.65a10 2010未加706.25ʃ30.14a4094.44ʃ274.42b873.51ʃ63.09e6.42ʃ1.21a添加705.19ʃ27.73a4769.58ʃ145.43c582.36ʃ43.33c2.98ʃ0.52a15未加702.50ʃ40.56a3064.31ʃ166.73a988.60ʃ29.63f18.46ʃ4.20b添加862.50ʃ14.54b4488.33ʃ211.85bc1072.91ʃ14.57f32.29ʃ4.31c㊀㊀同列不同字母表示植被土壤间差异显著(P<0.05)如表3所示,在增温和添加外源碳的条件下,0 10cm和10 20cm土壤铵态氮含量分别为231.61 756.99μg/g和582.36 1072.91μg/g,0 10cm和10 20cm土壤硝态氮含量分别为1.03 7.13μg/g和2.98 32.29μg/g㊂10 20cm土壤铵态氮和硝态氮含量明显高于0 10cm土壤铵态氮和硝态氮含量㊂温度对两种土壤深度的铵态氮㊁硝态氮含量有显著影响(表2),温度升高使得两种土壤深度的铵态氮含量分别增加了90.39%和13.18%㊂葡萄糖对两种土壤深度的铵态氮以及0 10cm土壤硝态氮均有显著影响(表2),在10ħ和15ħ两种温度条件下,葡萄糖添加使得0 10cm土壤铵态氮含量分别降低了41.75%和34.28%,同时使得0 10cm土壤硝态氮含量分别降低了56.90%和50.63%㊂温度和葡萄糖添加的交互作用对10 20cm土壤铵态氮㊁硝态氮含量有显著影响(表2),在外源碳输入的条件下,温度升高使得10 20cm土壤铵态氮㊁硝态氮含量显著增加,10 20cm土壤铵态氮含量增加了84.23%㊂同时,我们发现,产甲烷菌(mcrA)丰度与铵态氮和硝态氮含量均呈正相关(P<0.01,P<0.05,表4)㊂氨氧化细菌(amoA)丰度与铵态氮含量呈负相关(P<0.01,表4),铵态氮含量与微生物量碳含量呈负相关(P<0.05,表4)㊂2264㊀生㊀态㊀学㊀报㊀㊀㊀40卷㊀表4㊀Person相关性分析Table4㊀Personcorrelationanalysis细菌Bacteria产甲烷菌mcrA甲烷氧化菌pmoA氨氧化细菌amoA反硝化细菌nirK反硝化细菌nirS铵态氮Ammoniumnitrogen硝态氮Nitratenitrogen溶解性有机碳Dissolvedorganiccarbon微生物量碳Microbialbiomasscarbon细菌Bacteria1-0.3210.622∗∗0.2980.423∗0.701∗∗-0.277-0.0050.534∗∗0.21产甲烷菌mcrA㊀1-0.084-0.3480.091-0.358∗0.556∗∗0.523∗∗-0.18-0.087甲烷氧化菌pmoA㊀㊀10.405∗0.426∗0.348-0.2560.1140.454∗∗0.211氨氧化细菌amoA㊀㊀㊀1-0.0020.089-0.575∗∗-0.2120.0760.207反硝化细菌nirK㊀㊀㊀㊀10.490∗∗0.0530.230.520∗∗0.164反硝化细菌nirS㊀㊀㊀㊀㊀1-0.1610.1370.692∗∗0.128铵态氮Ammoniumnitrogen㊀㊀㊀㊀㊀㊀10.788∗∗-0.001-0.396∗硝态氮Nitratenitrogen㊀㊀㊀㊀㊀㊀㊀10.286-0.316溶解性有机碳Dissolvedorganiccarbon㊀㊀㊀㊀㊀㊀㊀㊀10.058微生物量碳Microbialbiomasscarbon㊀㊀㊀㊀㊀㊀㊀㊀㊀1㊀㊀∗∗.在0.01水平(双侧)上显著相关㊂∗.在0.05水平(双侧)上显著相关3㊀讨论3.1㊀土壤微生物对温度升高与外源碳输入的响应本研究发现,在无增温和外源碳添加的情况下,0 10cm土壤细菌㊁甲烷氧化菌以及反硝化细菌丰度高于10 20cm土壤细菌㊁甲烷氧化菌以及反硝化细菌丰度,并且在增温和外源碳输入的情况下,土壤细菌㊁甲烷氧化菌以及反硝化细菌丰度与溶解性有机碳含量呈显著正相关(图1,表4)㊂已有研究表明,10 20cm土壤碳比0 10cm土壤碳更加稳定[32⁃33]㊂鲁博权等[34]研究表明,土壤溶解性有机碳和微生物量碳含量随土壤深度的增加而降低,这与本文结果一致,进一步证实了下层土壤难溶解性有机碳抑制了土壤相关微生物丰度的增长㊂葡萄糖添加使得10 20cm土壤微生物量碳和15ħ条件下土壤溶解性有机碳含量均显著增加㊂对于这一现象,Fontaine等[35]提出了新鲜碳源的缺乏使得深层土壤有机碳稳定㊂倪玉雪等[36]研究表明,加入碳源能够使微生物量显著增加㊂外源碳输入增加了10 20cm土壤活性有机碳含量,进一步促进深层土壤微生物丰度的增长㊂土壤细菌丰度是反映土壤微生物活性的重要指标之一[37]㊂本研究发现温度对土壤细菌有显著影响,温度升高使得0 10cm土壤细菌丰度降低(图1),温度升高会改变土壤细菌的生理和活性[38],也有研究表明温度升高使得碳氮等活性基质有效性降低,从而导致土壤细菌丰度下降[39]㊂Wang等[22]和Hayden等[40]的研究结果与本文一致,细菌丰度会随温度升高而降低㊂但也有其他研究表明,细菌丰度在变暖条件下变化不大或没有变化[41⁃42],这主要与土壤微生物所处的生态系统类型㊁地理位置㊁土壤类型和植物的影响有关[43⁃44]㊂本研究中葡萄糖添加使得10 20cm土壤细菌丰度显著增加,这主要是由于葡萄糖作为可溶性有机碳为土壤微生物提供了充足的可利用的碳源[45]㊂Bastida等[46]也发现可溶性有机碳含量可以影响土壤细菌活性,与本文观点一致,这可能与土壤微生物受碳基质限制有关㊂甲烷是重要的温室气体,产甲烷菌(mcrA)和甲烷氧化菌(pmoA)参与甲烷循环过程[47],对北方高纬度泥炭地生态系统有重要作用㊂Conrad等[48]研究表明,产甲烷菌的丰度和群落结构功能与温度有关㊂本研究中,温度升高导致10 20cm土壤产甲烷菌和甲烷氧化菌丰度增加(图1)㊂Høj等[49]研究表明,温度升高使得产甲烷菌多样性和数量增加,加快产甲烷速率㊂产甲烷菌增加所产生的甲烷可能会导致甲烷氧化菌丰度的增3264㊀13期㊀㊀㊀高思齐㊀等:增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响㊀4264㊀生㊀态㊀学㊀报㊀㊀㊀40卷㊀加[42],进而加速土壤甲烷氧化过程,抑制土壤甲烷的排放[50]㊂葡萄糖添加对产甲烷菌有显著影响㊂Vizza等[51]研究表明,甲烷产生与底物有关,底物充足能促进产甲烷菌丰度增加㊂有外源碳输入的情况下,温度升高有利于土壤产甲烷菌以及甲烷氧化菌丰度的增加,说明温度和葡萄糖的交互作用可能会促进土壤甲烷循环过程㊂土壤氮循环在土壤物质循环过程中占有重要地位,其中硝化和反硝化过程是土壤氮循环的两个重要过程[52]㊂amoA㊁nirK和nirS是研究硝化和反硝化过程的重要功能基因[53⁃55]㊂Fierer等[56]研究表明,温度是影响氨氧化细菌的一个重要因素,温度升高会导致北方泥炭地土壤的氮氧通量增加[57]㊂本研究中,温度升高显著降低了0 10cm土壤氨氧化细菌丰度,显著提高了10 20cm土壤氨氧化细菌丰度,说明温度升高有利于10 20cm土壤氨氧化过程,可能会促进N2O的排放㊂而参与反硝化的0 10cm和10 20cm土壤nirK和nirS功能基因丰度随温度的升高而增加,说明温度升高有利于土壤反硝化过程㊂同时,葡萄糖添加使得0 10cm土壤氨氧化细菌丰度降低,10 20cm土壤氨氧化细菌丰度随温度的升高而增加(图1)㊂可能是此时微生物固定NH+4,或者在可溶性碳浓度较高的情况下,抑制了硝化作用[58]㊂无增温和外源碳输入的情况下,0 10cm土壤氨氧化细菌丰度远远高于10 20cm土壤氨氧化细菌丰度,0 10cm土壤铵态氮含量小于10 20cm土壤铵态氮含量㊂我们的结果表明在增温和外源碳输入的条件下,土壤氨氧化细菌丰度与土壤铵态氮含量呈负相关,说明土壤微生物受有效基质的影响㊂同时本研究也发现葡萄糖添加会导致10 20cm参与反硝化的nirS和nirK功能基因丰度增加㊂朱霞等[9]研究也发现,添加可溶性碳可间接增强土壤反硝化作用㊂葡萄糖添加能够促进微生物的生长和活性,使得产生的NO-3参与到反硝化过程中[59]㊂3.2㊀土壤活性有机碳和有效氮对温度升高和葡萄糖添加的响应土壤溶解性有机碳(DOC)和土壤微生物量碳(MBC)是土壤活性有机碳的重要组分[60⁃61]㊂温度升高导致10 20cm土壤微生物量碳含量显著降低了25.16%,说明增温条件下,微生物首先利用活性碳组分,尤其是在室内培养环境下,微生物快速分解活性有机碳,由于培养试验缺少外来新鲜碳源的供给,温度升高能通过提高微生物活性和分解速率,从而增加微生物对活性碳组分的消耗利用,引起土壤微生物碳限制㊂微生物的代谢活动也会因增温幅度过大而受到抑制,进而导致微生物生物量碳降低[16]㊂可利用有机碳输入显著提高了10 20cm土壤微生物量碳和15ħ条件下溶解性有机碳含量,说明外源碳输入有利于土壤活性有机碳的增加㊂土壤有效氮主要有铵态氮和硝态氮等,是土壤中最易被吸收利用的氮[62],对土壤氮循环有重要意义㊂土壤有效氮的变化能够通过改变碳的生物化学过程而对碳蓄积能力产生显著影响[63]㊂已有研究发现,温度升高加速了氮周转率,从而导致冻土㊁草地和森林土壤中无机氮含量升高[64⁃66]㊂我们的研究结果也发现温度升高显著提高了冻土区泥炭地土壤氨氮和硝氮含量,促进土壤氮周转速率,从而导致有效氮含量增加㊂施瑶等[67]研究发现,氮的有效性越强,土壤微生物活性越高㊂我们也发现产甲烷菌丰度与铵态氮和硝态氮含量均呈正相关,氨氧化细菌丰度与铵态氮含量呈负相关,因此温度升高使得土壤有效氮含量增加,进而间接使得相关微生物丰度发生变化,从而影响泥炭地碳氮平衡㊂而外源碳输入降低了10ħ条件下0 10cm和10 20cm以及15ħ条件下0 10cm土壤氨氮和硝氮含量,这主要是由于碳源的输入增加了微生物对有效氮的吸收和利用,进而导致微生物受氮限制㊂4㊀结论通过研究温度升高和外源碳输入条件下,北方泥炭地土壤微生物碳氮循环关键微生物基因丰度㊁活性碳组分含量以及无机氮含量的变化特征,我们发现:在温度升高和外源碳输入的条件下,0 10cm土壤微生物比10 20cm土壤微生物更加敏感㊂温度升高能够通过改变微生物的生理活性,也可以通过改变碳氮等活性基质间接影响土壤微生物丰度㊂外源碳输入为土壤微生物提供了更多可利用的碳源,整体提高深层土壤微生物丰度,使得10 20cm土壤细菌㊁产甲烷菌㊁甲烷氧化菌㊁氨氧化细菌以及反硝化细菌丰度显著增加㊂有外源碳输入的情况下,温度升高有利于土壤产甲烷菌和甲烷氧化菌丰度以及10 20cm土壤氨氧化细菌丰度增加㊂同时,相关性分析结果表明,在增温和外源碳输入条件下,土壤微生物受碳氮等可利用性活性基质的限制㊂然而,我们的研究结果只是基于短期的室内培养实验,对于北方泥炭地土壤微生物对温度升高的长期响应模式仍需要开展长期的野外监测及模拟研究㊂参考文献(References):[1]㊀王小平,杨雪,杨楠,辛晓静,曲耀冰,赵念席,高玉葆.凋落物多样性及组成对凋落物分解和土壤微生物群落的影响.生态学报,2019,39(17):6264⁃6272.[2]㊀李森森,马大龙,臧淑英,王璐璐,孙弘哲.不同干扰方式下松江湿地土壤微生物群落结构和功能特征.生态学报,2018,38(22):7979⁃7989.[3]㊀SongYY,SongCC,RenJS,MaXY,TanWW,WangXW,GaoJL,HouAX.Short⁃termresponseofthesoilmicrobialabundancesandenzymeactivitiestoexperimentalwarminginaborealpeatlandinNortheastChina.Sustainability,2019,11(3):590.[4]㊀TveitAT,UrichT,FrenzelP,SvenningMM.MetabolicandtrophicinteractionsmodulatemethaneproductionbyArcticpeatmicrobiotainresponsetowarming.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2015,112(19):E2507⁃E2516.[5]㊀WaldropGL,HoldenHM,MauriceMS.TheenzymesofbiotindependentCO2metabolism:Whatstructuresrevealabouttheirreactionmechanisms.ProteinScience,2012,21(11):1597⁃1619.[6]㊀强学彩,袁红莉,高旺盛.秸秆还田量对土壤CO2释放和土壤微生物量的影响.应用生态学报,2004,15(3):469⁃472.[7]㊀KemmittSJ,LanyonCV,WaiteLS,WenQ,AddiscottTM,BirdNRA,OᶄDonnellAG,BrookesPC.Mineralizationofnativesoilorganicmatterisnotregulatedbythesize,activityorcompositionofthesoilmicrobialbiomass anewperspective.SoilBiologyandBiochemistry,2008,40(1):61⁃73.[8]㊀徐鹏,邬磊,胡金丽,蒋梦蝶,赵劲松,胡荣桂.添加葡萄糖㊁乙酸㊁草酸对红壤旱地土壤氮素矿化及反硝化的影响.环境科学学报,2017,37(12):4740⁃4746.[9]㊀朱霞,韩晓增,乔云发,王守宇.外加可溶性碳氮对不同热量带土壤N2O排放的影响.农业环境科学学报,2009,28(12):2637⁃2644.[10]㊀FiererN,SchimelJP,HoldenPA.Variationsinmicrobialcommunitycompositionthroughtwosoildepthprofiles.SoilBiologyandBiochemistry,2003,35(1):167⁃176.[11]㊀XueK,YuanMM,ShiZJ,QinYJ,DengY,ChengL,WuLY,HeZL,VanNostrandJD,BrachoR,NataliS,SchuurEAG,LuoCW,KonstantinidisKT,WangQ,ColeJR,TiedjeJM,LuoYQ,ZhouJZ.Tundrasoilcarbonisvulnerabletorapidmicrobialdecompositionunderclimatewarming.NatureClimateChange,2016,6(6):595⁃600.[12]㊀JungJ,YeomJ,HanJ,KimJ,ParkW.Seasonalchangesinnitrogen⁃cyclegeneabundancesandinbacterialcommunitiesinacidicforestsoils.TheJournalofMicrobiology,2012,50(3):365⁃373.[13]㊀KimSY,FreemanC,FennerN,KangH.Functionalandstructuralresponsesofbacterialandmethanogencommunitiesto3⁃yearwarmingincubationindifferentdepthsofpeatmire.AppliedSoilEcology,2012,57:23⁃30.[14]㊀PeltoniemiK,LaihoR,JuottonenH,BodrossyL,KellDK,MinkkinenK,MakirantaP,MehtataloL,PenttiläT,SiljanenHMP,TuittilaES,TuomivirtaT,FritzeH.Responsesofmethanogenicandmethanotrophiccommunitiestowarminginvaryingmoistureregimesoftwoborealfens.SoilBiologyandBiochemistry,2016,97:144⁃156.[15]㊀KowalchukGA,StephenJR.Ammonia⁃oxidizingbacteria:amodelformolecularmicrobialecology.AnnualReviewofMicrobiology,2001,55:485⁃529.[16]㊀李娜,王根绪,高永恒,王俊峰,柳林安.模拟增温对长江源区高寒草甸土壤养分状况和生物学特性的影响研究.土壤学报,2010,47(6):1214⁃1224.[17]㊀宋长春,宋艳宇,王宪伟,郭跃东,孙丽,张新厚.气候变化下湿地生态系统碳㊁氮循环研究进展.湿地科学,2018,16(3):424⁃431.[18]㊀周文昌,索郎夺尔基,崔丽娟,王义飞,李伟.排水对若尔盖高原泥炭地土壤有机碳储量的影响.生态学报,2016,36(8):2123⁃2132.[19]㊀王娇月,韩耀鹏,宋长春,郗凤明.冻融作用对大兴安岭多年冻土区泥炭地土壤有机碳矿化的影响研究.气候变化研究进展,2018,14(1):59⁃66.[20]㊀蒋磊,宋艳宇,宋长春,任久生,王明全.大兴安岭冻土区泥炭地土壤碳㊁氮含量和酶活性室内模拟研究.湿地科学,2018,16(3):294⁃302.[21]㊀王亚男,曾希柏,王玉忠,白玲玉,李莲芳,吴翠霞,俄胜哲,马世铭.施肥模式对设施菜地根际土壤微生物群落结构和丰度的影响.生态学杂志,2015,34(3):826⁃834.[22]㊀WangH,YangJP,YangSH,YangZC,LvYM.Effectofa10ħ⁃elevatedtemperatureunderdifferentwatercontentsonthemicrobial5264㊀13期㊀㊀㊀高思齐㊀等:增温和外源碳输入对泥炭地土壤碳氮循环关键微生物功能基因丰度的影响㊀。
增温和降水减少对冬小麦和大豆生长季土壤氮素的影响

