第八章 天冬氨酸族氨基酸发酵机制
生物工程氨基酸工艺学考试试题

1.目前国内谷氨酸提取工艺主要有;等电点法、离子交换法和等电点-离子交法。
2.谷氨酸等电点为pH3.22 ,其溶解度受温度影响较大(较大、较小),其结晶型有2种,分为α-型结晶和β-型结晶,其中不易沉降分离的是β-型结晶型。
3.国内谷氨酸生产的碳源原料主要有淀粉糖和糖蜜两类,国内谷氨酸产生菌大多数为生物素(维生素H 或VH)缺陷型4.在生物合成氨基酸途径中,谷氨酸是谷氨酰胺(Gln)/鸟氨酸(Orn)/瓜氨酸(Cit)/精氨酸(Arg)(填氨基酸的名称,一种即可,下同)的前体,苏氨酸是异亮氨酸(Ile)的前体;.分枝酸是色氨酸(Trp)的前体。
5.谷氨酸的实际生产中,要求上柱发酵液的pH值控制在5.0~5.5 即可;上柱时,逆上柱流速控制在2~3 m3/(m3·h),顺上柱流速控制在1.5~2. 0 m3/(m3·h),同时注意控制流出的谷氨酸的量不要超过0.2 %,避免漏吸现象的发生。
6.氨基酸发酵产生菌种选育时,常规育种中一采用黄色短杆菌和谷氨酸棒杆菌为出发菌株。
7.要提高谷氨酸发酵产率,可以从选育高产菌种、采用流加液氨培养二级种子、采用大种量、高生物素、高初糖、连续流加高浓度糖的发酵工艺、通电发酵法、改进发酵设备、应用先进的计量和自控技术实现在线显示及自动操作等几个方面进行。
8.天冬氨酸族氨基酸生物合成共同途径上的第一个关键酶是天冬氨酸激酶(AK),在E.coli中,此酶由 3 种同工酶组成;从合成调节机制上看,天冬氨酸族氨基酸优先合成蛋氨酸(Met);在苏氨酸合成途径中,苏氨酸反馈抑制高丝氨酸脱氢酶(HD)三.名词解释(每题2分,共12分)1.末端产物阻遏:是指由某代谢途径末端产物的过量累积时而引起的反馈阻遏,是一种较为重要的反馈阻遏。
2.代谢调控:在发酵工业中,为了大量积累人们所需要的某一代谢产物,常人为地打破微生物细胞内的自动代谢调节机制,使代谢朝人们所希望的方向进行。
生物工艺原理习题集

⽣物⼯艺原理习题集《⽣物⼯艺原理》⼀、名词解释(共5题,每题3分,共15分)诱变选育⾃然选育⽣理碱性物质⽣理酸性物质前体⽣长因⼦产物促进剂代谢控制发酵巴斯德效应葡萄糖效应初级代谢产物次级代谢产物临界溶氧浓度发酵热⼆、填空题(,共20空,每空0.5分,共10分)1.⾃然界分离微⽣物的⼀般操作是_______________、_______________、________________、_________________________________。
2.从环境中分离⽬的微⽣物时,进⾏富集的⽬的是________________________________。
3.菌种选育分⼦改造的⽬的是_______________、______________、_________________、_____________、_________________。
4.培养基成分选择的原则是________________、________________、______________________、______________________________。
5.种⼦扩⼤培养的⽬的是_____________、_______________________、_______________________与要求__________________、_________________、_________________、____________________________。
6.种⼦扩⼤培养的⼀般步骤__________________、_________________、_______________________、_____________________________________。
7.在酿酒过程中影响杂醇油⽣成的原因是___________________、____________________、_______________________________________。
氨基酸发酵机制及过程

④沿着由柠檬酸至α–酮戊二酸的氧化途径,谷氨酸产 生菌有两种NADP专性脱氢酶,即异柠檬酸脱氢酶和L谷氨酸脱氢酶。 在谷氨酸的生物合成中,谷氨酸脱氢酶和异柠檬 酸脱氢酶在铵离子存在下,两者非常密切地偶联起 来,形成强固的氧化还原共轭体系,不与NADPH2 的末端氧化系相连接,使α–酮戊二酸还原氨基化生 成谷氨酸。
2.三羧酸循环(TCA循环)的调节
谷氨酸产生菌在代谢途径中,三羧酸循环的调节主要 是通过5种酶的调节进行的。这五种酶是磷酸烯醇式丙酮 酸羧化酶、柠檬酸合成酶、异柠檬酸脱氢酶、谷氨酸脱氢 酶和α-酮戊二酸脱氢酶。
草酰乙酸+谷氨酸
谷氨酸转氨酶
天冬氨酸+α -酮戊二酸
谷氨酸比天冬氨酸优先合成,谷氨酸合成过量后, 谷氨酸抑制谷氨酸脱氢酶的活力并阻遏柠檬酸合成酶 的合成,使代谢转向天冬氨酸的合成。
在谷氨酸发酵生产中,生物素缺陷型菌在 NH4+存在时,葡萄糖消耗速率快而且谷氨酸收率 高; NH4+不存在时,葡萄糖消耗速率很慢,生 成物是α–酮戊二酸、丙酮酸等物质,不产生谷氨 酸。
四、细胞膜通透性的调节
对谷氨酸发酵的重要性: 当细胞膜转变为有利于谷 氨酸向膜外渗透的方式, 谷氨酸才能不断地排出细 胞外,这样既有利于细胞 内谷氨酸合成反应的优先 性、连续性,也有利于谷 氨酸在胞外的积累。
4.CO2固定反应的调节
CO2固定反应主要通过以下途径完成:
C02的固定反应的作用:补充草酰乙酸; 在谷氨酸合成过程中,糖的分解代谢途径与C02固定 的适当比例是提高谷氨酸对糖收率的关键问题。
5.NH4+的调节
谷氨酸脱氢酶也能催化谷氨酸氧化脱氨反应,脱 氨过程以NAD+作为辅酶,该酶催化的反应虽然偏向 氨合成谷氨酸一边,但是脱氢过程产生的NADH被氧 化成NAD+,同时产生的NH3很容易被除去。 脱氨反应被NH4+和α–酮戊二酸所抑制,这对于谷 氨酸的积累也起到了很好的作用。
天冬氨酸族氨基酸发酵机制---赖氨酸(补充材料)

