5第五章 血红素蛋白
血红蛋白的生成、调节及相关的疾病

血红蛋白的生成、调节及相关的疾病血红蛋白合成血红蛋白结构:珠蛋白+血红素(Fe2+ +卟啉)血红素属于铁卟啉化合物,由Fe2+与卟啉环螯合而成。
根据同位素示踪法,其合成的基本原料是琥珀酰辅酶A、甘氨酸和Fe2+ 。
其合成过程包括:1.δ-氨基-γ-酮戊酸(ALA)的生成。
在线粒体内,琥珀酰CoA与甘氨酸在ALA合酶作用下合成ALA。
2.胆色素原的生成ALA进入胞质,在ALA脱水酶作用下,2分子ALA脱水合成胆色素原3.尿卟啉原与粪卟啉原合成胞质中胆色素原脱氨酶催化四分子胆色素原合成为一分子现状吡咯再由尿卟啉原Ⅲ同合酶转化为尿卟啉原Ⅲ,之后进一步脱羧变为粪卟啉原Ⅲ。
4.血红素合成粪卟啉原Ⅲ进入线粒体脱羧生成原卟啉原Ⅸ,经氧化得到卟啉原Ⅸ,此为血红素的直接前体。
之后Fe2+ 螯合形成血红素此为整体反映。
血红蛋白的另一部分珠蛋白按照中心法则,由基因转录出RNA再翻译成为蛋白质。
珠蛋白与血红素结合成为血红蛋白,而单个珠蛋白是利用它们的互补面和在红细胞中的高浓度自动结合到血红蛋白四聚体中的。
血红素对于血红蛋白的合成具有重要作用,它不仅为血红蛋白合成的底物,还可促进珠蛋白mRNA的合成聚集,对血红蛋白的合成起到调节作用。
血红蛋白合成调节:1.血红素的反馈调节当血红素的合成速度远大于珠蛋白的合成速度时,过多游离的血红素可以对ALA合酶具有别构抑制作用。
不仅如此,过多的血红素被氧化为高铁血红素对ALA酶也有很强的抑制作用。
2.促红细胞生成素促进血红蛋白合成EPO主要与晚期红系祖细胞上的EPO受体结合。
当机体缺氧或者红细胞减少时,肾红细胞生成素激活表达,释放入血到达骨髓,诱导ALA合酶合成,促进血红蛋白合成。
另外血红素可促进骨髓原始红细胞的增值与分化,加速有核红细胞成熟。
此外促红细胞生成素还可以减少红细胞的凋亡。
3.体内的类固醇激素(睾酮等)水平升高,诱导ALA酶合成,促进血红蛋白合成。
与血红蛋白相关的疾病血红蛋白病:由于血红蛋白分子结构异常或珠蛋白肽链合成速率异常所引起的一组遗传病。
生物化学 第5章 蛋白质结构与功能

第五章蛋白质结构和功能的关系一、、肌红蛋白的结构与功能:1、肌红蛋白的三级结构哺乳动物肌肉中储氧的蛋白质。
由一条多肽链(珠蛋白,153个aa残基)和一个血红素辅基组成。
亚铁离子形成六个配位健,四个与N原子,一个与组氨酸,一个与氧配位。
球状分子,单结构域。
8段直的α-螺旋组成,分别命名为A、B、C…H,拐弯处是由1~8个氨基酸组成的松散肽段(无规卷曲)。
4个Pro残基各自处在一个拐弯处,另外4个是Ser、Thr、Asn、Ile。
血红素辅基血红素辅基,扁平状,结合在肌红蛋白表面的一个洞穴内。
CO 中毒CO 与肌红蛋白有更高的亲和性2、肌红蛋白的氧合曲线OMb 解离平衡常数:][]][[22MbO K =][2PO Mb K ∙=][2MbO 氧饱和度:[]2MbO Y =][][2Mb MbO +PO 2Y =2PO K +Y=0.5时,肌红蛋白的一半被饱和,PO 2=K =P 50=2.8t torr(托)解离常数K 也称为P 50,即肌红蛋白一半被饱和时的氧压。
3、Hill 曲线和Hill 系数YY K PO YK PO Y log log 1log 122-=-=-Hill曲线Log[Y/(1-Y)]=0时的斜率称Hill 系数(n H )肌红蛋白的n H =1二血红蛋白的结构与功能蛋白的结构与功能1、血红蛋白的结构:成人成人:HbA:α2β298%,a亚基(141),β亚基(146)HbA2:α2δ22%胎儿:HbFα2γ2早期胚胎:α2ε2▲接近于球体,4个亚基分别在四面体的四个角上,每个亚基上有一个血红素辅基。
▲α、β链的三级结构与肌红蛋白的很相似,一级结构具有同源性。
氧合造成盐桥断裂42、血红蛋白的氧合曲线四个亚基之间具有正协同效应因此它的氧合曲四个亚基之间具有正协同效应,因此,它的氧合曲线是S 型曲线。
Hill 曲线和Hill 系数。