基金项目:公益性行业 (气象) 科研专项 (重大项目) (GYHY201506001-6) ;江苏省大学生实践创新计划项目 (201610300057; 201510300014Z) ; 作者简介:陈健(1995 年生),男,硕士研究生,主要研究方向为农业气象与生态气象。E-mail: nuistchenjian@ *通信作者,E-mail: zhhu@ 收稿日期:2017-05-30
全球气候变暖是当今非常严峻的环境问题,已 引起世界各国和学术界的广泛关注。IPCC 第五次 评估报告指出,近 100 年(1880—2012)全球地表 平均温度上升了大约 0.85 ℃,陆地比海洋增温更 快 。 1913 年 以 来 , 中 国 地 表 平 均 温 度 上 升 了 0.91 ℃,最近 60 年气温上升尤其明显,平均每 10 年升高 0.23 ℃, 几乎是全球的两倍 (IPCC, 2013) 。 据估计,气候变暖的趋势将会持续下去,到 21 世 纪中期中国平均气温可能升高 1.2~2.0 ℃,21 世纪 末将升高 2.2~4.2 ℃(丁一汇等,2006)。气温持 续升高势必影响农田土壤碳氮循环关键过程(刘艳 等,2013),氮素是重要的生命元素,是生命支持 系统不可替代的主要成分,也是农田土壤的主要养
1108
生态环境学报
表1 Table 1
第 26 卷第 7 期(2017 年 7 月)
1)
随着作物生长季的变化而变化,在北方干旱和半干 (刘微等, 2006) 。 旱地区作物氮吸收以 NO3--N 为主 而在生 小麦在其生长季的前期吸收 NO3 -N 比较多, 长季的后期则吸收 NH4+-N 比较多(谢经荣等, 1996)。有研究表明,增温和降水变化对土壤氮素 有影响。增温使土壤 NH4+-N 含量显著降低(Rinnan et al. , 2009 ),对土壤全氮无显著影响却增加了 NO3--N 含量(陈智等,2010)。土壤水分亏缺会导 致土壤全氮量提高(张步翀等,2007),降水减少 对 NO3--N 无显著影响 (武丹丹等, 使 NH4+-N 降低, 2016)。土壤全氮含量随着年平均温度的增加而降 低,随着年平均降水量的增加而增加(傅华等, 2005)。增温而降水量减少将使土壤底层全氮含量 增加, 增温而降水量不变将使土壤全氮含量减少 (衡 涛等,2011)。另外,增温还会影响土壤氮矿化作 用 (Rustad et al., 2001; Novem Auyeung et al., 2013) 。 然而,在以往研究中,关于增温和降水改变影 响土壤氮素的研究多见于森林和草地,而针对影响 农田土壤氮素的研究大多是把增温和降水变化这 两个因子分开来研究,对这两个因子的复合效应研 究并不多见。然而,增温和降水变化这两个因子并 不独立,往往共同作用,综合影响农田土壤氮素。 本研究利用红外辐射灯管模拟增温,并人工减少降 水量,通过测定冬小麦和大豆生长季土壤氮素的含 量,探讨增温和降水减少对冬小麦和大豆生长季土 壤氮素的影响,对评价气候变化的农业生态响应具 有一定意义,为气候变化背景下作物的栽培和管理 提供一定的科学依据。
东北典型森林土壤呼吸的模拟——IBIS模型的局域化应用

东北典型森林土壤呼吸的模拟——IBIS模型的局域化应用国庆喜;张海燕;王兴昌;王传宽【摘要】集成生物圈模拟器(IBIS)将陆地生态系统的生态学过程与相关的生物物理和生理学过程统一起来,代表了生态系统碳循环模拟模型的研究方向.将IBIS-2.6进行适当改造用于中国东北地区的6种典型森林类型(红松林、落叶松林、杨桦林、硬阔叶林、蒙古栎林和杂木林)的土壤呼吸、根际呼吸和异养呼吸估算,并以实测数据作验证.2004-2005年土壤呼吸、根际呼吸和异养呼吸年通量的模拟结果与实测值吻合较好,模拟偏差变动范围分别为:-5%-21%、-2%-16%和-16%-45%.土壤呼吸模拟值与实测值之间的差异不显著(P>0.05),两者间的相关系数以杂木林最低(0.362)、硬阔叶林最高(0.917).除了春末夏初的土壤呼吸迅速升高过程外,模型能较好的捕捉土壤呼吸的季节动态.研究为IBIS模型的局域化应用奠定了基础,并表明经过改造的IBIS可以用于特定的森林生态系统水平的土壤呼吸模拟估测.【期刊名称】《生态学报》【年(卷),期】2010(030)009【总页数】9页(P2295-2303)【关键词】土壤呼吸;集成生物圈模拟器;模型;温带森林【作者】国庆喜;张海燕;王兴昌;王传宽【作者单位】东北林业大学林学院,哈尔滨,150040;东北林业大学林学院,哈尔滨,150040;东北林业大学林学院,哈尔滨,150040;东北林业大学林学院,哈尔滨,150040【正文语种】中文地下过程对森林碳平衡而言至关重要。
森林土壤碳库占生态系统碳储量的30%—90%[1],根系生物量大约占森林总生物量的4%—41%[2]。
地下部分每年消耗掉35%—80%的光合产物[3-4]。
土壤呼吸作用是生态系统呼吸过程中最重要的部分[5],通常占生态系统呼吸作用的一半以上[6],而在全球尺度上每年释放50—75 Pg C[7],是化石燃料燃烧释放CO2的10倍[8]。
温度和水分对土壤碳、氮转化影响的研究

温度和水分对土壤碳、氮转化影响的研究
土壤是地球上最重要的生物圈之一,它是植物生长的基础,也是地球生命的重要来源。
随着温度和水分的变化,土壤中的微生物活动会发生变化,进而影响土壤中碳和氮的转化。
因此,探究土壤温度和水分对土壤碳和氮转化的影响具有重要的理论意义和应用价值。
土壤中的碳和氮是由微生物代谢产生的,在不同的温度和水分条件下,微生物的代谢活动会发生显著变化。
研究表明,随着温度的升高,土壤中微生物的代谢速率也会增加,这会促进土壤中碳和氮的转化。
然而,如果温度过高,微生物的代谢速率可能会降低,导致土壤碳和氮转化减慢。
因此,土壤温度对土壤碳和氮转化具有一定的影响。
土壤水分也是土壤中碳和氮转化的重要因素。
土壤水分过低会导致土壤中微生物的代谢减慢,从而减缓土壤中碳和氮的转化。
相反,土壤水分过高会导致土壤中微生物的代谢增快,从而促进土壤中碳和氮的转化。
因此,土壤水分也是土壤碳和氮转化的影响,不仅有助于我们更好地理解土壤动态,还可以为我们提供有效的管理方法,从而提高土壤的生产力,促进植物的生长。
在未来的研究中,还有必要进一步探究土壤温度和水分对土壤中碳和氮转化的具体机制,以及这两个因素对土壤中不同类型的碳和氮的影响。
此外,还有必要研究不同类型的土壤在不同的温度和水分条件下的碳和氮转化情况,为我们提供更为丰富的见解。
增温对南亚热带季风常绿阔叶林土壤微生物群落的影响

增温对南亚热带季风常绿阔叶林土壤微生物群落的影响赵建琪;吴建平;张慧玲;熊鑫;赵梦頔;褚国伟;孟泽;周国逸;张德强【摘要】土壤微生物是森林生态系统中重要的分解者,参与生物圈的物质循环和能量流动,对温度变化响应较为敏感.以鼎湖山南亚热带季风常绿阔叶林为研究对象,基于野外增温实验平台,采集0-10 cm和10-20 cm土层的土壤样品,采用磷脂脂肪酸(PLFA)方法并结合土壤理化性质的监测,探究气温上升对土壤微生物群落的影响.结果表明:(1)增温处理使0-10 cm和10-20 cm土层月均温分别显著上升1.24℃和1.17℃,土层湿度变化不显著;(2)增温显著增加了土壤硝氮含量,但对其他理化性质作用不明显;(3)增温组土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)、微生物生物量碳氮比(C/N)以及微生物总磷脂脂肪酸含量与对照组差异不显著;(4)增温显著改变了土壤微生物群落结构,使细菌相对丰度、细菌真菌之比(B/F)以及革兰氏阳性菌革兰氏阴性菌之比(G+/G-)显著增加,降低了真菌和丛枝菌根真菌的相对丰度;(5)进一步分析表明,土壤硝态氮和有机碳是影响土壤微生物群落结构变异的主要因子,两者共同解释了微生物群落结构60.5%的变异度.以上研究结果表明,尽管增温对南亚热带季风常绿阔叶林土壤微生物生物量作用不明显,但可通过对土壤硝氮和土壤有机碳含量的影响引起土壤微生物群落结构及其相对丰度的改变,微生物群落结构和相对丰度的变化又将通过影响微生物对土壤碳氮的同化作用,最终影响土壤的碳氮过程.【期刊名称】《生态环境学报》【年(卷),期】2019(028)005【总页数】9页(P881-889)【关键词】气候变暖;微生物生物量;微生物群落结构;南亚热带森林土壤【作者】赵建琪;吴建平;张慧玲;熊鑫;赵梦頔;褚国伟;孟泽;周国逸;张德强【作者单位】中国科学院华南植物园,广东广州 510650;中国科学院大学,北京100049;中国科学院华南植物园,广东广州 510650;中国科学院大学,北京 100049;中国科学院华南植物园,广东广州 510650;中国科学院大学,北京 100049;中国科学院华南植物园,广东广州 510650;中国科学院大学,北京 100049;中国科学院华南植物园,广东广州 510650;中国科学院大学,北京 100049;中国科学院华南植物园,广东广州 510650;中国科学院华南植物园,广东广州 510650;中国科学院华南植物园,广东广州 510650;中国科学院华南植物园,广东广州 510650【正文语种】中文【中图分类】S718.5;X17以全球变暖为主的气候变化问题一直备受关注。
全球气候变化下土壤呼吸对温度和水分变化的响应特征综述

全球气候变化下土壤呼吸对温度和水分变化的响应特征综述冉漫雪;丁军军;孙东宝;顾峰雪【期刊名称】《中国农业气象》【年(卷),期】2024(45)1【摘要】气候变暖、降水格局变化等是气候变化的主要表现形式,也是影响土壤呼吸主要的非生物因素,探明气象条件(温度、水分)对土壤呼吸影响及作用机制是理解陆地生态系统碳循环的重要内容之一。
本文对近年来国内外学者关于温度和水分对土壤呼吸的影响及机制的研究进展进行综述。
结果表明,(1)气候变暖与土壤呼吸存在正反馈调节,但温度适应性削弱了二者的反馈关系。
增温时长和土壤碳储量不同导致温度对土壤呼吸的影响具有时空差异。
土壤呼吸对温度适应性机制主要包括土壤微生物适应性、底物消耗和土壤矿物质活化等。
(2)降水对土壤呼吸的作用取决于土壤初始水分含量。
当土壤含水量低于萎蔫系数时,降水不仅增加土壤含水量还可促进土壤呼吸,在土壤含水量接近田间持水量时土壤呼吸达到最大值,当土壤含水量达到饱和值时土壤呼吸又会受到抑制。
水分对土壤呼吸影响机制主要为替代效应与阻滞效应、底物供给、微生物胁迫以及根系响应等。
(3)土壤呼吸与土壤温度、水分的耦合关系取决于土壤水热因子配比,当土壤温度成为胁迫因子时,降水引发的土壤水分含量升高对土壤呼吸的激发效应被低温的负面影响所抑制;当土壤水分成为胁迫因子时,气候变暖引发的土壤温度升高对土壤呼吸的促进作用被干旱的负面影响所抵消,进行土壤呼吸研究时需充分考虑土壤温度和水分的交互作用。
为更全面深入地明晰陆地生态系统土壤碳排放扰动因素,未来气候变化下土壤呼吸与环境关系等相关领域研究应为重点方向,一是加强多因素交互作用对土壤呼吸影响的研究,并定量化研究土壤呼吸组分;二是持续关注土壤呼吸对土壤初始温度和温度波动的响应特征,探索生物多样性或群落结构组成对土壤呼吸的影响。
【总页数】11页(P1-11)【作者】冉漫雪;丁军军;孙东宝;顾峰雪【作者单位】中国农业科学院农业环境与可持续发展研究所/农业农村部旱地节水农业重点实验室【正文语种】中文【中图分类】S15【相关文献】1.陆地生态系统土壤呼吸对全球气候变化响应的研究进展2.气候变化背景下陇东塬区麦田土壤水分变化及生产特征研究3.土壤微生物呼吸热适应性与微生物群落及多样性对全球气候变化响应研究4.气候变化条件下宁南山区旱地胡麻土壤水分变化特征及对发育期和产量的影响5.内蒙古克氏针茅草原土壤异养呼吸对土壤温度和水分变化的响应因版权原因,仅展示原文概要,查看原文内容请购买。
积雪变化对陆地生态系统植被特征和土壤碳氮过程的影响