一、苏 氨酸的 育种途 径
苏氨酸高产菌株应具备的生化特征
DDP合成酶活力极弱或欠缺 琥珀酰高丝氨酸合成酶酶活力极弱或欠缺 苏氨酸脱氨酶酶活力极弱或欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协 同反馈抑制不敏感 高丝氨酸脱氢酶活力强,不受苏氨酸反馈调节。
三、蛋氨酸育种途径
高产蛋氨酸应具备的生化特性
DDP合成酶活力极弱或欠缺 S-腺苷蛋氨酸酶活力极弱或欠缺 高丝氨酸激酶欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协同 反馈抑制不敏感 高丝氨酸脱氢酶活力强,苏氨酸和异亮氨酸的反 馈调节不敏感、不受蛋氨酸反馈调节。
方法:
①选育丙氨酸缺陷型; ②选育抗天冬氨酸结构类似物突变株; ③选育适宜的CO2固定酶/TCA循环酶活性比突变株; 1.通过TCA循环 葡萄糖→丙酮酸→草酰乙酸→天冬氨酸 赖氨酸
2.通过磷酸烯醇式丙酮酸羧化反应
葡萄糖→磷酸烯醇式丙酮酸→草酰乙酸→天冬氨酸 丙酮酸 赖氨酸
6.改变细胞膜的透过性 7.选育温度敏感突变株 天冬氨酸系分枝代谢途径中,末端产物 种类多,调节机制复杂,为高效率生产 赖氨酸,可以采取顺次解除各种调节机 制的诱变育种,获得多重标记的突变株。
第八章 天冬氨酸族氨 基酸发酵机制
第一节
生物合成途径(天冬氨酸族) 及代谢调控机制
一、生物合成途径
天冬氨酸族氨基酸合成可以以草酰乙酸 或天冬氨酸为原料,合成苏氨酸、异亮氨酸、 蛋氨酸和赖氨酸。
葡萄糖
EMP
丙酮酸
CO2 固定、氧化
氨基化反应
草酰乙酸
天冬氨酸激酶
8氨基酸发酵机制

芳香族氨基酸(Phe、Tyr、Trp)生物合 芳香族氨基酸(Phe、Tyr、Trp)生物合 成途径的主导关键酶DAHP(2成途径的主导关键酶DAHP(2-酮-3-脱氧-D脱氧阿拉伯糖型庚糖酸- 磷酸) 阿拉伯糖型庚糖酸-7-磷酸)合成酶在大肠杆 菌中也是有3 菌中也是有3种同功酶,分别受酪氨酸、苯 丙氨酸和色氨酸的反馈调节;
要想选育出某种目的产物生产菌,首先 要了解这种物质的生物合成途径、关键酶以 及关键酶受到怎样的反馈调节,再考虑如何 解除反馈调节,随之设计出正确的代谢改造 方案来。
图8-3可以看 出,参与氨基酸 生物合成的关键 酶主要有12种: 酶主要有12种: ①磷酸果糖激酶; ②柠檬酸合成酶; ③N-乙酰谷氨酸 激酶;④鸟氨酸 转氨基甲酰酶; ⑤天冬氨酸激酶; ⑥高丝氨酸脱氢 酶;⑦苏氨酸脱 水酶;⑧α 水酶;⑧α-乙酰 乳酸合成酶;⑨ DAHP合成酶; DAHP合成酶; ⑩分支酸变位酶; ⑾预苯酸脱水酶; ⑿预苯酸脱氢酶。
已经证明,微生物细胞具有高度适应 环境和繁殖的能力,细胞的各种机构能协 调地进行工作,对环境的刺激和信息作出 反应,进行自我调节。
可以说微生物活细胞是个远离平衡状态 的开放体系,从微生物细胞对能量和化学物 质的内外交换、增收节支、协调规律的客观 存在出发,可以把微生物细胞作为按特殊的 经济规律运行的经济实体看待,并且把这种 按特殊的经济规律运行的有利于生存竞争的 பைடு நூலகம்陈代谢特性称作细胞经济性。
微生物细胞的经济性是在自然选择的过 程中逐渐形成的。微生物细胞的生命过程就 是细胞经济运行的过程。一般地说,细胞经 济性可以以生成细胞的质量与消耗底物/ 济性可以以生成细胞的质量与消耗底物/基 质的质量之比值即细胞经济系数来衡量。
微生物在生存竞争中进化的方向是发展 自身的适应能力和提高细胞运行的经济系数。 经生存竞争而幸存下来的野生型微生物在其 所处的环境中是富有竞争能力的,并且它们 的代谢流量在代谢网络中的分布及细胞经济 运行状况有利于细胞生长、繁殖和在竞争中 获胜。
第九章天冬氨酸族氨基酸发酵