协同效应可增加血红蛋白在肌肉中的卸氧量,使它能有效地输送氧气。
血红蛋白异常所致的贫血及其实验诊断考点总结
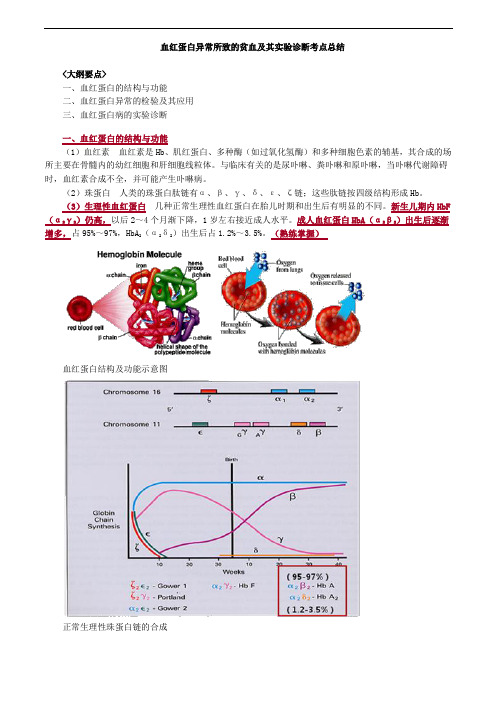
血红蛋白异常所致的贫血及其实验诊断考点总结<大纲要点>一、血红蛋白的结构与功能二、血红蛋白异常的检验及其应用三、血红蛋白病的实验诊断一、血红蛋白的结构与功能(1)血红素血红素是Hb、肌红蛋白、多种酶(如过氧化氢酶)和多种细胞色素的辅基,其合成的场所主要在骨髓内的幼红细胞和肝细胞线粒体。
与临床有关的是尿卟啉、粪卟啉和原卟啉,当卟啉代谢障碍时,血红素合成不全,并可能产生卟啉病。
(2)珠蛋白人类的珠蛋白肽链有α、β、γ、δ、ε、ζ链;这些肽链按四级结构形成Hb。
(3)生理性血红蛋白几种正常生理性血红蛋白在胎儿时期和出生后有明显的不同。
新生儿期内HbF (α2γ2)仍高,以后2~4个月渐下降,1岁左右接近成人水平。
成人血红蛋白HbA(α2β2)出生后逐渐增多,占95%~97%,HbA2(α2δ2)出生后占1.2%~3.5%。
(熟练掌握)血红蛋白结构及功能示意图正常生理性珠蛋白链的合成二、血红蛋白异常的检验及其应用1.血红蛋白电泳(原理了解,其余掌握)2.抗碱血红蛋白测定(熟练掌握)3.异丙醇沉淀试验(掌握)4.红细胞包涵体试验(掌握)5.HbA2测定(熟练掌握)6.珠蛋白肽链分析(掌握)7.红细胞镰变试验(掌握)1.血红蛋白电泳原理:根据不同的血红蛋白带有不同的电荷,等电点不同,在一定的pH缓冲液中,缓冲液的pH大于Hb的等电点时其带负电荷,电泳时在电场中向阳极泳动,反之,Hb带正电荷向阴极泳动。
经一定电压和时间的电泳,不同的血红蛋白所带电荷不同、相对分子质量不同,其泳动方向和速度不同,可分离出各自的区带,同时对电泳出的各区带进行电泳扫描,可进行各种血红蛋白的定量分析。
参考值:pH8.6TEB缓冲液醋酸纤维膜电泳,正常血红蛋白电泳区带:HbA>95%、HbF<2%、HbA2为1.0%~3.1%。
pH8.6TEB缓冲液适合于检出HbA、HbA2、HbS、HbC,但HbF不易与HbA分开,HbH与HbBarts不能分开和显示,应再选择其他缓冲液进行电泳分离。
第五章 蛋白质的三维结构

肌红蛋白的三级结构
肌红蛋白由一条多肽链和一个血红素(heme)辅基 构成,分子量为16700,含153个氨基酸残基。
抹香鲸肌红蛋白的三级结构
维系三级结构的化学键
盐键 氢键 疏水作用 范德华力 二硫键
六、蛋白质的四级结构
二个或二个以上具有独 立的三级结构的多肽链 (亚基),彼此借次级键 相连,形成一定的空间 结构,称为四级结构。
β-转角
RNase的分子结构
β-折叠
α-螺旋
无规卷曲
各 种 二 级 结 构 在 拉 氏 图 中 的 分 布
三、纤维状蛋白质
含大量的α-螺旋,β-折叠片,整个分子呈纤维状。 广泛分布于脊椎和无脊椎动物体内,起支架和保护 作用。分不溶性和可溶性两类。 不溶性纤维状蛋白质主要有: 1、 α-角蛋白 2、 β-角蛋白 3、 胶原蛋白 4、 弹性蛋白
第五章 蛋白质的三维结构
一、多肽主链折叠的空间限制
(一)肽平面与Cα原子的二面角: 相邻肽平面构成二面角 :一个Cα原子相连的两 个肽平面,由于N1—Cα和Cα—C2(羧基碳)两个键 为单键,肽平面可以分别围绕这两个键旋转,从而 构成不同的构象。一个肽平面围绕N1—Cα(氮原子 与α- 碳原子)旋转的角度,用Φ表示。另一个肽平 面围绕Cα—C2(α- 碳原子与羧基碳)旋转的角 度,用Ψ表示。这两个旋转角度叫二面角。
★ 从 Cα 沿 键 轴 方 向 观察,顺时针旋转的 Φ (Ψ)角为正值,反 之为负值。
Cα-N和Cα-C键旋转时将受到α-碳原子上的侧 链R基的空间阻碍影响,所以使肽链的构象受到限 制,只能形成一定的构象。如果每一个氨基酸残基 的Ψ和Φ角已知,多肽主链的构象就被完全确定。 (拉氏构象图)
拉氏构象图:可允许的Φ和Ψ值
上海师范大学本科课程教学大纲-化学教学中心.doc