积雪变化对陆地生态系统植被特征和土壤碳氮过程的影响党宁;马望;代泽成;胡玉香;王志瑞;王正文;姜勇;李慧【期刊名称】《生态学报》【年(卷),期】2024(44)1【摘要】在季节性积雪地区,冬季气候变暖导致积雪变薄、积雪不连续、融雪提前及雪盖面积缩小等现象。
然而相较于氮沉降、增温、降水变化等全球变化因子,目前尚缺乏积雪因子对陆地生态系统过程和功能影响的系统报道。
为加深人们对积雪特征变化生态后果的认知,综述了积雪深度和融雪时间变化对植被物候和群落组成、凋落物分解、土壤碳氮过程、温室气体排放和土壤微食物网(土壤动物和微生物)的影响。
由于模拟积雪变化手段不同和复杂的气候、土壤背景,生态系统各要素对积雪特征变化的响应规律存在较大的分异和不确定性。
例如,在未来气候变暖导致积雪变薄和融雪提前情景下,植被物候提前,生长季延长,导致生产力增加和凋落物数量增加,禾草比例减少导致凋落物质量增加,早春温度高刺激微生物活性,凋落物分解速率高,促进土壤碳氮周转过程。
但积雪减少和融雪提前导致的早春低温和夏季干旱也可能引起植被生产力下降,凋落物数量减少质量降低,土壤微生物活性低,分解速率低,从而减缓碳氮周转过程。
此外,积雪特征变化对植被特征和土壤碳氮过程影响相关研究目前还存在以下问题:1)积雪深度和融雪时间对生态系统的影响是否存在交互效应仍缺乏关注,且积雪变化对后续生长季是否存在持续效应也不明确;2)积雪因子对植被、土壤碳氮动态过程和土壤生物的影响,各生态要素研究相对较为独立;3)积雪变化引起对土壤地化循环过程影响的微生物驱动机制缺乏组学数据支撑;4)缺乏遥感手段反演各类影响生态系统功能和过程的积雪参数。
应加强植被群落-土壤碳氮过程-土壤微食物网生态关联研究、基于基因组学的土壤微生物群落组成和生态功能研究和遥感相关技术研究,以期为发展积雪生态学提供参考。
【总页数】18页(P18-35)【作者】党宁;马望;代泽成;胡玉香;王志瑞;王正文;姜勇;李慧【作者单位】中国科学院沈阳应用生态研究所;中国科学院大学;河北大学生命科学学院【正文语种】中文【中图分类】G63【相关文献】1.毛乌素沙地不同植被类型的土壤颗粒分形与土壤碳氮变化特征2.石漠化区植被恢复过程凋落叶分解特征及其对土壤有机碳/氮的影响——以重庆中梁山为例3.气候变化对陆地生态系统土壤有机碳储量变化的影响4.祁连山南坡不同海拔土壤与植被位移后土壤碳氮的短期变化特征5.八宝河流域3种植被类型下土壤碳氮变化特征及其影响因素因版权原因,仅展示原文概要,查看原文内容请购买。
人工灌丛总初级生产力和蒸散对气候变化的响应模拟——以宁夏盐池县荒漠草原区为例