谷氨酸
N-乙酰谷氨酸合成酶
N-乙酰谷氨酸 N-乙酰谷氨酸磷酸
N-乙酰谷氨酸和N-乙酰 鸟氨酸乙酰基转移酶
N-乙酰鸟氨酸半醛 共轭反应催化
N-乙酰鸟氨酸
N-乙酰鸟氨酸酶
鸟氨酸 瓜氨酸 精氨酸琥珀酸 精氨酸
精氨酸发酵
精氨酸是该代谢途径的最终产物,且合成 途径没有分支代谢。
不能采用阻断代谢流、选育缺陷型的方法 来选育生产菌。
改善膜的通透性 增加前体物质的合成 切断支路,增加前体物质。 1)选育丙氨酸缺陷型 2)选育抗天冬氨酸结构类似物 3)选育合适的co2固定和TCA循环比
选育温度敏感型突变株 选育脲酶回复突变株 利用基因工程技术构建工程菌。
第五节 苏氨酸发酵 苏氨酸高产菌具备的生化特征 1)二氧化碳固定反应强 2)天冬氨酸合成能力强 3)天冬氨酸激酶活性强 4)高丝氨酸脱氢酶活力强 5)二氢二吡啶二羧酸合成酶活力弱或丧失 6)琥珀酰高丝氨酸转琥珀酰酶活力弱或丧失
色氨酸合成酶、丝氨酸消旋酶 吲哚+DL-丝氨酸 L-色氨酸+水
色氨酸酶 吲哚+丙酮酸+氨 L-色氨酸+水
色氨酸合成酶、丝氨酸消旋酶 吲哚+L-丝氨酸 L-色氨酸+水
三种水解酶 IMH L-色氨酸+氨+水
2.微生物转化法 3.直接发酵法 4.其他方法 二、L-色氨酸育种策略 1.切断支路代谢 2.解除自身反馈抑制 3.增加前提物质的合成 4.切断进一步代谢 5.其他方法
7)谷氨酸脱氢酶活力弱 8)苏氨酸脱氨酶活力弱或丧失
切断支路 天冬氨酸 二氢吡啶-2,6-二羧酸 赖氨酸 天冬氨酸半醛 高丝氨酸 苏氨酸 Ο-琥珀酰-高丝氨酸 强化代谢
蛋氨酸
第八章天冬氨酸族氨基酸发酵机制ppt课件

第三节 苏氨酸、蛋氨酸、高丝氨酸和天冬氨 酸的育种途径
一、苏 氨酸的 育种途 径
苏氨酸高产菌株应具备的生化特征
DDP合成酶活力极弱或欠缺 琥珀酰高丝氨酸合成酶酶活力极弱或欠缺 苏氨酸脱氨酶酶活力极弱或欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协
按照起始物可将氨基酸的合成分成几个家族:
㈠谷氨酸族(α-酮戊二酸族)
包括:谷氨酸、谷氨酰胺、精氨酸、赖氨酸和 脯氨酸;
㈡丙酮酸族
包括:丙氨酸、缬氨酸、亮氨酸;
㈢天冬氨酸族(草酰乙酸族) 包括:天冬氨酸、天冬酰胺、苏氨酸和异亮
氨酸; ㈣磷酸甘油酸族
包括:甘氨、酪氨酸、色氨酸; 另外,组氨酸的合成为单独的一条途径。
4.解除代谢互锁
①选育亮氨酸缺陷型菌株,或者以抗AEC的 赖氨酸的生产菌为出发菌株,经诱变得到抗 AEC兼抗亮氨酸缺陷型菌株。
②选育抗亮氨酸结构类似物的突变株,从 遗传上解除亮氨酸对DDP合成酶的阻遏。
③选育对苯醌或喹啉衍生物敏感菌株,这 是一种寻找亮氨酸渗漏缺陷型菌株的有效方 法。
5.增加前体物的生物合成和阻塞产物的生成:
同反馈抑制不敏感 高丝氨酸脱氢酶活力强,不受苏氨酸反馈调节。
二、高丝氨酸菌种的选育途径
高产高丝氨酸菌株应具
DDP合成酶活力极弱或欠缺
琥珀酰高丝氨酸合成酶酶活力极弱或欠缺
高丝氨酸激酶欠缺
CO2固定能力强 天冬氨酸合成能力强
天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协同 反馈抑制不敏感
高丝氨酸脱氢酶活力强,不受苏氨酸和蛋氨酸反 馈调节。
HD HDⅠ:受苏氨酸反馈抑制 HDⅡ:受蛋氨酸阻遏
⑤苏氨酸脱氨酶(异亮氨酸合成途径的酶) 受异亮氨酸的反馈抑制和阻遏
天冬氨酸族氨基酸发酵机制

优化发酵条件提高产量
通过实验手段优化天冬氨酸族氨基酸发酵的条件,如温度、pH、溶氧浓 度、碳氮源等,以提高发酵效率和产物产量。
开发新型的细胞培养技术,如连续培养、高密度培养等,以实现细胞的高 密度生长和产物的高效积累。
结合代谢工程和基因工程技术,对菌株进行遗传改造,提高其发酵性能和 产物产量。
开发新型天冬氨酸族氨基酸生产菌株
营养强化剂
天冬氨酸族氨基酸可添加到婴儿食品、运动员食品等特殊食品中,以满足特定营养需求。
在农业领域的应用前景
植物生长调节剂
天冬氨酸族氨基酸可促进植物生长,提 高产量,可作为潜在的植物生长调节剂 。
VS
饲料添加剂
天冬氨酸族氨基酸可添加到动物饲料中, 提高动物生长性能和饲料转化率。
06
研究展望
深入探究天冬氨酸族氨基酸发酵机制
天冬氨酸族氨基酸发酵机 制
• 引言 • 天冬氨酸族氨基酸发酵机制概述 • 天冬氨酸族氨基酸发酵的生物合成途
径 • 天冬• 研究展望
01
引言
氨基酸发酵的重要性
生命活动的基础
氨基酸是构成蛋白质的基本单位,是 生物体进行生命活动不可或缺的物质。
04
天冬氨酸族氨基酸发酵的代谢调控
代谢调控机制
酶的活性调节
通过调节酶的活性来控制代谢过 程,例如通过调节天冬氨酸族的 合成酶活性来控制氨基酸的合成。
代谢流控制
通过控制代谢流的方向和流量来调 节氨基酸的合成,例如通过控制糖 酵解和三羧酸循环的代谢流来影响 天冬氨酸族的合成。
代谢物阻遏
通过调节细胞内代谢物的浓度来控 制基因的表达和酶的活性,从而影 响氨基酸的合成。
通过诱变育种、基因重组等手段,发掘和培育具有优良发酵性能的新型天冬氨酸族 氨基酸生产菌株。
氨基酸合成路径解析