上海师范大学本科课程教学大纲课程名称:化学与生命(Element and Life)学分:2 总学时:36开课专业:化学(师范)和应用化学一、课程性质、目的和培养目标《化学与生命》属于新兴边缘科学-生物无机化学的范畴。
它是无机化学与生物化学、材料科学、医学临床化学、营养化学、环境化学等学科相互渗透、相互融合的产物,应用其原理和规律为人类服务。
本课程为化学类专业的选修课,36学时,2学分。
该课程讨论与生命有关的金属酶和金属蛋白以外,还对金属离子在大脑中和神经系统中的功能研究加以注意。
希望通过本科的学习,使学生对金属离子对生命的重要性有个充分的了解并能解释其生活中的一些现象。
二、预修课程无机化学三、课程内容和建议学时分配第一章化学与生命概论第一节生命必需元素及其动态平衡第二节金属中心的结构与功能第二章生物大分子的结构与性质第一节蛋白质及其组成与结构第二节核酸第三节其他生物分子第三章生物无机化学中的物理化学方法第一节X射线单晶衍射法第二节X射线吸收光谱第三节核磁共振谱第四节电子顺磁共振光谱第五节其他物理化学方法简介第四章天然含铁氧载体及其模拟物第一节血红蛋白和肌红蛋白第二节蚯蚓血红蛋白第三节重要模型化合物一一合成氧载体第五章血红素蛋白及催化体系第一节细胞色素:电子传递第二节细胞色素P-450第三节甲烷单加氧酶、双加氧酶第四节过氧化物酶及过氧化氢酶第六章铁一硫蛋白和其他非血红素铁蛋白第一节铁一硫蛋白第二节铁一硫蛋白模拟物第三节含铁核昔酸还原酶及其他第七章铁的吸收、转运及储备第一节铁载体第二节运铁蛋白第三节运铁蛋白受体第四节储铁蛋白第五节铁的生物学调控第八章含锌酶和含锌蛋白第一节概述第二节锌肽酶第三节碳酸酎酶第四节核酸酶第五节锌指蛋白第六节金属硫蛋白第九章含铜蛋白及含铜酶第一节含铜电子传递蛋白一一型铜蛋白第二节铜锌超氧化物歧化酶一一型铜蛋白第三节含铜氧载体一一型铜蛋白第四节多铜蛋白第五节重要模型化合物第十章钳酶及含钳蛋白第一节固氮酶第二节双氮的配为化学第三节固氮酶模拟第四节钳转氧酶及其模拟第^一章镣酶第一节尿素及其模拟第二节氢化酶及其模拟第三节一氧化碳脱氢酶和乙酰辅酶A合成酶第四节超氧化歧化酶第十二章辅酶岛2及其相关化学第一节钻一碳键的形成与断裂第二节氧化-还原反应第三节重排反应第四节甲基转移反应第十三章神经生物无机化学第一节钠、钾、钙的生物无机化学和调控第二节离子通道第三节离子泵第四节神经系统中的过渡金属离子第十四章医药中的金属:药物和疾病第一节概述第二节治疗药物第三节诊断药物第四节金属离子与神经退行性疾病第十五章生物体系的矿化与自组装第一节生物矿化概论第二节主要的生物矿源第三节生物矿化机理第四节生物矿化的体外模拟第五节展望课时安排章―二三四五六七八讲授时数22422224章九十十一十二十三十四十五总计讲授时数422222236四、教材和参考书目1 .使用教材:郭子建,孙为银主编,生物无机化学科学教育出版社,2006 年第一版。
第5章血液y

血浆的化学成分
1、化学成分:由90%的水和100多种溶 质(蛋白质、脂类、糖类、氨基酸、维 生素、矿物质、气体、激素、各种细胞 代谢产物和电解质)组成。
2、血浆的酸碱平衡:正常人血浆的pH 为7.35—7.45 ,平均为7.45。
血浆是血液细胞间质,为浅黄色,有黏性的透明液体。
包括水(91%)、血浆蛋白、脂蛋白、酶、激素、维生 素、无机盐和各种代谢产物。 其功能为: ①运载血细胞、血小板、营养物质和机体代谢物; ②所含补体蛋白和免疫球蛋白参与机体的免疫机能; ③运输各种激素,参与机体体液调节; ④所含纤维蛋白原及凝血因子参与血凝;运输无机盐
这种内环境相对稳定的状态称为稳态。 生理意义:是机体维持正常生命活动的先决条件。
细胞内液﹙占总量的2/3﹚
体液
细胞外液﹙占总量的1/3﹚
血管内(1/5):血浆 细胞外液
血管外(4/5):组织液、淋巴液、脑脊液等
二、血液的基本组成与血量
(一)血液的组成
(二)、血量:
定义:机体中血液的总量称为血量,是血浆量和血细胞 的总和。