第44卷第8期2024年4月生态学报ACTAECOLOGICASINICAVol.44,No.8Apr.,2024基金项目:宁夏自然科学基金重点项目(2022AAC02011);国家自然科学基金项目(41967027);宁夏地质局财政项目(NXCZ20220203)收稿日期:2023⁃07⁃31;㊀㊀网络出版日期:2024⁃01⁃29∗通讯作者Correspondingauthor.E⁃mail:dult80@qq.comDOI:10.20103/j.stxb.202307311643袁洪艺,杜灵通,乔成龙,王玉霞,薛斌,魏采用,周峰,李明涛.人工灌丛总初级生产力和蒸散对气候变化的响应模拟 以宁夏盐池县荒漠草原区为例.生态学报,2024,44(8):3515⁃3524.YuanHY,DuLT,QiaoCL,WangYX,XueB,WeiCY,ZhouF,LiMT.SimulationofGPPandETresponsetoclimatechangefortheplantedshrubecosystemindesertsteppeareaofYanchicounty,Ningxia.ActaEcologicaSinica,2024,44(8):3515⁃3524.人工灌丛总初级生产力和蒸散对气候变化的响应模拟 以宁夏盐池县荒漠草原区为例袁洪艺1,2,杜灵通1,2,∗,乔成龙1,2,王玉霞1,2,薛㊀斌1,2,魏采用3,周㊀峰4,李明涛41宁夏大学西北土地退化与生态恢复省部共建国家重点实验室培育基地,银川㊀7500212宁夏大学西北退化生态系统恢复与重建教育部重点实验室,银川㊀7500213宁夏回族自治区遥感调查院,银川㊀7500214宁夏回族自治区矿产地质调查院,银川㊀750021摘要:荒漠草原区人工灌丛生态系统的总初级生产力(GPP)和蒸散(ET)如何响应全球气候变化,不仅是全球变化生态学研究的核心问题,也关乎干旱半干旱风沙区生态建设的可持续性㊂利用参数优化后的生物群区生物地球化学循环(Biome⁃BGC)模型和气象环境驱动数据,考虑不同气候变化情景和未来趋势,模拟了盐池荒漠草原区人工灌丛生态系统GPP和ET对气候变化的响应㊂结果表明:(1)增温会显著抑制生态系统的GPP,大幅度的增温(3ħ)会导致GPP急剧下降,但增温对ET的抑制作用非常微弱;(2)降水是限制ET变化的重要因素,相对于增温诱发干旱胁迫所引起的ET小幅下降,降水多寡则更直接地控制着生态系统的ET大小;(3)中国西北地区未来气候的 暖湿化 趋势和大气CO2浓度升高会对荒漠草原区人工灌丛生态系统产生综合驱动效应,增强陆地和大气间的碳水交换通量㊂研究成果可为干旱半干旱区应对全球变化及指导地方政府制定生态保护修复政策提供科学依据㊂关键词:人工灌丛生态系统;生物群区生物地球化学循环(Biome⁃BGC)模型;总初级生产力;蒸散;气候变化;盐池县SimulationofGPPandETresponsetoclimatechangefortheplantedshrubecosystemindesertsteppeareaofYanchicounty,NingxiaYUANHongyi1,2,DULingtong1,2∗,QIAOChenglong1,2,WANGYuxia1,2,XUEBin1,2,WEICaiyong3,ZHOUFeng4,LIMingtao41BreedingBaseforStateKeyLaboratoryofLandDegradationandEcologicalRestorationinNorthwestChina,NingxiaUniversity,Yinchuan750021,China2KeyLaboratoryforRestorationandReconstructionofDegradedEcosysteminNorthwestChinaofMinistryofEducation,NingxiaUniversity,Yinchuan750021,China3NingxiaInstituteofRemoteSensingSurvey,Yinchuan750021,China4NingxiaInstituteofGeologicalandMineralResourcesSurvey,Yinchuan750021,ChinaAbstract:Howthegrossprimaryproductivity(GPP)andevapotranspiration(ET)oftheplantedshrubecosystemindesertsteppeareasrespondstoglobalclimatechangeisnotonlyakeyscientificproblemofglobalchangeecology,butalsorelatestothesustainabilityofregionalecologicalrestorationinaridandsemi⁃aridareas.Consideringdifferentclimatechangescenariosandfuturetrends,thisstudyusedmeteorologicalandenvironmentaldatatodrivetheBiome⁃BGCmodel.TheresponseofGPPandEToftheplantedshrubecosysteminthedesertsteppeareaofYanchicountytoclimatechangewasstudied.Theresultsshowthat:(1)airtemperaturerisingsignificantlyinhibitstheGPPoftheecosystem.High⁃intensity6153㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀warming(3ħ)leadstoasharpdeclineinGPP,buttheinhibitoryeffectofwarmingonETisveryweak.(2)PrecipitationisacriticalfactorlimitingthechangeofET.ComparedwiththedecreaseofETcausedbydroughtstressunderregionalwarming,theamountofprecipitationcontrolstheETᶄsmagnitudeoftheecosystemmoredirectly.(3)ThewarmandhumidclimateandtheincreaseofatmosphericCO2concentrationinthefuturewillhavecomprehensiveeffectsontheecosystemandultimatelypromotethefluxofcarbonandwaterbetweenlandandatmosphere.Theresearchcanprovideascientificbasisforhumantoadapttoglobalchangesinaridandsemi⁃aridareasandforlocalgovernmentstoimplementecologicalprotectionandrestorationpolicies.KeyWords:plantedshrubecosystem;Biome⁃BGCmodel;grossprimaryproductivity;evapotranspiration;climatechange;Yanchicounty当前气候变化对陆地生态系统的影响愈加复杂,一方面因温室气体排放增加引起的气温上升,会改变植物生长和发育的环境条件并引发干旱胁迫[1],另一方面CO2的施肥作用又增强了生态系统的生产力[2],因CO2浓度升高引起的水文循环强迫,又会影响到干旱区生态系统的植被格局与过程[3],最终对生态系统的碳水循环产生深远影响[4 5]㊂干旱半干旱区占全球41%的陆地表面,草地是这一区域中最主要的生态系统类型,强烈的人类活动和气候变化对干旱区草地的生态系统过程影响显著[6]㊂气候变化通过改变植物碳输入和微生物代谢过程来影响草地生态系统的碳储量[7],因此,模拟不同气候变化情景下草地生态系统的碳水通量响应,对理解干旱区生态系统过程的未来变化至关重要[8]㊂近20年来气候学领域的进展为全球变化模拟研究提供了新途径,世界气候研究计划组织的耦合模式比较计划(CMIP)得到了国际社会的高度认可[3],其中,基于CMIP5排放情景模拟未来陆地生态系统的碳水通量是其重要应用领域,如康满春等[9]根据CMIP5中的RCP4.5和RCP6.0排放情景,研究了中国北方杨树人工林碳水通量对气候变化的响应㊂最新发布的CMIP6气候模式在分辨率等方面有进一步提升[10],其基于不同的共享社会经济路径(SSPs)及最新的人为排放趋势,提出了新的预估情景[11],能更好地衡量不同社会经济发展模式与气候变化风险的关系,并在生态系统过程模拟中得到了应用,如孙倩等[12]基于CMIP6气候情景数据驱动Daycent模型,分析了2015 2100年甘肃草地生态系统碳收支未来的时空特征和影响因素;桑春云等[13]利用CMIP6中加拿大地球系统模式(CanESM5)数据分析了三江源地区的作物参考蒸散(ET0)变化特征及在未来不同情景下的变化㊂在沙化㊁退化荒漠草原区,人工种植耐旱灌木进行防风固沙是中国西北重要的生态治理措施,这一措施在荒漠草原区形成的大量人工灌丛成为区域重要的生态系统类型和景观结构,该生态系统受人类活动干扰强烈,已引起科学界广泛关注[14 17]㊂但在全球气候变化背景下,荒漠草原区人工灌丛的碳水循环过程如何变化尚需开展模拟研究,特别是厘清其在不同气候变化模式和社会经济发展路径下的响应规律,对理解该人工生态系统的可持续性和稳定性至关重要,其不仅具有重要的科学意义,而且对干旱半干旱区生态治理具有指导价值㊂为此,本文以宁夏盐池荒漠草原区中间锦鸡儿(Caraganaliouana)人工灌丛为例,利用联合国政府间气候变化专门委员会(IPCC)第六次气候变化评估报告中提出的五种SSPs,结合盐池的降水㊁气温等变化情景,在生物群区生物地球化学循环(Biome⁃BGC)模型中模拟总初级生产力(GPP)和蒸散(ET)对未来气候变化的响应,以期揭示其生态过程的响应机制,为荒漠草原区应对气候变化提供依据㊂1㊀研究区概况及研究数据和方法1.1㊀研究区概况盐池县位于宁夏回族自治区东部,其幅员辽阔,总面积达6769km2,最高海拔1953m,地形由南到北逐渐降低,气候从半湿润区向半干旱区过渡,植被类型从典型草原向荒漠草原过渡[18 19]㊂气候以中温带半干旱大陆性气候为主,盐池气象站记录地近60年来的年平均气温8.3ħ,年平均降水量在297mm[20]㊂因处在华东季风气候的末界,全年降水分布不均,夏秋多而冬春少,其中7 9月的降水量约占全年的62%[15]㊂土壤结构松散,渗透性强,易受侵蚀,南部黄土丘陵区主要以黑垆土㊁黄绵土为主,北部风沙区以风沙土和灰钙土为主[21]㊂盐池县近几十年持续推进防沙治沙㊁造林种草㊁封山育林等生态建设工程,极大地改善了区域植被结构,在荒漠草原区形成约8.9ˑ104hm2以中间锦鸡儿等灌木为优势种的成林人工灌丛景观[19]㊂1.2㊀模型及驱动数据1.2.1㊀Biome⁃BGC模型Biome⁃BGC模型是一种典型的以日尺度为步长的生理生态过程模型[22],能够模拟水㊁碳㊁氮在陆地生态系统的植被㊁凋落物和土壤中的存储与流通过程,模型设计遵循物质与能量守恒定律,即进入系统的物质和能量等于留在系统中的物质和能量加上离开系统的物质和能量,具体模拟原理参见文献[22]㊂模拟过程包括初始化模拟和常规模拟两个阶段,初始化模拟基于设定的生理生态指标,利用大气CO2浓度㊁氮沉降值和研究区气象数据进行循环,直至模型状态变量㊁碳库和氮库等达到平衡态[23],初始化模拟需要需要输入描述站点属性的初始化文件㊁气象数据文件和生理生态参数文件,当模型初始化运行到模拟的年土壤碳储量差异小于0.5gCm-2a-1就达到了平衡态㊂之后运用被模拟时期的气象资料㊁CO2含量年际变化数据以及生态生理参数,来实现生态系统碳㊁氮㊁水的存储和通量模拟㊂1.2.2㊀数据来源Biome⁃BGC模型所需的日尺度气象数据来源于中国气象数据网(http://data.cma.cn/),研究获取了盐池县1986 2018年逐日最高温度㊁最低温度及降雨量㊂利用逐日降水㊁气温数据驱动山地小气候模拟模型(MTCLIM)模型,模拟获得驱动Biome⁃BGC模型所需的其他气象指标,大气CO2浓度数据来源于青海省瓦里关大气本地观测站㊂本文采用PEST参数优化方法和宁夏大学盐池荒漠草原定位研究站的涡度相关观测数据,对Biome⁃BGC模型中的28个生态生理参数进行了优化,各生态生理参数的优化结果及具体取值参照文献[20]㊂1.3㊀气候变化情景模拟本文分别模拟气温升高㊁降水增加与大气CO2浓度升高等单一因子和组合变化对生态系统总初级生产力(GPP)和蒸散(ET)的可能影响,以及预测在IPCC第六次气候变化评估报告中的五种SSPs下研究区GPP和ET未来的变化特征㊂1.3.1㊀增温与降水增加的组合情景模拟方案预计到21世纪末,中国区域气温会增加1.3 5ħ,北方地区的降水量可能将上升5% 15%[24 25]㊂因此,本研究基于1986 2018年的气温㊁降水和大气CO2浓度数据,通过设定气温升高1ħ㊁2ħ㊁3ħ和降水增加5%㊁10%㊁15%的气候变化幅度,由此构建出包括初始情况(T0P0)在内的2个变量组合的16种变化情景(表1),模拟各情景下研究区GPP和ET的变化㊂表1㊀气候变化情景设计7153㊀8期㊀㊀㊀袁洪艺㊀等:人工灌丛总初级生产力和蒸散对气候变化的响应模拟㊀1.3.2㊀未来气候情景设置基于IPCC第六次气候变化评估报告中所提出的五种SSPs[26],模拟未来全球变暖和大气CO2浓度增加对荒漠草原区人工灌丛生态系统GPP和ET的影响㊂在设计模拟情景时,气温与降水以盐池荒漠草原人工灌丛区1986 2018年的实际上升率为基准,每20a设置一个增加,CO2浓度来源于IPCC的预测排放情景(表2)㊂本研究选择五种SSPs中的低排放情景(SSP1⁃1.9)㊁中等排放情景(SSP2⁃4.5)和高排放情景(SSP5⁃8.5),模拟21世纪早期(2021 2040年)㊁中期(2041 2060年)和末期(2081 2100年)荒漠草原区人工灌丛生态系统的GPP与ET变化㊂表2㊀未来气候变化情景设计Table2㊀Scenariosdesignofclimatechangeinfuture年份Year增温/ħTemperatureincreasing降水增加/mmPrecipitationincreasing共享社会经济路径SharedSocio⁃economicPathwaysCO2浓度/(μmol/mol)CO2concentration2021 20400.4949.63SSP1⁃1.9433.79SSP2⁃4.5448.14SSP5⁃8.5458.472041 20600.9899.26SSP1⁃1.9436.94SSP2⁃4.5510.55SSP5⁃8.5573.812081 21001.95198.52SSP1⁃1.9403.95SSP2⁃4.5598.04SSP5⁃8.51012.792㊀结果和分析图1㊀盐池1986—2018年气候因子及CO2浓度变化情况Fig.1㊀ChangesofclimaticfactorsandCO2concentrationinYanchiCountyfrom1986to20182.1㊀盐池荒漠草原区气候变化基本特征盐池荒漠草原人工灌丛区1986 2018年的气温㊁降水及CO2浓度变化特征见图1㊂从中可以看出,研究期内的多年平均气温为12.44ħ,最低值出现在1986年,为11.43ħ,最高值出现在1998年,为13.63ħ,年平均气温呈波动上升趋势,上升率为0.02ħ/a㊂研究期内的多年平均降水量为296.40mm,2011年为丰水年,年降水量达到最高的402.80mm,2000年为极端干旱年份,年降水量仅仅只有160.80mm,与气温变化趋势一样,年降水量也表现出波动增加的趋势,上升率为2.48mm/a㊂由于盐池缺乏大气本底CO2浓度观测,故采用离本地最近的青海省瓦里关大气本底站观测数据来研究本区域CO2浓度变化,1986 2018年间区域CO2浓度呈极显著上升趋势,年增幅为2.02μmol/mol,其中CO2浓度在1986年为340.23μmol/mol,是这一期间的最小值,在2018年达到407.28μmol/mol,为这一期间的最大值㊂可见,在全球变化背景下,盐池荒漠草原人8153㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀工灌丛区表现出了大气CO2浓度持续增高㊁气候向 暖湿化 演变的基本特征㊂2.2㊀GPP和ET对不同增温与降水增加组合情景的响应基于1986 2018年期间的气象数据驱动参数优化后的Biome⁃BGC模型,模拟了当前大气CO2浓度背景下16种增温与降水增加组合情景(含基准情景T0P0)的生态系统GPP和ET(表3)㊂结果可以看出,在基准情景下,盐池荒漠草原区人工灌丛生态系统的GPP年均值为535.56gCm-2a-1,灌木入侵会增强原始荒漠草原生态系统的总初级生产力;ET年均值为293.28mm,ET基本与多年平均降水量持平㊂气温的增高会显著抑制荒漠草原区人工灌丛生态系统的GPP,当气温增加3ħ后,不同降水情景下的GPP平均值为63.91gCm-2a-1,仅为基准情景下的11.9%,可见气温升高的强烈胁迫可能会导致这种干旱区的人工灌丛生态系统生产力彻底毁灭,极有可能导致人工灌丛完全退化㊂气温的增高也会显著抑制荒漠草原区人工灌丛生态系统的ET,但增温导致ET降低的幅度并不大,当气温增加3ħ后,不同降水情景下的ET平均值为305.77mm㊂从不同增温与降水增加的组合情景来看,只有降水不变(P0)且气温增加的情况下,ET年均值才会比基准情景下(T0P0)略微有所降低,其它增温与降水增加组合均会导致ET增加㊂可见,盐池荒漠草原人工灌丛区在当前气候 暖湿化 变化趋势(图1),尤其是降水增加,会促进生态系统的水分消耗的增强㊂表3㊀不同气候变化情景下盐池荒漠草原区人工灌丛GPP和ET的变化Table3㊀ThevariationofGPPandETofplantedshrubinthedesertsteppeofYanchiCountyunderdifferentclimatechangescenarios气候变化情景模拟Climatechangescenariosimulation总初级生产力Grossprimaryproductivity(GPP)蒸散Evapotranspiration(ET)1986 2018年的总值Totalamount/(gC/m2)年均值Annualaverage/(gCm-2a-1)年均值变化量Amountofchange/(gCm-2a-1)1986 2018年的总值Totalamount/mm年均值Annualaverage/(mm/a)年均值变化量Amountofchange/(mm/a)T0P017673.32535.56/9678.33293.28/T0P118251.41553.0717.5110133.59307.0813.80T0P218685.67566.2330.6710580.05320.6127.33T0P318982.43575.2339.6711026.70334.1440.86T1P013414.83406.51-129.059593.82290.72-2.56T1P113507.86409.33-126.2310037.79304.1810.90T1P213366.99405.06-130.5010475.32317.4324.15T1P313704.55415.29-120.2710916.39330.8037.52T2P06270.28190.01-345.559532.19288.85-4.43T2P16268.48189.95-345.619971.05302.158.87T2P26266.28189.89-345.6710392.42314.9221.64T2P36267.85189.93-345.6310802.51327.3534.07T3P02109.3863.92-471.649505.23288.04-5.24T3P12109.1663.91-471.659895.78299.876.59T3P22108.7663.90-471.6610286.20311.7018.42T3P32108.7063.90-471.6610674.10323.4630.18盐池荒漠草原区人工灌丛生态系统的GPP和ET对增温和降水量增加的响应存在差异(图2)㊂GPP明显受控于气温,增温会显著抑制GPP,线性斜率显示,气温每增加1ħ,GPP会降低183.79gCm-2a-1㊂GPP受降水增加的影响微弱,在基准气温情景下(T0),降水量增加15%会导致GPP增加39.67gCm-2a-1,但在气温增加3ħ的情景下(T3),降水量增加不再对GPP产生任何影响,由此可见,随着增温幅度的增大,降水量变化对GPP的影响越来越微弱(图2)㊂与之相反,ET明显受控于降水变化,降水增加会显著增强ET;气温增加只能微弱的抑制ET,线性斜率显示,气温每增加1ħ,ET只降低2.65mm㊂在基准气温情景下(T0),降水量增加15%会导致ET增加40.86mm;在气温增加3ħ的情景下(T3),降水量增加15%依然会导致ET增加35.42mm(图2)㊂盐池荒漠草原区人工灌丛生态系统的生产力受控于气温,全球气候的变暖对这一生态系统9153㊀8期㊀㊀㊀袁洪艺㊀等:人工灌丛总初级生产力和蒸散对气候变化的响应模拟㊀的生产力将会产生致命性打击,极端升温3ħ会摧毁其生物生产过程,导致灌丛生态系统退化;但生态系统的水分消耗受气候变暖的影响不大,这是因为蒸散由土壤蒸发和植物蒸腾共同构成,人工灌丛生产力受损后,地表水分依然能通过土壤蒸发途径输送回大气,即气候变暖会改变人工灌丛生态系统的蒸腾与蒸发比例结构,但对地气水文循环强度的影响不大㊂图2㊀不同增温与降水增加组合情景下的总初级生产力(GPP)㊁蒸散(ET)模拟结果及影响差异Fig.2㊀SimulatedGrossPrimaryProductivity(GPP),Evapotranspiration(ET),andtheirvariationunderdifferentscenarioswithcombinedtemperatureandprecipitationincreasing2.3㊀未来气候变化对人工灌丛生态系统GPP和ET的影响2.3.1㊀未来气候变化对GPP的影响在未来的SSP1⁃1.9㊁SSP2⁃4.5和SSP5⁃8.5气候情景下(表2),盐池荒漠草原区人工灌丛生态系统在21世纪早期(2021 2040年)㊁中期(2041 2060年)和末期(2081 2100年)的年平均GPP基本呈现逐渐递增趋势(图3)㊂在低排放的SSP1⁃1.9情景下,早期GPP年均值与末期GPP年均值比较接近,中期GPP年均值则达到最大值1128.74gCm-2a-1,GPP出现先增后跌的现象㊂在中等排放的SSP2⁃4.5情景下,早期GPP年均值为980.56gCm-2a-1,中期到达1219.54gCm-2a-1,末期达到1362.09gCm-2a-1,GPP逐渐增强㊂在高排放的SSP5⁃8.5情景下,GPP也是逐渐增强,且比同时期中等排放情景下的GPP略高㊂因此,在未来 暖湿化 增温和降水的增加趋势下,中㊁高排放情景均会导致盐池荒漠草原区人工灌丛生态系统GPP的持续增加,而低排放情景则导致这一生态系统的GPP在21世纪中期达到峰值㊂同时期的三种排放情景间相比,高排放情景下盐池荒漠草原植被生产力的增加幅度更大,即在未来降水量增加㊁气温升高的情景下,大气CO2浓度升高会对荒漠草原人工灌丛生态系统带来施肥效应,促进其生产力增强(图3)㊂2021 2100年盐池荒漠草原区人工灌丛生态系统在不同排放情景下的GPP年内变化如图4所示,从中可以看出,不管哪种排放情景和哪一时期,GPP在年内都表现出一个规律的单峰增长形态,其年内变化特征0253㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀与以往研究得出的该区域历史时期GPP年内变化规律一致[20]㊂在4 10月的人工灌丛生长期内,5 9月的GPP相对较高,这几个月是人工灌丛生产力最旺盛的月份,其中8月GPP达到年内最高值㊂在21世纪早期,三种排放情景对GPP的影响差异较小;21世纪中期,不同排放情景下GPP的差异逐渐显现,即人类活动排放强度逐渐开始对人工灌丛生态系统的生产力产生影响;而到21世纪末期,中㊁高排放情景会显著增强GPP,明显高于低排放情景下的各月GPP,即人类活动排放强度对人工灌丛生态系统的生产力影响完全显现㊂图4㊀不同SSPs情景下GPP和ET的年内变化Fig.4㊀MonthlyGPPandETunderdifferentSSPsscenarios2.3.2㊀未来气候变化对ET的影响SSP1⁃1.9㊁SSP2⁃4.5和SSP5⁃8.5气候情景下未来各时期的ET变化规律如图3所示㊂由图可知,盐池荒漠草原区人工灌丛生态系统在21世纪早期㊁中期和末期的年平均ET值基本呈现逐期递增的趋势㊂在21世纪早期,ET在三种排放情景下的年均值模拟结果相差不大,在433.38 434.14mm/a之间;在21世纪中期,三种排放情景下的ET逐渐拉开差距,年均值在522.03 543.40mm/a之间;在21世纪末期,中㊁高排放情景与低排放情景下的ET年均值差异非常明显,高排放情景下的ET年均值比低排放情景下的ET年均值高出118.50mm/a(图3)㊂不同排放情景和不同时期的ET年内变化情况如图4所示,ET在年内也表现出了和GPP一样的变化规律,在4 10月的生长期内数值较高,尤以5 9月最为显著㊂与GPP不同的是ET在7月达到最顶峰,这是因为7月份气温最高,土壤蒸发年内最大,而8月份灌生长最为旺盛,植被生产力最大㊂不同排放情景对21世纪早期的逐月ET影响不大,到了中期开始有所显现,而到了末期则明显会影响逐月ET,这一规律与年尺度上的结果一致㊂综上所知,未来在持续增温和降水增加的趋势下,盐池荒漠草原区人工灌丛生态系统的GPP和ET均会持续增加的态势,虽然增温会抑制GPP,但降水量的成倍强烈增加依然会促进GPP的增加;而人类活动排放强迫对碳水循环的影响需要长时间积累,即在21世纪早期㊁中期和末期的三个阶段,排放强迫对碳水循环的影响会越来越强㊂1253㊀8期㊀㊀㊀袁洪艺㊀等:人工灌丛总初级生产力和蒸散对气候变化的响应模拟㊀2253㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀3㊀讨论3.1㊀增温会抑制碳水循环气候变化通过改变陆地生态系统的碳水循环过程,进而影响到生态系统GPP分布格局,并改变区域水资源分布状况,引发陆地生态系统对气候变化和CO2浓度增加等的进一步反馈㊂现有研究表明,增温会对草地生态系统的碳循环产生影响[27 28],但在不同地区得出结果不尽相同㊂Wu等[29]通过Meta分析得出,增温在大多数情况下对草地生产力产生负面影响㊂但高寒草地的一些研究结果显示,气温升高可以提高草地生产力㊂Mowll等[30]发现增温并不是简单的对草地生产力产生负面影响,其与降水具有强烈的交互作用,可见,草地生态系统的水分条件在很大程度上决定着增温对其生产力是正面影响还是负面影响㊂盐池荒漠草原区由于沙土的保水能力差,增温会进一步加剧人工灌丛生态系统的干旱胁迫,进而对GPP产生抑制作用㊂本研究模拟得出GPP随气温上升而降低,即增温会抑制荒漠草原区人工灌丛生态系统的生产力,当气温增加3ħ后,可能会导致人工灌丛生产力彻底奔溃,引起生态系统结构和功能的彻底退化(表3㊁图2)㊂叶兵等[31]认为气温上升会导致植物叶片内部水汽压的急剧增大,相比于稳定的大气水汽压,两者之间的饱和差会大大增加,进而使得植物的蒸腾作用显著地提高,但在干旱区植物的水分利用策略可能有所不同㊂从模型机理来看,Biome⁃BGC使用Farquhar叶片光合模型模拟GPP,由于受到增温的影响,与光合作用有关的酶活性降低,进而对GPP产生抑制作用㊂而Biome⁃BGC使用Penman-Monteith模型模拟水循环过程,在模拟增温条件下,理论上会引起水汽压亏缺增大,土壤蒸发和冠层截留蒸发的动力增强,但增温又会抑制植物的光合过程,减弱植物蒸腾量,进而导致增温有抑制ET的作用,只是作用非常微弱(图2)㊂增温抑制碳水循环的模拟结果也符合宁夏盐池县的实际,研究区处在半干旱的荒漠草原区,区域气候干旱,多年平均降水量为296.40mm,而模拟的多年ET均值为293.28mm,仅ET耗水就接近降水供给量,人工灌丛生态系统的总耗水量甚至在一些时期会超过供水量[17]㊂在这种极限耗水模式下,增温带来的蒸发动力对其促进作用微弱,反而因增温抑制光合,进而减弱蒸腾,并在一定程度上个抑制了生态系统的总ET㊂虽然Biome⁃BGC模型基于生态㊁生物及气象等领域长期实验得出的规律,对光合㊁分配㊁死亡㊁分解和呼吸等碳循环过程及降水分配㊁植物蒸腾㊁冠层截留蒸发和土壤蒸发等水循环过程进行定量模拟,具有较强的普适性㊂针对宁夏盐池荒漠草原区人工灌丛生态系统的特性,丹杨等[32]实测和改进了Biome⁃BGC模型中部分生理生态参数,袁洪艺等[20]又进一步利用涡度相关通量观测的结果,使用参数估计(PEST)对模型参数进行优化,这些改进极大地提升了模型本地化效果和模拟精度㊂当然,由于模型是对真实生态过程的定量描述,其从本质上无法完全逼近真实生态过程,因此发展和改进模型机理是提高模拟精度的根本路径,如植物⁃土壤⁃大气间碳交换(CEVSA)模型改进生态系统水碳交换关键过程的定量表达方法,在模拟陆地生态系统碳循环过程时有优异的表现[33 34]㊂3.2㊀持续的降水增加会加速碳水循环在干旱地区,降水以及由此产生的土壤水分动态是植被变化的主要限制性因素[35],强降水事件会通过补给土壤水来促进草地GPP[36]㊂Wu等[29]收集了全球85个站点的观测和模拟实验数据,得出降水增加可以增加光合和呼吸作用,加速生态系统碳水循环过程㊂但是,降水增加的强度和多长时间累积会对生态系统碳水循环过程产生质的改变尚有很大不确定性㊂本研究从当前气候背景下的模拟得出,降水增加会增强荒漠草原区人工灌丛生态系统的ET,但在增温胁迫条件(1 3ħ)下,降水增加15%也不会提升生态系统的生产力㊂在当前气温和和降水增加的线性趋势下(图1),经过近一个世纪的持续气候变化,21世纪末期将增温1.31ħ,降水将增加144.24%,这种长时间尺度的降水增加则会彻底加速荒漠草原区人工灌丛生态系统的碳水循环过程,即在未来长期气候变化情景下,降水增加会促进生态系统GPP和ET(图3)㊂从现实情况来看,中国西北地区的 暖湿化 会缓解干旱区的水资源短缺状况,增加土壤可利用水分,增强水资源匮乏区的各类生态系统的生产力,荒漠草原区人工灌丛生态系统也不例外㊂Biome⁃BGC模型所模拟的生态系统蒸散(ET)是蒸发(E)和蒸腾(T)的总和,气候变化对ET的影响实质上是对E和T影响的累积,康满春等[9]通过模拟气候变化对E和T的影响,发现降水增加导致E和T均增加,两者叠加后的ET对降水增加的响应也为正向,这与本研究的结果一致㊂在蒸发动力强盛㊁蒸腾需求大的盐池荒漠草原区,人工灌丛生态系统只有降水和凝结两种水分获取途径,故降水是限制ET的主要原因,降水的增加势必会增强ET,从而加速生态系统水循环强度㊂3.3㊀人为CO2排放增加对碳水循环的影响机制盐池荒漠草原人工灌丛入侵背景下的生态系统碳水循环特征及受气候变化影响已有报道[14 15,17,37],但鲜有考虑大气CO2浓度升高对生态系统碳水循环的影响㊂然而,大气CO2作为植物光合作用的原料,其浓度升高对植物起着 施肥 作用,能够在短期内促进植被生产力和生物量的增加[38]㊂本研究根据CMIP6气候情景,模拟了荒漠草原区人工灌丛生态系统GPP和ET对未来大气CO2浓度升高的响应,从21世纪早期的模拟结果来看,大气CO2浓度升高对碳水循环过程的影响均不明显,但随着气候变化的持续发展,在21世纪中期和末期,碳水循环过程开始显著响应大气CO2浓度升高,这种响应不仅表现年GPP和ET的总量上,也表现在月GPP和ET的变化上(图3㊁图4),也就是说在区域气候持续 暖湿化 的变化背景下,大气CO2浓度升高将成为盐池荒漠草原区人工灌丛碳水循环的另一控制因素㊂本研究发现长期的大气CO2浓度升高会促进生态系统生产力和蒸散耗水的增强,这与前人在不同地区㊁使用不同方法得出的结果一致㊂例如,Luo等利用陆地生物圈模型模拟了大气CO2浓度对青藏高原GPP年际变化和趋势的影响,结果表明,大气CO2浓度上升的施肥效应会增强植物光合作用和生态系统GPP[39 40]㊂在水循环方面,气孔作为蒸散过程中植物与大气进行水碳交换的通道,其开闭程度受控于植物生理和相关环境要素㊂一些植物的气孔导度会随着大气CO2浓度的升高而降低,导致蒸腾减少;也有一些植物总的叶面积会随着大气CO2浓度的升高而增加,从而抵消因气孔导度下降引起的蒸腾减少[40 41]㊂本研究发现,ET对长期的大气CO2浓度升高有响应,但其机制仍然不明㊂一方面,在气候持续 暖湿化 背景下,人工灌丛植被光合生产增强确实加速了植被蒸腾过程;另一方面 暖湿化 导致的降水量增加,引起土壤蒸发加大,这一过程与大气CO2浓度升高趋势同步,但这种同步不存在内在驱动机制㊂4㊀结论基于参数优化后的Biome⁃BGC模型,考虑不同气候变化情景和未来趋势,模拟了盐池荒漠草原区人工灌丛生态系统GPP和ET对气候变化的响应,得出结论如下:(1)增温会显著抑制生态系统的GPP,大幅度的增温(3ħ)会导致GPP急剧下降,但增温对ET的抑制作用非常微弱;(2)降水是限制ET变化的重要因素,相对于增温诱发干旱胁迫所引起的ET小幅下降,降水多寡则更直接地控制着生态系统的ET大小;(3)中国西北地区未来气候的 暖湿化 趋势和大气CO2浓度升高会对荒漠草原区人工灌丛生态系统产生综合影响,并增强生态系统的碳水通量㊂参考文献(References):[1]㊀PiaoSL,WangXH,ParkT,ChenC,LianX,HeYE,BjerkeJW,ChenAP,CiaisP,TømmervikH,NemaniRR,MyneniRB.Characteristics,driversandfeedbacksofglobalgreening.NatureReviewsEarth&Environment,2019,1(1):14⁃27.[2]㊀AhlströmA,RaupachMR,SchurgersG,SmithB,ArnethA,JungM,ReichsteinM,CanadellJG,FriedlingsteinP,JainAK,KatoE,PoulterB,SitchS,StockerBD,ViovyN,WangYP,WiltshireA,ZaehleS,ZengN.Thedominantroleofsemi⁃aridecosystemsinthetrendandvariabilityofthelandCO2sink.Science,2015,348(6237):895⁃899.[3]㊀CuiJP,PiaoSL,HuntingfordC,WangXH,LianX,ChevuturiA,TurnerAG,KoopermanGJ.VegetationforcingmodulatesgloballandmonsoonandwaterresourcesinaCO2⁃enrichedclimate.NatureCommunications,2020,11:5184.[4]㊀ZengZZ,PiaoSL,LiLZX,WangT,CiaisP,LianX,YangYT,MaoJF,ShiXY,MyneniRB.Impactofearthgreeningontheterrestrialwatercycle.JournalofClimate,2018,31(7):2633⁃2650.[5]㊀ReichPB,HobbieSE,LeeTD,RichR,PastoreMA,WormK.Synergisticeffectsoffourclimatechangedriversonterrestrialcarboncycling.NatureGeoscience,2020,13(12):787⁃793.[6]㊀LiCJ,FuBJ,WangS,StringerLC,WangYP,LiZD,LiuYX,ZhouWX.DriversandimpactsofchangesinChinaᶄsdrylands.NatureReviewsEarth&Environment,2021,2(12):858⁃873.[7]㊀BaiYF,CotrufoMF.Grasslandsoilcarbonsequestration:currentunderstanding,challenges,andsolutions.Science,2022,377(6606):603⁃608.3253㊀8期㊀㊀㊀袁洪艺㊀等:人工灌丛总初级生产力和蒸散对气候变化的响应模拟㊀。
全球气候变化与土壤氮循环的关联