氨基酸合成路径解析氨基酸是构成蛋白质的基本单位,对于生命活动至关重要。
了解氨基酸的合成路径不仅有助于我们深入理解生物化学的基本原理,还在医学、农业等领域具有重要的应用价值。
氨基酸的合成可以分为两大类:从头合成和补救合成。
从头合成是指利用简单的前体物质,如碳水化合物、无机氮等,通过一系列酶促反应逐步合成氨基酸。
补救合成则是利用已有的氨基酸或其降解产物重新合成所需的氨基酸。
首先来谈谈一些常见氨基酸的从头合成路径。
比如,丙氨酸的合成相对简单。
它可以由丙酮酸在谷丙转氨酶的催化下,与谷氨酸进行转氨基反应而生成。
谷氨酸的合成较为复杂。
在植物和微生物中,谷氨酸是通过谷氨酸脱氢酶催化α酮戊二酸的还原氨基化反应来合成的。
这个过程中,需要氨作为氮源,同时还原型辅酶Ⅱ(NADPH)提供氢。
赖氨酸的合成路径则涉及多个步骤和多种酶。
在细菌中,赖氨酸的合成起始于天冬氨酸,经过一系列反应生成高丝氨酸,然后再转化为二氨基庚二酸,最终形成赖氨酸。
对于芳香族氨基酸,如苯丙氨酸、酪氨酸和色氨酸,它们的合成前体是赤藓糖-4-磷酸和磷酸烯醇式丙酮酸。
这两种物质经过一系列复杂的反应,逐步形成芳香环结构,最终合成相应的氨基酸。
在氨基酸的合成过程中,关键的因素包括前体物质的供应、酶的活性以及代谢调节。
前体物质的充足供应是保证氨基酸合成的基础。
而酶的活性则直接影响反应的速率和效率。
代谢调节在氨基酸合成中起着至关重要的作用。
细胞通过多种方式对氨基酸的合成进行调节,以确保合成的氨基酸能够满足需求,同时避免过度合成造成浪费。
例如,通过反馈抑制机制,当细胞内某种氨基酸的浓度达到一定水平时,会抑制其合成途径中关键酶的活性,从而减少该氨基酸的合成。
除了从头合成,补救合成也在维持细胞内氨基酸的平衡中发挥着作用。
例如,嘌呤核苷酸循环可以将天冬氨酸转化为延胡索酸和氨,氨可以用于合成其他氨基酸。
在医学领域,对氨基酸合成路径的研究有助于理解某些疾病的发病机制,并为药物研发提供靶点。
氨基酸发酵机制及过程共70页文档

26、要使整个人生都过得舒适、愉快,这是不可能的,因为人类必须具备一种能应付逆境的态度。——卢梭
▪
27、只有把抱怨环境的心情,化为上进的力量,才是成功的保证。——罗曼·罗兰
▪
28、知之者不如好之者,好之者不如乐之者。——孔子
▪
29、勇猛、大胆和坚定的决心能够抵得上武器的精良。——在明眼的跛子肩上。——叔本华
谢谢!
70
氨基酸发酵机制及过程
56、死去何所道,托体同山阿。 57、春秋多佳日,登高赋新诗。 58、种豆南山下,草盛豆苗稀。晨兴 理荒秽 ,带月 荷锄归 。道狭 草木长 ,夕露 沾我衣 。衣沾 不足惜 ,但使 愿无违 。 59、相见无杂言,但道桑麻长。 60、迢迢新秋夕,亭亭月将圆。
▪
天冬氨酸族氨基酸发酵

二、高丝氨酸高产菌的定向育种策略
天冬氨酸发酵
一、天冬氨酸高产菌应具备的生化特征
1.天冬氨酸激酶丧失 2.谷氨酸脱氢酶活力微弱 3.丙酮酸氨化酶微弱或丧失 4.二氧化碳固定反应能力强 5.柠檬酸合成酶活力弱 6.草酰乙酸氨基反应强
二、天冬氨酸高产菌的育种策略
1.解除反馈调节
天冬氨酸对磷酸烯醇丙酮酸羧化酶存在着反馈抑制作用,天冬氨酸 合成过量后反馈抑制磷酸烯醇丙酮酸羧化酶的活性,是天冬氨酸生物 合成的速度减慢或停止,所以,必须解除天冬氨酸对磷酸烯醇丙酮酸 羧化酶的反馈抑制。选育天冬氨酸结构类似物抗性突变株。
③ 二氢吡啶-2,6-二羧酸还原酶 受赖氨酸的反馈抑制。 ④ O-琥珀酰高丝氨酸转琥珀酰酶和半胱氨酸脱硫化氢酶受的受蛋氨酸的反 馈阻遏
⑤ 高丝氨酸激酶受苏氨酸的反馈阻遏
⑥ 苏氨酸脱氨酶受异亮氨酸的反馈抑制 在大肠杆菌的天冬氨酸族氨基酸生物合成中,当某一终产物如 苏氨酸过量时,只能抑制AkⅠ和HDⅠ及自身分支点的高丝氨酸激 酶,限制苏氨酸的合成,并不影响蛋氨酸和赖氨酸的合成。因此, 大肠杆菌中这样的调节模式对氨基酸生产菌的选育是不利的。
3.增加前体物天冬氨酸的合成
天冬氨酸是苏氨酸生物合成的前体物,增加天冬氨酸的合成,是苏 氨酸得以大量积累的必要条件。 (1)选育天冬氨酸结构类似物抗性突变株 (2)选育丙氨酸缺陷突变株 措施(3)强化从丙酮酸得到苏氨酸的代谢流 (4)谷氨酸优先合成转换 (5)利用平衡合成
5.利用现代生物技术选育苏氨酸生产菌
二、选育苏氨酸生产菌的方法
1.切断支Leabharlann 代谢在苏氨酸发酵代谢控制育种上,切断或减弱除苏氨酸以外的能控 制共同酶的赖氨酸和蛋氨酸的合成途径,一方面降低了这些物质的浓 度,解除了它们对代谢途径中关键酶的反馈调节作用,另一方面还可 以节省碳源,使代谢流高效地转向所需的目的产物——苏氨酸,并使 之得以大量合成。因此,选育和利用营养缺陷型突变株(或营养渗漏 型突变株、营养缺陷回复突变株)是生产苏氨酸的有效措施。
氨基酸发酵