2. 功 能 : 特殊颗 粒含血小板因子 Ⅳ 、血小板源性生长因 子 (PDGF)等,致密颗粒含5-羟色胺、钙离子等,参与止血和凝血, 促进内皮细胞增殖、修复血管。
3.来源:是骨髓巨核细胞脱落的胞质小块。 4.寿命:7-14天。 5.正常值:15万-45万/微升血液
血涂片示聚集的血小板
四、免疫系统
血浆胶体渗透压: 主要来自白蛋白 约1.3 mmol/L
血液晶体渗透压的生理意义:
其相对稳定对维持细胞内外的水平衡及血细胞 的正常形态有重要意义;
红细胞
血管
血液胶体渗透压的生理意义:
其相对稳定对维持血管内外水平衡有重要意义。
血红素

含血红素蛋白的代谢在哺乳动物中需要:
①对卟啉环剪切产生的疏水性产物进行处理;
②所含铁的保留和动用,使其重新被利用。红细胞的生存周期大约为120天,衰老细胞通过膜的改变被识别, 并被血管外的状内皮系统吞噬。珠蛋白链变性后,将血红素释放于细胞质中;珠蛋白被降解为其组成的氨基酸, 重新被利用以满足一般代谢的需要。
(2)分离红细胞取抗凝猪血10mL于离心管中,以3000r/min离心15min,倾出上清液(血浆),收集红细胞, 用0.9% NaCl洗涤2次。洗涤方法为:用适量生理盐水悬浮红细胞,搅拌均匀,离心,收集红细胞沉淀,测定红细 胞体积(mL)。
(3)溶血加入相当于红细胞体积1倍的去离子水、0.25倍95%乙醇,搅拌30min,红细胞吸水胀裂,血红蛋白 释放出来。
除了运载氧,血红素还可以与二氧化碳、一氧化碳、氰离子结合,结合的方式也与氧完全一样,所不同的只 是结合的牢固程度,一氧化碳、氰离子一旦和血红素结合就很难离开,这就是煤气中毒和氰化物中毒的原理,遇 到这种情况可以使用其他与这些物质结合能力更强的物质来解毒,比如一氧化碳中毒可以用静脉注射亚甲基蓝的 方法来救治。
提取测定
试剂器材
提取原理
操作步骤
血红蛋白在pH低于3.0时,血红素与珠蛋白的结合最为疏松,此时加入有机溶剂丙酮,使珠蛋白变性凝固, 血红素则溶于丙酮中,在丙酮中加入适量的鞣酸或乙酸钠,可得到较纯的血红素结晶,然后用乙醇一乙醚洗涤, 可得到精制血红素。血红素在波长385处有最大吸收,可直接进行比色测定。
血红素分子结构由于协同效应,血红素与氧气的结合曲线呈S形,在特定范围内随着环境中氧含量的变化,血 红素与氧分子的结合率有一个剧烈变化的过程,生物体内组织中的氧浓度和肺组织中的氧浓度恰好位于这一突变 的两侧,因而在肺组织,血红素可以充分地与氧结合,在体内其他部分则可以充分地释放所携带的氧分子。可是 当环境中的氧气含量很高或者很低的时候,血红素的氧结合曲线非常平缓,氧气浓度巨大的波动也很难使血红素 与氧气的结合率发生显著变化,因此健康人即使呼吸纯氧,血液运载氧的能力也不会有显著的提高,从这个角度 讲,对健康人而言吸氧的所产生心理暗示要远远大于其生理作用。当血液内红细胞破坏过多,肝脏负荷增加肝细 胞内运送、结合和排泄障碍,或肝外胆道阻塞,都可引起血内胆红素浓度增高而出现黄疸。
第五六章铁-硫蛋白和其他非血红素铁蛋白

铁载体的结构类型
铁载体与铁形成非常稳定的低电荷配合物,有利于高电荷 铁(III)离子跨过亲脂性膜,实现铁(III)的吸收,运输过程。
氧肟酸盐型 (hydroxamate type); 邻苯二酚型(catechol type)
氧肟酸盐型 (hydroxamate type)
铁色素
Crystal structure of ferri普遍的看法是:在细胞 内,首先 Fe3+被还原为Fe2+,之后通过质子化作用将结合 不牢固的Fe2+从载体上置换下来。
第二节 铁蛋白(ferritin)
高等植物,微生物,哺乳动物的各种细胞中都含有储铁 蛋白。细胞类型不同储铁功能也有所不同。哺乳动物体内 主要储存铁的蛋白质,主要分布在动物的脾脏(spleen),肝 脏 (liver)和骨髓(bone marrow)中,植物的叶绿体和某些 细菌中也发现有铁蛋白。
铁传递蛋白的分类与结构
铁传递蛋白是一类金属键合蛋白,对于维持生命体