全球气候变化与土壤氮循环的关联随着全球气候变化的不断加剧,人们越来越关注气候变化对地球各个方面的影响。
其中,土壤是一个重要的环境要素,具有调节气候、维持生态平衡和支撑农作物生长的重要作用。
而氮循环作为土壤生态系统中至关重要的循环过程之一,与全球气候变化之间存在着密切的关联。
本文将探讨全球气候变化与土壤氮循环的关系,并分析其对生态环境与人类生活的影响。
一、全球气候变化对土壤氮循环的影响全球气候变化对土壤氮循环产生了许多直接和间接的影响。
首先,气温升高会加快土壤中有机质的分解速率,导致土壤中氮素的释放增加,进而影响氮的循环过程。
其次,降水变化对土壤湿度和通气性产生影响,进而改变土壤中微生物的活动水平,从而影响氮的转化和固定过程。
此外,全球气候变化还会导致风速和紫外线辐射的变化,这些因素也会直接或间接地影响土壤氮循环。
二、全球气候变化对土壤氮循环的生态环境影响全球气候变化对土壤氮循环的影响将进一步影响到生态环境。
首先,氮是植物生长的关键营养元素,土壤中氮素的供应与植物的生长和生产力密切相关。
全球气候变化导致土壤中氮素含量的变化,进而影响着植物的生长发育和生态系统的结构与功能。
其次,土壤中过量的氮排入水体会引发水体富营养化问题,导致水质恶化和生态环境破坏。
此外,氮氧化物的排放也会加剧大气污染,对大气环境和人体健康造成危害。
三、全球气候变化对土壤氮循环的农业影响全球气候变化对土壤氮循环的变化也将对农业生产产生重要影响。
首先,气候变化会改变季风气候的分布和强度,进而影响土壤水分状况和温度条件,从而影响植物对土壤中氮元素的吸收和利用效率。
其次,全球气候变化还会引起长期干旱和高温等极端气候事件的增加,这些气候事件将直接影响农作物生长的需水量和氮素的吸收利用能力。
此外,全球气候变化还会改变土壤中的微生物群落结构和功能,进而影响土壤氮转化过程和肥料的利用效率。
综上所述,全球气候变化与土壤氮循环之间存在着密切的关联。
气候变化对土壤碳循环的影响研究

气候变化对土壤碳循环的影响研究随着全球气候变化不断加剧,人们对土壤碳循环的关注度也随之提升。
土壤是地球上最大的碳库之一,其中存储着大量的有机碳。
而气候变化对土壤碳循环的影响将对生态系统的稳定性产生深远的影响。
气候变化对土壤碳循环的首要影响之一是土壤碳的释放。
温暖的气候条件下,有机质分解速度加快,导致更多的碳从土壤中释放到大气中,形成了温室气体,进一步加剧了气候变化。
此外,干旱和洪涝等极端气候事件也会干扰土壤氮循环,破坏土壤有机碳的稳定性,释放更多的碳到大气中。
然而,气候变化也会对土壤碳循环产生间接影响。
例如,气候变暖导致高温,增加土壤排水的需求,这可能导致农业灌溉加剧。
这种过度灌溉可能会导致土壤水分过剩,从而降低土壤氧气含量。
缺氧的土壤条件会加速有机物的分解,进一步释放更多的二氧化碳(CO2)。
此外,持续的气候变化也可能导致农作物种植区域的扩大和转移,这将对土壤碳循环产生更大的影响。
然而,不容忽视的是,土壤在抵抗气候变化中也发挥着积极的作用。
土壤是碳的重要储存库,通过吸收和固定大量的二氧化碳,可以减缓全球变暖的速度。
在自然生态系统中,植物通过光合作用将大气中的二氧化碳转化为有机碳,将一部分有机碳固定在土壤中。
土壤中的微生物也参与到碳循环中,它们通过分解有机物质释放二氧化碳,同时也通过与植物的共生关系促进土壤碳的固定。
因此,保护土壤碳循环的稳定性,对于应对气候变化至关重要。
要实现这一目标,我们首先需要采取措施减少温室气体的排放,以限制气候变化。
同时,合理利用土地资源,实施科学的农业耕作方式,减少土壤有机碳的流失和释放。
采用养分管理和旋转种植等可持续农业实践,有助于增加土壤有机质的含量,提升土壤碳的固定能力。
此外,应加强对土壤微生物的研究,了解其与土壤碳循环的关系,并寻求相应的调控方法。
总之,气候变化对土壤碳循环产生深远影响。
了解这种影响的机制,是为了采取科学有效的措施,保护和管理土壤碳循环,抵御气候变化。
碳排放的环境影响气候变化对农业产量的影响

碳排放的环境影响气候变化对农业产量的影响碳排放的环境影响以及气候变化对农业产量的影响随着人类工业化和现代化的发展,大量的碳排放已成为地球面临的严重问题之一。
碳排放的增加不仅对环境造成了巨大的影响,还对气候变化产生了重要的影响。
气候变化对农业产量的影响更是直接关系到人类的粮食安全和可持续发展。
本文将探讨碳排放的环境影响以及气候变化对农业产量的影响,并提出解决方案。
一、碳排放的环境影响1. 大气负荷的增加:工业和交通等活动导致了大量的二氧化碳和其他温室气体排放到大气中,形成大气负荷的增加。
这些温室气体能够吸收地球表面辐射的一部分,导致地球的温度上升。
2. 气候系统的紊乱:温室气体的增加导致了气候系统的紊乱。
全球变暖引发了海平面上升、极端天气事件的增加、洪涝灾害和干旱等破坏性气候现象的发生。
这些现象对生态系统、动植物种群和人类社会产生了重大影响。
3. 生态平衡的破坏:气候变化导致生态平衡的破坏,一些物种的栖息地消失或变得不适宜生存。
物种的迁徙、繁殖和生态系统的相互作用都将受到严重影响,从而进一步危及生物多样性。
二、气候变化对农业产量的影响1. 气温升高:气候变化导致了气温的升高,高温对农作物生长和发育产生了不利影响。
高温会导致作物的花粉发育异常,降低授粉率和结实率,从而减少农作物的产量。
2. 降水模式变化:气候变化引起了降水模式的变化,使得干旱和洪涝等极端天气事件的频率和强度增加。
长期的干旱会导致作物缺水,使其生长不良;而洪涝会破坏农作物的生长环境,导致作物减产甚至死亡。
3. 病虫害的爆发:气候变暖和气候异常引发了病虫害的爆发。
温暖湿润的气候条件有利于病虫害的繁殖和传播,给农作物产量带来了巨大的损失。
三、应对气候变化对农业产量影响的解决方案1. 科学适应农业技术:开发和应用适应气候变化的农业技术,如改良农作物品种、推广节水灌溉技术、建设温室等。
这些技术能够提高农作物的抗旱性和抗病虫害能力,有助于保证农作物的高产稳产。
气候变化对全球生态系统碳循环的影响