• 典型的例子就是谷氨酸发酵。 • 改变培养条件的发酵转换法中,有变化铵离
子浓度、磷酸浓度,使谷氨酸转向谷氨酰 胺和缬氨酸发酵
共一百零六页
2.2 用营养缺陷 变异株的 (quēxiàn) 方法
• 这一方法是诱变出菌体内氨基酸生物合成
某步反应阻遏的营养缺陷型变异(biànyì)体,使 生物合成在中途停止,不让最终产物起控 制作用。
共一百零六页
3.1.2 载体 的构建 (zàitǐ)
• 有效的载体需要有在受体菌中可启动的复
制起始位点,这可从棒状杆菌家族内源小 质粒中获得;
• 载体所需的筛选标记(biāojì)及外源基因插入的
多克隆位点,可从常用的克隆载体中获得。
共一百零六页
3.1.3 基因 转移手段 (jīyīn)
• 由于棒状杆菌是革兰氏阳性菌,CaCl2转化法对它不
今后的发展是采用诱变、细胞工程、基因工程的手段选育出从遗传角度解 除了反馈调节和遗传性稳定的更理想菌种,提高产酸;采用过程控制,进行最优化控制, 连续化、自动化,稳产、高产;探求新工艺、新设备,以提高产率和收得率;研究发酵 机制等问题,以便能更好地控制氨基酸这样微生物中间代谢产物的发酵。
共一百零六页
• 专性需氧菌,控制环境(huánjìng)条
件可改变代谢途径和产物。
共一百零六页
控制 细胞渗透性 (kòngzhì)
•生物素、油酸和表面活性剂, 引起细胞膜的脂肪酸成分的改变。 •青霉素:抑制(yìzhì)细胞壁的合 成,由于细胞面内外的渗透压 而泄露出来。
共一百零六页
控制 旁路代谢 (kòngzhì)
育成的氨基酸生物合成高能力的菌株。
共一百零六页
第8章发酵机制 51-100
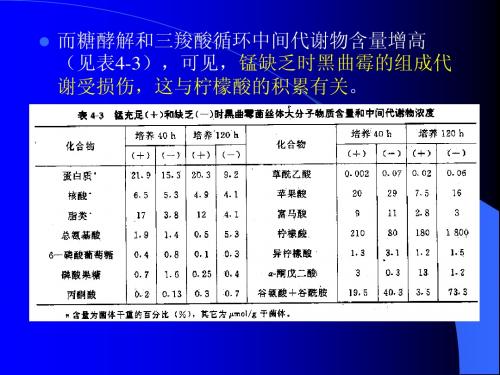
实验证明,谷氨酸产生菌的谷氨酸生物合成途 径与一般微生物是一样的,没有特殊的途径, 就是糖经过酵解途径(EMP)和单磷酸己糖途 径(HMP)生成丙酮酸。一方面,丙酮酸氧化 脱羧生成乙酰-CoA;另一方面,经CO2固定作 用生成草酰乙酸;两者合成柠檬酸进入三羧酸 循环(TCA循环),由三羧酸循环的中间产物 α–酮戊二酸,在谷氨酸脱氢酶催化下,还原氨 基化合成谷氨酸。
Mn2+ 对棒杆菌核苷酸积累的专一性影响 和上述例子极为相似。关于核苷酸的积 累,人们推断是因为催化核糖核苷酸形 成聚合阶段第一步反应的酶需要锰。
某些真菌的丙酮酸是酵解过程第二个调 节点已被证明,但是关于黑曲霉尚未被 证明。用交换定理未能确定这一步反应 是否调节性。测定柠檬酸发酵时酵解中 间代谢物浓度可推断流经丙酮酸激酶的 通量增加。
当乙酰 -CoA 氧化,使 ATP 水平提高,结 果丙酮酸激酶受 ATP抑制, PEP又向 CO2 固定。当生物合成和需能反应使 ATP 浓 度降低,则 ATP 对丙酮酸激酶的抑制被 解除,此酶被 AMP激活,使乙酰-CoA生 成量增加。
经过深入研究,他们通过实验证明在黄 色短杆菌中存在α-酮戊二酸脱氢酶络合 物,并发现该酶络合物极不稳定。该酶 的活力很弱,使糖代谢流进入TCA循环 后受阻在α-酮戊二酸处,在有NH存在下, 由谷氨酸脱氢酶催化还原氨基化生成谷 氨酸。
这一点也为木下等对TCA循环中有机酸的氧化 能的研究结果所证实,如表5-2所示。柠檬酸、 顺乌头酸、异柠檬酸和α-酮戊二酸的氧化能力 几乎没有,即α-酮戊二酸脱氢酶活性很弱。
二、TCA、DCA和CO2固定作用 1. TCA循环 糖经过 EMP、HMP 途径生成丙酮酸,在 有氧条件下,氧化脱羧生成乙酰 -CoA, 另一方面经CO2固定生成草酰乙酸,两者 在柠檬酸合成酶催化下生成柠檬酸,进 入TCA循环。
氨基酸发酵工艺学