中铁的含量以及铁的新陈代谢起着重要作用。根据来源 不同可将其分为四类:血清运铁蛋白(serum transferrin), 伴清运铁蛋白 (ovotransferrin), 乳运铁蛋白 (lactoferrin), 黑素运铁蛋白(melanotransferrin)。
邻苯二酚型 (catechol type)
肠杆菌素, Enterobactin
Enterobactin + Fe
铁载体的性质
根据软硬酸碱规则,铁载体中属于硬碱的配位原子(氧) 可与属于硬酸的Fe3+配位,形成稳定的配合物。研究表明, 含有三个双齿配体的铁载体与 Fe3+ 的结合常数大于 1030 。 虽然铁载体也可与Ga3+及人造錒元素结合,但是与Al3+及所 有二价金属离子的结合能力是相当弱的,因此自然界出现 的金属离子中, 铁载体仅对Fe3+选择性性结合。 所结合的 Fe3+处于高自旋态 (HS), 动力学性质是活泼的,使铁载体 结合的Fe3+能向细胞内其它配体或蛋白质释放铁。
血红素与胆色素代谢

问题
§1 血红素的化学
CH2
血红素是一种铁卟啉化
CH
CH3
合物, Fe2+与氧结合,运输 至全身各组织细胞,与细胞
H3C
CH=CH2 氧的供给和利用密切相关
NN
Fe
NN
H3C
CH3
Fe
正常C情H况2 下,血CH红2 素与氧结合与否,都不会引起
结合胆红素20-25mg/dl (正常浓度 0.1-1.0mg/dl )
生理意义:
增加了胆红素在血浆中的溶解度,便于运输
限制胆红素自由透过各种生物膜,避免对组织 细胞产生毒性作用
胆红素的转运 胆红素 + 清蛋白
胆红素—清蛋白
1) 清蛋白的结合,暂时限制胆红素毒性并促进其运输, 2) 不能透过肾,在尿中不出现,只存在于血液中,称
结合胆红素 肠菌 葡萄糖醛酸基 游离胆红素
+8H
尿胆素原 +2H
-2H 尿胆素
中胆素原 +4H -2H
中胆素
粪胆素原 -2H
粪胆素
胆素(黄色)
结合胆红素 体循环
原型
重新吸收 10 ~ 20% 胆素排泄 80%~90%
尿胆素
排0.5~4mg/d
胆素原
( 2 ) 胆 素 原 的 肠 肝 循 环
生理意义: 可排出大部胆素原
血胆红素, 3) 胆红素与清蛋白以非共价结合,不牢固、易解离,称
游离胆红素, 4) 以上胆红素还未经肝处理,统称为未结合胆红素。
2、影响胆红素-清蛋白结合的因素
有机阴离子: 脂肪酸、胆汁酸、磺胺类、抗生素、水杨酸 与胆红素竞争与清蛋白结合使胆红素游离出来, 增加其透入细胞的可能性
血红素的生物合成

血红素的生物合成佚名成熟红细胞中,血红蛋白(hemoglolin,Hb)占红细胞内蛋白质总量的95%,它是血液运输O2的最重要物质,和CO2的送输亦有一定关系。
血红蛋白是由4个亚基组成的四聚体,每一亚基由一分子珠蛋白(globin)与一分子血红素(heme)缔合而成。
由于珠蛋白的生物合成与一般蛋白质相同,因此本节重点介绍血红素的生物合成。
血红素也是其它一些蛋白质,如肌红蛋白(myoglobin),过氧化氢酶(catalase),过氧化物酶(peroxidase)等的辅基。
因而,一般细胞均可合成血红素,且合成通路相同。
在人红细胞中,血红素的合成从早动红细胞开始,直到网织红细胞阶段仍可合成。
而成熟红细胞不再有血红素的合成。
(一)血红素的合成通路(过程)血红素合成的基本原料是甘氨酸、琥珀酰辅酶A及Fe++。
合成的起始和终末过程均在线粒体,而中间阶段在胞液中进行。
合成过程分为如下四个步骤:1.δ-氨基-γ-酮戊酸(δ-aminplevulinic acid,ALA)的生成:在线粒体中,首先由甘氨酸和琥珀酰辅酶A在ALA合成酶(ALa synthetase)的催化下缩合生成ALA。
ALA合成酶由两个亚基组成,每个亚基分子量为60,000。
其辅酶为磷酸吡哆醛。
此酶为血红素合成的限速酶,受血红素的反馈抑制。
2.卟胆原的生成:线粒体生成的ALA进入胞液中,在ALA脱水酶(ALa dehydrase)的催化下,二分子ALA脱水缩合成一分子卟胆原(prophobilinogen,PBG)。