气候变化对全球生态系统碳循环的影响王天华;孟素昕;崔桂善【摘要】气候变化是全球变化的主要表现之一,它对全球陆地生态系统碳循环有着巨大的影响,具体表现在气温和降水两个方面.全球生态系统碳循环有多种影响因子,本文主要阐述NPP和土壤碳呼吸与气温和降水之间的关系.研究表明,气温与NPP 表现出一定的纬度地带性;气候变暖会增加土壤呼吸,不过超过最适温度,土壤呼吸减弱,表现出一定的适应性;随着降水格局的改变,在不同时间尺度和地区上,NPP与降水关系也不同;在一定降水范围内,降水量对土壤呼吸的作用是不同的.水分对土壤呼吸的抑制作用会随降水的增加而减小.【期刊名称】《中国资源综合利用》【年(卷),期】2019(037)001【总页数】4页(P105-108)【关键词】气候变化;生态系统;碳循环;净初级生产力;土壤呼吸【作者】王天华;孟素昕;崔桂善【作者单位】延边大学地理系;延边大学地理系;延边大学地理系;延边大学湿地研究中心,吉林延吉 133000【正文语种】中文【中图分类】Q148工业革命以来,人类活动产生了大量CO2,它是过去一百年气候变暖的重要驱动因子[1]。
当气温升高时,陆地生态系统呈现出一个微弱的碳汇[2]。
当净初级生产力的增加速率小于土壤呼吸速率时,生态系统则从一个微弱的碳汇转变为碳源[3]。
气候变暖同样会影响降水格局,降水量的增加会提高生态系统生产力,降水格局的改变也会直接影响碳循环。
所以,准确量化气候变化对陆地生态系统碳收的影响,既是维持区域生态系统碳收支平衡的关键,也是应对气候变化的科学依据。
植被在对气候变化的响应过程中主要表现为NPP的变化。
NPP 是指绿色植物在单位面积和时间内所累积的有机物数量,可以反映植物对大气二氧化碳的固定能力。
气温在对NPP影响的过程中促进了植物生长,同时也会促进植物呼吸,这是因为呼吸作用同样属于酶促反应。
所以,当气温变暖时,其通过影响植物光合作用,从而影响蒸发和植物呼吸。
第7~8题 碳排放和生态安全-备战2024年6月浙江高考地理真题题号押题
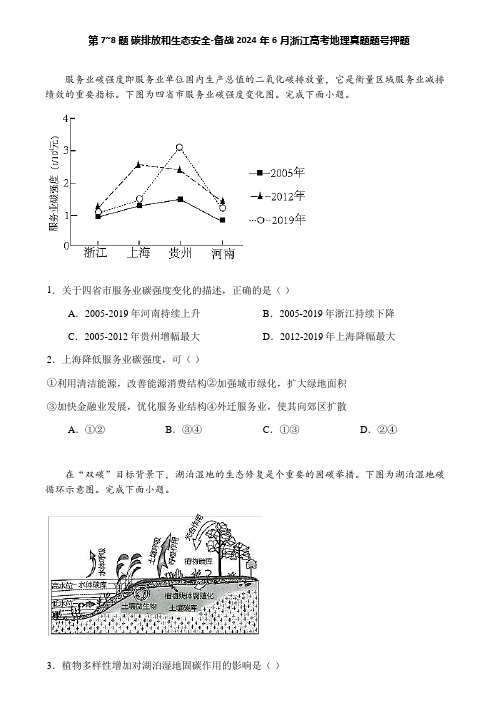
第7~8题碳排放和生态安全-备战2024年6月浙江高考地理真题题号押题服务业碳强度即服务业单位国内生产总值的二氧化碳排放量,它是衡量区域服务业减排绩效的重要指标。
下图为四省市服务业碳强度变化图。
完成下面小题。
1.关于四省市服务业碳强度变化的描述,正确的是()A.2005-2019年河南持续上升B.2005-2019年浙江持续下降C.2005-2012年贵州增幅最大D.2012-2019年上海降幅最大2.上海降低服务业碳强度,可()①利用清洁能源,改善能源消费结构②加强城市绿化,扩大绿地面积③加快金融业发展,优化服务业结构④外迁服务业,使其向郊区扩散A.①②B.③④C.①③D.②④在“双碳”目标背景下,湖泊湿地的生态修复是个重要的固碳举措。
下图为湖泊湿地碳循环示意图。
完成下面小题。
3.植物多样性增加对湖泊湿地固碳作用的影响是()A.土壤碳含量降低B.根系吸碳量减少C.碳净排放量降低D.微生物活性减弱4.下列对湖泊湿地生态修复的措施,合理的是()A.减少湖滨植物,清除入湖污染物B.放生外来物种,增加生物多样性C.降低湖面水位,重建微生物群落D.改变湿地地形,建设生态缓冲岛生态系统生产总值(GEP)是指生态系统能够为人类生活和社会发展提供的最终产品与服务价值的总和,由物质产品、调节服务和文化服务构成,2019年海南热带雨林国家公园GEP 高达2045亿元,其各项价值量占比如图所示。
完成下面小题。
5.属于图中“其它”项目的是()A.农林产品B.科普教育C.休闲旅游D.净化空气6.该公园GEP高的主要原因是()A.土壤肥力高B.科研价值高C.森林面积大D.纬度跨度大人工湿地是建立在生态学基础上,主要利用微生物、植物等对污水进行自然净化的处理技术。
下图为某人工湿地处理微污染水的流程图。
完成下面小题。
7.与污水处理厂相比,该人工湿地系统可以()A.降低污水处理成本B.减少土地占用面积C.不受季节变化影响D.加快污水处理速度8.在湿地保护生态修复中,较为合理的途径是()A.引进外来物种,扩大滩涂种植B.河道种植芦苇,修复水生植物C.增加水域面积,扩大养殖规模D.河道整治清游,清除全部河滩下表为我国某研究机构发布的农田、湿地、森林、草原四种生态系统的单位面积生态服务价值,数值越大表示生态服务价值越高。
草地土壤磷循环及其对全球变化的响应

*通信作者,E -m a i l :l i u js 606@n e n u .e d u .c n 收稿日期:2020-10-06;修回日期:2021-02-25基金项目:国家自然科学基金面上项目 氮沉降背景下放牧对草地土壤线虫的影响及其作用机制 (31971744)作者简介:吴金凤(1996-),女,山东曹县人,在读硕士生,研究方向为草地生态学,E -m a i l :w u jf 046@n e n u .e d u .c n .D O I :10.16742/j .z gc d x b .20200326草地土壤磷循环及其对全球变化的响应吴金凤,刘鞠善*,李梓萌,王德利(植被生态科学教育部重点实验室/东北师范大学草地科学研究所,吉林 长春 130024)摘要:磷是生物体的必需元素,也是草地生态系统生产力仅次于氮的限制元素㊂土壤磷循环主要受地球化学和生物过程控制,并受到全球变化的影响㊂通过从大气氮沉降㊁C O 2浓度升高㊁温度升高㊁降水变化4个全球变化因子及其交互作用对土壤磷循环的影响进行分析,并从土壤理化性质㊁土壤磷酸酶活性和丛枝菌根真菌等途径揭示全球变化影响土壤磷循环的作用机制,指出全球变化能够通过这些作用途径加快土壤磷循环,为提高土壤有效磷含量及全球变化背景下草地可持续利用提供科学的指导㊂最后总结当前研究的不足,即球变化对草地土壤磷循环的影响仍需开展系统深入的研究,并需考虑全球变化与刈割和放牧等人类活动交互作用对土壤磷循环的影响㊂关键词:氮沉降;C O 2浓度升高;温度升高;降水变化;磷循环;草地中图分类号:S 812 文献标志码:A 文章编号:1673-5021(2021)06-0102-10草地是地球上最大的陆地生态系统,约占陆地总面积的25%,我国草地面积为3.31ˑ106k m 2,约占国土面积的41%[1]㊂磷是所有生物的必需元素,在植物生长发育和繁殖过程中发挥关键作用,也是许多生态系统中的主要限制元素[2]㊂磷元素缺乏会限制生态系统净初级生产力㊁氮固定和碳储存[3~4]㊂土壤磷循环是重要的地球化学循环之一,是生态系统养分循环的重要组成部分,对生态系统结构㊁过程和功能具有关键的调节作用㊂土壤磷循环受地球化学和生物过程的控制,易受植被状况㊁土壤类型㊁利用方式以及全球变化等因素的影响[5~6]㊂全球变化是由自然因素或环境因素所驱动的环境变化,已对生态系统产生重要影响[7]㊂近年来由于化石燃料的燃烧和氮肥的使用,使大气氮沉降逐年加剧,同时温室气体排放量的增加导致温度升高和降水格局的变化[8]㊂综合国内外研究表明,氮沉降和C O 2浓度升高会增加土壤氮㊁碳含量,改变土壤NʒP ㊁C ʒP ,而参与土壤磷循环的微生物的主要功能基因的丰度主要由土壤碳氮磷化学计量比决定[9]㊂气温升高通过增加土壤温度改变微生物生物量㊁刺激土壤磷酸酶活性,加速土壤磷循环㊂降水变化主要通过改变土壤湿度和土壤团聚体结构,引起土壤淋失,降低土壤磷含量[10],从而导致土壤磷元素匮乏㊂近年来,已开展不同全球变化因子间交互作用对草地土壤磷循环影响的研究,通过增温和施氮对土壤磷循环影响的研究发现,两因素共同作用减少土壤有效磷的含量,加剧了草地磷限制[11]㊂而施氮和水分添加实验则表明二者相互作用增加土壤中有效磷含量,加快土壤磷循环[10]㊂因此,需要加强全球变化对草地生态系统磷循环过程及影响因素的理解以提高土壤有效磷含量,实现土壤磷元素的可持续利用㊂目前国内外对土壤磷循环的研究比较分散,且对全球变化影响土壤磷循环的机制尚不清楚㊂本文以大气氮沉降㊁C O 2浓度升高㊁温度升高㊁降水变化4个全球变化因子对草地土壤磷循环的影响进行综述,阐释草地土壤磷循环对全球变化的响应机制,为提高土壤有效磷含量及全球变化背景下草地可持续利用提供科学的指导㊂1 草地土壤磷循环磷循环属于沉积型循环,生态系统中的磷元素主要源于矿物岩石的风化作用,最常见的原生磷矿物是磷灰石,而次生磷矿物包括各种磷酸钙和无定形的铝和铁磷酸盐[5]㊂岩石风化中的一部分磷酸盐会随地表径流进入湖泊或海洋中沉积下来㊂由于岩石风化速率和沉积作用缓慢,磷循环在全球尺度上的周转较慢,长达107~108年[12~13]㊂因此,在较短的时间尺度上,草地土壤磷循环主要指磷元素的输入㊁在植物-土壤间的迁移与转化以及在生态系统的输出过程㊂草地土壤磷元素的输入主要包括植物凋落物分解㊁矿物风化和有机磷肥施用等途径,磷元素201 第43卷 第6期V o l .43 N o .6 中 国 草 地 学 报C h i n e s e J o u r n a l o fG r a s s l a n d2021年6月J u n .2021的迁移与转化指土壤中各个磷库之间磷元素形态的转化过程,土壤磷元素的输出指植物对可利用磷的吸收利用以及磷在土壤中的淋溶[13]㊂土壤磷循环主要围绕土壤中各磷库之间磷元素的相互转化以及对植物生长的影响展开研究㊂土壤中的磷可归纳总结为土壤溶液中的磷㊁微生物生物量磷㊁无机磷和非生物有机磷4个磷库,并以有机磷和无机磷两种形式存在[14]㊂其中有机磷占土壤全磷的20%~80%[15],主要包括正磷酸盐单脂和正磷酸盐二脂㊁有机多磷酸盐和磷酸盐等形式[16]㊂有机磷矿化包括物理化学(吸附/解吸和沉淀/溶解)和生物生化(矿化/固定和细胞外酶的水解)两个过程[14]㊂有机磷转化过程中所释放的可溶性磷酸盐,一部分被植物和微生物吸收,另一部分被土壤中的矿质元素固定,即土壤中有机磷矿化与生物固定同时进行[17]㊂微生物和植物通过矿化有机磷㊁吸收磷㊁合成新的有机磷以及通过分泌物影响磷矿物的溶解度来驱动磷循环㊂2全球变化对草地土壤磷循环的作用2.1氮沉降对草地土壤磷循环的影响在陆地生态系统中,大气氮沉降日趋严重,已成为全球变化的重要特征之一㊂在过去150年中,人为活性氮的年输入量增加10倍以上,并预计未来几年大气氮沉降速度将会增加2~3倍[18]㊂氮沉降能够提高草地生产力,改变凋落物分解速率,并通过土壤酸化和提高磷酸酶活性调节土壤有效磷含量,进而影响草地土壤磷库和循环过程㊂氮沉降通过改变凋落物积累量和养分归还影响草地土壤磷输入㊂氮沉降能够增加草地凋落物积累量并提高其分解速率,进而增加草地生态系统的磷元素输入㊂T a n g等[19]对氮添加的56项研究进行m e t a分析发现,在草甸草原上每年施氮量为7.5~ 20.0g/m2时,可以提高植物生产力;高于或低于该阈值时,植物生物量都会降低㊂多数草地生态系统受到氮元素的限制,当施氮量过低时,微生物和非生物过程对氮的竞争超过了植物本身,使植物可利用氮含量减少而降低植物生物量;而高施氮量减少生物量可能是由于过量氮的输入会引起其他因素限制植物生长㊂适量的施氮量可以通过提高土壤可利用氮含量,促进植物生长并提高初级生产力,从而增加草地凋落物积累量[20]㊂赵浩波等[21]通过研究不同氮素水平对羊草生长的影响发现,随着施氮水平的增加,羊草生物量呈现先上升后下降的趋势㊂氮沉降加快营养元素循环,改变凋落物形成过程中的营养元素再分配,增加凋落物中的氮㊁磷含量㊂氮沉降对凋落物分解速率的作用受施氮肥的种类和数量的影响,表现为促进效应[22]㊁抑制效应[23]和没有影响[24]㊂研究表明当凋落物CʒN低于5~15时或CʒP低于200~480时,凋落物氮㊁磷元素出现净释放㊂当凋落物氮㊁磷含量不足时,微生物会固定土壤中的氮㊁磷来维持自身的化学计量平衡[25]㊂因此,氮沉降可通过改变凋落物数量和质量调节草地土壤磷元素输入,其作用效果主要受到氮沉降量的影响㊂氮沉降对土壤磷元素迁移与转化的作用主要表现在对有效磷含量的影响,土壤磷元素只有转化成可利用磷才可以被植物和微生物吸收㊂在多数草地生态系统中,氮沉降可增加酸性磷酸酶的活性,提高土壤有效磷含量[26]㊂这是由于适量的氮添加能够促进植物生长,提高微生物活性,当土壤处于氮饱和时,微生物会将多余的氮用于合成磷酸酶,同时植物根系也会释放磷酸酶加速有机磷矿化过程,提高土壤有效磷含量,加快磷元素养分循环㊂此外,长期大气氮沉降会导致土壤酸化,能够加快被矿物吸附的无机磷的溶解释放,对微生物群落也会产生影响㊂刘红梅等[27]在内蒙古典型草原进行模拟不同梯度氮沉降的研究表明,随着施氮强度的增加,土壤p H 及全磷含量逐渐降低,但有效磷含量逐渐增加㊂这与土壤酸化会溶解释放矿物吸附的无机磷㊁促进迟效状态的磷向有效态包括可溶性磷的转化有关㊂氮沉降对土壤磷元素输出的影响主要表现在影响植物对磷元素的吸收㊂在植物生长初期,大气氮沉降增加土壤中有效氮浓度,提高植物体氮含量和NʒP㊂后期随着氮的持续输入,会引起土壤酸化,降低土壤磷元素矿化速率和有效磷含量,最终提高植物NʒP[28]㊂植物NʒP的增加会导致植物体内养分失衡,引起植物生长的磷限制㊂植物会加快对磷元素的吸收以平衡氮含量㊂L i等[29]在半干旱草地进行氮添加和凋落物对羊草养分重吸收的实验,该研究发现氮添加提高了羊草叶片和茎秆对磷的重吸收能力㊂但随着氮沉降量持续增加,植物叶片内的磷含量会呈现下降趋势,这是因为高浓度氮添加减少了细根生物量,降低植物根冠比[20],从而减少细根对有效磷的吸收㊂氮沉降导致草地生态系统土壤氮饱和,引起草地生态系统中植物和土壤的CʒNʒP化学计量比发生变化㊂植物㊁凋落物和土301吴金凤刘鞠善李梓萌等草地土壤磷循环及其对全球变化的响应壤C ʒNʒP 化学计量比的差异,不仅代表了植物和微生物为维持生态系统平衡而面临的养分竞争关系,还是反映了土壤碳㊁氮㊁磷元素循环的重要指标[30]㊂2.2 C O 2浓度升高对草地土壤磷循环的作用自工业化以来,大气中的C O 2浓度大幅增加,2017年初C O 2浓度为400p pm ,预计2050年将达到550p pm [31]㊂大气中C O 2浓度增加能够改变生态系统的营养循环过程㊂有研究表明,C O 2浓度升高促进植物生长,增加植物磷限制[32]㊂同时,C O 2浓度升高可以增加土壤微生物生物量和碱性磷酸酶活性,提高土壤有效磷含量,促进植物对磷吸收㊂C O 2浓度升高可以通过改变凋落物积累量及其养分归还影响草地土壤磷元素输入㊂C O 2浓度升高促进植物光合作用及生物量积累㊂A n d r e s e n等[33]在温带草原进行长期增加C O 2浓度实验发现,C O 2浓度升高能够显著提高地上生物量㊂C O 2浓度升高产生的施肥效应可以提高植物养分利用率,降低土壤的矿化速率,从而降低凋落物养分含量和分解速率,即养分限制假说[34]㊂但A l l a r d 等[35]研究表明,在高C O 2浓度下草地土壤有机质含量表现出增加趋势㊂这是因为C O 2浓度升高增加植物向地下的碳分配,植物细根增多并提高根部养分吸收能力,同时增加土壤微生物活性并加速土壤养分矿化,从而抵消因C O 2浓度升高引起的养分限制对凋落物分解的不利影响,即C O 2施肥假说[34]㊂C O 2浓度升高对凋落物分解影响持有两种不同的观点,这也影响了凋落物中磷元素向土壤释放的研究㊂C O 2浓度升高主要通过改变土壤有效磷含量影响磷元素的迁移和转化㊂目前C O 2浓度升高对土壤磷元素转化影响的研究相对较少,这些研究趋向于认为为了满足植物生长需求,C O 2浓度升高能够加速磷元素矿化,增加土壤有效磷含量㊂D e n g 等[36]通过m e t a 分析112项研究发现,C O 2浓度升高能够提高磷酸酶活性,从而加快有机磷转化为植物可利用磷的矿化过程,有效缓解土壤有效磷缺乏的状况㊂C O 2浓度主要通过土壤CʒN ㊁有机质和pH 间接影响磷酸酶活性㊂此外,土壤微生物通常受到土壤碳限制而影响磷固持过程,大气C O 2浓度升高可以通过刺激植物生长改变土壤碳库,促进土壤微生物对养分的固定[37],进而提高磷固持能力㊂有研究表明在高C O 2浓度环境下的土壤比周围土壤具有更大的微生物生物量,这是由于大气C O 2浓度升高将产生的额外碳分配给土壤中的微生物生物量,并增加土壤有机质中的碳及其他养分的积累[38]㊂C O 2浓度升高通过影响植物磷吸收调节土壤磷元素的输出,主要指对植物体磷含量的作用,而关于植物对磷的吸收和NʒP 化学计量的影响关注较少㊂C O 2浓度升高会引起植物体内营养元素变化,从而导致C ʒP ㊁NʒP 发生变化㊂C O 2浓度升高能促进植物对磷元素的吸收,降低植物组织NʒP [9]㊂有研究表明随着C O 2浓度升高植物体磷浓度逐渐升高,植物C ʒP 也有所提高[39]㊂这是由于C O 2是植物光合作用的底物,而磷是光合作用的重要参与者,植物需要相对更多富含磷的R N A 支持蛋白质快速合成以满足植物生长需要[40],生长快速的植物体内会具有更高的磷含量㊂2.3 温度升高对草地土壤磷循环的影响近100年来,全球地表温度升高0.74ʃ0.18ħ,I P C C 预测到2100年全球气温将上升1.4~5.8ħ[41]㊂温度升高能提高植物光合作用效率及植物生产力[42],同时也会加速水分流失,引起植物干旱胁迫[43]㊂这些作用会通过影响微生物活性和土壤酶活性影响草地生态系统磷循环㊂温度升高对土壤磷元素输入的影响主要表现为对凋落物产量及其分解速率的作用㊂温度升高可通过促进植物光合作用,增加植物生物量[44],也会由于温度升高提高蒸腾作用和加快土壤水分流失,抑制植物生长,降低生物量[43]㊂草地凋落物分解是草地生态系统养分循环的关键环节,对调节土壤养分可利用性具有重要作用[45]㊂温度是影响凋落物分解的重要环境因子,温度升高能够提高微生物活性和分解酶活性,促进土壤有机质分解,加快凋落物的分解速率进而加速磷元素的返还[46]㊂而林巧燕等[47]进行增温对高寒草甸凋落物质量变化影响的实验发现,增温显著促进了凋落物的分解速率并提高了凋落物中的磷含量,这表明温度升高增加了凋落物中磷元素的固定㊂在未来温度升高的条件下,有可能会导致草地生态系统的磷缺乏,进而引起草地磷限制㊂温度升高能够通过改变磷酸酶活性和微生物生物量磷影响土壤磷元素的迁移与转化过程,进而改变植物和土壤中的有效磷含量㊂研究表明增温处理显著提高磷酸酶活性,即温度升高加快有机质的分解引起磷酸酶活性增强,磷酸酶活性的增强促进有401 中国草地学报 2021年 第43卷 第6期机磷矿化,提高有效磷含量[48]㊂G o n g等[49]研究发现增温处理提高土壤总磷含量,这是因为温度升高降低了土壤含水量和磷的扩散性,减少植物和微生物对磷元素的吸收㊂此外,温度升高还会影响土壤微生物活性,土壤微生物生物量磷的周转期较短,且能够释放活性态的磷元素,可被看作土壤磷元素转化的中转站和植物有效磷的主要来源[50]㊂在目前温度升高对微生物生物量磷的研究中,一部分研究者认为增温处理降低了微生物生物量磷[49]㊂另一部分研究发现增温处理对微生物生物量磷没有显著影响[51]㊂导致结果不一致的原因可能是由于增温处理的时间长短不同,长期增温会降低土壤湿度,通过减缓磷元素解吸和溶解过程减少对微生物的磷元素供应㊂温度升高能够通过改变植物养分吸收影响磷元素输出,温度升高对植物磷元素吸收的影响可能与植物自身特性和土壤性质有关㊂大部分研究表明温度升高会降低植物磷含量,并提高磷元素吸收效率㊂Y a n g等[52]在青藏高原冻土区研究高山莎草叶片对增温的响应发现,适度的增温降低了叶片磷浓度,更大程度的增温会增加根系磷浓度㊂这是因为温度升高促进土壤有机质释放磷元素,提高磷元素吸收效率,植物有效磷含量增加㊂而R e n等[53]在荒漠草地研究干㊁湿年份养分吸收对增温的响应,结果发现在湿润年份,增温显著提高土壤和植物体的磷含量,降低了磷元素吸收效率;而在干旱年则没有影响,这可能是由于降雨解除了增温引起的干旱的负面效应㊂总的来说,温度升高会通过降低土壤磷含量和增加植物养分吸收效率,进一步加剧生态系统磷限制,而降水可能在一定程度上能够缓解因温度升高而导致的磷限制㊂2.4降水变化对草地土壤磷循环的调节作用全球气候变化导致极端降水和干旱事件频发,对生态系统过程产生重要影响㊂降水变化与土壤养分状态及植物光合生理过程密切相关[54],是控制草地初级生产力的关键因素㊂降水过多则会引起土壤磷元素的淋失作用,进一步影响土壤磷含量㊂降水通过改变凋落物产量及其养分归还影响土壤磷元素输入㊂降水变化主要通过提高土壤水分可利用性增加植物生物量㊂有研究表明地上生物量随着降水量的增加而增加,且在降水量增加60%时达到最大[55]㊂相邻年份降水量增加也会提高植物生物量㊂降水是干旱和半干旱区凋落物分解及养分释放的重要驱动因素㊂短期降水增多可加快表层凋落物的碎裂和水溶性物质的淋溶,加速凋落物质量损失和分解过程[56]㊂不同物种凋落物分解速率和养分释放对降水变化的响应存在差异㊂W a n g等[57]在半干旱草地研究了两种禾草和一种杂类草的凋落物分解及养分释放对水分添加的响应发现,杂类草的凋落物质量损失和养分释放快于禾草,且水分添加加速了杂类草凋落物中磷元素的释放㊂由于半干旱草原不同物种凋落物养分释放的不同步性,长期增加降水可能导致土壤养分供应和有效性的变化㊂降水主要通过控制有机磷矿化的生物化学过程影响磷元素的迁移和转化,以及土壤磷元素的组成和有效性㊂W a n g等[10]在干旱草地研究水分添加对土壤团聚体磷有效性的影响发现,水分添加能够改善土壤结构,加速土壤团聚体中磷元素的溶解与解吸过程,增加土壤有效磷含量㊂降水变化主要通过增加土壤有机碳含量,使微生物生物量上升㊂微生物生物量磷作为土壤磷库的重要组成部分,可以为植物提供大部分磷元素营养的需求㊂有研究表明降水能够提高微生物生物量磷的周转速率,从而提高土壤全磷含量[58]㊂D i j k s t r a等[59]用32P同位素标记法研究干旱在短期内对微生物磷吸收的影响,结果表明干旱抑制微生物磷吸收,这可能是由于水分减少降低了磷迁移率,进而限制了无机磷向微生物的供应㊂在降水对磷酸酶活性影响的研究中,一部分结果表明降水增多提高了磷酸酶活性,另一部分研究发现降水增多降低了磷酸酶活性,除了降水对微生物活性的影响外,造成结果差异的原因可能与植物生长发育的时间㊁土壤类型以及不同取样测定的时间有关㊂降水对磷元素输出的影响主要表现在植物吸收和土壤磷元素的淋失作用㊂由于研究地点㊁植物种类以及土壤含水量对土壤养分可利用性和植物营养元素影响等差异,降水变化对植物NʒP的影响尚不清楚㊂例如在地中海地区[60],适度干旱会增加土壤有效磷含量,却降低了植物体内的磷浓度㊂而在荒漠草原降雨量减少则会提高植物体内全磷含量[61]㊂目前关于降水变化对土壤-植物之间磷元素传递机制的研究较为薄弱,仍需进一步研究㊂在较高水分条件下,土壤磷扩散率会随着土壤湿度的增加而增加,加快植物和微生物吸收磷的速率[62]㊂降水事件频发会引起地表径流,加速土壤中的速效养分发生淋失作用㊂如S c h l e s i n g e r[63]认为在灌丛草地上有机磷化合501吴金凤刘鞠善李梓萌等草地土壤磷循环及其对全球变化的响应物占可溶性磷流失量的70%以上,大多数无机磷与碳酸钙矿物存在于不可利用的络合物中㊂2.5多因子交互作用对草地土壤磷循环的作用规律全球变化是由大气氮沉降㊁C O2浓度升高㊁温度升高㊁降水变化等因子共同作用的结果,且各个变化因子间存在复杂的交互作用㊂研究表明增温和施氮的综合效应显著提高土壤NʒP,提高了土壤全磷与微生物生物量磷的相关性,二者通过影响土壤和微生物CʒNʒP进而改变碳㊁氮㊁磷的循环[64]㊂D i j k s t r a等[65]在半干旱草原进行的增温和增加C O2浓度的实验表明,增温和C O2浓度升高可以通过改变土壤湿度来影响植物和微生物有效NʒP,温度升高可以减缓在高C O2浓度下植物的氮限制,并进一步加剧磷限制㊂降水变化可以解除温度升高引起土壤水分减少所导致的负面效应,在降水多的年份可在一定程度上缓解温度升高对土壤有效磷的不利影响㊂而施氮和降水的累加效应可提高土壤有效磷含量[9]㊂T h a k u r等[66]研究发现,C O2浓度㊁温度㊁降水三者交互作用影响土壤磷酸酶活性,当温度升高㊁降水增加时,磷酸酶活性会随着C O2浓度升高而降低㊂总之,全球变化因子及其复杂的交互作用会加剧草地磷限制,并通过调节地上植物群落㊁土壤环境和微生物活性,改变土壤有效磷含量,进而加快土壤磷循环(图1)㊂因此,研究磷元素的动态变化对预测草地生态系统对全球气候变化的响应至关重要㊂图中土壤磷循环部分参考赵琼等[12]和Bün e m a n n[14]的文献T h e p a r t o f s o i l p h o s p h o r u s c y c l i n g i n t h e f i g u r e r e f e r s t o t h e r e s e a r c ho f Z h a oQ i o n g e t a l.[12]a n dBün e m a n n[14]图1全球变化对草地土壤磷循环的作用机制F i g.1 T h em e c h a n i s mo f g l o b a l c h a n g e o n g r a s s l a n d s o i l p h o s p h o r u s c y c l i n g3全球变化影响土壤磷循环的机理3.1全球变化通过改变土壤理化性质影响磷循环土壤p H是影响磷循环的主要环境因子㊂土壤p H能够影响土壤磷元素的化学形态,但对土壤磷元素的作用主要与土壤中存在的离子和矿物含量有关[67]㊂在低p H值的土壤中,磷酸盐容易吸附在F e㊁A l㊁M n氧化物表面形成沉淀㊂在高p H值的碱性土壤或者石灰性土壤中,磷离子易与C a2+形成沉淀[12]㊂T y l e r[68]研究表明,在高p H值土壤中无机磷组分以钙磷酸盐为主,而在中p H值土壤中无机磷组分减少,在低p H值土壤中无机磷组分变化较大㊂这项研究还表明,磷酸铁和磷酸铝随着p H值的降低而增加㊂T u r n e r等[69]在研究土壤p H值与有机磷之间的关系时发现,在酸性土壤中,无机磷与游离的铝离子和锰离子呈显著负相关㊂全球变化对土壤p H也会产生一定的影响,长期大气氮沉降会引起土壤酸化,加速土壤阳离子(F e3+㊁A l3+㊁M n2+)从矿物中释放到土壤溶液中,与土壤中的活性磷酸盐结合或者形成沉淀,进而影响土壤磷元素601中国草地学报2021年第43卷第6期。
氮循环过程对土壤健康和农业可持续发展影响探讨