第二节、氨基酸发酵菌株的育种
是氨基酸代谢控制发酵的基本策略之一
发酵工程要求微生物大量地合成特定的代 谢产物,这一目的只有当微生物的部分代 谢调控机制遭到破坏时才能达到。用人工 诱变的方法有目的地改变微生物固有的调 节机制,使合成产物的途径畅通无阻,最 大限度地积累特定产物,这种发酵称为代 谢控制发酵。
1、 用野生菌株的方法
分离的野生菌株具备积累产物的特性,可用 于直接发酵(产率低)。如谷氨酸发酵。
通过转换发酵,可延伸获得其它产物。主要 采用改变培养条件。如谷氨酸发酵中改变铵 离子浓度、磷酸浓度,使谷氨酸转向谷氨酰 胺和缬氨酸发酵。
2.
用营养缺陷变异株的方法
通过诱变出 85000 105000 45000 5000 4000 4000 55000
谷氨酸
谷氨酸 谷氨酸 谷氨酸 色氨酸 色氨酸 色氨酸 色氨酸 色氨酸 色氨酸 甘氨酸
味之素
日本旭化成 协和发酵 日本武田药品 味之素 昭和电工 三井东压 田造制药 日本化药 协和发酵 日本有机合成化学
第一节、氨基酸概论
1、氨基酸简介
– 构成蛋白质的基本分子单元。 – α 碳原子分别以共价键连接氢原子、羧基和氨
基及侧链。侧链不同,氨基酸的性质不同。 – 目前世界上可用发酵法生产氨基酸有20多种。
NH2 CH R COOH + NH3 CH R COO -
2、氨基酸的用途
(1)食品工业: 强化食品:赖氨酸, 苏氨酸,色氨酸于小麦 中 增鲜剂:谷氨酸单钠 和天冬氨酸 苯丙氨酸与天冬氨酸 可用于制造低热量二肽 甜味剂(α-天冬酰苯丙 氨酸甲酯),此产品1981 年获FDA批准,现在每年 产量已达数万吨。
第三节
氨基酸发酵的代谢控制
第三篇第四章天冬氨酸族氨基酸发酵机制

§第七章天冬氨酸族氨基酸发酵机制第一节天冬氨酸族氨基酸生物合成途径及代谢调节机制一、天冬氨酸族氨基酸生物合成途径GlucoseEMP丙酮酸草酰乙酸Asp天冬氨酸激酶(AK)天冬氨酰磷酸(asp-p)天冬氨酸β-半醛DDP合成酶(PS)高丝氨酸脱氢酶(HD)二羟吡啶羧酸(DDP)高丝氨酸(Hos)琥珀酰高丝氨酸合成酶高丝氨酸激酶二氨基庚二酸(DAP)琥珀酰高丝氨酸ThrLys 苏氨基酸脱氨酶Met Ile二、天冬氨酸族氨基酸生物合成的代谢调节机制1、大肠杆菌中天冬氨酸族氨基酸生物合成的调节机制GlucoseEMP丙酮酸草酰乙酸Asp(天冬氨酸激酶AK,同功酶)天冬氨酸磷酸(asp-p)天冬氨酸β-半醛(同功酶)二羟吡啶羧酸高丝氨酸(Hos)Lys琥珀酰高丝氨酸 O-磷酸高丝氨酸Met Thr大肠杆菌天冬氨酸族氨基酸代谢特点:生物合成途径要比黄色短杆菌、谷氨酸棒杆菌、乳糖发酵短杆菌的代谢调控要复杂,其过程如下:关键酶:天冬氨酸激酶是一个同功酶,分别受三个代谢产物的抑制,这三个终产物分别是:Lys、Met和Thr,只有当这三个代谢产物同时过量时,Asp激酶的活性才能完全被抑制。
同功酶:几种在同一细胞中催化同一反应的酶,但其活性受不同代谢产物体调节。
2、谷氨酸棒杆菌,黄色短杆菌天冬氨酸族氨基酸生物合成的调控GlucoseEMP丙酮酸草酰乙酸Asp(天冬氨酸激酶,AK)天冬氨酸磷酸(asp-p)天冬氨酸β-半醛二羟吡啶羧酸高丝氨酸LysO-琥珀酰高丝氨酸 O-磷酸高氨酸Met Thr黄色短杆菌与大肠杆菌(E.coli)的区别:(1)天冬氨酸激酶(AK),在黄色短杆菌中是一个变构酶,并有两个活性中心,分别受Lys、Thr的协同反馈抑制(2)黄色短杆菌中,存在两个分支点的优先合成机制:P75如图所示),即优先合成Hos,然后再优先合成Met,当Met过量时,阻遏:催化Hos 琥珀酰高丝氨酸所需要的酶的合成(即,琥珀酰高丝氨酸合成酶),使代谢流向合成Thr的方向进行,当Thr过量时,反馈抑制:Asp-β-半醛 Hos所需要的酶的的活性(即高丝氨酸脱氢酶),使代谢流向Lys的合成上。
发酵制品学代谢调控发酵机制培训课件