ALA脱水酶由八个亚基组成,分子量为26万。
为含巯基酶。
3.尿卟啉原和粪卟啉原的生成:在胞液中,四分子PBG脱氨缩合生成一分子尿卟啉原Ⅲ(uroporphyrinogen Ⅲ,UPG Ⅲ)。
此反应过程需两种酶即尿卟啉原合酶(uroporphyrinogen synthetase)又称卟胆原脱氨酶(PBG deaminase)和尿卟啉原Ⅲ同合酶(uroporphyrinogen Ⅲ cosynthase)。
从血红蛋白血红素结构角度阐述一氧化碳中毒的生物化学原理

从血红蛋白血红素结构角度阐述一氧化碳中毒的生物化
学原理
一氧化碳中毒指的是人体吸入一氧化碳气体所引起的中毒现象。
一氧化碳是一种具有高度毒性的无色、无味、无臭的气体,它能和血红蛋白结合形成一氧化碳血红蛋白(COHb),导致血红蛋白无法有效地携带氧气,从而引起中毒。
血红蛋白是一种由四个氧气分子和四个血红素分子组成的复合物,它是红细胞中最主要的质量成分,也是氧气在体内的主要携带者。
血红蛋白的结构决定了它可以与氧气发生强烈的亲和力,从而形成氧合血红蛋白。
而一氧化碳通过与血红蛋白中的血红素结合,形成COHb,从而阻碍了血红蛋白与氧气的结合,导致氧气无法及时地携带和运输到体内各个组织和器官中,引起中毒反应。
血红素是血红蛋白中的一个重要组成部分,它是一种由四个呼吸色素分子(使血红蛋白呈现红色的分子)和单个铁离子组成的分子。
当一氧化碳与血红素相结合时,它会占据血红蛋白分子中一个铁离子的位置,并与血红蛋白分子中的其他组分相互作用,形成COHb,这样血红蛋白结构中的一个重要组成部分就被扰乱了,氧气的结合就会受到阻碍,因此会发生中毒反应。
总之,一氧化碳中毒的生物化学原理是指,一氧化碳通过与血红蛋白中的血红素发生结合,形成COHb,从而阻碍了氧气的结合,导致氧气无法及时地携带和运输到体内各个组织和器官中,引起中毒反应。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
电子传递的顺序为: 细胞色素b 细胞色素c1 细胞色素c
细胞色素氧化酶(细胞色素a 细胞色素a3) 氧分子
一、细胞色素c 细胞色素c是相对分子质量比较小且 相对比较容易结晶的蛋白,它的分布非常 广,存在于从细菌、酵母、植物、无脊椎 动物、脊椎动物到高等动物和人等所有的 原核生物和真核生物中。
细胞色素c是少数几个被详细研究的含 金属的生物大分子之一,现在已经确定了 几十种不同来源的细胞色素c的氨基酸序列, 每个细胞色素c分子含有一条蛋白链 和一
细胞色素b5 中铁离子除了与 卟啉环平面的四 个氮原子配位之 外,轴向位置被 两个组氨酸侧链 上的咪唑氮原子 占据。
X射线衍射晶体结构表明,细胞色素b5 中的血红素位于疏水袋中,原卟啉的两个丙 酸基指向蛋白表面,处在与水可接触的区域。 血红素周围有四个α螺旋链存在,其底部有β 折叠片存在,这样就构成了一个桶状疏水空 腔,血红素辅基正好位于其中。
图5.7 未结合底物分子的细胞色素P-450 血红素活性中心的结构
图5.7 结合樟脑底物分子的细胞色素P-450 血红素活性中心的结构
小 结: 细胞色素P-450是以血红素b为辅基的 一类金属蛋白,它是一种金属单加氧酶,可 以催化某些有机底物分子的加氧反应,使有 机底物分子发生羟基化反应。该酶对于生物 体系的代谢具有重要作用。
细胞色素P-450显示出非常广泛的底 物分子特异性,底物分子可以包括脂肪环、 脂肪链类碳氢化合物以及它们的衍生物、 芳香族碳氢化合物及其衍生物等。
细胞色素P-450催化的主要反应类型
某些有机化合物由于不溶于水,使其 在体内难以代谢,而在细胞色素P-450单 加氧酶作用下,使这些难代谢物进行羟基 化,将其转化成水溶性化合物。然后进一 步与其他水溶性物质结合,代谢后排出体 外,这一代谢过程对哺乳动物十分重要。