氮循环过程对土壤健康和农业可持续发展影响探讨引言:氮素是植物生长中的关键营养元素之一,对于维持土壤肥力和农作物的生长发育至关重要。
然而,过度或不当的氮素施用会导致氮素积累、土壤酸化和环境污染等问题。
因此,了解氮循环过程对土壤健康和农业可持续发展的影响是至关重要的。
一、氮循环过程概述氮循环过程是指氮在土壤和生物体之间的转化和迁移过程,包括氮素的吸收、固定、转化和释放等环节。
主要的氮循环途径包括农艺循环(作物吸收、农业副产品回收利用等)、自然循环(大气氮通过闪电、与植物共生固氮菌固定、酸雨氮素沉降等)和人为循环(化肥施用、有机肥施用等)。
二、氮循环对土壤健康的影响1. 土壤肥力:氮素是植物生长的重要营养元素之一,适量的氮素供应可以促进植物体的生长发育,提高农作物产量和品质。
合理施肥和农艺管理可以提高土壤氮素利用率,减少氮素的损失,维持土壤肥力水平。
2. 土壤酸化:氮素转化过程中,氨化作用和硝化作用会释放出氢离子,导致土壤酸化。
长期过量施氮会显著增加土壤酸度,降低土壤pH值,影响土壤微生物活性和养分循环,对土壤健康产生不利影响。
3. 土壤生物多样性:氮素的施用和氮素循环过程会影响土壤微生物群落结构和功能,进而影响土壤生物多样性的维持。
适度氮素供应可以促进土壤微生物多样性,增强土壤生态系统功能。
三、氮循环对农业可持续发展的影响1. 环境保护:过量的氮素施用和不当的农业废弃物管理会造成氮素积累和流失,导致土壤和水体的污染,甚至引发蓝藻暴发等环境问题。
因此,合理使用肥料、优化氮素施用方式等措施对于减少氮素污染具有重要意义。
2. 农业可持续性:氮素作为农业生产中的关键要素,适度的氮素施用可以提高农作物产量和品质,增加农民收益。
同时,合理的氮素管理可以减少氮素的损失和浪费,降低生产成本,提高农业的可持续性。
3. 土壤保护:氮素的循环过程不仅影响土壤肥力,还与土壤有机质的储存和分解密切相关。
合理管理土壤的氮素循环可以促进土壤有机质的稳定性,改善土壤结构和保护土壤健康。
气候变化对土地生态系统碳循环的影响