有过量的NH4+ 存在,-酮戊二酸经氧化还原共轭氨基化反应而 生成谷氨酸却不形成蛋白质,从而分泌泄漏于菌体外;
同时,谷氨酸生产菌应不利用体外的谷氨酸,使谷氨酸成为最
终产物。
发酵制品学代谢调控发酵机制
18
从前图还可以看出: 生产菌株还应该具有生物素合成缺陷、油酸合成
缺陷和甘油合成缺陷等特点。
发酵制品学代谢调控发酵机制
发酵制品学代谢调控发酵机制
12
2. 谷氨酸代谢调节机制
①谷氨酸脱氢酶 ②-酮戊二酸脱氢酶 ③磷酸烯醇丙酮酸羧化酶 ④柠檬酸合成酶
NH4+
在黄色短杆菌中谷氨酸、天冬氨酸生物合成的调节机制
发酵制品学代谢调控发酵机制
13
▪ 在微生物的代谢中,Glu比Asp优先合成; 合成过量时则抑制谷氨酸脱氢酶,使代谢转向合成Asp; Asp过量时反馈抑制PEP羧化酶的活力,停止合成草酰乙酸。
发酵制品学代谢调控发酵机制
35
在黄色短杆菌、谷氨酸棒杆菌等微生物中,AK是单一的, 并且受Lys 和 Thr的协同反馈抑制,反馈调节易于解除,使 育种简单化,所以常常被用作氨基酸发酵育种的出发菌株。
黄色短杆菌的AK受Lys和Thr协同反馈情况
发酵制品学代谢调控发酵机制
36
▪ 乳糖发酵短杆 菌中赖氨酸及其 前体物生物合成 的代谢调节
▪ NH4+的导入不仅仅证明Glu是氮素同化发酵,它还会抑制 Glu生成的逆反应,因此当NH4+存在时,葡萄糖的消耗速度 很快, Glu的生成很高;但是当生物素充足时,NH4+几乎 不影响糖代谢。
发酵制品学代谢调控发酵机制
14
• Glu生产菌大多是生物素缺陷型,发酵时控制生物素亚适 量,使细胞变形拉长,改变了细胞膜的通透性引起代谢失 调使Glu得以积累。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
方法:
①选育丙氨酸缺陷型; ②选育抗天冬氨酸结构类似物突变株; ③选育适宜的CO2固定酶/TCA循环酶活性比突变株; 1.通过TCA循环 葡萄糖→丙酮酸→草酰乙酸→天冬氨酸 赖氨酸
2.通过磷酸烯醇式丙酮酸羧化反应
葡萄糖→磷酸烯醇式丙酮酸→草酰乙酸→天冬氨酸 丙酮酸 赖氨酸
6.改变细胞膜的透过性 7.选育温度敏感突变株 天冬氨酸系分枝代谢途径中,末端 产物种类多,调节机制复杂,为高效率 生产赖氨酸,可以采取顺次解除各种调 节机制的诱变育种,获得多重标记的突 变株。
四、天冬氨酸的育种途径
高产天冬氨酸菌株应有的生化特征
天冬氨酸激酶丧失 丙酮酸氨基化酶丧失 CO2固定酶系丙酮酸氧化脱羧要强 草酰乙酸氨基化反应强。
复习思考题
简述黄色短杆菌赖氨酸生物合成的调节 机制 简述赖氨酸生产菌的选育原则
二氢吡啶二羧酸(DDP)合成酶
高丝氨酸脱氢酶
高丝氨酸
高丝氨酸激酶
蛋氨酸 苏氨酸
苏氨酸脱氨酶
异亮氨酸
二、天冬氨酸族生物合成的代谢调节机制
在细菌中,虽然天冬氨酸族氨基酸生 物合成途径是相同的,但是其代谢调节机 制是多种多样的。 1.大肠杆菌K12
①天冬氨酸激酶( AK)
AKⅠ:受苏氨酸、异亮氨酸的多价抑制和阻遏 天冬氨酸激酶 AKⅡ:受蛋氨酸专一性抑制和阻遏 AKⅢ:受赖氨酸专一性抑制和阻遏
4.解除代谢互锁 ①选育亮氨酸缺陷型菌株,或者以抗AEC的赖 氨酸的生产菌为出发菌株,经诱变得到抗 AEC兼抗亮氨酸缺陷型菌株。 ②选育抗亮氨酸结构类似物的突变株,从遗 传上解除亮氨酸对DDP合成酶的阻遏。 ③选育对苯醌或喹啉衍生物敏感菌株,这是 一种寻找亮氨酸渗漏缺陷型菌株的有效方法。
5.增加前体物的生物合成和阻塞产物的生成:
3.乳糖发酵短杆菌赖氨酸合成调节 P75,图65 1)存在代谢优先向苏氨酸和蛋氨酸方向进行。 2)与黄色短杆菌区别: AK受赖氨酸+苏氨酸的反馈抑制 存在代谢互锁,DDP合成酶的合成受亮氨酸阻 遏。DDP还原酶受半胱氨酸和丙氨酸抑制。
三、酵母和霉菌的赖氨酸生物合成途径和 调节机制
所谓同型TCA循环途径进行,但生物合 成酶与TCA循环酶完全两样。
三、蛋氨酸育种途径
高产蛋氨酸应具备的生化特性
DDP合成酶活力极弱或欠缺 S-腺苷蛋氨酸酶活力极弱或欠缺 高丝氨酸激酶欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协同 反馈抑制不敏感 高丝氨酸脱氢酶活力强,苏氨酸和异亮氨酸的反 馈调节不敏感、不受蛋氨酸反馈调节。