例如,在肝微粒体中已经发现多种细 胞色素P-450,可将外来有毒化合物、污 染物等催化氧化成水溶性物种排出体外, 从而起到保护机体不受伤害的作用。但是, 也有一些本来没有毒性或毒性较小的化合 物经过细胞色素P-450催化引入羟基之后 反而变成毒性更大的化合物。
生物体系中,在许多重要的代谢和生物 合成中,细胞色素P-450起了非常重要的作 用。例如,在肾上腺皮质中,细胞色素P- 450参与脂的代谢、胆甾醇的氧化等重要过 程。这些反应中的氢供体主要有还原型烟酰 胺腺嘌呤二核苷酸(NADH)、还原型烟酰 胺腺嘌呤核苷酸磷酸盐(NADPH)、还原 型黄素蛋白、维生素C等。
通过ESR、EXAFS等谱学表征,细胞色 素P-450的活性中心含有铁和原卟啉,还 原型的细胞色素P-450中铁为高自旋,易 与O2、CO、CN-等分子或离子结合,并转 变为低自旋,这一现象与肌红蛋白类似。
X射线衍射晶体结构证明,血红素铁的 轴向第五个配体来自于蛋白链上半胱氨酸残 基侧链的硫原子。氧化型细胞色素P-450中 在没有底物分子存在的条件下,第六配体为 水分子,而且在配位水分子附近聚集了数个 水分子形成一个水分子簇和物。当有底物分 子进入之后,水分子簇和物即被破坏,血红 素上方的空间被底物分子所占据。
细胞色素广泛存在于动物和植物组织 中。在动物中细胞色素的含量,以心脏和 其它活跃的运动肌(如鸟类和昆虫类的飞翔 肌)较高,肝、脑和非横纹肌次之,皮肤和 肺最低。故一般多从动物(如猪、牛)的心脏 提取。
1961年,国际生物化学学会把细胞色 素定义为“一种血红素蛋白,它的基本生 物功能是通过分子中血红素铁的价态的可 逆变化在生物体中起电子及氢的传递作 用”。目前已知的细胞色素有50种以上。
细胞色素b5的蛋白链由133个氨基酸残 基组成,分子质量约16000 Da。细胞色素 b5与血红蛋白、肌红蛋白有一定的相似之 处,也有明显的不同。
在细胞色素b5中,谱学测试、晶体结 构、溶液结构研究证实铁离子为配位饱和 的六配位构型,因此细胞色素b5难以自动 氧化,不易与CO、CN-、N3-等外来配体 结合。细胞色素b5中铁无论是氧化态还是 还原态都处于低自旋态。
血红素a3中的铁在还原态时为高自旋 的五配位构型,除了血红素平面四个氮原 子与铁配位外,一个轴向配体是组氨酸的 咪唑氮原子,另一个轴向位置为空,因此 很容易与CO、CN-等配体结合。谱学及晶 体结构研究推测,CuB中心的配位环境中有 组氨酸的咪唑氮原子参与配位,在还原态 时可能也有一个空位。
三、细胞色素b5 细胞色素b是一类含有原卟啉的电子 传递蛋白,主要存在于线粒体内膜。还原 型细胞色素b的α、β和γ特征吸收带分别在 555~563 nm、523~533 nm和420~430 nm。细胞色素b5是b类细胞色素中研究最 多、了解最清楚的一种蛋白,多见于真核 细胞,特别是动物的肝细胞中。
第五章
血红素蛋白及催化体系
在生物体内,除了含铁蛋白的氧载 体之外,还有多种含铁蛋白,在生物体 内起着传递电子、参与各种酶催化体系 以及铁自身的吸收、储存、运送等。
第一节 细胞色素:电子传递 第二节 细胞色素 P-450 第三节 甲烷单加氧酶、双加氧酶 第四节 过氧化物酶及过氧化氢酶
第一节
细胞色素:电子传递
自然界存在两种加氧酶:一种是单加氧 酶,即将氧分子中两个氧原子中的一个氧原 子催化加合到底物分子中,而另一个氧原子 被转换为水分子;另一种是双加氧酶,即将 氧分子中的两个氧原子都加合到底物分子中。
细胞色素P-450属于单加氧酶,催化单 加氧反应。反应式为: SH + O2 + 2H+ + 2e SOH + H2O 其中SH为底物分子。 细胞色素P-450催化的反应是在底物分 子中引入一个羟基 羟பைடு நூலகம்,即底物分子进行羟基化 羟基 反应,这种羟基化反应对各种有机分子在生 物体系中的代谢具有重要作用。
蛋白链 蛋白链
血红素c辅基除 了由两个轴向 配体与蛋白链 连接外,卟啉 环2,4-位以共价 键形式与蛋白 链中的两个半 胱氨酸连接。