气候变化对土地生态系统碳循环的影响气候变化是当前全球面临的一大挑战,它不仅对人类社会造成了直接的影响,也对地球上的土地生态系统带来了巨大的变化。
其中,气候变化对土地生态系统碳循环的影响尤为重要。
本文将深入探讨气候变化对土地生态系统碳循环的影响,并分析其可能带来的后果及采取的应对措施。
一、气候变化对土地生态系统碳固定的影响气候变化对土地生态系统碳循环的重要影响之一是对碳固定的影响。
土地上的植物通过光合作用将二氧化碳转化为有机物质,并通过根系将部分有机物质转移到土壤中,从而完成碳的固定过程。
然而,随着气候变化的加剧,降水量和温度等环境因素发生了明显的变化,导致植物的生长状况和分布范围发生了改变。
一方面,气候变暖导致冻土融化,使得之前无法生长的植物得以在冻土层下的土壤中扎根生长,增加了碳的固定量;另一方面,干旱和热浪等气候极端事件频发,使得一些植物无法正常生长,减少了土地生态系统的碳固定能力。
因此,气候变化对土地生态系统碳固定能力的影响是双重的,不同地区的影响程度也存在差异。
二、气候变化对土地生态系统碳释放的影响除了对碳固定能力的影响,气候变化还会对土地生态系统碳循环的另一个重要过程——碳释放造成影响。
土壤中的有机质在一定条件下会分解并释放出二氧化碳和甲烷等温室气体,这就是所谓的土壤呼吸过程。
气候变化会通过影响土壤湿度、氧气含量等条件,进而改变土壤中的呼吸速率。
例如,气候变暖导致土壤湿度下降、干旱加剧,会抑制土壤中的呼吸作用,从而减少碳的释放量;而强降雨会导致土壤湿度增加,加快土壤中的呼吸速率,增加碳的释放量。
因此,气候变化对土地生态系统碳释放量的影响也存在一定的复杂性和多样性。
三、气候变化对土地生态系统碳储存的影响碳固定和碳释放是土地生态系统中的两个关键过程,它们共同决定了土地生态系统中碳的储存量。
气候变化的影响使得土地生态系统中的碳储存量发生了变化。
虽然碳固定和碳释放受到气候变化的相互作用,并不是简单的线性关系,但总体而言,随着全球气候变暖和极端气候事件频发,土地生态系统中的碳储存量整体上呈现出减少的趋势。
土壤碳循环过程及其气候调节效应

土壤碳循环过程及其气候调节效应土壤是地球重要的碳库之一,土壤中的碳含量远远大于大气中的碳含量。
土壤中的碳循环过程在全球碳循环中起着至关重要的作用,并且对地球气候具有调节作用。
本文将对土壤碳循环过程及其气候调节效应进行详细的介绍和分析。
首先,土壤碳循环过程主要包括碳输入、碳固定和碳释放三个环节。
碳输入是指有机质的输入、植物残渣的输入以及以其它方式输入土壤中的碳。
有机质的输入是通过植物残渣、动物排泄物、植物死亡等形式将有机化合物输入土壤。
此外,大气中的二氧化碳也可以通过植物的光合作用输入土壤。
碳固定是指土壤中有机碳的积累过程,主要通过土壤中的微生物的代谢作用来实现。
这些微生物可以利用有机质进行呼吸作用和分解作用,将其转化为溶解有机碳和胶体有机碳,并稳定在土壤中。
碳释放是指土壤中的有机碳被微生物呼吸消耗,同时由于土壤通气性差,土壤中的有机质会发生缺氧分解,产生二氧化碳和甲烷等气体释放到大气中。
土壤碳循环过程对气候具有重要的调节作用。
首先,土壤中的碳含量可以影响大气中的二氧化碳浓度。
随着全球温室气体的排放增加,土壤中的有机质被破坏和挥发进而释放出大量的二氧化碳,增加了大气中的二氧化碳浓度。
其次,土壤中的有机质含量可以影响土壤的水分保持能力。
有机质具有一定的吸湿性,可以增加土壤的保水能力,从而减少水分的蒸发和流失,对维持地表水分平衡起到了重要的作用。
此外,土壤中的有机质可以增加土壤的肥力,改善植物生长环境,进而增加植物的光合作用和碳固定能力,有助于减少大气中的二氧化碳浓度。
此外,土壤碳循环过程还受到一系列因素的影响。
首先,气候因素对土壤碳循环具有重要影响。
气温和降水是影响土壤中有机质分解速率的重要因素。
较高的温度和湿度有利于土壤中有机质的分解和呼吸过程,导致有机碳释放增加。
其次,土壤类型也对碳循环有影响。
不同类型的土壤对有机质的固定和稳定能力不同,影响着土壤中碳的积累。
再次,植被类型和覆盖度也对土壤碳循环有重要影响。
农田土壤有机碳,氮转化及其与温室气体排放的关系

农田土壤有机碳,氮转化及其与温室气体排放的关系农田土壤有机碳和氮的转化与农业生产和土地利用紧密相关,并对温室气体排放产生影响。
土壤有机碳是土壤中最重要的有机质组分之一,影响土壤生态系统的功能和气候变化。
农业活动会影响农田土壤有机碳的含量和分布,通过改变土地利用方式、施肥和耕作方式等来影响农田土壤有机碳的积累。
随着土壤有机碳的下降,土壤对二氧化碳的吸收也逐渐减少,从而导致温室气体排放增加。
因此,通过科学管理土地资源和耕作方式,适当增加土壤有机质含量,可以降低农田温室气体排放。
土壤中的氮元素是植物生长所必需的营养元素,但过量的氮元素会引起农田土壤氮素累积,导致氮素的硝化和反硝化反应频繁发生。
反硝化反应会通过产生亚氧环境导致氧化亚氮的生成和释放,从而增加温室气体氮氧化物的排放。
因此,科学合理的氮肥管理,减少氮素的过度利用,有助于减少农田土壤氮素累积和温室气体排放。
总而言之,农田土壤有机碳、氮转化与温室气体排放之间存在着密不可分的关系,科学的土地资源管理和农业生产方式,都将在减少温室气体排放中发挥着重要作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
模拟增温对小麦田土壤碳氮循环过程的影响一、研究背景1.气候变化由于化石燃料的燃烧和土地利用方式的变化,大气中温室气体浓度快速上升,加快了气候变暖趋势,由温室效应导致的全球变暖已经成为最热门的环境问题之一。
在全球温度变暖的气候条件下,我国在最近100年来,地表平均温度表现出明显上升趋势,其升温幅度大约在0.5—0.8℃之间,这比同时期的全球温度变化上升幅度的平均值(0.6±0.2℃)略高[1]。
我国这近100年的气温上升在冬季和春季最为明显,而夏季的气温变化并不明显,这与全球以及北半球的平均气温变化情况相一致[2]。
根据中国科学家利用气候模式预估的结果表明,未来50—100年全球地表温度将继续上升。
在21世纪,全球和东亚地区表现出明显的气温上升,在中高纬度地区的气湿上升又大于中低绅度地区,尤其在冬季和夏季气温上升现象更为明显。
未来气温变暖的幅度将随着温室气体排放的情景和模式产生一定的差异。
根据以上的预测结果,全球气温将出现在过去一万内从未有过的变化,这对全球环境变化以及生态系统造成巨大的冲击作用,从而将导致一系列的环境问题,比如海平面上升、生态系统生物种群的变化、降水的时空分布和格局变化等,直接或间接的对人类赖以生存的陆地生态系统产生重大影响(IPCC,2001)。
20世纪50年代以来,观测到的气候系统的许多变化是过去几十年甚至千年以来史无前例的,包括大气和海洋的温度升高、冰雪覆盖面积减少。
(1)大气观测事实过去30年,每10年地表温度的增暖幅度高于1850年以来的任何时期。
在北半球,1983-2012年可能是最近1400年来气温最高的30年,21世纪的第一个10年是最暖的10年。
全球地表持续升温,结合陆地与海洋的地表温度数据,通过线性趋势计算1880-2012年全球平均温度已升温0.85℃。
基于最长的数据库资料,1885-1900年平均和2003-2013年平均相差0.78℃。
20世纪末的升温是横跨全球的,虽然在大陆尺度恢复的中世纪暖期(AD 950-1250)在一些地区的气温与20世纪末一样高,但仅限于区域性升温,并未像20世纪末一样是跨地域全球性的升温自20世纪中叶以来,可以肯定的是全球对流层已经升温。
更完整的观测表明,对流层温度变化在北半球温带的估算比其他地方的信度更高。
自1901年以来,北半球中纬度陆地的降水量已经在增加。
(2)海洋观测事实海洋变暖导致气候系统中储存的能量增加,占1971-2010年储存能量的90%以上。
海洋上层(0-700m)在1971-2010年几乎肯定变暖,而在19世纪70年代至1970年则有可能变暖。
从全球尺度来讲,海洋升温最大的是在近表层,1971-2010年期间海面至水深75米之间的表层海水以每10年升温0.11℃的速度在升温。
在1957-2009年期间,700-2000米海深的海水可能已经变暖。
但1992-2005期间,在2000-3000m海深间没有观测到明显的升温趋势,而在水深3000m以下直到海床的海温这段期间却在升温,深层海水温度上升最多的地区是在南极海;依据1971-2011年40年相对完善的采样,超过60%气候系统所增加的能量被储存在0-700m的表层海水中,30%储存于700m以下的海洋,这说明90%新增的气候系统能量储存在变热的海洋。
从线性趋势估计,1971-2010年表层海水的热含量可能增加了17×1022J。
自1950年代以来,海洋上蒸发占优势的高盐度区盐度更加高了,而降水占优势的低盐度区海水变的更淡了,这提供的间接证据说明海洋上蒸发和降水已经发生了变化。
(3)冰冻圈观测事实过去20年,格陵兰岛和南极冰盖已大量消失,世界范围内的冰川继续萎缩,而北极海冰和北半球春季积雪已呈持续减少的程度。
全球除冰盖周边冰川之外的冰川冰量损失,在1971-2009年间平均速率很可能为每年损失226×109t·a-1,但在1993-2009年期间平均速率很可能为每年达到275×109t·a-1,显示出全球冰川退缩速度在增加;格陵兰冰盖的冰量损失平均速率已经从1992-2001年每年34×109t·a-1,持续增加到2002-2011年的每年215×109t·a-1,而在南极冰盖的冰量损失平均速率从1992-2001年的每年30×109t·a-1,增加到2002-2011年的每年147×109t·a-1。
在1979-2012年间,年均北极海冰范围很可能每10年以3.5%~4.1%的速度在减少(相当于每10年减少45×104~51×104km2海冰面积)。
到了夏季,海冰面积最小,但这时段的多年冰的面积每10年也以9.4%~13.6%的速度在减少(相当于每10年减少73×104~107×104km2海冰面积)。
过去30年来北极夏季海冰退缩与海水温度上升,至少在1450年以来均属异常的。
而南极海冰面积反而在增加,在1979-2012年期间,南极海冰范围很可能每10年平均以1.2%~1.8%的速度增加(相当于每10年增加13×104~20×104km2海冰面积)。
自1980年代初期以来,大多区域的多年冻土层温度在增加。
已观测到阿拉斯加北部在1980年代初期至2000年代中期增温达3℃,俄罗斯欧洲北部在1971-2010年增温达2℃。
其中,在俄罗斯欧洲北部多年冻土的厚度及面积在1975-2005年间有明显减小[3]。
2.温室气体变化就目前的研究而言,导致全球温度升高的气体总结为下几种主要气体:二氧化碳(CO2)、甲烷(CH4)和氧化亚氮(N2O)。
这些气体被称为溫室气体,它们能够吸收太阳的长波热辐射,从而导致地球表面的温度上升,造成气候变暖现象[4]。
这些气体具有可使大气温度升高的温室效应,称为温室气体(green-house gas)。
温室气体浓度的升高,将使全球气候发生显著的变化。
这种变化将对生态系统的结构、功能及分布产生重要影响,同时会扩大和加深物理反馈过程,比如地球物理学过程(水分蒸发、反射率等)、热力学过程(大气循环、洋流和气体的溶解性等)、生物地球化学过程(甲烷的氢代谢、陆地和海洋有机物的生产和分解等)。
另外,还会对人类活动产生作用。
据记载,在过去的一个世纪中陆地和海洋温度增加了0.3~0.6℃,其中最近10a增加速度最快。
在温室气体中,CO2对温室热效应作用最大占56%,尽管其它气体对热辐射吸收能力大大高于CO2,但由于其浓度基数低,作用较小(表1),其中CH4占11%,N2O占6%,CFCS占24%,水汽占3%[5].3.气候变化对碳氮循环的影响碳、氮作为生物体必需的营养元素,同时也是研究生态系统中物质循环的重要元素。
在IPCC最新研究报告中显示,大气中二氧化碳浓度从1750年至今增加了31%,经计算预测在2100年大气中二氧化碳浓度可达到540-970μmol·mol-1[6]。
土壤是陆地生态系统的主要碳库,而土壤碳库的变化对碳氮循环有重要影响。
碳氮循环是农田生态系统最基本的生态过程,对农田生态系统的稳定性、生产力及其环境效应具有关键性的影响作用[7]。
大气中CO2浓度的升高既能导致温室效应增强进而影响全球气候变化,又能通过施肥效应影响绿色植物的生长,进一步影响整个生态系统的物质循环过程。
土壤碳氮循环研究中表明,土壤呼吸释放出来的CO2对大气二氧化碳浓度上升产生重要影响,同时作为陆地生态系统碳收支的重要环节,研究土壤呼吸强度以及主要影响因子成为探究土壤碳循环问题的重中之重;N2O是重要的温室气体,一方面,作为一种效应仅次于CO2和CH4的含氮温室气体,N2O对大气的福射强迫作用约占总长存留温室气体的6%,另一方面,在平流层中能光解NO进而破坏臭氧层。
大气中N2O的浓度也以每年0.2%—0.3的速度在増加[8]。
研究表明,作为温室气体的N2O主要来源于土壤的反硝化作用过程,而反硝化过程中的氮一部分来源于土壤硝化作用产生的氮,所以N2O的产生和排放与土壤氮循环过程密切相关。
因此,土壤氮循环的研究在当今气候变暖条件下受到各国学者的重视,研究土壤硝化、反硝化作用是当前氮循环研究中的重要议题之一。
对我国这样的农业大国而言,农田是极为重要的温室气体排放的来源,在农业发展中,农田土壤中的有机质经过微生物的分解,都以二氧化碳的形式释放到大气环境中去,同时在农田中经过长时间淹水发酵易产生甲烷,农田土地利用面积广泛,全球大气中一半上的氧化亚氮都来自土壤的硝化及反硝化过程[9]。
目前关于水稻田土壤碳氮循环的研究已经有很多了,但是关于小麦田的论文相对较少,而小麦在我国分布很广,南起海南的热带地区,北至黑龙江漠河的严寒地带,西自新疆的西界,东到台湾及沿海诸岛屿,均有小麦栽培。
但主要分布在20-41"N。
占全国麦播总面积80%、总产量90%的产区是河南、山东、河北、黑龙江、安徽、甘市、江苏、陕西、四川山西湖此内家方等13个名、市、自治区,而尤以河南和山东为最自1月到10用全国不同地区月月都有小麦收获。
生存期短者70天,长的可达300多天,西藏有些可达周年之久,是我国最重要的粮食作物之一,其产量占我国粮食总产量的22.5%。
冬小麦是我国小麦的主体,占全国小麦播种面积的80%以上。
因此,在全球气候变化的前提下,本研究通过观测小麦田模拟增温观测土壤呼吸速率、硝化速率、反硝化速率及相关土壤碳氮指标,分析了模拟增温对小麦田土壤呼吸、土壤碳氮循环关键过程的影响规律,以期为了解全球变暖情境下,增温对土壤碳氮循环的潜在影响提供基础资料和科学依据。
土壤CO2、N2O排放与土壤碳氮循环过程有关,全球变暖对生态系统具有重要影响,通过增温试验来研究和探讨我国小麦田温室气体的排放量及减排措施,这对于未来气候变暖情形下生态系统物质循环和能量流动的规律具有重要意义。
二、研究现状及进展气候变暖作为全球变化的主要表现之一(IPCC,1991,1997,2001),己经成为一个不争的事实。
温度对陆地生态系统碳循环过程有着重要的影响,比如植物的光合作用、土壤的呼吸作用等,这都能引起土壤、生物以及大气中的碳收支的重要变化[3],从而对大气中二氧化碳浓度产生一定影响。
总之,气候变暖对陆地生态系统的影响是目前全球气候变化问题研究中的核心之一,气候变暖对陆地生态系统碳循环过程的影响及其反馈于气候变暖的作用是全球气候变化问题研究的重中之重(IPCC,2001)。
由于温度是控制和调节许多生态学过程的关键因素,气候变暖势必会对生态系统生产力、植物群落结构以及土壤生化过程都产生非常深刻的影响。