按照起始物可将氨基酸的合成分成几个家族:
㈠谷氨酸族(α-酮戊二酸族) 包括:谷氨酸、谷氨酰胺、精氨酸、赖氨酸和 脯氨酸; ㈡丙酮酸族 包括:丙氨酸、缬氨酸、亮氨酸;
㈢天冬氨酸族(草酰乙酸族) 包括:天冬氨酸、天冬酰胺、苏氨酸和异亮氨 酸; ㈣磷酸甘油酸族 包括:甘氨酸、丝氨酸和半胱氨酸; ㈤芳香族 包括:苯丙氨酸、酪氨酸、色氨酸; 另外,组氨酸的合成为单独的一条途径。
二、高丝氨酸菌种的选育途径
高产高丝氨酸菌株应具备的生化特征
DDP合成酶活力极弱或欠缺 琥珀酰高丝氨酸合成酶酶活力极弱或欠缺 高丝氨酸激酶欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协同 反馈抑制不敏感 高丝氨酸脱氢酶活力强,不受苏氨酸和蛋氨酸反 馈调节。
第八章 天冬氨酸族氨 基酸发酵机制
第八章 天冬氨酸族氨基酸发酵机制
重点:天冬氨酸族氨基酸生物合成途径及代谢
调节机制;天冬氨酸族氨基酸生物合成的代谢 调节机制;酵母和霉菌的赖氨酸生物合成途径 和调节机制;赖氨酸生产菌的育种途径;
难点:天冬氨酸族氨基酸生物合成的代谢
调节机制;
氨基酸本身的合成在不同生物体中,有较大的差 异,然而许多氨基酸的合成途径在不同生物体中 也有共同之处。
8.应用细胞工程和遗传工程育种 9.防止高产菌株回复突变 选育遗传性稳定的菌株 定向赋加生产菌多个遗传标记 菌种培养和保藏时,培养基要丰富,尤其有足 够的要求营养物 培养时添加抗生素,抑制恢复突变株的生长, 达到分纯的目的。
第三节 苏氨酸、蛋氨酸、高丝氨酸和天冬氨 酸的育种途径
第二节
赖氨酸生产菌的育种途径
出发菌株的选择:要求代谢比较简单的细 菌(如黄色短杆菌、谷氨酸棒杆菌、乳酸发酵 短杆菌等) 1.优先合成的转换:渗漏缺陷型的选育。 降低高丝氨酸脱氢酶活性,使代谢流转向优 先合成赖氨酸。 高丝氨酸脱氢酶渗漏缺陷型。
2.切断支路代谢:营养缺陷型的选育。 黄色短杆菌、谷氨酸棒杆菌,对天冬 氨酸激酶的反馈抑制是赖氨酸+苏氨酸的协 同作用,通过诱变使高丝氨酸脱氢酶缺失, 切断通向苏氨酸、蛋氨酸的代谢流,控制 高丝氨酸(或苏氨酸+蛋氨酸)量,降低苏 氨酸浓度,解除协同反馈作用,过量积累 赖氨酸。
一、苏 氨酸的 育种途 径
苏氨酸高产菌株应具备的生化特征
DDP合成酶活力极弱或欠缺 琥珀酰高丝氨酸合成酶酶活力极弱或欠缺 苏氨酸脱氨酶酶活力极弱或欠缺 CO2固定能力强 天冬氨酸合成能力强 天冬氨酸激酶活力强,对苏氨酸和赖氨酸的协 同反馈抑制不敏感 高丝氨酸脱氢酶活力强,不受苏氨酸反馈调节。
第一节
生物合成途径(天冬氨酸族) 及代谢调控机制
一、生物合成途径
天冬氨酸族氨基酸合成可以以草酰乙酸 或天冬氨酸为原料,合成苏氨酸、异亮氨酸、 蛋氨酸和赖氨酸。
葡萄糖
EMP
丙酮酸
CO2 固定、氧化
氨基化反应
草酰乙酸
天冬氨酸激酶
天冬氨酸
天冬氨酸β-半醛脱氢酶ห้องสมุดไป่ตู้
天冬氨酸β-半醛 赖氨酸
琥珀酰高丝氨 酸合成酶
2.黄色短杆菌
其赖氨酸生物合成调节机制比大肠杆 菌简单,其天冬氨酸激酶只有一种,该酶具 有两个变构部位,可以与终产物结合,当只 有一种终产物(赖氨酸或苏氨酸)与酶结合, 酶活性不受影响。当两种终产物(赖氨酸和 苏氨酸)同时过量时,与酶的两个变构部位 结合,该酶活性受到抑制,这种终产物的反 馈抑制称为协同反馈抑制。P74图6-3
3.抗结构类似物突变株的选育(代谢调节突变)。 优点:突变株遗传性地解除了终产物对自身合成 途径的酶的调节控制,不受培养基中所要求物 质浓度的影响,生产较稳定。 通过诱变使天冬氨酸激酶编码的结构基因 发生突变,对赖氨酸及结构类似物不敏感,即 使有过量苏氨酸存在,该酶也不与赖氨酸或结 构类似物结合,但酶活性中心不变。
②天冬氨酸-β-半醛脱氢酶 专一性的受赖氨酸不完全的抑制 ③二氢吡啶二羧酸(DDP)合成酶(赖氨酸分 支的第一个酶) DDP合成酶受赖氨酸反馈抑制。 二氨基庚二酸(DAP)脱羧酶受赖氨酸阻遏
④高丝氨酸脱氢酶(HD)(通向苏氨酸、蛋氨 酸分支的第一个酶) HD HDⅠ:受苏氨酸反馈抑制 HDⅡ:受蛋氨酸阻遏 ⑤苏氨酸脱氨酶(异亮氨酸合成途径的酶) 受异亮氨酸的反馈抑制和阻遏