二、细胞色素c氧化酶-- 细胞色素a和细胞色素a3 细胞色素c氧化酶是细胞色素c的氧化剂, 它是需氧生物呼吸链中的最后一步氧化还 原反应,它将还原态的细胞色素c催化氧化 到氧化态的细胞色素c,同时将一分子氧还 原为两分子水,反应式为:
在细胞色素P-450参与的各种催化羟化 反应中,研究最多、了解最清楚的是茨酮-5单氧酶,反应式为:
在黄素蛋白、铁硫蛋白和细胞色素P- 450的共同作用下,由烟酰胺腺嘌呤二核苷 酸(NADH)作为氢供体,在樟脑(2-茨酮) 分子的5-位上进行立体选择性羟基化反应, 催化该反应的细胞色素P-450简称为P- 450cam。
半胱氨酸
组氨酸
蛋氨酸
细胞色素c氧化酶中CuA中心的配位结构
2、细胞色素a3 磁性、ESR等研究发现氧化型细胞色素 a3中的Fe和Cu之间存在较强的相互作用, 推测Fe和Cu之间可能有某种桥连基团存在。 这两个非常接近的Fe和Cu氧化还原中心对 氧分子的四电子还原过程的顺利完成非常 氧分子的四电子还原过程 有利。
4Fe(II)-cyt c+ O2+4H+ 4Fe(III)-cyt c+2H2O
细胞色素c氧化酶存在于细胞的线粒体 膜上,该酶在催化氧化还原反应的同时还 伴随着质子通过膜的转移,即细胞色素c氧 化酶同时具有电子转移和质子泵的双重功 能。
细胞色素c氧化酶是由多个亚基组成的 复合蛋白,催化反应活性中心集中在I、II和 III三个亚基中,活性中心由低氧化还原电位 的细胞色素 细胞色素a和高氧化还原电位的细胞色素 细胞色素 细胞色素 a3组成。其中细胞色素a是接受还原态细胞 色素c提供电子的部位,而细胞色素a3则是 还原氧分子到水分子的部位。此外,细胞色 素c氧化酶中还含有锌离子 镁离子 锌离子和镁离子 锌离子 镁离子。
小 结: 细胞色素a、细胞色素b、细胞色素c是 包含血红素辅基的一类金属蛋白,它们依靠 血红素铁的价态变化来进行电子传递。 在线粒体内膜上电子传递的顺序为: 细胞色素b 细胞色素c1 细胞色素c 细胞色素氧化酶(细胞色素a 细胞色素a3) 氧分子
第二节 细胞色素 P-450
细胞色素P-450是于1958年在鼠肝微粒 体中发现的一种以血红素b为辅基的蛋白, 它也是b类细胞色素中的一种。与细胞色素c、 细胞色素b5等不同的是,细胞色素P-450在 生物体系中不是电子传递蛋白,而是一种金 属酶,它催化某些有机底物分子的加氧反应。
个血红素c辅基。马心的细胞色素c的分子
质量为12400 Da,它由104个氨基酸残基组 成。
图5.1 金枪鱼细胞色素c (第一个测定结构的细胞色素c)
用核磁共振(NMR)和X射线衍射单 晶结构分析对细胞色素c的研究表明,血红 素c中的铁离子具有六配位的八面体构型。 铁离子分别和卟啉环中的四个吡咯氮原子 以及一个蛋氨酸80的硫原子、一个组氨酸 18的咪唑氮原子配位。
在生物体系中,电子传递反应是一系列 重要生物过程的基本反应,所有的氧化还原 反应都会涉及电子的转移。由于铁、铜等过 渡金属离子有可变的化合价,通过其价态之 间的变化,如Fe(III)/Fe(II)、Cu(II)/Cu(I)等, 可以有效地进行电子传递。所以在生物体内 形成了一些含有铁、铜等过渡金属离子的蛋 白分子作为生物过程中的电子传递体。
1、细胞色素a 拉曼光谱、ESR等谱学研究表明,血 红素a中的铁无论是氧化态还是还原态都处 于六配位低自旋状态。铁离子除了与血红 素平面的四个氮原子配位之外,轴向两个 位置分别被来自于蛋白链中组氨酸侧链上 的咪唑氮原子占据。
细胞色素c氧化酶中血红素a铁的配位结构
CuA中心的结构由EXAFS、ESR以及利 用同位素标记的氨基酸研究结果,并最终根 据X射线衍射晶体结构结果确定为双核结构。 双核铜通过蛋白链中两个半胱氨酸侧链上的 硫原子桥连,一个铜离子还与一个组氨酸和 一个蛋氨酸配位,另一个铜离子还与一个组 氨酸和一个主链上的羰基氧原子配位。
在生物体内,根据所含血红素辅基结 构的不同,将细胞色素分为:细胞色素a (包含血红素a辅基)、细胞色素b(包含 血红素b辅基)、细胞色素c(包含血红素 c辅基)等。